
Biodegradable Material for Oyster Reef Restoration: First-Year Performance and Biogeochemical Considerations in a Coastal Lagoon


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Approach
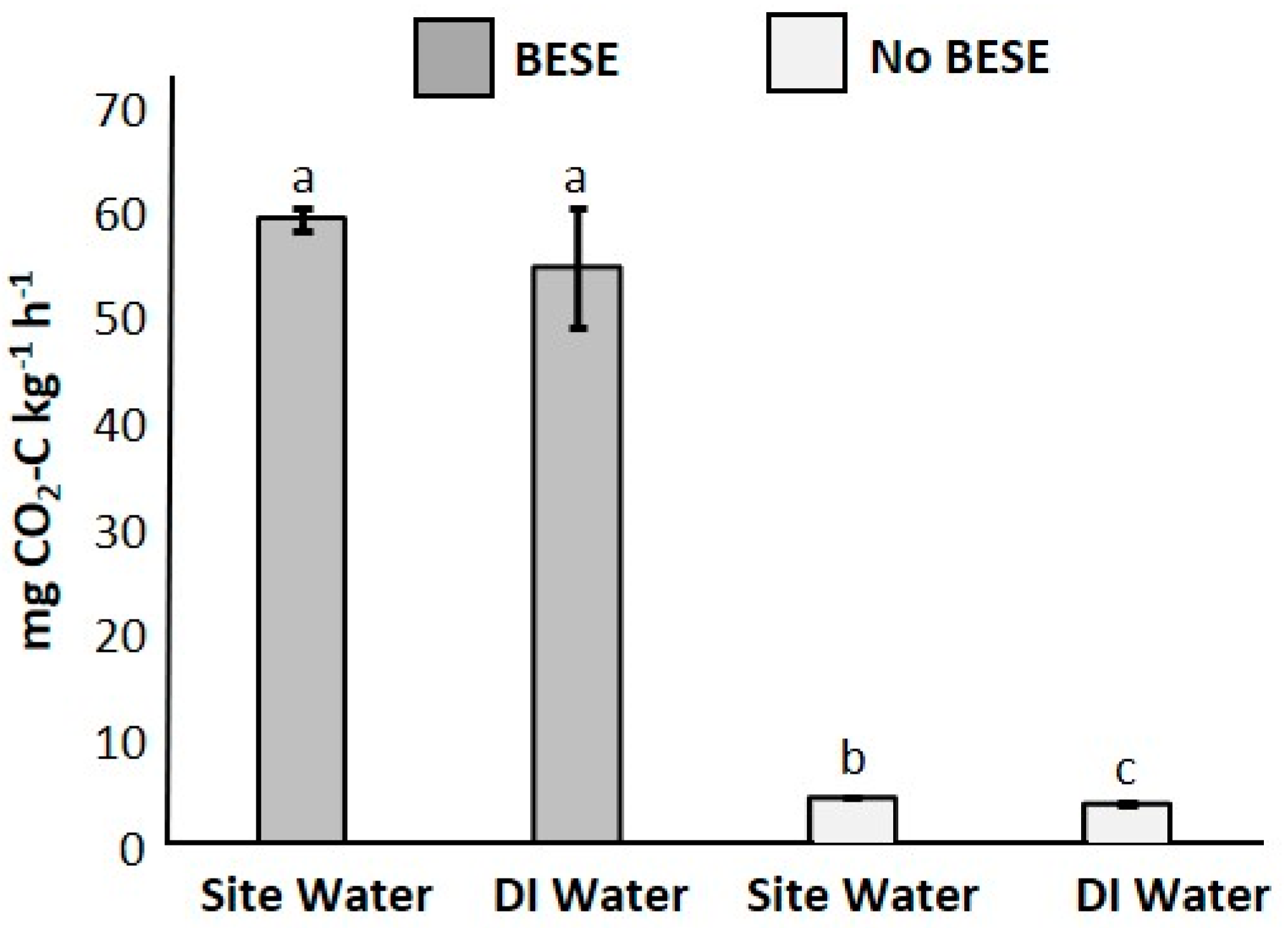
2.2. Laboratory Respiration Experiment
Statistical Approach for Respiration Experiment
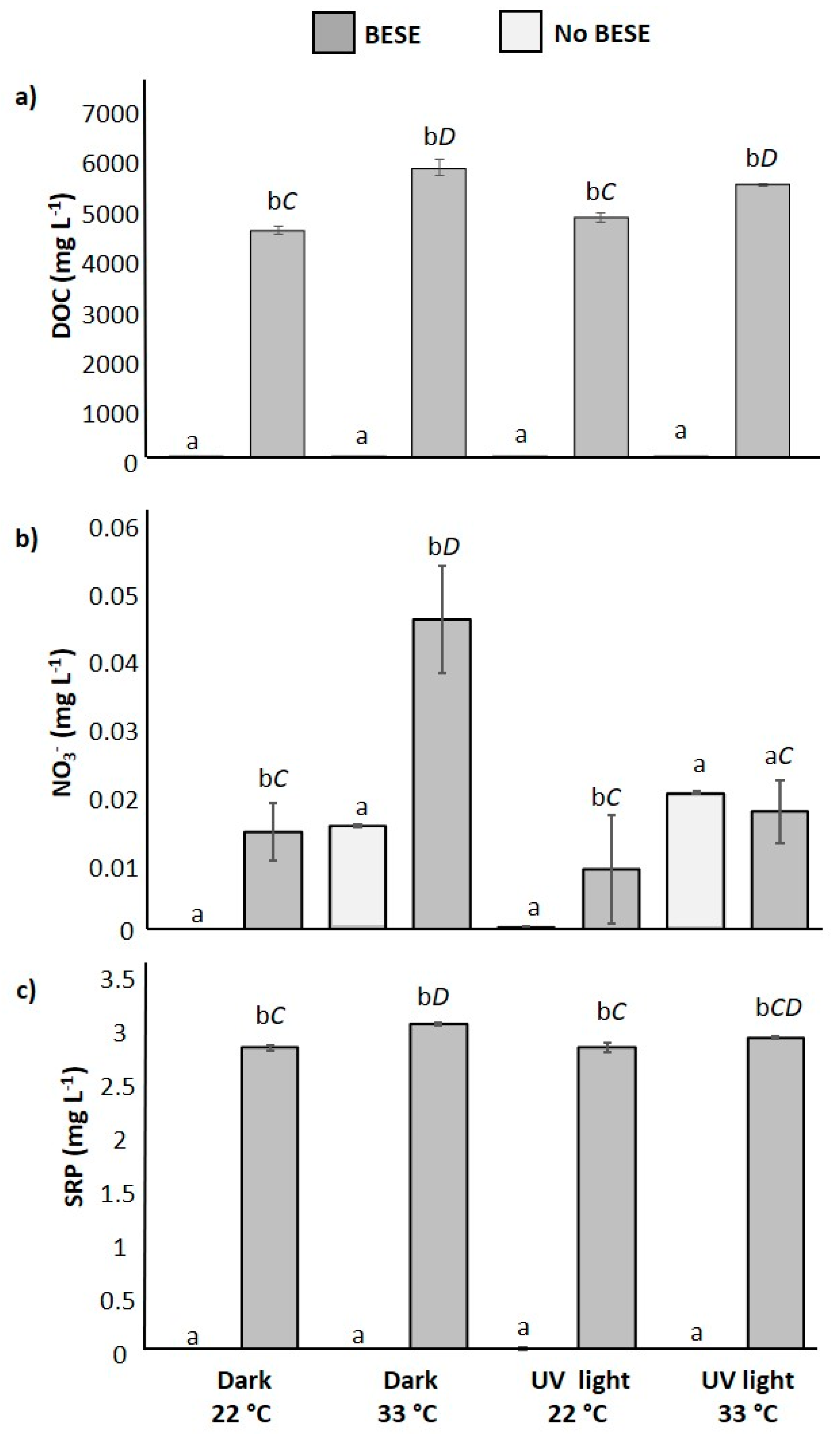
2.3. Laboratory Nutrient Release Experiment
Statistical Approach for Nutrient Release Experiment
2.4. Field BACI Experimental Design
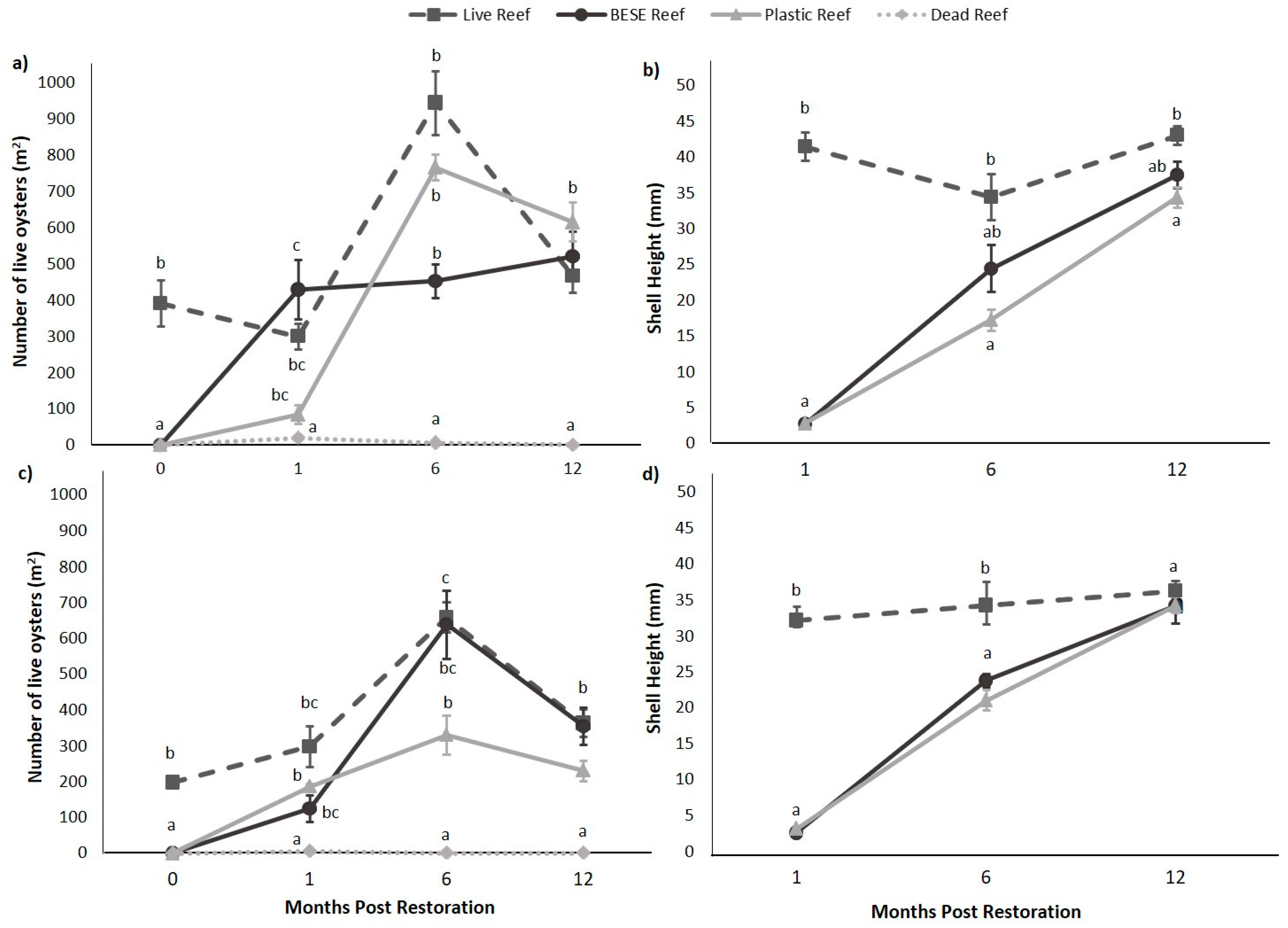
2.4.1. Field BACI Experiment—Oyster Monitoring
Statistical Approach for Oyster Monitoring
2.4.2. Field BACI Experiment—Sediment Biogeochemistry
Statistical Approach for Sediment Biogeochemistry
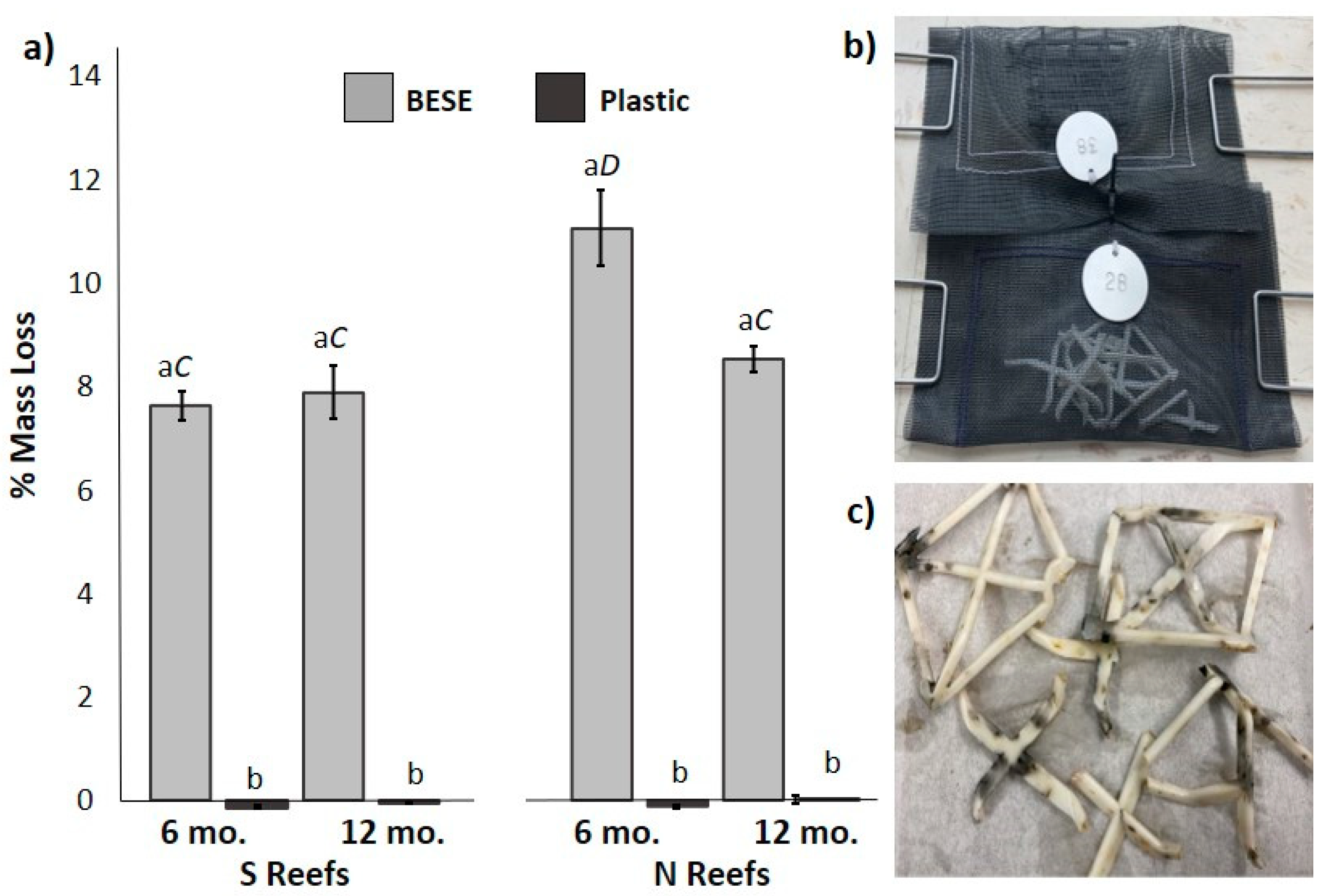
2.5. Field BESE Degradation Experiment
Statistical Approach for Degradation Experiment
2.6. Laboratory Analysis of Aqueous Nutrients
2.7. Laboratory Analysis of Sediment Properties
3. Results
3.1. Laboratory Respiration Experiment
3.2. Laboratory Nutrient Release Experiment
3.3. Field BACI Experiment
3.4. Field BESE Degradation Experiment
4. Discussion
4.1. BESE Impacts on Biogeochemical Properties and Processes under Laboratory Conditions
4.2. Performance of BESE under Field Conditions
4.3. Consideration for Wide-Scale Adoption of Biodegradable Material for Oyster Reef Restoration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Encomio, V.G.; Stickler, S.M.; Allen, S.K.; Chu, F.L. Performance of “natural dermo-resistant” oyster stocks-Survival, disease, growth, condition and energy reserves. J. Shellfish Res. 2005, 24, 143–155. [Google Scholar] [CrossRef]
- Carlsson, J.; Carnegie, R.B.; Cordes, J.F.; Hare, M.P.; Leggett, A.T.; Reece, K.S. Evaluating recruitment contribution of a selectively bred aquaculture line of the oyster, Crassostrea virginica used in restoration efforts. J. Shellfish Res. 2008, 27, 1117–1124. [Google Scholar] [CrossRef] [Green Version]
- Garvis, S.K.; Sacks, P.E.; Walters, L.J. Formation, Movement, and Restoration of Dead Intertidal Oyster Reefs in Canaveral National Seashore and Mosquito Lagoon, Florida. J. Shellfish Res. 2015, 34, 251–258. [Google Scholar] [CrossRef]
- Walters, L.J.; Sacks, P.E.; Campbell, D.E. Boating impacts and boat-wake resilient restoration of the eastern oyster Crassostrea virginica in Mosquito Lagoon, Florida, USA. Florida Sci. 2021, in press. [Google Scholar]
- Herbert, D.; Astrom, E.; Bersoza, A.C.; Batzer, A.; McGovern, P.; Angelini, C.; Wasman, S.; Dix, N.; Sheremet, A. Mitigating erosional effects induced by boat wakes with living shorelines. Sustainability 2018, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Safak, I.; Norby, P.L.; Dix, N.; Grizzle, R.E.; Southwell, M.; Veenstra, J.J.; Acevedo, A.; Cooper-Kolb, T.; Massey, L.; Sheremet, A.; et al. Coupling breakwalls with oyster restoration structures enhances living shoreline performance along energetic shorelines. Ecol. Eng. 2020, 158, 106071. [Google Scholar] [CrossRef]
- Gilby, B.L.; Olds, A.D.; Chapman, S.; Goodridge Gaines, L.A.; Henderson, C.J.; Ortodossi, N.L.; Didderen, K.; Lengkeek, W.; van der Heide, T.; Schlacher, T.A. Attraction versus production in restoration: Spatial and habitat effects of shellfish reefs for fish in coastal seascapes. Restor. Ecol. 2021. [Google Scholar] [CrossRef]
- Johnson, E.E.; Medina, M.D.; Hernandez, A.C.B.; Kusel, G.A.; Batzer, A.N.; Angelini, C. Success of concrete and crab traps in facilitating Eastern oyster recruitment and reef development. PeerJ 2019, 2019, 1–19. [Google Scholar] [CrossRef]
- Scyphers, S.B.; Powers, S.P.; Heck, K.L. Ecological Value of Submerged Breakwaters for Habitat Enhancement on a Residential Scale. Environ. Manag. 2015, 55, 383–391. [Google Scholar] [CrossRef]
- Dunn, R.P.; Eggleston, D.B.; Lindquist, N. Oyster-sponge interactions and bioerosion of reef-building substrate materials: Implications for oyster restoration. J. Shellfish Res. 2014, 33, 727–738. [Google Scholar] [CrossRef]
- Moody, J.A.; Gentry, M.J.; Bouboulis, S.A.; Kreeger, D.A. Effects of Substrate (Protection and Type) on Ribbed Mussel (Geukensia demissa) Recruitment for Living Shoreline Applications. J. Coast. Res. 2020, 36, 619–627. [Google Scholar] [CrossRef]
- George, L.M.; De Santiago, K.; Palmer, T.A.; Beseres Pollack, J. Oyster reef restoration: Effect of alternative substrates on oyster recruitment and nekton habitat use. J. Bus. Psychol. 2015, 30, 13–22. [Google Scholar] [CrossRef]
- Soniat, T.M.; Burton, G.M. A comparison of the effectiveness of sandstone and limestone as cultch for oysters, Crassostrea virginica. J. Shellfish Res. 2005, 24, 483–485. [Google Scholar] [CrossRef]
- Quan, W.; Fan, R.; Wang, Y.; Humphries, A.T. Long-Term Oyster Recruitment and Growth are not Influenced by Substrate Type in China: Implications for Sustainable Oyster Reef Restoration. J. Shellfish Res. 2017, 36, 79–86. [Google Scholar] [CrossRef]
- Frederick, P.; Vitale, N.; Pine, B.; Seavey, J.; Sturmer, L. Reversing a Rapid Decline in Oyster Reefs: Effects of Durable Substrate on Oyster Populations, Elevations, and Aquatic Bird Community Composition. J. Shellfish Res. 2016, 35, 359–367. [Google Scholar] [CrossRef]
- Soucy, B. Alternative Material Selection for Oyster Restoration with an Emphasis on Living Docks. Master’s Thesis, Florida Institute of Technology, Melbourne, FL, USA, 2020; p. 91. [Google Scholar]
- Morris, R.L.; Bilkovic, D.M.; Boswell, M.K.; Bushek, D.; Cebrian, J.; Goff, J.; Kibler, K.M.; La Peyre, M.K.; McClenachan, G.; Moody, J.; et al. The application of oyster reefs in shoreline protection: Are we over-engineering for an ecosystem engineer? J. Appl. Ecol. 2019, 56, 1703–1711. [Google Scholar] [CrossRef]
- McClenachan, G.M.; Donnelly, M.J.; Shaffer, M.N.; Sacks, P.E.; Walters, L.J. Does size matter? Quantifying the cumulative impact of small-scale living shoreline and oyster reef restoration projects on shoreline erosion. Restor. Ecol. 2020, 28, 1365–1371. [Google Scholar] [CrossRef]
- Goelz, T.; Vogt, B.; Hartley, T. Alternative Substrates Used for Oyster Reef Restoration: A Review. J. Shellfish Res. 2020, 39, 1–12. [Google Scholar] [CrossRef]
- Chambers, L.G.; Gaspar, S.A.; Pilato, C.J.; Steinmuller, H.E.; McCarthy, K.J.; Sacks, P.E.; Walters, L.J. How Well Do Restored Intertidal Oyster Reefs Support Key Biogeochemical Properties in a Coastal Lagoon? Estuaries Coasts 2018, 41, 784–799. [Google Scholar] [CrossRef]
- Shaffer, M.; Donnelly, M.; Walters, L. Does intertidal oyster reef restoration affect avian community structure and behaivor in a shallow estuarine system? A post-restoration analysis. Florida Field Nat. 2019, 47, 37–98. [Google Scholar]
- Lewis, D.; Durham, K.; Walters, L.; Cook, G. Resident fishes as higher trophic level indicators of oyster reef restoration success. Estuaries Coasts. in review.
- Harris, K. Oyster Reef Restoration: Impacts on Infaunal Communities in a Shallow Water Estuary. Univ. Cent. Florida Undergrad. Res. J. 2020, 11, 7–17. [Google Scholar]
- Copertino, J. Bird Abundance and Diversity and the Impact of Oyster Reef Restoration on the Bird Community in Mosquito Lagoon, Florida, USA. Master’s Thesis, University of Central Florida, Orlando, FL, USA, 2021; p. 106. [Google Scholar]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Cole, M.; Lindeque, P.; Halsband, C.; Galloway, T.S. Microplastics as contaminants in the marine environment: A review. Mar. Pollut. Bull. 2011, 62, 2588–2597. [Google Scholar] [CrossRef]
- Waite, H.R.; Donnelly, M.J.; Walters, L.J. Quantity and types of microplastics in the organic tissues of the eastern oyster Crassostrea virginica and Atlantic mud crab Panopeus herbstii from a Florida estuary. Mar. Pollut. Bull. 2018, 129, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Walters, L.; Craig, C. Microplastics, Oysters, and the Indian River Lagoon–Final Report to IRL NEP; CEELAB Research Data 4; University of Central Florida Libraries: Orlando, FL, USA, 2021. [Google Scholar]
- Jambeck, J.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. The Ocean. Mar. Pollut. 2015, 347, 768. [Google Scholar]
- Guo, X.; Wang, J. The chemical behaviors of microplastics in marine environment: A review. Mar. Pollut. Bull. 2019, 142, 1–14. [Google Scholar] [CrossRef]
- Carlin, J.; Craig, C.; Little, S.; Donnelly, M.; Fox, D.; Zhai, L.; Walters, L. Microplastic accumulation in the gastrointestinal tracts in birds of prey in central Florida, USA. Environ. Pollut. 2020, 264, 114633. [Google Scholar] [CrossRef]
- Zhang, D.; Cui, Y.; Zhou, H.; Jin, C.; Yu, X.; Xu, Y.; Li, Y.; Zhang, C. Microplastic pollution in water, sediment, and fish from artificial reefs around the Ma’an Archipelago, Shengsi, China. Sci. Total Environ. 2020, 703, 134768. [Google Scholar] [CrossRef]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Goïc, N.L.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [Green Version]
- Dybas, C.L. Florida’s Indian River Lagoon: An Estuary in Transition. Bioscience 2002, 52, 554–559. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Lasi, M.A.; Chamberlain, R.; Green, W.C.; Hall, L.M.; Hart, J.A.; Lockwood, J.C.; Miller, J.D.; Morris, L.J.; et al. From Red Tides to Green and Brown Tides: Bloom Dynamics in a Restricted Subtropical Lagoon Under Shifting Climatic Conditions. Estuaries Coasts 2015, 38, 886–904. [Google Scholar] [CrossRef]
- Smith, N.P. Tidal and nontidal flushing of Florida’s Indian River Lagoon. Estuaries 1993, 16, 739–746. [Google Scholar] [CrossRef]
- McClenachan, G.; Witt, M.; Walters, L.J. Replacement of oyster reefs by mangroves: Unexpected climate-driven ecosystem shifts. Glob. Chang. Biol. 2020, 1–13. [Google Scholar] [CrossRef]
- Redfield, A.C. The biolgoical control of chemical factors in the environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- BESE. Ecosystem Restoration Products. Available online: http://www.bese-products.com/ (accessed on 18 May 2021).
- Locher, B.; Hurst, N.R.; Walters, L.J.; Chambers, L.G. Juvenile Oyster (Crassostrea virginica) Biodeposits Contribute to a Rapid Rise in Sediment Nutrients on Restored Intertidal Oyster Reefs (Mosquito Lagoon, FL, USA). Estuaries Coasts 2020, 44. [Google Scholar] [CrossRef]
- Wieder, R.K.; Lang, G.E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef] [Green Version]
- Ashton, E.C.; Hogarth, P.J.; Ormond, R. Breakdown of mangrove leaf litter in a managed mangrove forest in Peninsular Malaysia. Hydrobiologia 1999, 413, 77–88. [Google Scholar] [CrossRef]
- USEPA. Methods for the Determination of Inorganic Substances in Environmental Samples; US Environmental Protection Agency: Washington, DC, USA, 1993. [Google Scholar]
- Andersen, J.M. An ignition method for determination of total phosphorus in lake sediments. Water Res. 1976, 10, 329–331. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Duga, E.; Witt, M.; Filyaw, R.; Friedland, N.; Donnelly, M.J.; Walters, L.J.; Chambers, L.G. Carbon and nutrient fluxes from seagrass and mangrove wrack are mediated by soil interactions. Estuar. Coast. Shelf Sci. 2019, 229. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- Blagodatsky, S.; Blagodatskaya, E.; Yuyukina, T.; Kuzyakov, Y. Model of apparent and real priming effects: Linking microbial activity with soil organic matter decomposition. Soil Biol. Biochem. 2010, 42, 1275–1283. [Google Scholar] [CrossRef]
- Fontaine, S.; Bardoux, G.; Abbadie, L.; Mariotti, A. Carbon input to soil may decrease soil carbon content. Ecol. Lett. 2004, 7, 314–320. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Blodau, C.; Canadel, J.G.; Freeman, C.; Holden, J.; Roulet, N.; Rydin, H.; Schaepman-Strub, G. Peatlands and the carbon cycle: From local processes to global implications- a synthesis. Biogeosciences 2008, 5, 1739. [Google Scholar] [CrossRef] [Green Version]
- Katsoyiannis, A.; Samara, C. The fate of Dissolved Organic Carbon (DOC) in the wastewater treatment process and its importance in the removal of wastewater contaminants. Environ. Sci. Pollut. Res. 2007, 14, 284–292. [Google Scholar] [CrossRef]
- Lønborg, C.; Carreira, C.; Jickells, T.; Álvarez-Salgado, X.A. Impacts of Global Change on Ocean Dissolved Organic Carbon (DOC) Cycling. Front. Mar. Sci. 2020, 7, 1–24. [Google Scholar] [CrossRef]
- Benner, R.; Amon, R.M.W. The size-reactivity continuum of major bioelements in the Ocean. Ann. Rev. Mar. Sci. 2015, 7, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.; Gilbert, P.; Burkholder, J. Harmful Algal Blooms and Eutrophication Nutrient Sources, Composition, and Consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Strock, K.E.; Theodore, N.; Gawley, W.G.; Ellsworth, A.C.; Saros, J.E. Increasing dissolved organic carbon concentrations in northern boreal lakes: Implications for lake water transparency and thermal structure. J. Geophys. Res. Biogeosci. 2017, 122, 1022–1035. [Google Scholar] [CrossRef]
- Katsoyiannis, A.; Samara, C. Persistent organic pollutants (POPs) in the conventional activated sludge treatment process: Fate and mass balance. Environ. Res. 2005, 97, 245–257. [Google Scholar] [CrossRef]
- Evans, C.D.; Monteith, D.T.; Cooper, D.M. Long-term increases in surface water dissolved organic carbon: Observations, possible causes and environmental impacts. Environ. Pollut. 2005, 137, 55–71. [Google Scholar] [CrossRef] [PubMed]
- EPA. Water Quality Standards for the State of Florida’s Lakes and Flowing Waters. Fed. Regist. 40 CFR Part 131 2010, 75, 75761–75807. [Google Scholar]
- Reddy, K.R.; DeLaune, R. Biogeochemistry of Wetlands: Science and Applications; CRC Press: New York, NY, USA, 2008. [Google Scholar]
- Dame, R.; Spurrier, J.; Wolaver, T. Carbon, nitrogen and phosphorus processing by an oyster reef. Mar. Ecol. Prog. Ser. 1989, 54, 249–256. [Google Scholar] [CrossRef]
- Kellogg, M.L.; Smyth, A.R.; Luckenbach, M.W.; Carmichael, R.H.; Brown, B.L.; Cornwell, J.C.; Piehler, M.F.; Owens, M.S.; Dalrymple, D.J.; Higgins, C.B. Use of oysters to mitigate eutrophication in coastal waters. Estuar. Coast. Shelf Sci. 2014, 151, 156–168. [Google Scholar] [CrossRef]
- Fodrie, F.J.; Rodriguez, A.B.; Gittman, R.K.; Grabowski, J.H.; Lindquist, N.L.; Peterson, C.H.; Piehler, M.F.; Ridge, J.T. Oyster reefs as carbon sources and sinks. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Kitsikoudis, V.; Kibler, K.M.; Walters, L.J. In-situ measurements of turbulent flow over intertidal natural and degraded oyster reefs in an estuarine lagoon. Ecol. Eng. 2020, 143, 105688. [Google Scholar] [CrossRef]
- Newell, R.I.E.; Fisher, T.R.; Holyoke, R.R.; Cornwell, J.C. Influence of Eastern Oysters on Nitrogen and Phosphorus Regeneration in Chesapeake Bay, USA. In The Comparative Roles of Suspension Feeders in Ecosystems; Dame, R., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 47, pp. 93–120. ISBN 1568-1238. [Google Scholar]
- Smyth, A.R.; Murphy, A.E.; Anderson, I.C.; Song, B. Differential Effects of Bivalves on Sediment Nitrogen Cycling in a Shallow Coastal Bay. Estuaries Coasts 2018, 41, 1147–1163. [Google Scholar] [CrossRef] [Green Version]
- Dame, R.; Dankers, N.; Prins, T.; Jongsma, H.; Smaal, A. The Influence of Mussel Beds on Nutrients in the Western Wadden Sea and Eastern Scheldt Estuaries. Estuaries 1991, 14, 130–138. [Google Scholar] [CrossRef]
- Dame, R.F.; Spurrier, J.D.; Zingmark, R.G. In situ metabolism of an oyster reef. J. Exp. Mar. Bio. Ecol. 1992, 164, 147–159. [Google Scholar] [CrossRef]
- Newell, R.I.E.; Cornwell, J.C.; Owens, M.S. Influence of simulated bivalve biodeposition and microphytobenthos on sediment nitrogen dynamics: A laboratory study. Limnol. Oceanogr. 2002, 47, 1367–1379. [Google Scholar] [CrossRef]
- Bayne, B.L.; Hawkins, A.J.S. Ecological and Physiological Aspects of Herbivory in Benthic Suspension-Feeding Molluscs. In Plant-Animal Interactions in the Marine Benthos; John, D., Hawkins, S., Price, J., Eds.; Clarendon Press: Oxford, UK, 1992; pp. 265–288. [Google Scholar]
- Chae, H.M.; Choi, S.H.; Lee, S.H.; Cha, S.; Yang, K.C.; Shim, J.K. Effect of litter quality on needle decomposition for four pine species in Korea. Forests 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Melillo, J.M.; Aber, J.D.; Linkins, A.E.; Ricca, A.; Fry, B.; Nadelhoffer, K.J. Carbon and nitrogen dynamics along the decay continuum: Plant litter to soil organic matter. Plant Soil 1989, 115, 189–198. [Google Scholar] [CrossRef]
- Stagg, C.L.; Baustian, M.M.; Perry, C.L.; Carruthers, T.J.B.; Hall, C.T. Direct and indirect controls on organic matter decomposition in four coastal wetland communities along a landscape salinity gradient. J. Ecol. 2018, 106, 655–670. [Google Scholar] [CrossRef]
- Loría-Naranjo, M.; Sibaja-Cordero, J.A.; Cortés, J. Mangrove Leaf Litter Decomposition in a Seasonal Tropical Environment. J. Coast. Res. 2019, 35, 122–129. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Wall, L.M.; Walters, L.J.; Grizzle, R.E.; Sacks, P.E. Recreational boating activity and its impact on the recruitment and survival of the oyster Crassostrea virginica on intertidal reefs in Mosquito Lagoon, Florida. J. Shellfish Res. 2005, 24, 965–973. [Google Scholar] [CrossRef]
- Grizzle, R.; Adams, J.; Walters, L. Historical changes in intertidal oyster (Crassostrea virginica) reefs in a Florida lagoon potentially related to boating activities. J. Shellfish Res. 2002, 21, 749–756. [Google Scholar]
- Temmink, R.J.M.; Christianen, M.J.A.; Fivash, G.S.; Angelini, C.; Boström, C.; Didderen, K.; Engel, S.M.; Esteban, N.; Gaeckle, J.L.; Gagnon, K.; et al. Mimicry of emergent traits amplifies coastal restoration success. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Primary Research Question | Primary Hypothesis |
|---|---|---|
| Laboratory respiration | Can BESE serve as a substrate for microbial respiration? | CO2 production will increase with the addition of BESE. |
| Laboratory nutrient release | Can BESE release labile nutrients given idealized laboratory conditions? | Dissolved inorganic N, P, and organic C will increase when BESE is incubated under warm temperatures and UV light. |
| Field BACI study—oyster monitoring | Are oyster reef restoration success metrics comparable between BESE and plastic materials? | Oyster density will be the same on BESE and plastic restored reefs at 12 months post-restoration. |
| Field BACI study—sediment biogeochemistry | Does the rate or degree to which biogeochemical hot spots develop under recently restored oyster reefs differ between BESE and plastic materials? | BESE will accelerate the development of biogeochemical hot spots in sediments beneath oyster reefs. |
| Field BESE degradation | What is the initial rate of mass loss for BESE material under field conditions? | Significant mass loss of BESE will be observed within the first year of field deployment. |
| Month | ||||
|---|---|---|---|---|
| 0 | 1 | 6 | 12 | |
| OM (g kg−1) | 89.9 ± 5.3 a | 86.2 ± 4.7 a,b | 94.6 ± 5.5 b | 109.4 ± 4.6 c |
| DBD (g cm−3) | 1.02 ± 0.04 a | 0.92 ± 0.04 a | 1.03 ± 0.04 ab | 0.90 ± 0.03 b |
| Total P (g kg−1) | 0.81 ± 0.02 | |||
| Total C (g kg−1) | 40.1 ± 1.24 | |||
| Total N (g kg−1) | 1.58 ± 0.11 a | 1.50 ± 0.11 a,b | 1.75 ± 0.10 b | 2.33 ± 0.12 c |
| Extr. DOC (g kg−1) | 71.8 ± 6.4 a | 77.2 ± 6.4 a,b | 57.8 ± 3.5 b | 87.8 ± 6.5 c |
| Extr. NO3− (g kg−1) | 1.83 ± 0.11 a | 0.51 ± 0.05 b | 1.05 ± 0.08 c | 2.42 ± 0.10 a,b |
| Extr. NH4+ (g kg−1) | 1.1 ± 0.2 a | 0.9 ± 0.4(D) 9.0 ± 1.0(L,B,P) b | 1.7 ± 0.4 c | 0.9 ± 0.5(D) 9.7 ± 1.0(L,B,P) c |
| Extr. SRP (g kg−1) | 0.81 ± 0.11 a | 0.67 ± 0.10 a,b | 0.40 ± 0.06 b,c | 0.97 ± 0.12 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nitsch, C.K.; Walters, L.J.; Sacks, J.S.; Sacks, P.E.; Chambers, L.G. Biodegradable Material for Oyster Reef Restoration: First-Year Performance and Biogeochemical Considerations in a Coastal Lagoon. Sustainability 2021, 13, 7415. https://doi.org/10.3390/su13137415
Nitsch CK, Walters LJ, Sacks JS, Sacks PE, Chambers LG. Biodegradable Material for Oyster Reef Restoration: First-Year Performance and Biogeochemical Considerations in a Coastal Lagoon. Sustainability. 2021; 13(13):7415. https://doi.org/10.3390/su13137415
Chicago/Turabian StyleNitsch, Chelsea K., Linda J. Walters, Joshua S. Sacks, Paul E. Sacks, and Lisa G. Chambers. 2021. "Biodegradable Material for Oyster Reef Restoration: First-Year Performance and Biogeochemical Considerations in a Coastal Lagoon" Sustainability 13, no. 13: 7415. https://doi.org/10.3390/su13137415
APA StyleNitsch, C. K., Walters, L. J., Sacks, J. S., Sacks, P. E., & Chambers, L. G. (2021). Biodegradable Material for Oyster Reef Restoration: First-Year Performance and Biogeochemical Considerations in a Coastal Lagoon. Sustainability, 13(13), 7415. https://doi.org/10.3390/su13137415