Effect of Urbanization on Vegetation in Riparian Area: Plant Communities in Artificial and Semi-Natural Habitats


Abstract
:1. Introduction
- (1)
- Study the changes in plant species richness and composition in artificial and semi-natural habitats along an urbanization gradient. We expected that the species richness and abundance of alien and ruderal species might benefit from the urbanization.
- (2)
- Analyze which urbanization and environmental variables influence plant species richness and species composition in artificial and semi-natural habitats along an urbanization gradient. We hypothesized that the effects of urbanization factors on vegetation might vary with habitat type and that the dominant factor of plant species would also vary in habitat type.
2. Materials and Methods
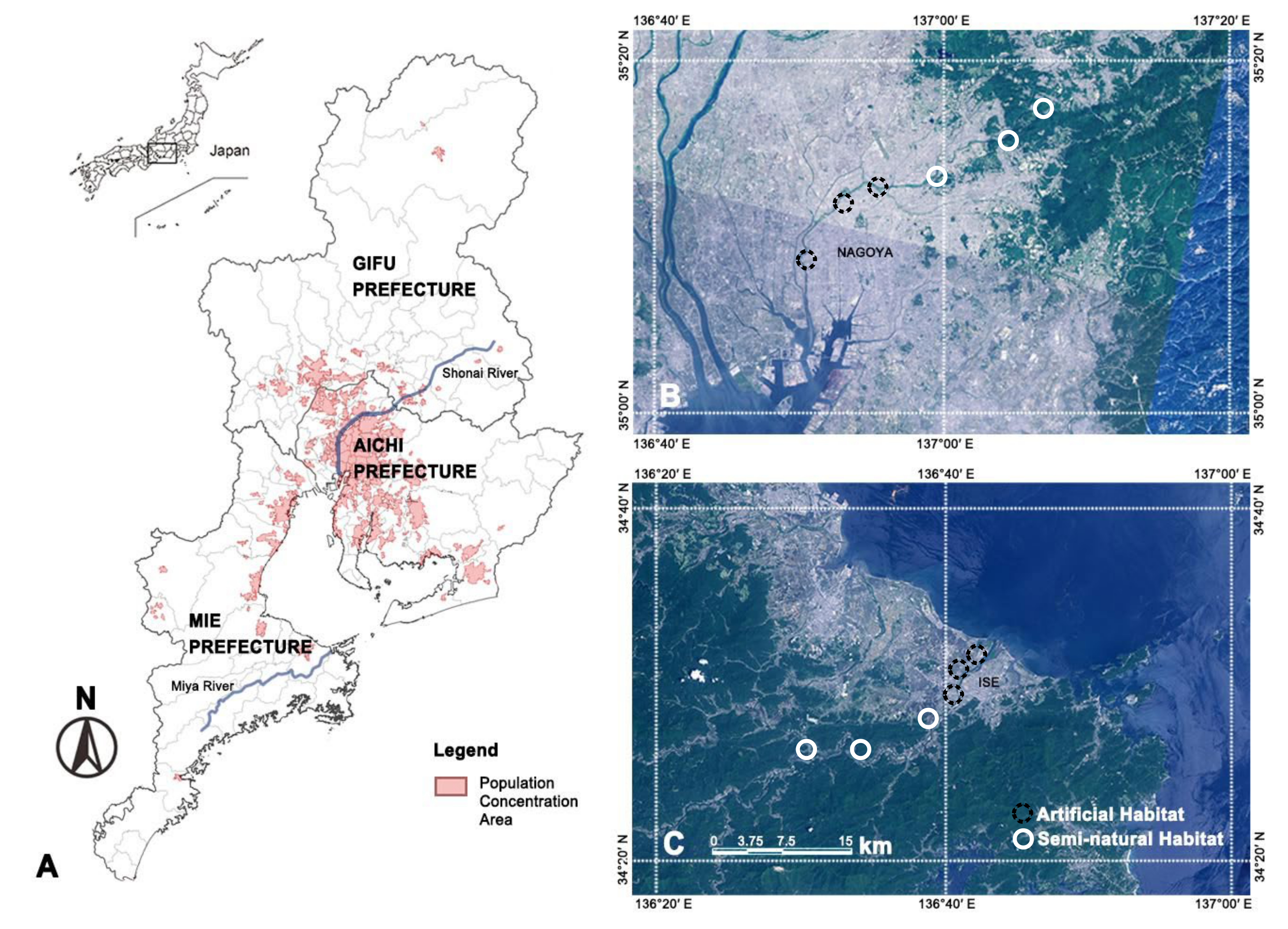
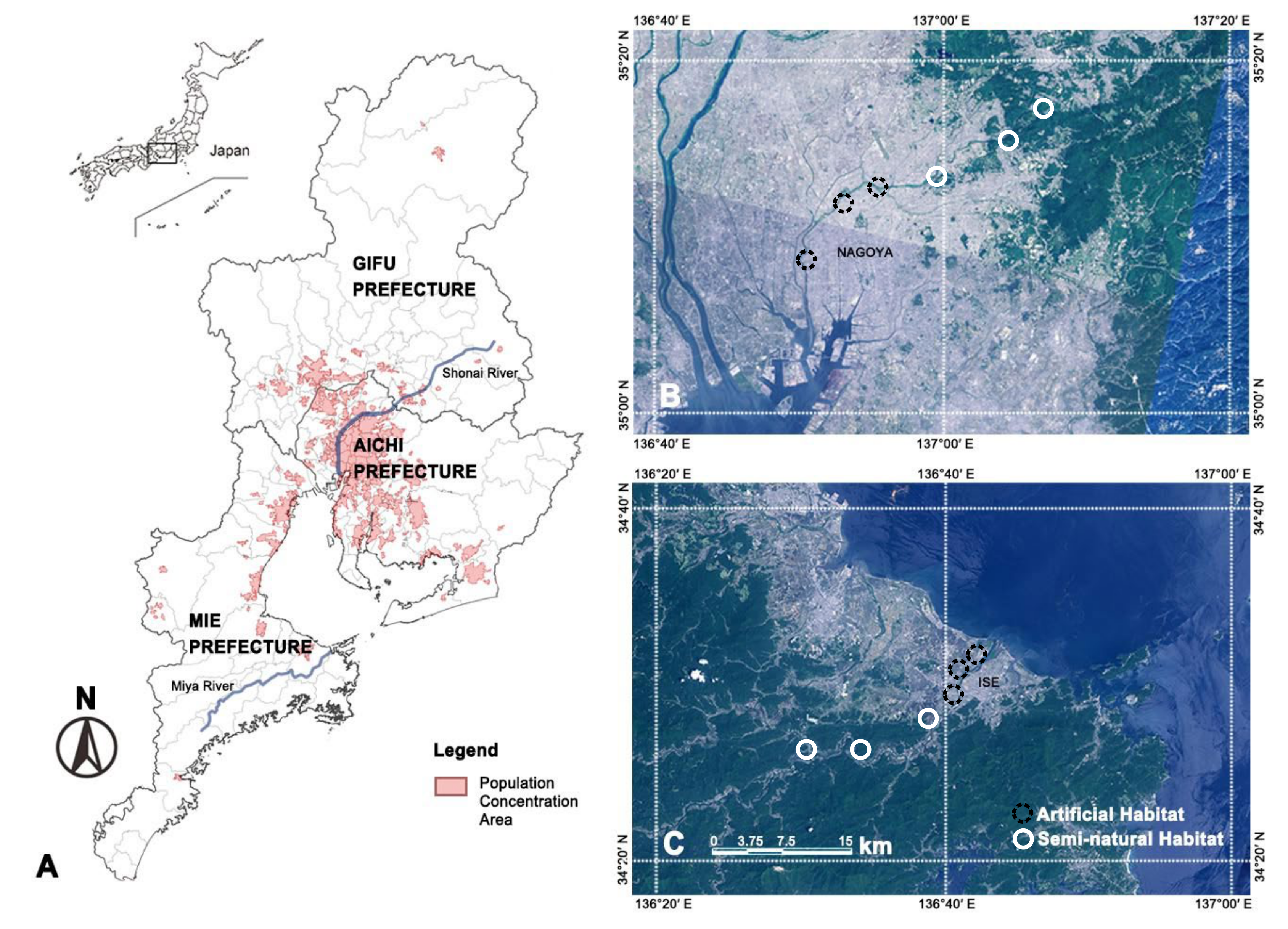
2.1. Study Sites
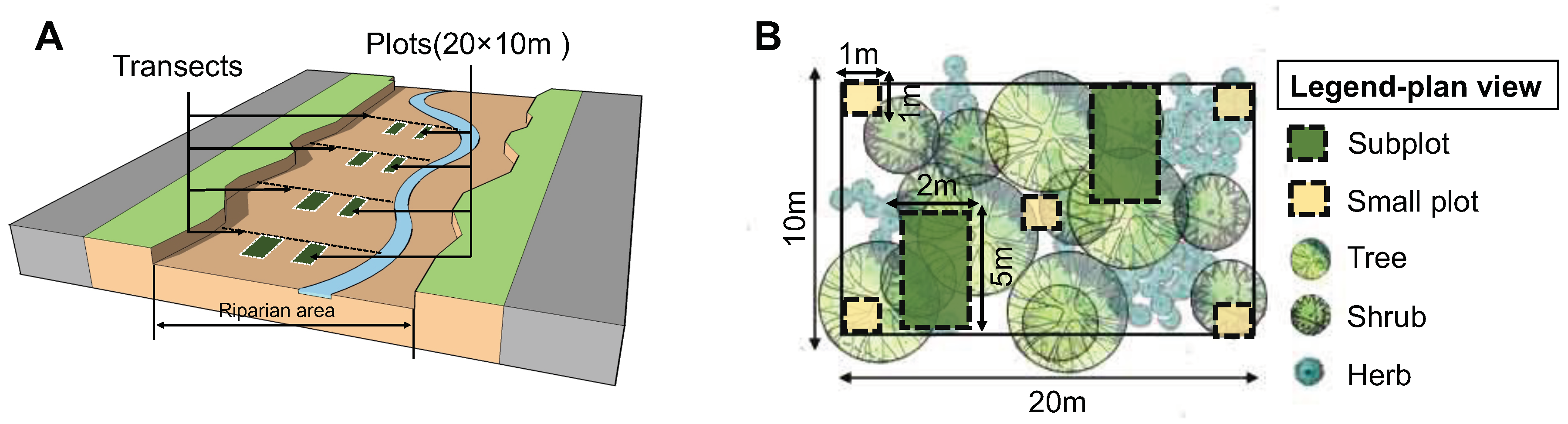
2.2. Vegetation Sampling
2.3. Data Collection
2.4. Statistical Analysis
3. Results
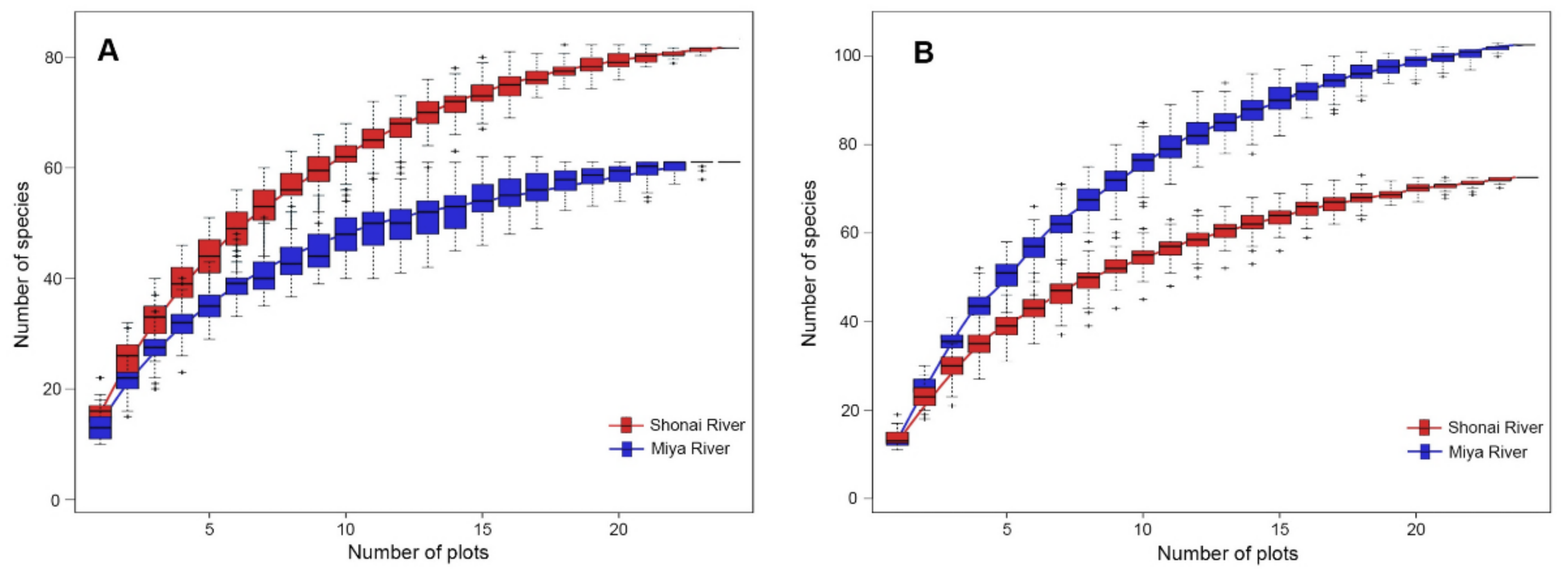
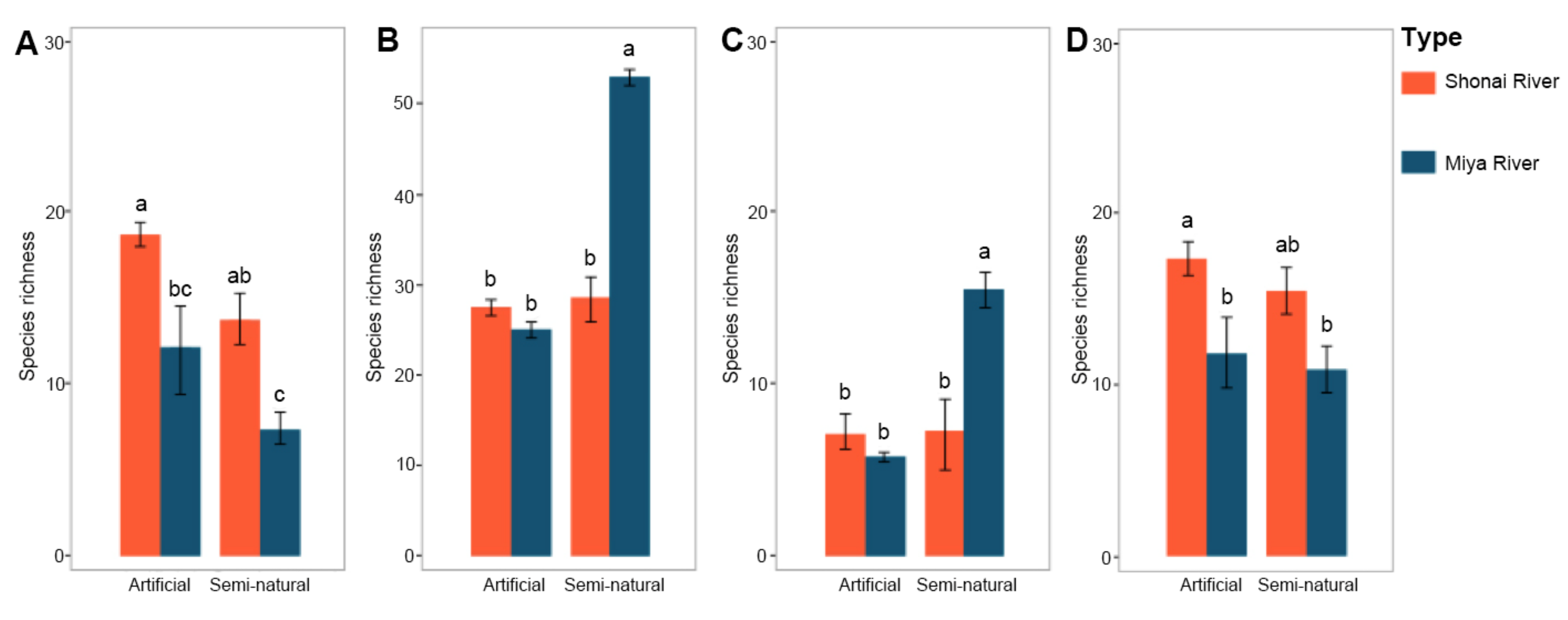
3.1. Plant Species Richness in Artificial and Semi-Natural Habitats
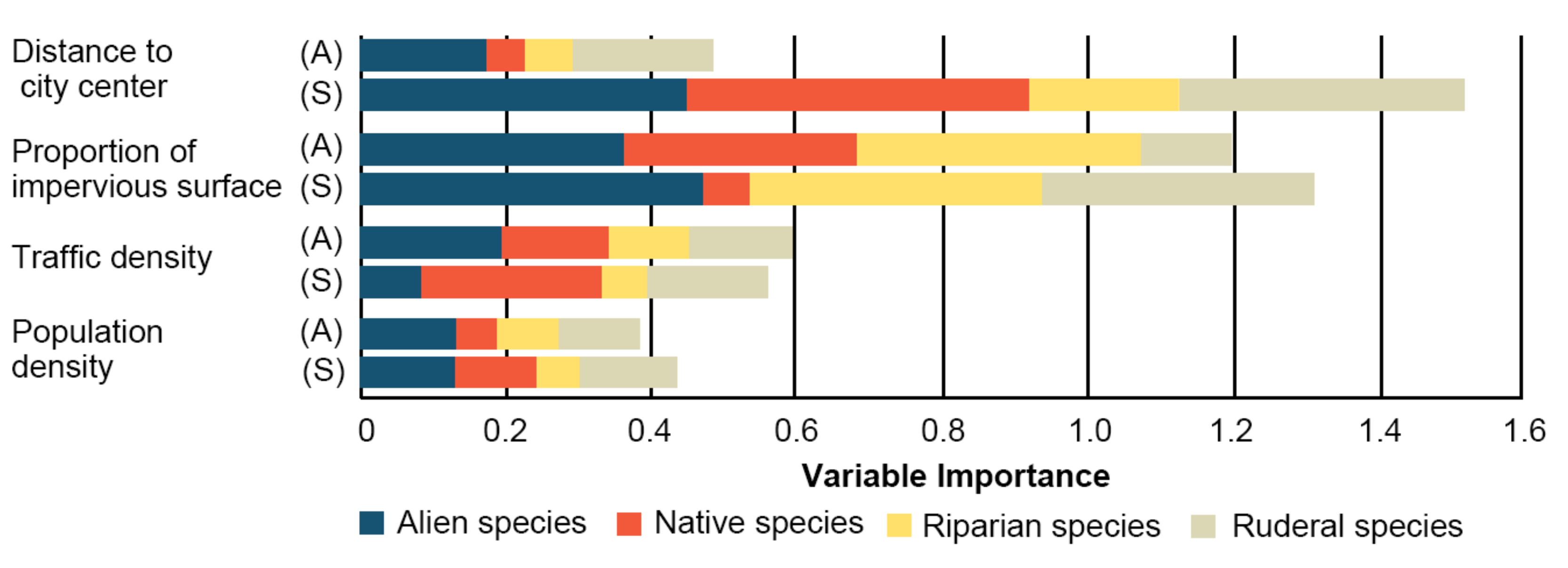
3.2. Components of Urbanization
3.3. Effects of Urbanization on Species Richness
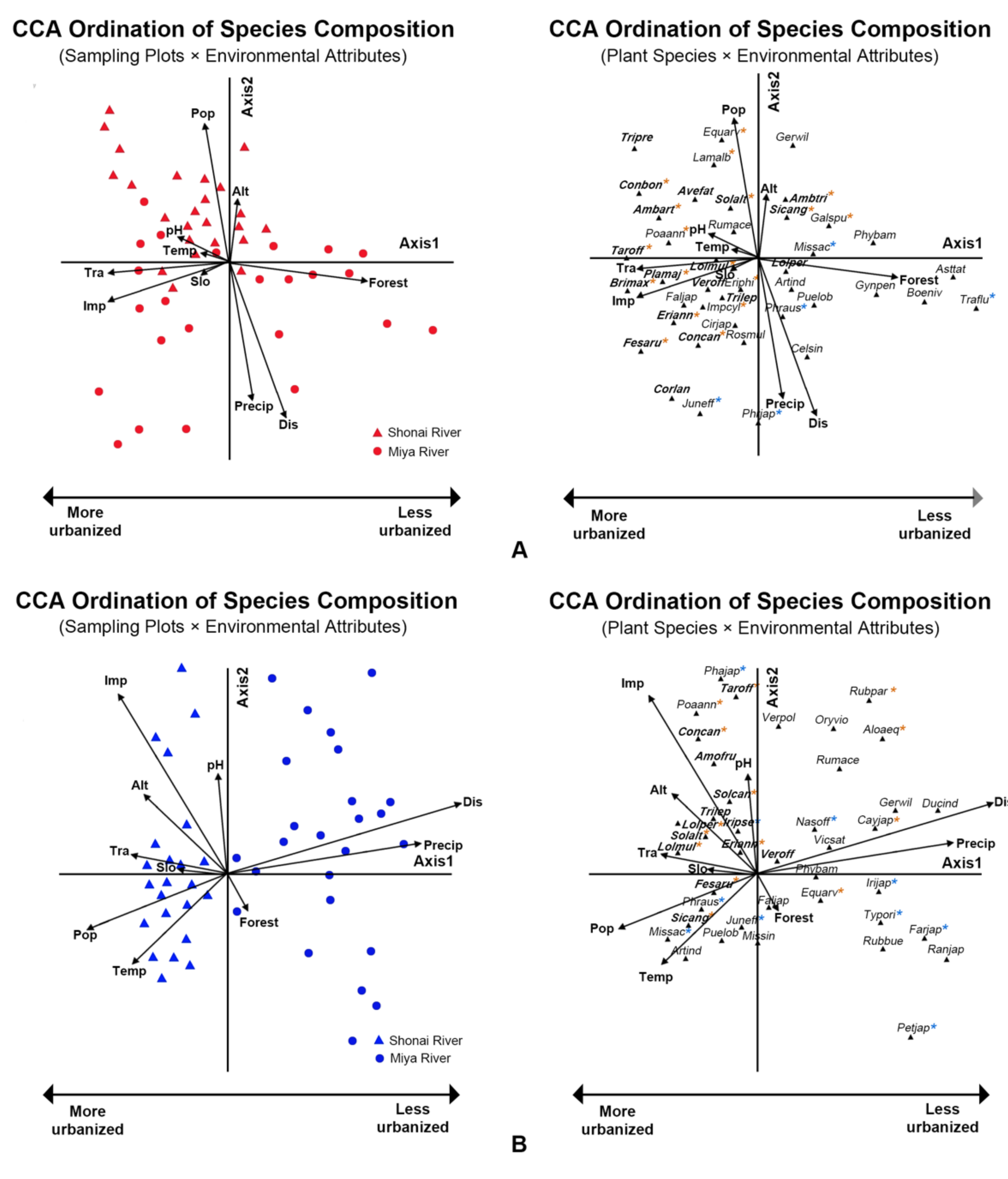
3.4. Effects of Urbanization on Plant Community Composition
4. Discussion
4.1. Plant Species Richness in Artificial and Semi-Natural Habitats
4.2. Driving Factors Affecting Different Types of Vegetation
4.3. Effects on Plant Community Composition
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Naiman, R.; Decamps, H. The Ecology of Interfaces: Riparian Zones. Annu. Rev. Ecol. Syst. 1997, 28. [Google Scholar] [CrossRef] [Green Version]
- Ewel, K.; Cressa, C.; Kneib, R.; Levin, L.; Palmer, M.; Snelgrove, P.; Wall, D. Managing Critical Transition Zones. Ecosystems 2001, 4, 452–460. [Google Scholar] [CrossRef]
- Higgins, S.L.; Thomas, F.; Goldsmith, B.; Brooks, S.J.; Hassall, C.; Harlow, J.; Stone, D.; Völker, S.; White, P. Urban freshwaters, biodiversity and human health and well-being: Setting an interdisciplinary research agenda. Wiley Interdiscip. Rev. Water 2019, 6, 1339. [Google Scholar] [CrossRef]
- Niemelä, J. Urban Ecology: Patterns, Process and Applications; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Hanski, I. Landscape fragmentation, biodiversity loss and the societal response. EMBO Rep. 2005, 6, 388–392. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; Fraedrich, K.; Guan, Y.; Guo, S.; Zhang, C.; Zhu, X. Urbanization and climate change: Insights from eco-hydrological diagnostics. Sci. Total Environ. 2019, 647, 29–36. [Google Scholar] [CrossRef]
- Cai, D.F.K.; Guan, Y.; Guo, S.; Zhang, C.; Carvalho, L.M.; Zhu, X. Causality of Biodiversity Loss: Climate, Vegetation, and Urbanization in China and America. Sensors 2019, 20, 4499. [Google Scholar] [CrossRef] [Green Version]
- White, M.D.; Greer, K.A. The effects of watershed urbanization on the stream hydrology and riparian vegetation of Los Peñasquitos Creek, California. Landsc. Urban Plan. 2006, 74, 125–138. [Google Scholar] [CrossRef]
- Barnosky, A.D.; Hadly, E.A.; Bascompte, J.; Berlow, E.L.; Brown, J.H.; Fortelius, M.; Getz, W.M.; Harte, J.; Hastings, A.; Marquet, P.A.; et al. Approaching a state shift in Earth’s biosphere. Nature 2012, 486, 52–58. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Jiao, F.; Li, Y.H.; Kallenbach, R.L. Anthropogenic disturbances are key to maintaining the biodiversity of grasslands. Sci. Rep. 2016, 6, 22132. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, J.T.; Marlin, A. Habitat origins and microhabitat preferences of urban plant species. Urban Ecosyst. 2006, 9, 139–159. [Google Scholar] [CrossRef]
- Cameron, G.N.; Culley, T.M.; Kolbe, S.E.; Miller, A.I.; Matter, S.F. Effects of urbanization on herbaceous forest vegetation: The relative impacts of soil, geography, forest composition, human access, and an invasive shrub. Urban Ecosyst. 2015, 18, 1051–1069. [Google Scholar] [CrossRef]
- Washitani, I. Plant conservation ecology for management and restoration of riparian habitats of lowland Japan. Popul. Ecol. 2001, 43, 189–195. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Porter, E.E.; Forschner, B.R.; Blair, R.B. Woody vegetation and canopy fragmentation along a forest-to-urban gradient. Urban Ecosyst. 2001, 5, 131–151. [Google Scholar] [CrossRef]
- Duguay, S.; Eigenbrod, F.; Fahrig, L. Effects of surrounding urbanization on non-native flora in small forest patches. Landsc. Ecol. 2007, 22, 589–599. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Pickett, S.T.A. Ecosystem Structure and Function along Urban-Rural Gradients: An Unexploited Opportunity for Ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- Naiman, R.J.; Bechtold, J.S.; Drake, D.C.; Latterell, J.J.; O’Keefe, T.C.; Balian, E.V. Origins, Patterns and Importance of Heterogeneity in Riparian Systems. In Ecosystem Function in Heterogeneous Landscapes; Lovett, G.M., Turner, M.G., Jones, C.G., Weathers, K.C., Eds.; Springer: New York, NY, USA, 2005; pp. 279–309. [Google Scholar] [CrossRef]
- Alberti, M.; Botsford, E.; Cohen, A. Quantifying the urban gradient: Linking urban planning and ecology. In Avian Ecology and Conservation in an Urbanizing World; Marzluff, J.M., Bowman, R., Donnelly, R., Eds.; Springer: Boston, MA, USA, 2001; pp. 89–115. [Google Scholar] [CrossRef]
- Gerstner, K.; Dormann, C.F.; Stein, A.; Manceur, A.M.; Seppelt, R. Editor’s Choice: Review: Effects of land use on plant diversity—A global meta-analysis. J. Appl. Ecol. 2014, 51, 1690–1700. [Google Scholar] [CrossRef]
- Amici, V.; Rocchini, D.; Filibeck, G.; Santi, E.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Landscape structure effects on forest plant diversity at local scale: Exploring the role of spatial extent. Ecol. Complex. 2015, 21, 52–56. [Google Scholar] [CrossRef]
- Inoue, M.; Nakagoshi, N. The effects of human impact on spatial structure of the riparian vegetation along the Ashida river, Japan. Landsc. Urban Plan. 2001, 53, 111–121. [Google Scholar] [CrossRef]
- Guzy, J.C.; Eskew, E.A.; Halstead, B.J.; Price, S.J. Influence of damming on anuran species richness in riparian areas: A test of the serial discontinuity concept. Ecol. Evol. 2018, 8, 2268–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lyons, J.; Kanehl, P.; Bannerman, R. Impacts of Urbanization on Stream Habitat and Fish Across Multiple Spatial Scales. Environ. Manag. 2001, 28, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Damgaard, C. Estimating mean plant cover from different types of cover data: A coherent statistical framework. Ecosphere 2014, 5, 1–7. [Google Scholar] [CrossRef]
- Muranaka, T. Naturalization and invasion of alien plants in Japan: Relationships among their origin, use and time of introduction. Jpn. J. Conserv. Ecol. 2008, 13, 89–101. [Google Scholar] [CrossRef]
- Miyawaki, S. Invasive alien plant species in riparian areas of Japan: The contribution of agricultural weeds, revegetation species and aquacultural species. Glob. Environ. Res. 2004, 10, 89–101. [Google Scholar]
- Kasahara, Y. The Species Grouping of Wild Herbs, Ruderals, Naturalized Plants, Weeds and Crops on the Basis of Their Habitats. J. Weed Sci. Technol. 1971, 12, 23–27. [Google Scholar] [CrossRef]
- Muranaka, T.; Washitani, I. Alien plant invasions and gravelly floodplain vegetation of the Kinu River. Ecol. Civ. Eng. 2001, 4, 121–132. [Google Scholar] [CrossRef]
- Lawton, J.H. What Do Species Do in Ecosystems? Oikos 1994, 71, 367–374. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Multimodel Inference: A Practical Information—Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef] [Green Version]
- Stadler, J.; Trefflich, A.; Klotz, S.; Brandl, R. Exotic plant species invade diversity hot spots: The alien flora of northwestern Kenya. Ecography 2000, 23, 169–176. [Google Scholar] [CrossRef]
- Wania, A.; Kühn, I.; Klotz, S. Plant richness patterns in agricultural and urban landscapes in Central Germany—Spatial gradients of species richness. Landsc. Urban Plan. 2006, 75, 97–110. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Bacher, S.; Chiron, F.; Didžiulis, V.; Essl, F.; et al. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, P.; Yu, F.; Ren, Y.; Liu, D.; Li, J.; Ouyang, Z.; Wang, X. Response of Ruderal Species Diversity to an Urban Environment: Implications for Conservation and Management. Int. J. Environ. Res. Public Health 2018, 15, 2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, J.; Cheptou, P.-O. Effects of fragmentation on plant adaptation to urban environments. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, S.A.O.; Ohlson, H.; Eriksson, O. Effects of historical and present fragmentation on plant species diversity in semi-natural grasslands in Swedish rural landscapes. Landsc. Ecol. 2007, 22, 723–730. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef] [Green Version]
- McKinney, M.L. Urbanization, Biodiversity and Conservation: The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Selmants, P.; Knight, D.H. Understory plant species composition 30–50 years after clearcutting in southeastern Wyoming coniferous forests. For. Ecol. Manag. 2003, 185, 275–289. [Google Scholar] [CrossRef]
- Zhu, J.; Mao, Z.; Hu, L.; Zhang, J. Plant diversity of secondary forests in response to anthropogenic disturbance levels in montane regions of northeastern China. J. For. Res. 2007, 12, 403–416. [Google Scholar] [CrossRef]
- Paul, M.J.; Meyer, J.L. Streams in the Urban Landscape. Annu. Rev. Ecol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Nilsson, C.; Andersson, E.; Merritt, D.M.; Johansson, M.E. Differences in riparian flora between riverbanks and river lakeshores explained by dispersal traits. Ecology 2002, 83, 2878–2887. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Patel, M.V.; O’Neill, K.M.; Ehrenfeld, J.G. Urban riparian systems function as corridors for both native and invasive plant species. Biol. Invasions 2017, 19, 3645–3657. [Google Scholar] [CrossRef]
- Christen, D.C.; Matlack, G.R. The habitat and conduit functions of roads in the spread of three invasive plant species. Biol. Invasions 2009, 11, 453–465. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C. Human-caused environmental change: Impacts on plant diversity and evolution. Proc. Natl. Acad. Sci. USA 2001, 98, 5433–5440. [Google Scholar] [CrossRef] [Green Version]
- Salinitro, M.; Alessandrini, A.; Zappi, A.; Melucci, D.; Tassoni, A. Floristic diversity in different urban ecological niches of a southern European city. Sci. Rep. 2018, 8, 15110. [Google Scholar] [CrossRef] [Green Version]
- Dingaan, M.; Tsubo, M.; Walker, S.; Newby, T. Soil chemical properties and plant species diversity along a rainfall gradient in semi-arid grassland of South Africa. Plant Ecol. Evol. 2017, 150, 35–44. [Google Scholar] [CrossRef]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Levine, J.M. Species Diversity and Biological Invasions: Relating Local Process to Community Pattern. Science 2000, 288, 852. [Google Scholar] [CrossRef] [Green Version]
- Burton, M.L.; Samuelson, L.J.; Pan, S. Riparian woody plant diversity and forest structure along an urban-rural gradient. Urban Ecosyst. 2005, 8, 93–106. [Google Scholar] [CrossRef]
- Tikka, P.M.; Högmander, H.; Koski, P.S. Road and railway verges serve as dispersal corridors for grassland plants. Landsc. Ecol. 2001, 16, 659–666. [Google Scholar] [CrossRef]
- Yan, Z.; Teng, M.; He, W.; Liu, A.; Li, Y.; Wang, P. Impervious surface area is a key predictor for urban plant diversity in a city undergone rapid urbanization. Sci. Total Environ. 2018, 650. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biological Conservation. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Charbonneau, N.C.; Fahrig, L. Influence of canopy cover and amount of open habitat in the surrounding landscape on proportion of alien plant species in forest sites. Écoscience 2004, 11, 278–281. [Google Scholar] [CrossRef]
- McAlpine, K.; Lamoureaux, S.; Westbrooke, I. Ecological impacts of ground cover weeds in New Zealand lowland forests. N. Z. J Ecol. 2015, 39, 50–60. Available online: https://newzealandecology.org/nzje/3215 (accessed on 26 March 2017).
- Lososová, Z.; Chytry, M.; Kühn, I.; Hájek, O.; Horáková, V.; Pyšek, P.; Tichý, L. Plant trait patterns in annual vegetation of Central European human-made habitats. Perspect. Plant Ecol. Evol. Syst. 2006, 81, 69–81. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. Available online: http://www.jstor.org/stable/2460262 (accessed on 8 October 2019).
- Washitani, I. Invasive Alien Species Problems in Japan: An Introductory Ecological Essay. Glob. Environ. Res. 2004, 8, 1–12. [Google Scholar]
- Patamsytė, J.; Rančelis, V.; Čėsnienė, T.; Kleizaitė, V.; Tunaitienė, V.; Naugžemys, D.; Vaitkūnienė, V.; Žvingila, D. Clonal structure and reduced diversity of the invasive alien plant Erigeron annuus in Lithuania. Cent. Eur. J. Biol. 2013, 8, 898–911. [Google Scholar] [CrossRef]
- Djurdjević, L.; Mitrović, M.; Gajić, G.; Jarić, S.; Kostić, O.; Oberan, L.; Pavlović, P. An allelopathic investigation of the domination of the introduced invasive Conyza canadensis L. Flora-Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 921–927. [Google Scholar] [CrossRef]
- Yura, H. Comparative ecophysiology of Chrysanthemum pacificum Nakai and solidago altissima L. 1. why S. altissima cannot be established on the seashore. Ecol. Res. 1997, 12, 313–323. [Google Scholar] [CrossRef]
- Turner, K.; Lefler, L.; Freedman, B. Plant communities of selected urbanized areas of Halifax, Nova Scotia, Canada. Landsc. Urban Plan. 2005, 71, 191–206. [Google Scholar] [CrossRef]
- Stewart, G.H.; Ignatieva, M.E.; Meurk, C.D.; Buckley, H.; Horne, B.; Braddick, T. URban Biotopes of Aotearoa New Zealand (URBANZ) (I): Composition and diversity of temperate urban lawns in Christchurch. Urban Ecosyst. 2009, 12, 233–248. [Google Scholar] [CrossRef]
- Pino, J.; Font, X.; De Cáceres, M.; Molowny-Horas, R. Floristic homogenization by native ruderal and alien plants in north-east Spain: The effect of environmental differences on a regional scale. Glob. Ecol. Biogeogr. 2009, 18, 563–574. [Google Scholar] [CrossRef]
- Martínez-Ramos, M.; Ortiz-Rodríguez, I.A.; Piñero, D.; Dirzo, R.; Sarukhán, J. Anthropogenic disturbances jeopardize biodiversity conservation within tropical rainforest reserves. Proc. Natl. Acad. Sci. USA 2016, 113, 5323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, S.M.; Thompson, K.; Marrs, R.H.; Le Duc, M.G.; Maskell, L.C.; Firbank, L.G. Biotic homogenization and changes in species diversity across human-modified ecosystems. Proc. Biol. Sci. 2006, 273, 2659–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Štajerová, K.; Šmilauer, P.; Brůna, J.; Pyšek, P. Distribution of invasive plants in urban environment is strongly spatially structured. Landsc. Ecol. 2017, 32, 681–692. [Google Scholar] [CrossRef]
- Alston, K.P.; Richardson, D.M. The roles of habitat features, disturbance, and distance from putative source populations in structuring alien plant invasions at the urban/wildland interface on the Cape Peninsula, South Africa. Biol. Conserv. 2006, 132, 183–198. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shonai River | Miya River | |
|---|---|---|
| Latitude | 35°04′–35°24′ N | 34°10′–34°20′ N |
| Longitude | 136°49′–137°20′ E | 136°14′–136°45′ E |
| Climate zone | Warm, temperate climate | Warm, temperate climate |
| Basin area | 1010 km2 | 920 km2 |
| Length | 96 km | 91 km |
| topography | Inclined from the northeast to the southwest | Inclined from the southwest to the northeast |
| Temperature | 27.8 °C (August) and 4.5 °C (January) | 25.3 °C (August) and 5.1 °C (January) |
| Population | 2,461,000 | 140,000 |
| Altitude | 0–843 m | 0–1695 m |
| Precipitation | 1400–2000 mm | 1700–2500 mm |
| Major Cities | Nagoya, Tajimi, Tasugai | Ise |
| Land use | Forest 44%, agricultural field 16%, urban area 40% | Forest 84%, agricultural field 12%, urban area 4% |
| Characteristic | Artificial Habitats | Semi-Natural Habitats |
|---|---|---|
| Number of sampling sites | Shonai River: 3 Miya River: 3 | Shonai River: 3 Miya River: 3 |
| Location of sampling sites | Shonai River: Shonai Greenspace Park, Biwajima Park, Araizeki Park. Miya River: Miyagawatutumi Park, Love River Park, Miyagawashinsui Park. | Shonai River: Sakurasa Cho area, Kanori Cho area, Shimohadagawa Cho area. Miya River: Souchi Cho area, Watarai Cho area, Taiki Cho area. |
| Number of plots in each sampling site | 8 | 8 |
| Total number of plots | 48 | 48 |
| Predictor | Definition | Data Source |
|---|---|---|
| Population density | Number of human inhabitants (residents) per km2 | Statistics Bureau of Japan, https://www.stat.go.jp/data/jinsui/new.html (2015) |
| Traffic density | Daily vehicle flow of the nearest road beside the riparian area (cars/day) | Ministry of Land, Infrastructure, Transport, and Tourism, http://www.mlit.go.jp/road/census/h27/index.html (2015) |
| Mean annual temperature | Average value of monthly mean temperatures (°C) | Japan Meteorological Agency, https://www.data.jma.go.jp/obd/stats/etrn/index.php (2018) |
| Annual precipitation | Sum of monthly precipitation (mm) | Japan Meteorological Agency, https://www.data.jma.go.jp/obd/stats/etrn/index.php (2018) |
| Imperviousness | The rigid pavement area in a 100 m radius from plot center within the riparian area (%) | Geospatial Information Authority of Japan, https://www.gsi.go.jp/tizu-kutyu.html |
| Distance to the city center | Distance from plots to the nearest neighbor city center (m) | Geospatial Information Authority of Japan, https://www.gsi.go.jp/tizu-kutyu.html |
| Forest | The proportion of the plot area occupied by trees (%) | Geospatial Information Authority of Japan, https://www.gsi.go.jp/tizu-kutyu.html |
| Altitude and slope | The elevation and degree of inclination in each plot | Measured using GPS and declinometer, respectively |
| pH | The pH of the soil within a depth of 20 cm in each plot | Measured using an electric pH meter |
| (a) | |||
| Urbanization Factors | Shonai River | Miya River | Difference between Rivers |
| Mean ± SD | Mean ± SD | ||
| Population density | 6707.33 ± 391.40 | 610 | p < 0.001 |
| Proportion of impervious surface | 22.22 ± 2.14 | 20.88 ± 1.62 | p = 0.34 |
| Distance to the city center | 3.33 ± 0.08 | 3.72 ± 0.07 | p = 0.53 |
| Traffic density | 6810.66 ± 337.71 | 4865.83 ± 444.98 | p = 0.002 |
| (b) | |||
| Urbanization Factors | Shonai River | Miya River | Difference between Rivers |
| Mean ± SD | Mean ± SD | ||
| Population density | 2222.67 ± 246.85 | 160 ± 32.48 | p < 0.001 |
| Proportion of impervious surface | 7.7 ± 0.63 | 5.86 ± 0.96 | p = 0.23 |
| Distance to the city center | 4.32 ± 0.19 | 15.49 ± 1.44 | p < 0.001 |
| Traffic density | 3334.54 ± 307.80 | 2083.79 ± 380.27 | p = 0.041 |
| Adj.R2 | AIC | Pop. | Imp. | Dis. | Tra. | Temp. | Precip. | Alt. | Slo. | Forest | pH | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alien Species Richness | ||||||||||||
| Artificial | 0.660 *** | 182.24 | 0.364 | 0.315 | ||||||||
| Semi-natural | 0.646 *** | 144.65 | 0.466 | 0.432 | 0.225 | |||||||
| Native Species Richness | ||||||||||||
| Artificial | 0.287 *** | 205.29 | 0.327 | 0.156 | ||||||||
| Semi-natural | 0.500 *** | 216.17 | 0.464 | |||||||||
| Riparian Species Richness | ||||||||||||
| Artificial | 0.329 *** | 141.90 | 0.397 | |||||||||
| Semi-natural | 0.460 ** | 164.36 | 0.423 | 0.250 | 0.260 | |||||||
| Ruderal Species Richness | ||||||||||||
| Artificial | 0.423 *** | 185.78 | 0.201 | 0.305 | ||||||||
| Semi-natural | 0.566 ** | 175.81 | 0.381 | 0.419 | 0.197 | 0.256 | 0.328 | |||||
| Artificial Habitats | Semi-Natural Habitats | |||
|---|---|---|---|---|
| Axis 1 | Axis 2 | Axis 1 | Axis 2 | |
| Eigenvalue | 0.462 | 0.374 | 0.659 | 0.423 |
| Constrained proportion | 0.261 | 0.212 | 0.281 | 0.177 |
| Cumulative proportion | 0.261 | 0.473 | 0.281 | 0.459 |
| Species–environment correlations | 0.859 | 0.737 | 0.946 | 0.828 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Natuhara, Y. Effect of Urbanization on Vegetation in Riparian Area: Plant Communities in Artificial and Semi-Natural Habitats. Sustainability 2020, 12, 204. https://doi.org/10.3390/su12010204
Cao Y, Natuhara Y. Effect of Urbanization on Vegetation in Riparian Area: Plant Communities in Artificial and Semi-Natural Habitats. Sustainability. 2020; 12(1):204. https://doi.org/10.3390/su12010204
Chicago/Turabian StyleCao, Yang, and Yosihiro Natuhara. 2020. "Effect of Urbanization on Vegetation in Riparian Area: Plant Communities in Artificial and Semi-Natural Habitats" Sustainability 12, no. 1: 204. https://doi.org/10.3390/su12010204
APA StyleCao, Y., & Natuhara, Y. (2020). Effect of Urbanization on Vegetation in Riparian Area: Plant Communities in Artificial and Semi-Natural Habitats. Sustainability, 12(1), 204. https://doi.org/10.3390/su12010204