Abstract
Sonchus oleraceus is becoming a hard-to-control weed in Australian cropping systems, especially in glyphosate-tolerant cotton and during summer fallows. Several biotypes of this weed have developed resistance to glyphosate as a result of common management practices under conservation agriculture systems in the country. A series of pot experiments were conducted to evaluate the effect of temperature on glyphosate efficacy and performance of several post-emergence and pre-emergence herbicides on a glyphosate-resistant (GR) and a glyphosate-susceptible (GS) biotype of S. oleraceus. At low temperatures (19–24 °C), no plants of the GS biotype survived glyphosate application at 570 g/ha; however, in the high-temperature regime (28–30 °C), 83% of the plants survived this rate of glyphosate. Similarly, for the GR biotype, up to 58% of the plants survived at 2280 g/ha of glyphosate when applied during the high-temperature regime and no plants survived this rate during the low-temperature regime. A number of post-emergence herbicides were found to be effective for S. oleraceus control. However, herbicide application delayed to the six-leaf stage compared with the four-leaf stage reduced control, especially for bromoxynil and saflufenacil herbicides. Glufosinate and paraquat were the most effective herbicides for S. oleraceus control, resulting in no seedling survival for both biotypes. Isoxaflutole, pendimethalin or s-metolachlor efficacy was not reduced by the presence of crop residue, suggesting that these herbicides could be used to control S. oleraceus in conservation agriculture systems. The results of this study suggest that growers will need to reduce over-reliance on glyphosate for weed control in summer fallows and use alternative post-emergence herbicides.
1. Introduction
Weeds are among the most important biological constraints to crop production in Australian agricultural systems. They cost Australian grain growers more than AUD 3.3 billion/year in yield losses and control measures. Sonchus oleraceus L. is a major weed of global importance and is threatening the sustainability of conservation agriculture systems of Australia. The national ranking of S. oleraceus is eighth in terms of acreage infested, with a revenue loss of AUD 5 million/annum [1].
In a recent study conducted in the state of Queensland, 43 to 52 plants/m2 caused a yield reduction of 50% in wheat [2]. In the same study, this weed produced up to 193,000 seeds/m2. S. oleraceus seeds are known to have little to no dormancy, enabling them to germinate immediately after dispersal [3]. More than two decades ago, it was mainly a winter weed, but now, it is present throughout the year, especially in the southeastern part of Australia. In competition with 164 plants/m2 of mungbean (a summer crop), each plant of S. oleraceus produced around 4000 seeds [4], suggesting that crop competition may not be able to provide effective control of this weed species. In another summer crop, soybean, 18 to 20 S. oleraceus plants/m2 caused a 50% yield loss [5]. High competitiveness, prolific seed production, low seed dormancy and wind-blown dispersal have led to an increased prevalence of this weed in conservation agriculture systems of Australia. Lack of tillage in these systems has favoured small-seeded species like S. oleraceus, which generally emerge at or close to the soil surface [6]. As a result, S. oleraceus is rapidly increasing in prevalence throughout the cotton-growing areas of subtropical Australia [6].
A fallow phase between two crops is common in Southeast Australia, which may last from 6 to 15 months, depending on soil moisture [7,8]. This fallow phase enables soils to accumulate moisture and nutrients but weeds growing in fallow fields can use a considerable amount of these resources, resulting in reduced yield potential of subsequent crops [9]. Therefore, glyphosate is commonly used to control these weeds during this fallow phase. However, the continuous use of this herbicide has led to the evolution of glyphosate-resistant biotypes of S. oleraceus [10]. Surveys in the glyphosate-tolerant cotton system also showed its prevalence as a hard-to-control weed using glyphosate [11]. Glyphosate efficacy can be affected by environmental factors, including temperature. For example, glyphosate efficacy was reduced on Echinochloa colona (L.) Link at 30 °C compared with 20 °C [12]. As S. oleraceus can emerge throughout the year, applications of glyphosate may experience different temperatures in different months. However, information on glyphosate efficacy on S. oleraceus when applied at different times of the year is very limited.
Once a weed biotype has developed resistance to glyphosate, it becomes very difficult to control it without using an alternative herbicide program. For example, different post-emergence herbicides provided up to 70% control of glyphosate-resistant Amaranthus rudis L., which was otherwise not controlled by glyphosate [13]. Therefore, there is a need to evaluate different post-emergence herbicides that provide effective control of S. oleraceus. Herbicide efficacy can be affected by weed growth stage. Setaria viridis (L.) Beauv. and Avena fatua L. control was greater when tralkoxydim was applied at the 2–3 leaf stage compared with the 4–5 leaf stage [14]. Similarly, the efficacy of different herbicides was found to be lower when applied at the eight-leaf stage of four grass weed species compared with the four or six-leaf stage [15]. These results indicate the importance of early herbicide application. However, such information is very limited on S. oleraceus.
As mentioned above, conservation agriculture systems are common in Australia, wherein S. oleraceus has become a problematic weed species. In these farming systems, crop residues are retained on the soil surface. Depending on rainfall predictions, growers usually apply pre-emergence herbicides to keep the field weed-free during the fallow phase or before crop planting. The crop residue present on the soil surface may adsorb a significant amount of pre-emergence herbicides, resulting in reduced herbicide efficacy [16,17,18,19]. A significant gap exists for information on the performance of pre-emergence herbicides on S. oleraceus under different amounts of crop residue. Knowledge gained from such studies can be used to develop effective weed management programs for S. oleraceus control in conservation agriculture systems.
A study was conducted to evaluate glyphosate efficacy at different times of the year, the performance of different post-emergence herbicides at different growth stages and the interaction effect of sorghum residue and pre-emergence herbicides on the emergence and biomass of S. oleraceus.
2. Materials and Methods
2.1. Seed Collection
Sonchus oleraceus seeds were collected from two locations: Gatton, Queensland (glyphosate-susceptible, GS) and Gunnedah, New South Wales (glyphosate-resistant, GR). Glyphosate at 720 g a.e./ha was sprayed at the 4-leaf stage of both biotypes (GR and GS) to confirm their resistance status. These biotypes were grown in the same condition at Gatton in 2017 and seeds collected from these plants were used in this study. All experiments were conducted in the Weed Science Screenhouse Facility of the University of Queensland at Gatton. Plants in all experiments were watered four times per day using an automated sprinkler system. However, watering was stopped 24 h before herbicide spray and continued 24 h after spray.
2.1.1. Experiment 1. Effect of Temperature on Glyphosate Efficacy
To determine the effect of temperature on glyphosate efficacy, a pot trial was conducted four times in 2019–20 in a naturally lit screenhouse. Temperature was not regulated in the screenhouse but average and minimum temperatures during the trial duration (planting to harvest) were measured (Table 1). Based on these temperatures, the four runs were assigned as high temperature (Run 1 and 2) and low temperature (Run 1 and 2) experiments (Table 1). In preliminary trials, this experiment was conducted in growth chambers set at different temperatures (low and high). However, the growth of S. oleraceus was very slow at low temperatures and plants were lanky. Therefore, this experiment was conducted in a screenhouse.

Table 1.
Dates (planting, spray and observation), temperature (minimum and average) and assigned details for the temperature and glyphosate resistance study.
Ten seeds of each biotype were planted in 20-cm diameter plastic pots containing standard potting mix (Centenary Landscaping, Mt Ommaney, Queensland). Immediately after emergence, seedlings were thinned to keep three plants per pot. At the 4–5 leaf stage, seedlings were treated with glyphosate at rates of 0 to 2280 g a.e./ha, with four replicates of each rate. Glyphosate was applied using a research track sprayer with a water volume of 108 L/ha. Standard flat fan nozzles (110015) were used in the sprayer. At 4 weeks after herbicide application, plant survival was recorded with the criterion of new green leaf tissues on treated plants. Surviving plants were cut at the surface level, placed in paper bags and dried in an oven at 70° C for 72 h. Biomass was weighed and converted to biomass/plant.
2.1.2. Experiment 2. Performance of Different Post-Emergence Herbicides
Fifteen seeds of each S. oleraceus biotype were planted in 20 cm diameter pots containing standard potting mix (Centenary Landscaping, Mt Ommaney, Queensland) and after emergence, eight seedlings/pot were maintained. Nine post-emergence herbicides were sprayed on S. oleraceus plants at two growth stages: 4-leaf and 6-leaf. The height of these plants was 7 to 9 cm and 11 to 13 cm, respectively. Each herbicide was applied at two rates (Table 2) and each treatment was replicated three times. There was a control treatment in which plants were not sprayed (non-treated control). Herbicides were applied as mentioned above. At 4 weeks after herbicide application, plant survival and biomass were determined as mentioned above. This experiment was conducted twice. An additional trial was conducted to evaluate the effect of different rates (0, 175, 350, 700, 1400 and 2800 g/ha) of 2,4-D on both biotypes of S. oleraceus. The herbicide was applied at the 4–5 leaf stage of S. oleraceus. Survival and biomass data were collected as described above. These experiments were conducted outdoors from March to August 2019, when the minimum and maximum average temperatures were 10.8° C and 26.0° C, respectively.
Table 2.
Post-emergence and pre-emergence herbicides and their rates used in Experiment 2 and 3.
2.1.3. Experiment 3. Effect of Sorghum Residue Amount on Efficacy of Pre-Emergence Herbicides
Twenty seeds of each S. oleraceus biotype were planted on the surface of potting mix in 20-cm diameter pots and covered with small pieces (1–2 cm) of sorghum plant residue (cv. MR Bazley) at rates equivalent to 0 (no cover), 3 and 6 tons/ha. These plant residues had been oven-dried at 70 °C for 72 h before use in this experiment. These pots were then irrigated using a sprinkler system. After 24 h, three pre-emergence herbicides were applied at two rates (Table 2). There was also a control treatment in which no herbicide was applied. Herbicides were applied as mentioned above in the temperature experiment. At 7 weeks after herbicide application, plant survival and biomass were determined as described above. In this experiment, the observation date was extended (7 weeks vs. 4 weeks for post-emergence herbicides) to allow plants to accumulate sufficient biomass. This experiment was conducted twice and each treatment was replicated three times in each run. Temperatures during the experimental duration are given in the previous section.
2.2. Statistical Analyses
All experiments were conducted using a randomized complete block design. The data for seedling emergence or plant survival and plant biomass were pooled over the two runs for Experiment 2 and 3 as there was no significant difference between the two experimental runs [20]. For the temperature experiment (Experiment 1), data are presented separately for each run to show the impact of temperature, occurring in different months, on glyphosate efficacy. The Fisher’s protected least significant difference (LSD) was used to determine the difference among means at the 5% level of significance.
3. Results and Discussion
3.1. Experiment 1. Effect of Temperature on Glyphosate Efficacy
As described above, this experiment was conducted four times in a screenhouse in different months. Two trials (Trial 1 and 4) experienced high average and high minimum temperatures (high-temperature runs) and the other two runs (Trial 2 and 3) experienced low average and low minimum temperatures (low-temperature runs) during their growing duration (Table 1).
In both runs in the low-temperature regime, there was no survival for the GS biotype at 570 g glyphosate/ha; however, in the high-temperature regime, 83% plants survived glyphosate application at this rate (Figure 1). Similarly, for the GR biotype, 42 to 58% plants survived the application of glyphosate at 2280 g/ha (three times the recommended rate) when applied during the high-temperature regime (Figure 1). However, no plants of the GR biotype survived this rate when glyphosate was applied during the low-temperature regime. Similar results were found for S. oleraceus biomass (Figure 2). The GR biotype produced 4 to 10% biomass of the control treatment at a glyphosate rate of 2280 g/ha during the high-temperature regime but no biomass was produced at this glyphosate rate during the low-temperature regime. Similarly, at the lower glyphosate rate (1140 g/ha) too, the GR biotype produced greater biomass during the high-temperature regime (12 to 15% of their non-treated control) compared with the low-temperature regime (4 to 5% of their non-treated control). The GS biotype did not produce biomass at a glyphosate rate of 570 g/ha during the low-temperature regime but produced 17 to 28% biomass of the non-treated control treatment during the high-temperature regime at that glyphosate rate.
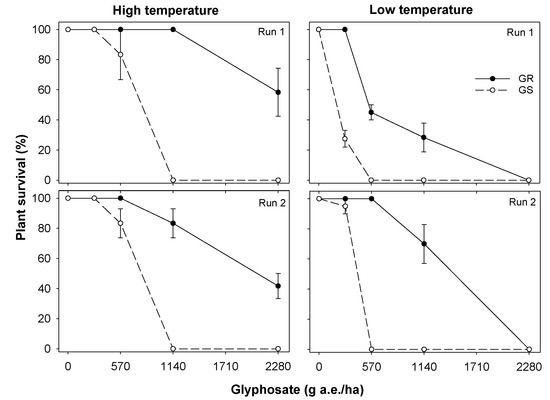
Figure 1.
Effect of temperature (high and low) and glyphosate rate on seedling survival of glyphosate-resistant (GR) and glyphosate-susceptible (GS) biotypes of Sonchus oleraceus. The vertical bars are the standard error of means (n = 3). These experiments were conducted in a screenhouse four times (high temperature—Run 1 and 2; low temperature—Run 1 and 2) in 2019/20.
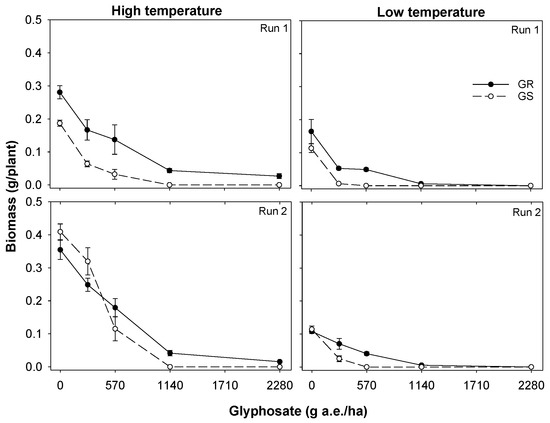
Figure 2.
Effect of temperature (high and low) and glyphosate rate on seedling biomass of glyphosate-resistant (GR) and glyphosate-susceptible (GS) biotypes of Sonchus oleraceus. The vertical bars are the standard error of means (n = 3). These experiments were conducted in a screenhouse four times (high temperature—Run 1 and 2; low temperature—Run 1 and 2) in 2019/20.
Undoubtedly, these results suggest that glyphosate efficacy is reduced at high temperatures. Similar results were reported for E. colona in South Australia, in which glyphosate resistance increased at 30 °C compared with 20 °C [12]. The authors suggested that a reason for this response at the high temperature was reduced glyphosate absorption, which would reduce the herbicide concentration in the leaf and its ability to enter the chloroplast. This explanation could be true for our study also; however, we did not evaluate the absorption and translocation of glyphosate. A recent study in the USA also reported that all the GR E. colona plants treated with glyphosate at 840 g/ha died when subjected to 15/10 °C alternating day/night temperature but the GR plants did not show any phytotoxicity at this herbicide rate when subjected to higher temperatures, 25/20 °C and 35/30 °C [21].
Another study on broadleaf weed species also indicated that the control of Chenopodium album L. and Conyza canadensis (L.) Cronquist by glyphosate could be reduced under projected future climatic conditions as both weed species were less sensitive to glyphosate under the higher temperature regime (32/26 °C) compared with the lower temperature regime (18/12 °C) [22]. These authors also suggested that altered glyphosate translocation might be the basis for reduced weed sensitivity at high temperatures. Our study was conducted in a naturally lit screenhouse, in which photoperiod and light intensity also differed during the four runs. Light intensity may affect the thickness of the leaf and therefore herbicide absorption and efficacy.
Sonchus oleraceus, considered mainly a winter weed in the past, is now a common weed throughout the year in Australia, especially in the Southeast Australian region, where rainfall is distributed throughout the year. In Queensland and New South Wales, growers usually grow one crop in a year, depending on rainfall; therefore, they need to manage weeds during the fallow phase. Due to sustainability of conservation agriculture systems, growers do not like to till their farm to control weeds and they rely on glyphosate during the fallow phase. Our results suggest that summer fallows will experience more glyphosate failures compared with winter fallows. S. oleraceus plants may survive glyphosate application during the summer months and produce seeds. As this weed can grow throughout the year, plants produced from these seeds may also become harder to control during the winter months, especially the GR populations. Therefore, over-reliance on glyphosate for weed control in summer fallows may result in more weed control failures [22], especially for weeds like S. oleraceus, which occur throughout the year.
Additionally, weather conditions in Southeast Australia are quite variable [23]. There can be hot days during autumn and winter months in this region, which may affect glyphosate efficacy. Therefore, growers need to check temperature conditions before glyphosate applications. Higher rates may be recommended to improve glyphosate activity on plants exposed to high temperatures.
3.2. Experiment 2. Performance of Different Post-Emergence Herbicides
As expected, glyphosate at both rates failed to provide effective control of the GR biotype (Table 3 and Table 4). All seedlings of the GS biotype were killed by glyphosate application when applied at the four-leaf stage but a delayed application to the six-leaf stage resulted in 36 to 88% seedling survival (Table 3). These seedlings, however, produced only 3 to 10% of the biomass in the non-treated control (Table 4). Similar results were reported for C. canadensis, which was found to be more susceptible to glyphosate at the seedling stage than at the large rosette stage [24].
Table 3.
Performance of post-emergence herbicides on seedling survival of glyphosate-resistant (GR) and glyphosate-susceptible (GS) biotypes of Sonchus oleraceus when applied at 4- and 6-leaf stages. The means (n = 6) were separated using least significant differences (LSD) at the 5% level of significance.
Table 4.
Performance of post-emergence herbicides on seedling biomass of glyphosate-resistant (GR) and glyphosate-susceptible (GS) biotypes of Sonchus oleraceus when applied at 4- and 6-leaf stages. The means (n = 6) were separated using LSD at the 5% level of significance. Values in parentheses are percent biomass reduction compared with the control treatment.
Irrespective of growth stage and herbicide rate, glufosinate and paraquat were the most effective post-emergence herbicides for S. oleraceus control, resulting in no seedling survival (Table 3) and biomass production (Table 4). In the USA, glufosinate has been reported as an alternative herbicide option for control of GR Ambrosia trifida L. [25]. A later study also reported glufosinate (700 g/ha) as an option for controlling GR Sorghum halepense L., with 77% control at soybean harvest [26]. An early study reported that control of C. album with glufosinate was most effective when applied at the 10-cm weed height compared to the 15-cm weed height [27]. In another study, glufosinate provided greater control (88%) of GR C. canadensis compared with paraquat (55 to 63%) [28]. Paraquat in our study provided excellent control of S. oleraceus but weeds may develop resistance to this herbicide due to restricted translocation [29]. Therefore, plants need to be monitored after paraquat application for possible escapes in the field.
No seedlings of S. oleraceus survived when 2,4-D + picloram, bromoxynil and saflufenacil were applied at the four-leaf stage. However, a significant number of seedlings survived when these herbicides were applied at the six-leaf stage. The results were more evident for bromoxynil, resulting in 100% seedling survival at both herbicide rates for both biotypes (Table 3). Compared with the non-treated control, bromoxynil at both rates reduced biomass by 27 to 46% for the GR biotype and 66 to 68% for the GS biotype (Table 4).
Similarly, late application of saflufenacil resulted in 49 to 89% seedling survival for the GR biotype and 64 to 71% for the GS biotype. However, these surviving plants did not grow very vigorously after herbicide application. Compared with the non-treated control, saflufenacil at both rates reduced 71 to 81% biomass for the GR biotype and 84 to 88% biomass for the GS biotype. A study from the USA suggested that saflufenacil could provide 90% control of rosette and bolted Parthenium hysterophorus L. at 6 to 27 g/ha, a range similar to our study [30]. The author also suggested that saflufenacil could be an effective burndown herbicide for control of P. hysterophorus populations resistant to glyphosate. In another study, the height of C. canadensis had little effect on saflufenacil efficacy, which provided 95 to 99% control of the weed when applied to small plants or >25-cm tall plants [31]. The authors suggested that the time of day of application had a greater effect on weed control with saflufenacil than weed height or weed density; however, we did not evaluate the effect of time of application on saflufenacil efficacy.
Irrespective of herbicide rate and growth stage, fluroxypyr and metsulfuron did not reduce plant survival compared with the non-treated control treatment (Table 3). These herbicide treatments resulted in 0 to 44% biomass reduction for fluroxypyr and 0 to 88% biomass reduction for metsulfuron (Table 4). The results show that there was a differential response between the two biotypes to metsulfuron. No reduction in biomass was observed for the GR biotype, indicating that the GR biotype may also be resistant to metsulfuron. Both fluroxypyr and metsulfuron are recommended for S. oleraceus control in Australia but they did not provide effective control of this weed. S. oleraceus biotypes resistant to chlorsulfuron (acetolactate synthase inhibitor) and 2,4-D (synthetic auxins) have been reported in Australia but no resistant biotypes to fluroxypyr (synthetic auxins) and metsulfuron (acetolactate synthase inhibitor) are known [10]. Our results suggest that there is a need to screen different populations of S. oleraceus against these herbicides to evaluate their resistance status.
Except for the higher herbicide application rate at the four-leaf stage for the GS biotype, 2,4-D also proved ineffective in reducing the survival percentage of S. oleraceus (Table 3). The effect of 2,4-D was more suppressive on S. oleraceus biomass than on its seedling survival. At the lower rate, 2,4-D application at the four-leaf stage reduced biomass by 34 and 71% for the GR and GS biotype, respectively (Table 4). This biomass was further reduced by 66 and 91% for the GR and GS biotypes, respectively, at the higher 2,4-D rate (i.e., 1050 g/ha). Delayed 2,4-D application, however, resulted in only 20 to 44% reduction in biomass, depending on biotype and 2,4-D rate.
As 2,4-D is used to manage a range of broadleaf weeds, its failure at the highest rate warranted a dose–response study. Fortunately, both biotypes were controlled completely at 1400 and 2800 g/ha of 2,4-D (data not shown). The herbicide 2,4-D on its own is not recommended for S. oleraceus control in fallows and our results support this recommendation. Therefore, 2,4-D needs to be mixed with other compatible herbicides to achieve complete control of S. oleraceus.
In general, a delayed herbicide application reduced S. oleraceus control. This was particularly true for bromoxynil and saflufenacil. Our study found a number of alternative herbicide options to control GR biotypes of S. oleraceus; however, these herbicides need to be applied at an early stage to achieve effective weed control. Growers tend to delay herbicide application to maximise weed seedling emergence from the seed bank so that all the weeds can be treated at the same time but this tendency may result in the build-up of herbicide-resistant weed seed banks.
3.3. Experiment 3. Effect of Sorghum Residue Amount on Efficacy of Pre-Emergence Herbicides
The interaction effect of herbicide treatments and sorghum residue amount was significant for the seedling emergence (Table 5) and biomass (Table 6) of both biotypes of S. oleraceus. In the non-treated control treatment, covering the seeds with sorghum residue did not affect the seedling emergence or biomass of both biotypes. Compared with the non-treated control, across residue amount, only a few herbicide treatments were able to reduce seedling emergence but a greater number of herbicide treatments were able to reduce seedling biomass. These results suggest that seedlings that survived herbicide application were not able to grow vigorously.
Table 5.
Interaction of pre-emergence herbicides and sorghum residue amounts (0, 3 and 6 t/ha) on seedling emergence of glyphosate-resistant (GR) and glyphosate-susceptible (GS) biotypes of Sonchus oleraceus. The means (n = 6) were separated using LSD at the 5% level of significance.
Table 6.
Interaction of pre-emergence herbicides and sorghum residue amounts (0, 3 and 6 t/ha) on seedling biomass of glyphosate-resistant (GR) and glyphosate-susceptible (GS) biotypes of Sonchus oleraceus. The means (n = 6) were separated using LSD at the 5% level of significance. Values in parentheses are percent biomass reduction compared with the control treatment.
Isoxaflutole at 75 g/ha provided effective control of seedling emergence of both biotypes when applied without crop residue cover; however, the addition of sorghum residue reduced the efficacy of isoxaflutole at this rate, resulting in an increased amount of seedling emergence (Table 5). Isoxaflutole at 150 g/ha was required to overcome the problem of increased seedling emergence with the addition of crop residue. These differential responses were not found for seedling biomass of both biotypes (Table 6). Seedling biomass was similar across both rates of isoxafluotole. Regardless of sorghum residue amount, herbicide rate and weed biotype, isoxaflutole provided 81 to 100% suppression of S. oleraceus biomass.
Isoxaflutole is a soil-applied isoxazole herbicide used for the control and suppression of selective broadleaf and grass weeds in sugarcane, chickpea and fallows by inhibiting the 4-hydroxyphenyl-pyruvate-dioxygenase (4-HPPD) biochemical pathway. In a previous study, lower degradation of isoxaflutole was found under conservation tillage than under conventional tillage [32]. Our results suggest that the presence of crop residue in conservation agriculture systems will not affect isoxaflutole efficacy on S. oleraceus biomass. In fallows, surviving seedlings may grow and produce seeds if follow-up treatments are not applied. S. oleraceus seedlings may emerge following isoxaflutole application but may not be able to produce enough biomass in competitive sugarcane or chickpea crops. In the USA, isoxaflutole was found to be an option for use in maize, especially for atrazine-resistant weeds [33]. In Australia, this herbicide needs to be evaluated for weed control in maize; however, a carryover from isoxaflutole applications in maize may require plant back restrictions for certain sensitive crops [34].
Pendimethalin at 910 g/ha did not affect the seedling emergence of either biotype (Table 5). Increasing the rate to 1820 g/ha also did not affect seedling emergence in the no-residue treatment; however, integration of pendimethalin at 1820 g/ha with residue retention resulted in decreased seedling emergence of both biotypes compared with the non-treated control treatments. The response was different between the biotypes for seedling biomass (Table 6). Pendimethalin application resulted in significant reductions (81 to 94%) in the seedling biomass of the GR biotype compared with the non-treated control treatments and there was no difference between the two rates of the herbicide. For the GS biotype, however, seedling biomass was similar between the non-treated control and pendimethalin treatments. These differential responses of the two biotypes to pendimethalin suggest a need to evaluate the performance of this herbicide on several biotypes of S. oleraceus.
Pendimethalin is a soil-applied dinitroaniline herbicide used for the control of some broadleaf and grass weeds by inhibiting mitosis. A study from the USA suggested that the use of biochar as a soil amendment could decrease pendimethalin efficacy by adsorbing a significant amount of the herbicide [35]. In another study similar to our study, pendimethalin application (1 and 2 kg ai/ha) in the presence of rice residue cover resulted in lower control of Cyperus iria L. than in the absence of residue15. This study suggested that some weed species may escape pendimethalin application in conservation agriculture systems. However, such adverse results of the crop residue on pendimethalin efficacy were not found in our study with S. oleraceus.
The herbicide s-metolachlor at 960 g/ha did not affect the seedling emergence of either biotype at different residue levels compared to their respective non-treated control treatments (Table 5). However, increasing the herbicide rate to 1920 g/ha resulted in decreased emergence in the no-residue cover condition. Covering the seeds with residue cover negated the effect of this herbicide rate on seedling emergence. A similar response was found for seedling biomass; however, both biotypes behaved differently (Table 6). Application of s-metolachlor resulted in greater suppression of biomass for the GR biotype compared with the GS biotype. Biomass of both biotypes was similar across s-metolachlor rates.
S-metolachlor is a soil-applied amide herbicide used for the control of some broadleaf and grass weeds in certain crops by inhibiting very long-chain fatty acid biosynthesis. In a recent study, using sorghum residue resulted in lower s-metolachlor efficacy on E. colona and Chloris virgata Sw., grass weed species [19]. In our study, s-metolachlor efficacy was not reduced by the addition of sorghum residue. Differential responses between the two studies could be due to the different irrigation systems used in these studies. Plants in the previous study were sub-irrigated while plants in the current study were irrigated using an overhead sprinkler system. A sprinkler irrigation system could have resulted in washing off the herbicide from crop residue, which was then available to S. oleraceus. A study from the USA suggested that integration of high-residue cover crops with s-metolachlor increased Amaranthus species control [36], suggesting that residue retention in conservation agriculture systems may not reduce the efficacy of s-metolachlor.
4. Conclusions
Temperature influenced the efficacy of glyphosate on both biotypes (GR and GS) of S. oleraceus. At high temperatures, glyphosate rates had to be increased to achieve a similar level of control obtained at low temperatures using low glyphosate rates. Therefore, growers may need to increase glyphosate rates for weed control in summer fallows to preserve the sustainability of conservation agriculture systems. Over-reliance on glyphosate for control of fallow-weeds needs to be reduced to avoid weed control failures in the future. There is a need to evaluate the efficacy of glyphosate on a range of GR weeds at different temperature regimes. This study also found a range of alternative herbicides for control of S. oleraceus, especially for GR biotypes. However, these herbicides need to be applied at an early stage to achieve effective control. The efficacy of pre-emergence herbicides was mostly not affected by the presence of crop residue, suggesting that effective control of S. oleraceus can be obtained with pre-emergence herbicides in high crop residue retention systems. Surviving plants, however, need to be managed with other weed management tools to avoid the build-up of weed seed banks. This trial was conducted in pots using a potting mix under an automated sprinkler system. Performance of soil-active herbicides is greatly affected by soil moisture and soil type; therefore, there is a need to evaluate the performance of these herbicides in field conditions.
Author Contributions
Conceptualization, B.S.C.; data curation, B.S.C.; formal analysis, B.S.C.; funding acquisition, B.S.C.; methodology, B.S.C.; resources, B.S.C.; writing—original draft, B.S.C.; writing—review and editing, P.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors have declared that no competing interests exist.
References
- Llewellyn, R.; Ronning, D.; Clarke, M.; Mayfield, A.; Walker, S.; Ouzman, J. Impact of Weeds on Australian Grain Production: The Cost of Weeds to Australian Grain Growers and the Adoption of Weed Management and Tillage Practices; Report for GRDC; CSIRO: Canberra, ACT, Australia, 2016; p. 112. [Google Scholar]
- Manalil, S.; Ali, H.H.; Chauhan, B.S. Interference of annual sowthistle (Sonchus oleraceus) in wheat. Weed Sci. 2020, 68, 98–103. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Gill, G.; Preston, C. Factors affecting seed germination of annual sowthistle (Sonchus oleraceus) in southern Australia. Weed Sci. 2006, 54, 854–860. [Google Scholar] [CrossRef]
- Mobli, A.; Matloob, A.; Chauhan, B.S. The response of glyphosate-resistant and glyphosate-susceptible biotypes of annual sowthistle (Sonchus oleraceus) to mungbean density. Weed Sci. 2019, 67, 642–648. [Google Scholar] [CrossRef]
- Song, J.-S.; Kim, J.W.; Im, J.H.; Lee, K.J.; Lee, B.W.; Kim, D.S. The effects of single- and multiple-weed interference on soybean yield in the far-eastern region of Russia. Weed Sci. 2017, 65, 371–380. [Google Scholar] [CrossRef]
- Manalil, S.; Werth, J.; Jackson, R.; Chauhan, B.S.; Preston, C. An assessment of weed flora 14 years after the introduction of glyphosate-tolerant cotton in Australia. Crop Past. Sci. 2017, 68, 773–780. [Google Scholar] [CrossRef]
- Thomas, G.; Felton, W.; Radford, B. Tillage and crop residue management. In Sustainable Crop Production in the Sub-Tropics: An Australian Perspective; Clarke, A.L., Wylie, P.B., Eds.; Queensland Department of Primary Industries: Roma, QLD, Australia, 1997; pp. 195–213. [Google Scholar]
- Webb, A.; Grundy, M.; Powell, B.; Littleboy, M. The Australian sub-tropical cereal belt: Soils, climate and agriculture. In Sustainable Crop Production in the Sub-Tropics: An Australian Perspective; Clarke, A.L., Wylie, P.B., Eds.; Queensland Department of Primary Industries: Roma, QLD, Australia, 1997; pp. 8–23. [Google Scholar]
- Widderick, M.; Sindel, B.; Walker, S. Distribution, importance and management of Sonchus oleraceus (common sowthistle) in the northern cropping region of Australia. In Proceedings of the 12th Australian Weeds Conference, Hobart, TAS, Australia, 12–16 September 1999; p. 198. [Google Scholar]
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 1 June 2020).
- Manalil, S.; Coast, O.; Werth, J.; Chauhan, B.S. Weed management in cotton (Gossypium hirsutum L.) through weed-crop competition: A review. Crop Prot. 2017, 95, 53–59. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Malone, J.M.; Boutsalis, P.; Shirley, N.; Preston, C. Temperature influences the level of glyphosate resistance in barnyardgrass (Echinochloa colona). Pest Manag. Sci. 2016, 72, 1031–1039. [Google Scholar] [CrossRef]
- Sarangi, D.; Sandell, L.D.; Kruger, G.R.; Knezevic, S.Z.; Irmak, S.; Jhala, A.J. Comparison of herbicide programs for season-long control of glyphosate-resistant common waterhemp (Amaranthus rudis) in soybean. Weed Technol. 2017, 31, 53–66. [Google Scholar] [CrossRef]
- Harker, K.N.; Blackshaw, R.E. Influence of growth stage and broadleaf herbicides on tralkoxydim activity. Weed Sci. 1991, 39, 650–659. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Abugho, S.B. Effect of growth stage on the efficacy of postemergence herbicides on four weed species of direct-seeded rice. Sci. World J. 2012, 2012, 7. [Google Scholar] [CrossRef]
- Banks, P.A.; Robinson, E.L. The influence of straw mulch on the soil reception and persistence of metribuzin. Weed Sci. 1982, 30, 164–168. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Abugho, S.B. Interaction of rice residue and PRE herbicides on emergence and biomass of four weed species. Weed Technol. 2012, 26, 627–632. [Google Scholar] [CrossRef]
- Khalil, Y.; Flower, K.; Siddique, K.H.M.; Ward, P. Rainfall affects leaching of pre-emergent herbicide from wheat residue into the soil. PLoS ONE 2019, 14, e0210219. [Google Scholar] [CrossRef] [PubMed]
- Mobli, A.; Rinwa, A.; Sahil; Chauhan, B.S. Effects of sorghum residue in presence of pre-emergence herbicides on emergence and biomass of Echinochloa colona and Chloris virgata. PLoS ONE 2020, 15, e0229817. [Google Scholar] [CrossRef] [PubMed]
- Genstat for Windows, 20th ed.; VSN International: Hemel Hempstead, UK, 2019.
- Shrestha, A.; Budhathoki, S.; Steinhauer, K. Temperature effects on glyphosate resistance in California populations of junglerice. Agron. J. 2018, 110, 1624–1626. [Google Scholar] [CrossRef]
- Matzrafi, M.; Brunharo, C.; Tehranchian, P.; Hanson, B.D.; Jasieniuk, M. Increased temperatures and elevated CO2 levels reduce the sensitivity of Conyza canadensis and Chenopodium album to glyphosate. Sci. Rep. 2019, 9, 2228. [Google Scholar] [CrossRef]
- Ali, H.H.; Kebaso, L.; Manalil, S.; Chauhan, B.S. Emergence and germination response of Sonchus oleraceus and Rapistrum rugosum to different temperatures and moisture stress regimes. Plant Species Biol. 2020, 35, 16–23. [Google Scholar] [CrossRef]
- VanGessel, M.J.; Scott, B.A.; Johnson, Q.R.; White-Hansen, S.E. Influence Of glyphosate-resistant horseweed (Conyza canadensis) growth stage on response to glyphosate applications. Weed Technol. 2009, 23, 49–53. [Google Scholar] [CrossRef]
- Kaur, S.; Sandell, L.D.; Lindquist, J.L.; Jhala, A.J. Glyphosate-resistant giant ragweed (Ambrosia trifida) control in glufosinate-resistant soybean. Weed Technol. 2014, 28, 569–577. [Google Scholar] [CrossRef]
- Landry, R.L.; Stephenson, D.O.; Woolam, B.C. Glufosinate rate and timing for control of glyphosate-resistant rhizomatous Johnsongrass (Sorghum halepense) in glufosinate-resistant soybean. Int. J. Agron. 2016, 8040235. [Google Scholar] [CrossRef]
- Steckel, G.J.; Wax, L.M.; Simmons, F.W.; Phillips, W.H. Glufosinate efficacy on annual weeds is influenced by rate and growth stage. Weed Technol. 1997, 11, 484–488. [Google Scholar] [CrossRef]
- Eubank, T.W.; Poston, D.H.; Nandula, V.K.; Koger, C.H.; Shaw, D.R.; Reynolds, D.B. Glyphosate-resistant horseweed (Conyza canadensis) control using glyphosate-, paraquat-, and glufosinate-based herbicide programs. Weed Technol. 2008, 22, 16–21. [Google Scholar] [CrossRef]
- Yu, Q.; Cairns, A.; Powles, S.B. Paraquat resistance in a population of Lolium rigidum. Funct. Plant Biol. 2004, 31, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Odero, D.C. Response of ragweed parthenium (Parthenium hysterophorus) to saflufenacil and glyphosate. Weed Technol. 2012, 26, 443–448. [Google Scholar] [CrossRef]
- Budd, C.M.; Soltani, N.; Robinson, D.E.; Hooker, D.C.; Miller, R.T.; Sikkema, P.H. Efficacy of saflufenacil for control of glyphosate-resistant horseweed (Conyza canadensis) as affected by height, density, and time of day. Weed Technol. 2017, 65, 275–284. [Google Scholar] [CrossRef]
- Alletto, L.; Coquet, Y.; Bergheaud, V.; Benoit, P. Water pressure head and temperature impact on isoxaflutole degradation in crop residues and loamy surface soil under conventional and conservation tillage management. Chemosphere 2012, 88, 1043–1050. [Google Scholar] [CrossRef]
- Stephenson, D.O.; Bond, J.A. Evaluation of thiencarbazone-methyl- and isoxaflutole-based herbicide programs in corn. Weed Technol. 2012, 26, 37–42. [Google Scholar] [CrossRef]
- Nelson, E.A.; Penner, D. Sensitivity of selected crops to isoxaflutole in soil and irrigation water. Weed Technol. 2005, 19, 659–663. [Google Scholar] [CrossRef]
- Soni, N.; Leon, R.G.; Erickson, J.E.; Ferrell, J.A.; Silveira, M.L. Biochar decreases atrazine and pendimethalin preemergence herbicidal activity. Weed Technol. 2015, 29, 359–366. [Google Scholar] [CrossRef]
- Price, A.J.; Balkcom, K.S.; Duzy, L.M.; Kelton, J.A. Herbicide and cover crop residue integration for Amaranthus control in conservation agriculture cotton and implications for resistance management. Weed Technol. 2012, 26, 490–498. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).