Effects of Different Nutrient and Trace Metal Concentrations on Growth of the Toxic Dinoflagellate Gymnodinium catenatum Isolated from Korean Coastal Waters


 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
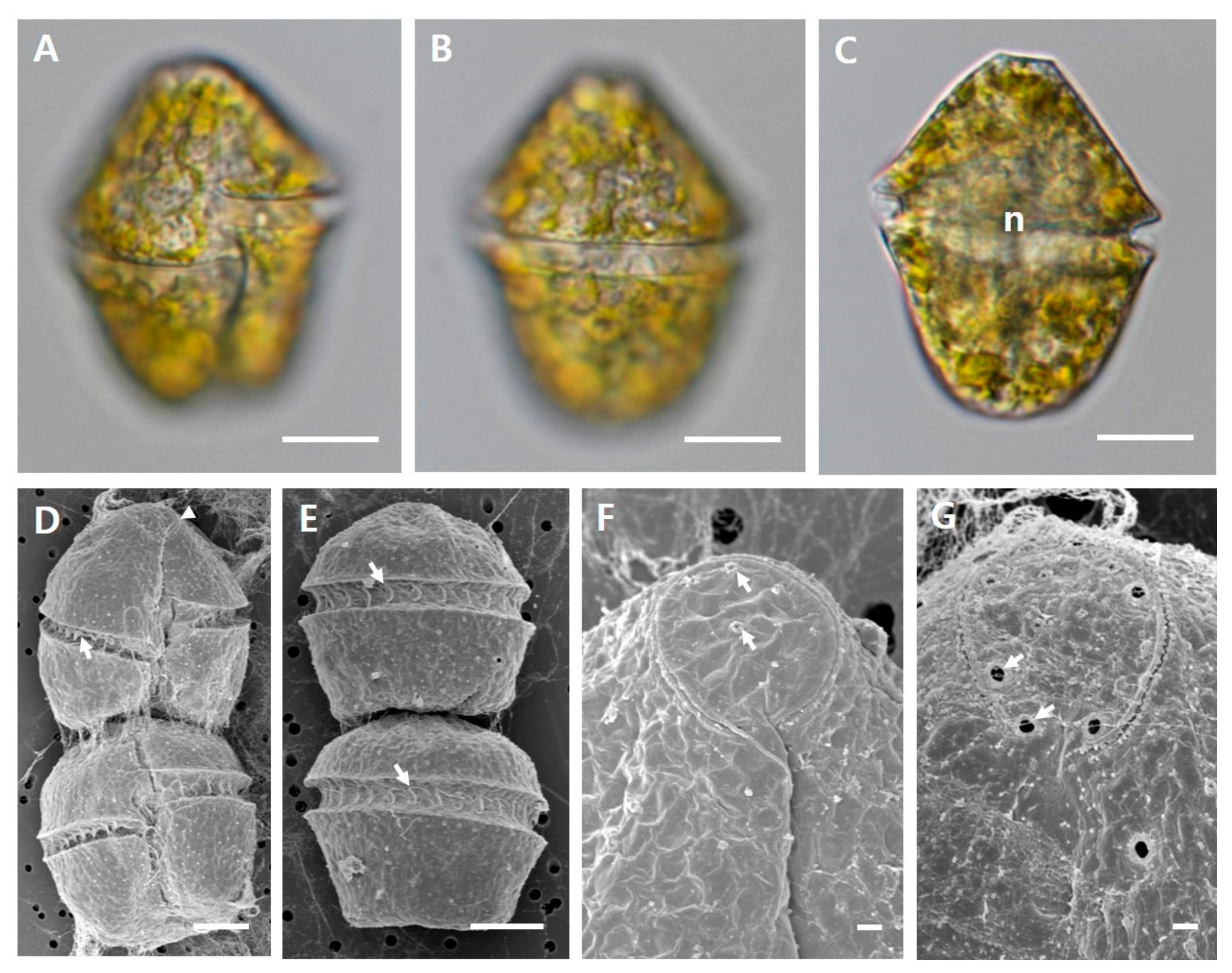
2.1. Culture of Gymnodinium catenatum
2.2. Growth Experiment
2.3. Calculation of Growth Rate
3. Results and Discussion
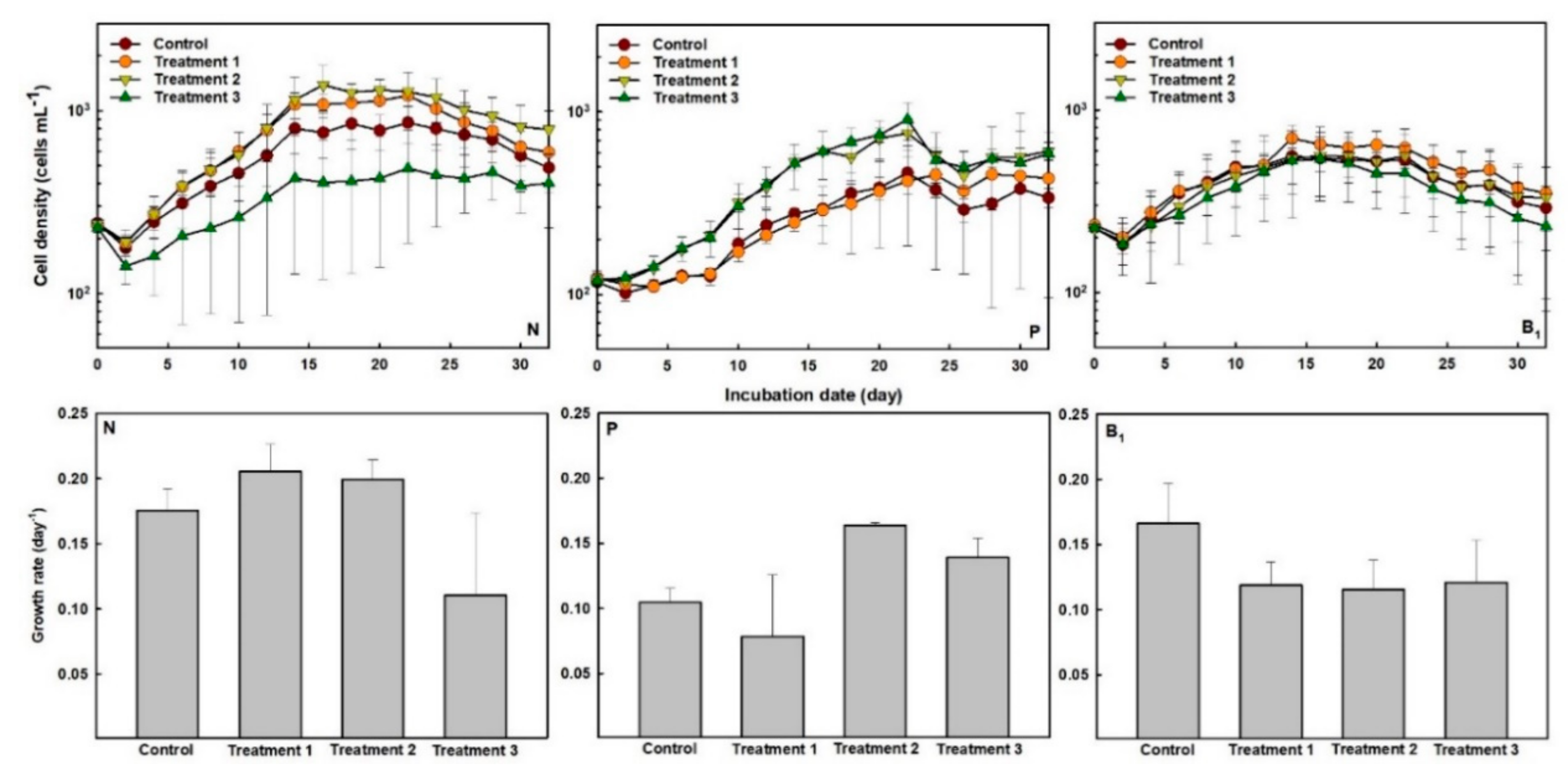
3.1. Growth of Gymnodinium catenatum under Different Concentrations of N, P and Vitamin B1
3.2. Growth of Gymnodinium catenatum under Different Concentrations of Fe, Cu and Se
3.3. Proliferation Potential of Gymnodinium catenatum and Nutritional Conditions in Korean Coastal Waters
Author Contributions
Funding
Conflicts of Interest
References
- Lagos, N.; Onodera, H.; Zagatto, P.A.; Andrinolo, D.; Azevedo, S.M.F.Q.; Oshima, Y. The first evidence of paralytic shellfish toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon 1999, 37, 1359–1373. [Google Scholar] [CrossRef]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [PubMed]
- Band–Schmidt, C.J.; Durán–Riveroll, L.M.; Bustillos–Guzmán, J.J.; Leyva–Valencia, I.; López–Cortés, D.J.; Núñez–Vázquez, E.J.; Hernández–Sandoval, F.E.; Ramirez–Rodriguez, D.V. Paralytic toxin producing dinoflagellates in Latin America: Ecology and physiology. Front. Mar. Sci. 2019, 6, 42. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Landsberg, J.H. The Effects of Harmful Algal Blooms on Aquatic Organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Yñiguez, A.T.; Lim, P.T.; Leaw, C.P.; Jipanin, S.J.; Iwataki, M.; Benico, G.; Azanza, R.V. Over 30 years of HABs in the Philippines and Malaysia: What have we learned? Harmful Algae 2020, 101776. [Google Scholar] [CrossRef]
- Chang, D.S.; Shin, I.S.; Pyeun, J.H.; Park, Y.H. A study on paralytic shellfish poison of sea mussel Mytilus edulis. Korean J. Fish. Aquat. Sci. 1987, 20, 293–299. [Google Scholar]
- Azanza, R.V.; Taylor, F.J. Are Pyrodinium blooms in the Southeast Asian region recurring and spreading? A review at the end of the millennium. Ambio 2001, 39, 356–364. [Google Scholar] [CrossRef]
- Harada, T.; Oshima, Y.; Kamiya, H.; Yasumoto, T. Confirmation of paralytic shellfish toxins in the dinoflagellate Pyrodinium bahamense var. compressa and bivalves in Palau. Nippon Suisan Gakkaishi 1982, 48, 821–825. [Google Scholar] [CrossRef]
- Oshima, Y.; Hasegawa, M.; Yasumoto, T.; Hallegraeff, G.; Blackburn, S. Dinoflagellate Gymnodinium catenatum as the source of paralytic shellfish toxins in Tasmanian shellfish. Toxicon 1987, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Doucette, G.J.; Gallagher, J.C.; Balech, E. Biogeography of toxic dinoflagellates in the genus Alexandrium from the northeastern United States and Canada. Mar. Biol. 1994, 120, 467–478. [Google Scholar] [CrossRef]
- Taylor, F.J.R.; Fukuyo, Y.; Larsen, J.; Hallegraeff, G.M. Taxonomy of Harmful Dinoflagellates. In Manual on Harmful Marine Microalgae. Monographs on Oceanographic Methodology, 11th ed.; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; Amsterdam: Paris, France, 2003; pp. 389–432. [Google Scholar]
- Moestrup, Ø.; Akselmann-Cardella, R.; Churro, C.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Larsen, J.; Lundholm, N.; Zingone, A. IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. Available online: http://www.marinespecies.org/hab (accessed on 20 February 2020).
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonomy: The Introduction of five species with emphasis on molecular–based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef]
- Murray, S.A.; Diwan, R.; Orr, R.J.; Kohli, G.S.; John, U. Gene duplication, loss and selection in the evolution of saxitoxin biosynthesis in alveolates. Mol. Phylogenetics Evol. 2015, 92, 165–180. [Google Scholar] [CrossRef]
- Lassus, P.; Chomérat, N.; Hess, P.; Nézan, E. Toxic and Harmful Microalgae of the World Ocean (Micro-Algues Toxiques et Nuisibles de l’océan Mondial; IOC Manuals and Guides 68 (Bilingual English/French); International Society for the Study of Harmful Algae; Intergovernmental Oceanographic Commission of UNESCO: Copenhagen, Denmark, 2016; p. 524. [Google Scholar]
- Hallegraeff, G.M.; Stanley, S.O.; Bolch, C.J.; Blackburn, S.I. Gymnodinium catenatum blooms and shellfish toxicity in Southern Tasmania, Australia. In Red Tide: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemato, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 77–80. [Google Scholar]
- Franca, S.; Almeida, J.K. Paralytic shellfish poisons in bivalve molluscs on the Portuguess coast caused by a bloom of the dinoflagellate Gymnodinium catenatum. In Red Tide: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemato, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 93–96. [Google Scholar]
- Anderson, D.M.; Sullivan, J.J.; Reguera, B. Paralytic shellfish poisoning in northwest Spain: The toxicity of the Gymnodinium catenatum. Toxicon 1989, 27, 665–674. [Google Scholar] [CrossRef]
- Amanhir, R.; Benhadouch, A.; Taleb, H.; Leblad, R.; Vale, P.; Blaghen, M. Weekly occurrence of Gymnodinium catenatum and paralytic shellfish poisoning in the Mediterranean shore of Morocco. Int. J. Biochem. Res. Rev. 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Ikeda, T.; Matsuno, S.; Sato, S. First report on paralytic shellfish poisoning caused by Gymnodinium catenatum Graham (Dinophyceae) in Japan. In Red Tide: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemato, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 411–414. [Google Scholar]
- Park, T.G.; Kim, C.H.; Oshima, Y. Paralytic shellfish toxin profiles of different geographic population of Gymnodinium catenatum (Dinophyceae) in Korean coastal waters. Phycol. Res. 2004, 52, 300–305. [Google Scholar] [CrossRef]
- Smayda, T.J. Ecological features of harmful algal blooms in coastal upwelling ecosystems. Afr. J. Mar. Sci. 2000, 22, 219–253. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton bloom in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Harmful algal blooms: A global overview. In Manual on Harmful Marine Microalgae, 2nd ed.; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO: Paris, France, 1995; pp. 25–50. [Google Scholar]
- Global Ecology and Oceanography of Harmful Algal Blooms (GEOHAB). Global Ecology and Oceanography of Harmful Algal Blooms. In Global Ecology and Oceanography of Harmful Algal Blooms: Eutrophication and HABs; Gilbert, P., Ed.; Scientific Committee on Oceanic Research (SCOR: Paris, France; Intergovernmental Oceanographic Commission (IOC): Baltimore, MD, USA, 2006; p. 74. [Google Scholar]
- Kudela, R.M.; Seeyave, S.; Cochlan, W.P. The role of nutrients in regulation and promotion of harmful algal blooms in upwelling systems. Prog. Oceanogr. 2010, 85, 122–135. [Google Scholar] [CrossRef]
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Gilbert, P.M.; Gobler, C.J.; Hell, C.A.; Kudela, R.M.; Parsons, M.L.; Rensel, J.E.J.; Townsend, D.W.; et al. Harmful algal blooms and eutrophication: Examining linkages from selected coastal regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef]
- Heisler, J.P.; Gilbert, J.; Burkholder, J.; Anderson, D.; Cochlan, W.; Dennison, W.; Dortch, Q.; Gobler, C.J.; Heil, C.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Bruland, K.W.; Donat, J.R.; Hutchins, D.A. Interactive influences of bioactive trace metals on biological production in oceanic waters. Limnol. Oceanogr. 1991, 36, 1555–1577. [Google Scholar] [CrossRef]
- Maldonado, M.T.; Allen, A.E.; Chong, J.S.; Lin, K.; Leus, D.; Karpenko, N.; Harris, S.L. Copper-dependent iron transport in coastal and oceanic diatoms. Limnol. Oceanogr. 2006, 51, 1729–1743. [Google Scholar] [CrossRef]
- Doblin, M.A.; Blackburn, S.I.; Hallegraeff, G.M. Intraspecific variation in the selenium requirement of different geographic strains of the toxic dinoflagellate Gymnodinium catenatum. J. Plankton Res. 2000, 22, 421–432. [Google Scholar] [CrossRef]
- Boyer, G.L.; Brand, L.E. Trace metals and harmful algal blooms. In Physiological Ecology of Harmful Algal Blooms; NATO-ASI Series 41; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998; pp. 489–508. [Google Scholar]
- Doblin, M.A.; Blackburn, S.I.; Hallegraeff, G.M. Growth and biomass stimulation of the toxic dinoflagellate Gymnodinium catenatum (Graham) by dissolved organic substances. J. Exp. Mar. Biol. Ecol. 1999, 236, 33–47. [Google Scholar] [CrossRef]
- Couet, D.; Pringault, O.; Bancon-Montigny, C.; Briant, N.; Poulichet, F.E.; Delpoux, S.; Yahia, O.K.; Hela, B.; Charaf, M.; Hervé, F.; et al. Effects of cooper and butyltin compounds on the growth, photosynthetic activity and toxin production of two HAB dinoflagellates: The planktonic Alexandrium catenella and the benthic Ostreopsis cf. ovata. Aquat. Toxicol. 2018, 196, 154–167. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Koch, F.; Gobler, C.J. Most harmful algal bloom species are vitamin B1 and B12 auxotrophs. Proc. Natl. Acad. Sci. USA 2010, 107, 20756–20761. [Google Scholar] [CrossRef]
- Lee, C.K.; Kim, H.C.; Lee, S.G.; Jung, C.S.; Kim, H.G.; Lim, W. Abundance of harmful algae, Cochlodinium polykrikoides, Gyrodinium impudicum and Gymnodinium catenatum in the coastal area of South Sea of Korea and their effects of temperature, salinity, irradiance and nutrient on the growth in culture. Korean J. Fish. Aquat. Sci. 2001, 34, 536–544. [Google Scholar]
- Han, K.H.; Li, Z.; Kang, B.J.; Youn, J.Y.; Shin, H.H. Toxic dinoflagellate Gymnodinium catenatum Graham (Dinophyceae) from the southern coast of Korea: Morphology, phylogeny and effects of temperature and salinity on growth. Korean J. Environ. Biol. 2019, 37, 31–41. [Google Scholar] [CrossRef]
- Band–Schmidt, C.J.; Morquecho, L.; Lechuga-Devéze, C.H.; Anderson, D.M. Effects of growth medium, temperature, salinity and seawater source on the growth of Gymnodinium catenatum (Dinophyceae) from Bahía Concepción, Gulf of California, Mexico. J. Plankton Res. 2004, 26, 1459–1470. [Google Scholar] [CrossRef]
- Bustillos-Guzmán, J.J.; Band-Schmidt, C.J.; López-Cortés, D.J.; Gárate-Lizárraga, I.; Núñez-Vázquez, E.J.; Hernández-Sandoval, F.E. Variations in growth and toxicity in Gymnodinium catenatum Graham from the Gulf of Calinifornia under different ratios of nitrogen and phosphorus. Cienc. Mar. 2012, 38, 101–117. [Google Scholar] [CrossRef]
- Oh, S.J.; Yamamoto, T.; Kataoka, Y.; Matsuda, O.; Matsuyama, Y.; Kotani, Y. Utilization of dissolved organic phosphorus by the two toxic dinoflagellates, Alexandrium tamarense and Gymnodinium catenatum (Dinophyceae). Fish. Sci. 2002, 68, 416–424. [Google Scholar] [CrossRef]
- Mather, R.L.; Reynolds, S.E.; Wolff, G.A.; Williams, R.G.; Torres-Valdes, S.; Woodward, E.M.S.; Landolfi, A.; Pan, X.; Sanders, R.; Achterberg, E.P. Phosphorus cycling in the north and South Atlantic Ocean subtropical gyres. Nat. Geosci. 2008, 1, 439–443. [Google Scholar] [CrossRef]
- Yamamoto, T.; Oh, S.J.; Kataoka, Y. Growth and uptake kinetics for nitrate, ammonium and phosphate by the toxic dinoflagellate Gymnodinium catenatum isolated from Hiroshima Bay, Japan. Fish. Sci. 2004, 70, 108–115. [Google Scholar] [CrossRef]
- Gobler, C.J.; Norman, C.; Panzeca, C.; Taylor, G.T.; Sañudo-Wilhelmy, S.A. Effect of B-Vitamins (B1, B12) and inorganic nutrients on algal bloom dynamics in a coastal ecosystem. Aquat. Microb. Ecol. 2007, 49, 181–194. [Google Scholar] [CrossRef]
- Croft, M.T.; Warren, M.J.; Smith, A.G. Algae need their vitamins. Eukaryot. Cell 2006, 5, 1175–1183. [Google Scholar] [CrossRef]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.G.; Ades, D.R. The role of iron nutrition in photosynthesis and nitrogen assimilation in Scenedemus quadricauda (Chlorophyceae). J. Phycol. 1987, 23, 452–457. [Google Scholar] [CrossRef]
- Geider, R.J.; La Roche, J. The role of iron in phytoplankton photosynthesis, and the potential for iron limitation of primary productivity in the sea, Mini review. Photosynth. Res. 1994, 39, 275–301. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Amandi, M.F.; Mckenzie, L.; Furey, A.; James, K.J. Effects of selenium, iron and cobalt addition to growth and yessotoxin production of the toxic marine dinoflagellate Protoceratium reticulatum in culture. J. Exp. Mar. Biol. Ecol. 2004, 313, 337–351. [Google Scholar] [CrossRef]
- He, H.; Chen, F.; Li, H.; Xiang, W.; Li, Y.; Jiang, Y. Effect of iron on growth, biochemical composition and paralytic shellfish poisoning toxins production of Alexandrium tamarense. Harmful Algae 2010, 9, 98–104. [Google Scholar] [CrossRef]
- Qin, X.M.; Zhou, J.Z.; Qian, P.Y. Effects of Fe and Mn on the growth of a red tide dinoflagellate Scrippsiella trochoidea (Stein) Loeblch III. Chin. J. Oceanol. Limnol. 1997, 15, 173–180. [Google Scholar] [CrossRef]
- Herzi, F.; Jean, N.; Zhao, H.; Mounier, S.; Mabrouk, H.H.; Hlaili, A.S. Cooper and cadmium effects on growth and extracellular exudation of the marine toxic dinoflagellate Alexandrium catenaella: 3D-fluorescence spectroscopy approach. Chemosphere 2013, 93, 1230–1239. [Google Scholar] [CrossRef]
- Doblin, M.A.; Blackburn, S.I.; Hallegraeff, G.M. Comparative study of selenium requirements of three species: Gymnodinium catenatum, Alexandrium minutum (Dinophyta) and Chaetoceros cf. tenuissimus (Bacillariophyta). J. Plankton Res. 1999, 21, 1153–1169. [Google Scholar] [CrossRef]
- Boisson, F.; Gnassia-Barelli, M.; Romeo, M. Toxicity and accumulation of selenite and selenate in the unicellular marine alga Cricosphaera elongata. Arch. Environ. Contam. Toxicol. 1995, 28, 487–493. [Google Scholar] [CrossRef]
- Price, N.M.; Thompson, P.A.; Harrison, P.J. Selenium: An essential element for growth of the coastal marine diatom Thalassiosira pseudonana (bacillariophyceae) 1, 2. J. Phycol. 1987, 23, 1–9. [Google Scholar] [CrossRef]
- Band-Schmidt, C.J.; Bustillos-Guzmán, J.J.; López-Cortés, D.J.; Gárate-Lizárraga, I.; Núñez-Vázquez, E.J.; Hernández-Sandoval, F.E. Ecological and physiological studies of Gymnodinium catenatum in the Mexican Pacific: A review. Mar. Drugs 2010, 8, 1935–1961. [Google Scholar] [CrossRef]
- Koo, J.S. A Study on the Relation between Nutrient and HAB in the South Sea of Korea. Master’s Thesis, Pusan National University, Busan, Korea, 2004. [Google Scholar]
- Cho, E.S. A comparative study on outbreak and non-outbreak of Cochlodinium polykrikoides Margalef in South Sea of Korea in 2007–2009. Korean Soc. Mar. Environ. Saf. 2010, 16, 31–41. [Google Scholar]
- Jang, P.G.; Shin, H.H.; Baek, S.H.; Jang, M.C.; Lee, T.S.; Shin, K. Nutrient distribution and effects on phytoplankton assemblages in the western Korea/Tsushima Strait. N. Z. J. Mar. Freshw. Res. 2013, 47, 21–37. [Google Scholar] [CrossRef]
- Lee, Y.S. Factors affecting outbreaks of high-density Cochlodinium polykrikoides red tides in the coastal seawaters around Yeosu and Tongyeong, Korea. Mar. Pollut. Bull. 2006, 52, 1249–1259. [Google Scholar] [CrossRef]
- Wang, H.; Lu, D.; Huang, H.; Xia, P.; Dai, X. Comparison of morphological structure and ITS sequence between two Prorocentrum strains from the East China Sea and Masan Bay of Korea. J. Mar. Sci. 2011, 29, 42–48. [Google Scholar]
- Shin, H.H.; Li, Z.; Kim, E.S.; Park, J.W.; Lim, W.A. Which species, Alexandrium catenella (Group I) or A. pacificum (Group IV), is really responsible for past paralytic shellfish poisoning outbreaks in Jinhae-Masan Bay, Korea? Harmful Algae 2017, 68, 31–39. [Google Scholar] [CrossRef]
- Shin, H.H.; Li, Z.; Mertens, K.N.; Seo, M.H.; Gu, H.; Lim, W.A.; Yoon, Y.H.; Soh, H.Y.; Matsuoka, K. Prorocentrum shikokuense Hada and P. donghaiense Lu are junior synonyms of P. obtusidens Schiller, but not of P. dentatum Stein (Prorocentrales, Dinophyceae). Harmful Algae 2019, 89, 101686. [Google Scholar] [CrossRef]
- Kim, D.I.; Matsuyama, Y.; Nagasoe, S.; Yamaguchi, M.; Yoon, Y.H.; Oshima, Y.; Imada, N.; Honjo, T. Effects of temperature, salinity and irradiance on the growth of the harmful red tide dinoflagellate Cochlodinium polykrikoides Margalef (Dinophyceae). J. Plankton Res. 2004, 26, 61–66. [Google Scholar] [CrossRef]
- Hu, Z.; Mulholland, M.R.; Duan, S.; Xu, N. Effects of nitrogen supply and its composition on the growth of Prorocentrum donghaiense. Harmful Algae 2012, 13, 72–82. [Google Scholar] [CrossRef]
- Oh, S.J.; Park, J.A.; Kwon, H.K.; Yang, H.S.; Lim, W.A. Ecophysiological studies on the population dynamics of two toxic dinoflagellates Alexandrium tamarense and Alexandrium catenella isolated from the southern coast of Korea. I. Effects of temperature and salinity on the growth. J. Korean Soc. Mar. Environ. Energy 2012, 15, 133–141. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | Treatment 1 | Treatment 2 | Treatment 3 | Treatment 4 | Treatment 5 | |
|---|---|---|---|---|---|---|
| Nutrient | ||||||
| N | None added | 8.82 101 (2.6) | 8.82 102 (24.2) | 1.76 103 (48.1) | - | - |
| P | None added | 3.62 1 (210.9) | 3.62 101 (24.2) | 7.24 101 (12.2) | - | - |
| Vitamin B1 | None added | 2.96 10−2 | 2.96 10−1 | 5.92 10−1 | - | - |
| Trace Metal | ||||||
| Fe | None added | 1.17 1 | 1.17 101 | 2.34 101 | - | - |
| Cu | None added | 3.93 10−3 | 3.93 10−2 | 7.86 10−2 | - | - |
| Se | None added | 10−5 | 10−4 | 10−3 | 10−2 | 10−1 |
| Control | Treatment 1 | Treatment 2 | Treatment 3 | Treatment 4 | Treatment 5 | |
|---|---|---|---|---|---|---|
| Nutrient | ||||||
| N | 0.18 (862) | 0.21 (1214) | 0.20 (1389) | 0.11 (485) | - | - |
| P | 0.10 (464) | 0.08 (455) | 0.16 (764) | 0.14 (909) | - | - |
| Vitamin B1 | 0.17 (545) | 0.12 (699) | 0.12 (561) | 0.12 (540) | - | - |
| Trace Metal | ||||||
| Fe | 0.14 (851) | 0.15 (1125) | 0.16 (1310) | 0.14 (852) | - | - |
| Cu | 0.17 (1055) | 0.14 (986) | 0.15 (1071) | 0.17 (1058) | - | - |
| Se | 0.17 (993) | 0.16 (1391) | 0.16 (1388) | 0.15 (1811) | 0.16 (2021) | 0.15 (1862) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, K.H.; Kim, H.J.; Li, Z.; Youn, J.Y.; Kwak, K.Y.; Seo, M.H.; Hwang, J.; Lee, S.D.; Yun, S.M.; Oh, S.J.; et al. Effects of Different Nutrient and Trace Metal Concentrations on Growth of the Toxic Dinoflagellate Gymnodinium catenatum Isolated from Korean Coastal Waters. Sustainability 2020, 12, 4992. https://doi.org/10.3390/su12124992
Han KH, Kim HJ, Li Z, Youn JY, Kwak KY, Seo MH, Hwang J, Lee SD, Yun SM, Oh SJ, et al. Effects of Different Nutrient and Trace Metal Concentrations on Growth of the Toxic Dinoflagellate Gymnodinium catenatum Isolated from Korean Coastal Waters. Sustainability. 2020; 12(12):4992. https://doi.org/10.3390/su12124992
Chicago/Turabian StyleHan, Kyong Ha, Hyun Jung Kim, Zhun Li, Joo Yeon Youn, Kyeong Yoon Kwak, Min Ho Seo, Jinik Hwang, Sang Deuk Lee, Suk Min Yun, Seok Jin Oh, and et al. 2020. "Effects of Different Nutrient and Trace Metal Concentrations on Growth of the Toxic Dinoflagellate Gymnodinium catenatum Isolated from Korean Coastal Waters" Sustainability 12, no. 12: 4992. https://doi.org/10.3390/su12124992
APA StyleHan, K. H., Kim, H. J., Li, Z., Youn, J. Y., Kwak, K. Y., Seo, M. H., Hwang, J., Lee, S. D., Yun, S. M., Oh, S. J., Park, J. W., Lim, W.-A., & Shin, H. H. (2020). Effects of Different Nutrient and Trace Metal Concentrations on Growth of the Toxic Dinoflagellate Gymnodinium catenatum Isolated from Korean Coastal Waters. Sustainability, 12(12), 4992. https://doi.org/10.3390/su12124992