
Legume Nitrogen Fixation and Symbioses in Low-Inputs Rainfed Rice Rotations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
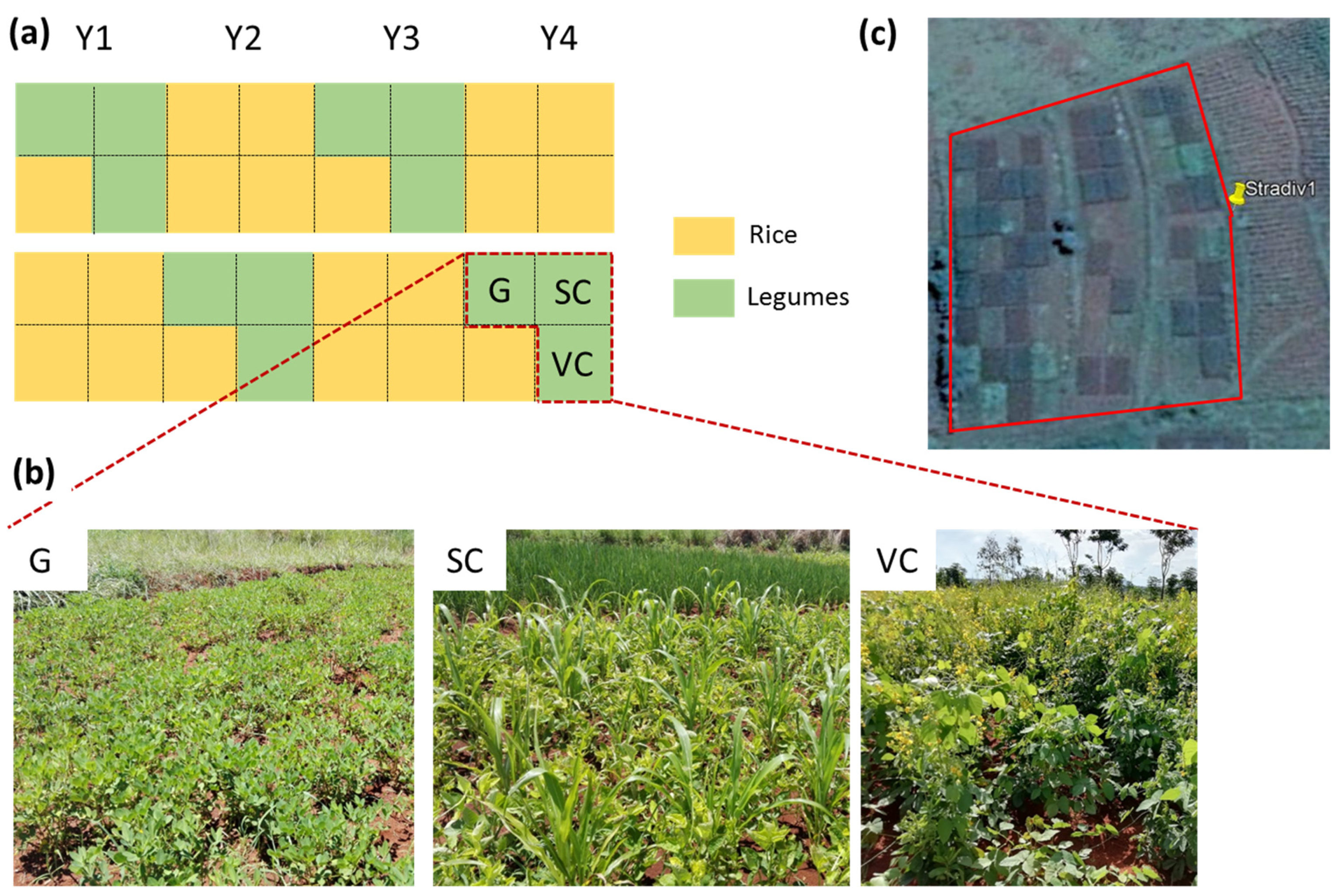
2.1. Study Site, Experimental Design, and Crop Management
2.2. Plant Sampling and Analyses
2.3. Statistical Analysis
3. Results
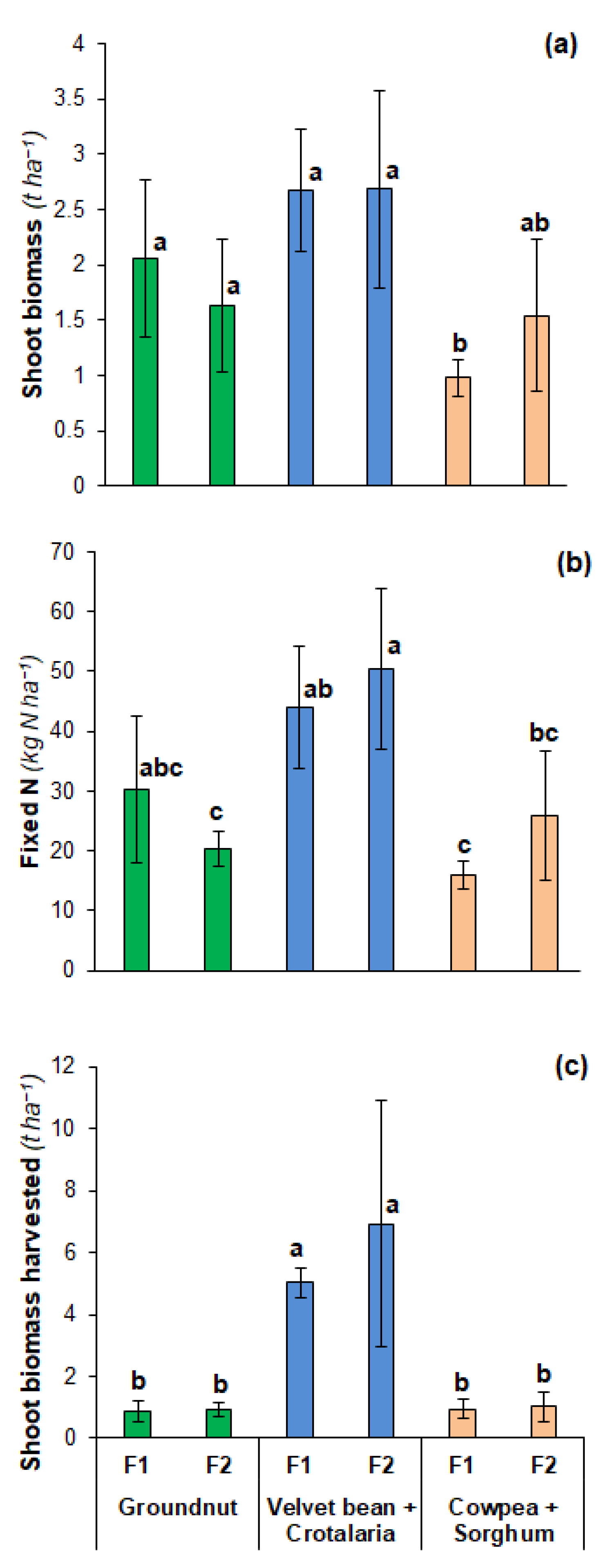
3.1. Legume Growth and Biological N2 Fixation
3.2. Legume Symbioses
3.3. Legume Response to Symbioses
3.4. Biomass Production and N Restitution at the Plot Scale
4. Discussion
4.1. Importances of Rhizobial and AMF Symbionts on Plant N2 Fixation in Acid Ferralsols
4.2. Amount of N Fixed by Legumes in SSA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scopel, E.; Triomphe, B.; Affholder, F.; Da Silva, F.A.M.; Corbeels, M.; Xavier, J.H.V.; Lahmar, R.; Recous, S.; Bernoux, M.; Blanchart, E.; et al. Conservation agriculture cropping systems in temperate and tropical conditions, performances and impacts. A review. Agron. Sustain. Dev. 2013, 33, 113–130. [Google Scholar] [CrossRef]
- Kleijn, D.; Bommarco, R.; Fijen, T.P.; Garibaldi, L.A.; Potts, S.G.; van der Putten, W.H. Ecological intensification: Bridging the gap between science and practice. Trends Ecol. Evol. 2019, 34, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Beillouin, D.; Ben-Ari, T.; Makowski, D. Evidence map of crop diversification strategies at the global scale. Environ. Res. Lett. 2019, 14, 123001. [Google Scholar] [CrossRef]
- Sauvadet, M.; Trap, J.; Damour, G.; Plassard, C.; Van den Meersche, K.; Achard, R.; Allinne, C.; Autfray, P.; Bertrand, I.; Blanchart, E.; et al. Agroecosystem diversification with legumes or non-legumes improves differently soil fertility according to soil type. Sci. Total Environ. 2021, 795, 148934. [Google Scholar] [CrossRef] [PubMed]
- Mango, N.; Makate, C.; Mapemba, L.; Sopo, M. The role of crop diversification in improving household food security in central Malawi. Agric. Food Secur. 2018, 7, 7. [Google Scholar] [CrossRef]
- ten Berge, H.F.; Hijbeek, R.; van Loon, M.P.; Rurinda, J.; Tesfaye, K.; Zingore, S.; Craufurde, P.; van Heerwaardenb, J.; Brentrupf, F.; Schrödera, J.J.; et al. Maize crop nutrient input requirements for food security in sub-Saharan Africa. Glob. Food Secur. Agric. Policy 2019, 23, 9–21. [Google Scholar] [CrossRef]
- Tittonell, P.; Giller, K.E. When yield gaps are poverty traps: The paradigm of ecological intensification in African smallholder agriculture. Field Crop. Res. 2013, 143, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Chianu, J.N.; Chianu, J.N.; Mairura, F. Mineral fertilizers in the farming systems of sub-Saharan Africa. A review. Agron. Sustain. Dev. 2012, 32, 545–566. [Google Scholar] [CrossRef] [Green Version]
- Franke, A.C.; Van den Brand, G.J.; Vanlauwe, B.; Giller, K.E. Sustainable intensification through rotations with grain legumes in Sub-Saharan Africa: A review. Agric. Ecosyst. Environ. 2018, 261, 172–185. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Hungria, M.; Kanampiu, F.; Giller, K.E. The role of legumes in the sustainable intensification of African smallholder agriculture: Lessons learnt and challenges for the future. Agric. Ecosyst. Environ. 2019, 284, 106583. [Google Scholar] [CrossRef]
- Liu, L.; Knight, J.D.; Lemke, R.L.; Farrell, R.E. A side-by-side comparison of biological nitrogen fixation and yield of four legume crops. Plant Soil 2019, 442, 169–182. [Google Scholar] [CrossRef]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. Appl. Physiol. Nutr. Metab. 2014, 39, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Vaz Patto, M.C.; Amarowicz, R.; Aryee, A.N.; Boye, J.I.; Chung, H.J.; Martín-Cabrejas, M.A.; Domoney, C. Achievements and challenges in improving the nutritional quality of food legumes. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Duchene, O.; Vian, J.F.; Celette, F. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric. Ecosyst. Environ. 2017, 240, 148–161. [Google Scholar] [CrossRef]
- Kocira, A.; Staniak, M.; Tomaszewska, M.; Kornas, R.; Cymerman, J.; Panasiewicz, K.; Lipińska, H. Legume cover crops as one of the elements of strategic weed management and soil quality improvement. A review. Agriculture 2020, 10, 394. [Google Scholar] [CrossRef]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of legume-based grassland–livestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef] [PubMed]
- Farrow, A.; Ronner, E.; Van Den Brand, G.J.; Boahen, S.K.; Leonardo, W.; Wolde-Meskel, E.; Adjei-Nsiah, S.; Chikowo, R.; Baijukya, F.; Ebanyat, P.; et al. From best fit technologies to best fit scaling: Incorporating and evaluating factors affecting the adoption of grain legumes in sub-Saharan Africa. Exp. Agric. 2019, 55, 226–251. [Google Scholar] [CrossRef] [Green Version]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Sahrawat, K.L.; Upadhyaya, H.D.; Mengoni, A.; Galardini, M.; Bazzicalupo, M.; Biondi, E.G.; Hungria, M.; Kaschuk, G.; Blair, M.W.; et al. Advances in host plant and rhizobium genomics to enhance symbiotic nitrogen fixation in grain legumes. Adv. Agron. 2015, 129, 1–116. [Google Scholar]
- Augusto, L.; Delerue, F.; Gallet-Budynek, A.; Achat, D.L. Global assessment of limitation to symbiotic nitrogen fixation by phosphorus availability in terrestrial ecosystems using a meta-analysis approach. Glob. Biogeochem. Cycle 2013, 27, 804–815. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Nitrogen fixation in perspective: An overview of research and extension needs. Field Crop. Res. 2000, 65, 93–106. [Google Scholar] [CrossRef]
- Larimer, A.L.; Bever, J.D.; Clay, K. The interactive effects of plant microbial symbionts: A review and meta-analysis. Symbiosis 2010, 51, 139–148. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Razakatiana, A.T.E.; Trap, J.; Baohanta, R.H.; Raherimandimby, M.; Le Roux, C.; Duponnois, R.; Ramanankierana, H.; Becquer, T. Benefits of dual inoculation with arbuscular mycorrhizal fungi and rhizobia on Phaseolus vulgaris planted in a low-fertility tropical soil. Pedobiologia 2020, 83, 150685. [Google Scholar] [CrossRef]
- Deckers, J. Soil fertility and environmental problems in different ecological zones of the developing countries in Sub-Saharan Africa. In The Role of Plant Nutrients for Sustainable Food Crop Production in Sub-Saharan Africa; Van Reuler, H., Prins, R.W., Eds.; Vereniging Van Kunstmest Producenten: Wageningen, The Netherlands, 1993; pp. 37–52. [Google Scholar]
- Ripoche, A.; Autfray, P.; Blanchart, E.; Rabary, B.; Randriamanantsoa, R.; Trap, J.; Sauvadet, M.; Becquer, T.; Letourmy, P. Increasing plant diversity promotes ecosystem functions in rainfed rice based short rotations in Malagasy highlands. Agric. Ecosyst. Environ. 2021, 320, 107576. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; Vandenboom, T.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Okalebo, R.J.; Gathua, K.W.; Woomer, P.L. Laboratory Methods of Soil and Plant Analysis: A working Manual, 2nd ed.; Sacred Africa: Nairobi, Kenya, 2002. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Unkovich, M.; Herridge, D.; Peoples, M.; Cadisch, G.; Boddey, B.; Giller, K.; Alves, B.; Chalk, P. Measuring Plant-Associated Nitrogen Fixation in Agricultural Systems; Australian Centre for International Agricultural Research: Canberra, Australia, 2008; 258p. [Google Scholar]
- Samago, T.Y.; Anniye, E.W.; Dakora, F.D. Grain yield of common bean (Phaseolus vulgaris L.) varieties is markedly increased by rhizobial inoculation and phosphorus application in Ethiopia. Symbiosis 2018, 75, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Okito, A.; Alves, B.R.J.; Urquiaga, S.; Boddey, R.M. Isotopic fractionation during N2 fixation by four tropical legumes. Soil Biol. Biochem. 2004, 36, 1179–1190. [Google Scholar] [CrossRef]
- Nyemba, R.C.; Dakora, F.D. Evaluating N2 fixation by food grain legumes in farmers’ fields in three agro-ecological zones of Zambia, using 15N natural abundance. Biol. Fertil. Soils 2010, 46, 461–470. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piche, Y. Ink and vinegar, a simple staining technique for arbuscular mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA Press: Paris, France, 1986; pp. 217–221. [Google Scholar]
- White, H. A heteroskedasticity-consistent covariance matrix estimator and a direct test for heteroskedasticity. Econometrica 1980, 48, 817–838. [Google Scholar] [CrossRef]
- Pule-Meulenberg, F.; Belane, A.K.; Krasova-Wade, T.; Dakora, F.D. Symbiotic functioning and bradyrhizobial biodiversity of cowpea (Vigna unguiculata L. Walp.) in Africa. BMC Microbiol. 2010, 10, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belane, A.K.; Asiwe, J.; Dakora, F.D. Assessment of N2 fixation in 32 cowpea (Vigna unguiculata L. Walp) genotypes grown in the field at Taung in South Africa, using 15N natural abundance. Afr. J. Biotechnol. 2011, 10, 11450–11458. [Google Scholar]
- Unkovich, M.J.; Baldock, J.; Peoples, M.B. Prospects and problems of simple linear models for estimating symbiotic N2 fixation by crop and pasture legumes. Plant Soil 2010, 329, 75–89. [Google Scholar] [CrossRef]
- Blesh, J. Functional traits in cover crop mixtures: Biological nitrogen fixation and multifunctionality. J. Appl. Ecol. 2018, 55, 38–48. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; Hammond, J.P. Phosphorus nutrition of terrestrial plants. In The Ecophysiology of Plant-Phosphorus Interactions; White, P.J., Hammond, J.P., Eds.; Springer: Amsterdam, The Netherlands, 2008; Volume 7, pp. 51–81. [Google Scholar]
- Raminoarison, M.; Razafimbelo, T.; Rakotoson, T.; Becquer, T.; Blanchart, E.; Trap, J. Multiple-nutrient limitation of upland rainfed rice in ferralsols: A greenhouse nutrient-omission trial. J. Plant Nutr. 2020, 43, 270–284. [Google Scholar] [CrossRef]
- Gérard, F. Clay minerals, iron/aluminum oxides, and their contribution to phosphate sorption in soils-A myth revisited. Geoderma 2016, 262, 213–226. [Google Scholar] [CrossRef]
- Kouas, S.; Labidi, N.; Debez, A.; Abdelly, C. Effect of P on nodule formation and N fixation in bean. Agron. Sustain. Dev. 2005, 25, 389–393. [Google Scholar] [CrossRef]
- Santachiara, G.; Salvagiotti, F.; Rotundo, J.L. Nutritional and environmental effects on biological nitrogen fixation in soybean: A meta-analysis. Field Crop. Res. 2019, 240, 106–115. [Google Scholar] [CrossRef]
- Peoples, M.B.; Brockwell, J.; Hunt, J.R.; Swan, A.D.; Watson, L.; Hayes, R.C.; Li, G.D.; Hackney, G.B.; Nuttall, J.G.; Davies, S.L.; et al. Factors affecting the potential contributions of N2 fixation by legumes in Australian pasture systems. Crop Pasture Sci. 2012, 63, 759–786. [Google Scholar] [CrossRef]
- Sanginga, N.; Ibewiro, B.; Houngnandan, P.; Vanlauwe, B.; Okogun, J.A.; Akobundu, I.O.; Versteeg, M. Evaluation of symbiotic properties and nitrogen contribution of mucuna to maize grown in the derived savanna of West Africa. Plant Soil 1996, 179, 119–129. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Nwoke, O.C.; Diels, J.; Sanginga, N.; Carsky, R.J.; Deckers, J.; Merckx, R. Utilization of rock phosphate by crops on a representative toposequence in the Northern Guinea savanna zone of Nigeria: Response by Mucuna pruriens, Lablab purpureus and maize. Soil Biol. Biochem. 2000, 32, 2063–2077. [Google Scholar] [CrossRef]
- Bagayoko, M.; Buerkert, A.; Lung, G.; Bationo, A.; Römheld, V. Cereal/legume rotation effects on cereal growth in Sudano-Sahelian West Africa: Soil mineral nitrogen, mycorrhizae and nematodes. Plant Soil 2000, 218, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi: A model and field test. New Phytol. 2002, 155, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.; Johnson, D.E. Legumes as dry season fallow in upland rice-based systems of West Africa. Biol. Fertil. Soils 1998, 27, 358–367. [Google Scholar] [CrossRef]
- Rose, T.J.; Kearney, L.J.; Erler, D.V.; Rose, M.T.; Van Zwieten, L.; Raymond, C.A. Influence of growth stage and seed nitrogen on B values and potential contributions to error in estimating biological N2 fixation using the 15N natural abundance method. Plant Soil 2018, 425, 389–399. [Google Scholar] [CrossRef]
- Pacheco, R.S.; Boddey, R.M.; Alves, B.J.R.; Straliotto, R.; Araújo, A.P. Growth patterns of common bean cultivars affect the ‘B’value required to quantify biological N2 fixation using the 15N nat-ural abundance technique. Plant Soil 2017, 419, 293–304. [Google Scholar] [CrossRef]
- Balboa, G.R.; Ciampitti, I.A. Estimating biological nitrogen fixation in field-grown soybeans: Impact of B value. Plant Soil 2020, 446, 195–210. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean ± Standard Error for the Main Effects | p-Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Legume Species | Fertilization | Legume Species | Fertilization | Legume × Fertilization | ||||||
| Groundnut | Crotalaria | Velvet Bean | Cowpea | F1 | F2 | |||||
| Shoot DW (g m−2) | 185 ± 65 a | 181 ± 52 a | 87 ± 36 b | 109 ± 19 b | 143 ± 68 A | 138 ± 57 A | <0.001 | 0.79 | 0.50 | |
| Root DW (g m−2) | 54.3 ± 10.5 a | 34.0 ± 8.3 b | 6.5 ± 2.8 d | 19.7 ± 6.0 c | 30.6 ± 21.2 A | 26.6 ± 17.7 A | <0.001 | 0.17 | 0.51 | |
| Shoot: Root | 3.34 ± 0.56 c | 5.30 ± 0.70 b | 14.38 ± 3.99 a | 5.82 ± 1.45 b | 6.46 ± 3.75 A | 7.96 ± 5.65 A | <0.001 | 0.08 | 0.42 | |
| % N Shoot (%) | Total | 2.55 ± 0.18 | 3.07 ± 0.36 | 3.06 ± 0.44 | 2.83 ± 0.31 | 2.84 ± 0.41 | 2.92 ± 0.36 | 0.007 | 0.44 | 0.04 |
| F1 | 2.52 ± 0.26 a | 2.78 ± 0.18 a | 3.24 ± 0.46 a | 2.80 ± 0.44 a | - | - | 0.09 | - | - | |
| F2 | 2.59 ± 0.10 b | 3.36 ± 0.22 a | 2.89 ± 0.39 ab | 2.85 ± 0.17 ab | - | - | 0.006 | - | - | |
| ‰ P Shoot (‰) | 1.54 ± 0.22 a | 1.85 ± 0.41 a | 1.83 ± 0.28 a | 1.53 ± 0.31 a | 1.55 ± 0.32 B | 1.83 ± 0.30 A | 0.06 | 0.01 | 0.98 | |
| Shoot N:P | 16.8 ± 2.2 a | 17.1 ± 3.0 a | 17.1 ± 3.8 a | 19.0 ± 3.9 a | 18.8 ± 3.5 A | 16.2 ± 2.4 B | 0.48 | 0.03 | 0.46 | |
| δ15N (‰) | 2.02 ± 0.44 ab | 3.14 ± 1.29 a | 0.8 ± 0.72 b | 1.82 ± 0.83 b | 2.15 ± 1.10 A | 2.36 ± 1.27 A | <0.001 | 0.22 | 0.94 | |
| %Ndfa (%) | 58.6 ± 5.1 ab | 49.7 ± 10.6 b | 66.2 ± 8.0 a | 59.9 ± 9.0 ab | 56.7 ± 11.4 A | 60.5 ± 8.4 A | 0.005 | 0.20 | 0.32 | |
| Fixed N (g m−2) | 2.53 ± 0.98 a | 2.72 ± 0.95 a | 1.94 ± 0.56 a | 1.83 ± 0.38 a | 2.15 ± 0.93 A | 2.36 ± 0.71 A | 0.05 | 0.42 | 0.07 | |
| Nodule numb (m−2) * | 1158 ± 767 a | 118 ± 109 b | 82 ± 39 b | 61 ± 64 b | 456 ± 782 A | 254 ± 329 A | <0.001 | 0.15 | 0.62 | |
| Nodule DW (mg m−2) * | 5.83 ± 2.76 a | 0.55 ± 0.52 b | 2.94 ± 1.37 a | 0.43 ± 0.37 b | 2.98 ± 3.23 A | 1.90 ± 1.97 A | <0.001 | 0.18 | 0.57 | |
| F. Myc (%) | 8.8 ± 17.0 a | 6.7 ± 9.9 a | 11.3 ± 8.9 a | 5.4 ± 14.0 a | 7.71 ± 9.2 A | 8.33 ± 15.3 A | 0.40 | 0.79 | 0.10 | |
| M. Myc (%) | 0.56 ± 1.51 | 1.07 ± 1.98 | 1.13 ± 2.0 | 1.01 ± 2.84 | 1.06 ± 1.94 | 0.82 ± 2.20 | ||||
| Nodule Nb | Nodule DW | F. Myc | Shoot DM | Root DM | Shoot:Root | % N Shoot | ‰ P Shoot | Shoot N:P | %Ndfa | |
|---|---|---|---|---|---|---|---|---|---|---|
| Nodule DW | 0.86 *** | |||||||||
| F. Myc | –0.14 | –0.04 | ||||||||
| Shoot DW | 0.49 * | 0.30 | –0.11 | |||||||
| Root DW | 0.74 *** | 0.50 * | –0.15 | 0.82 *** | ||||||
| Shoot:Root | –0.38 | –0.11 | 0.17 | –0.49 * | –0.74 *** | |||||
| % N Shoot | –0.42. | –0.33 | –0.11 | –0.27 | –0.45 * | 0.22 | ||||
| ‰ P Shoot | –0.30 | –0.15 | 0.17 | –0.09 | –0.28 | 0.26 | 0.49 * | |||
| Shoot N:P | 0.00 | –0.10 | –0.27 | –0.09 | –0.04 | –0.11 | 0.21 | –0.74 *** | ||
| %Ndfa | –0.01 | 0.12 | –0.24 | –0.36 | –0.32 | 0.42 | 0.00 | –0.17 | 0.21 | |
| Fixed N | 0.34 | 0.22 | –0.29 | 0.73 *** | 0.50 * | –0.16 | 0.04 | 0.08 | –0.03 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razafintsalama, H.; Sauvadet, M.; Trap, J.; Autfray, P.; Ripoche, A.; Becquer, T. Legume Nitrogen Fixation and Symbioses in Low-Inputs Rainfed Rice Rotations. Sustainability 2021, 13, 12349. https://doi.org/10.3390/su132212349
Razafintsalama H, Sauvadet M, Trap J, Autfray P, Ripoche A, Becquer T. Legume Nitrogen Fixation and Symbioses in Low-Inputs Rainfed Rice Rotations. Sustainability. 2021; 13(22):12349. https://doi.org/10.3390/su132212349
Chicago/Turabian StyleRazafintsalama, Harimenja, Marie Sauvadet, Jean Trap, Patrice Autfray, Aude Ripoche, and Thierry Becquer. 2021. "Legume Nitrogen Fixation and Symbioses in Low-Inputs Rainfed Rice Rotations" Sustainability 13, no. 22: 12349. https://doi.org/10.3390/su132212349