
Citron Watermelon Potential to Improve Crop Diversification and Reduce Negative Impacts of Climate Change




Abstract
:1. Introduction
2. Materials and Methods
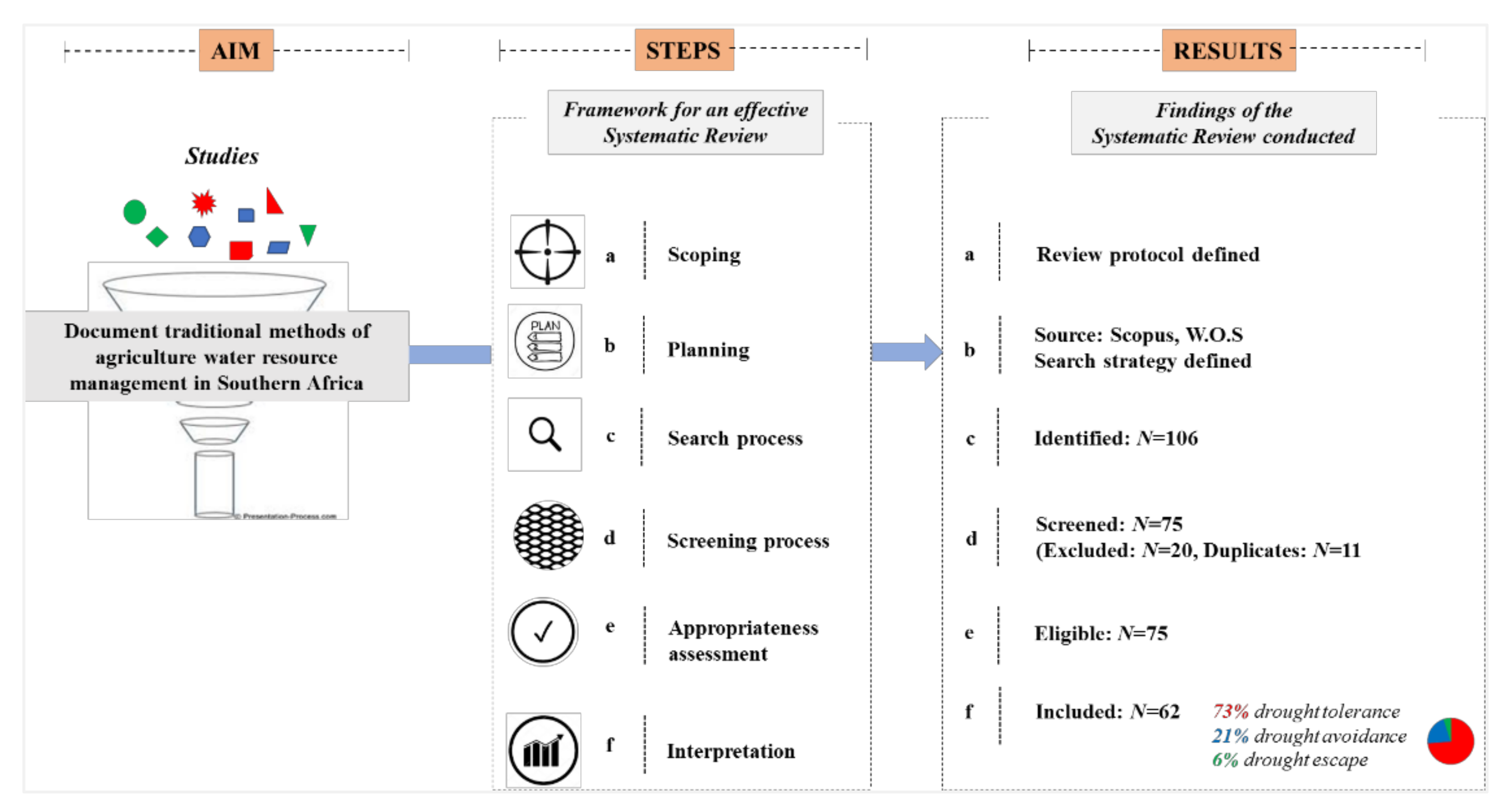
- Scoping
- Planning
- Identification/search
- Screening
- Eligibility/assessment and
- Presentation/interpretation
2.1. Research Question
2.2. Data Sources and Search Strategy
- Scopus (www.scopus.com) (accessed on 8 February 2020)
- Web of Science (www.webofknowledge.com) (accessed on 8 February 2020)
- Science Direct (www.sciencedirect.com) (accessed on 13 February 2020)
- Science.gov (www.science.gov) (accessed on 23 February 2020) and
- Google Scholar (scholar.google.com) (accessed on 10 March 2020)
2.3. Citation Management
2.4. Relevance Screening and Eligibility Criteria
2.5. Data Charting
2.6. Summarizing and Reporting
3. Results
4. Discussion
4.1. Citron Watermelon Contribution to Human Nutrition and Health
4.2. The Preservation of Citron Watermelon Biodiversity
5. Future Research Perspectives
5.1. Synchronization of Research and Methods
5.2. Integrating Phenomics to Harness the Potential of Developed Genomic Resources
5.3. Exploring Alternative Uses
5.4. Remote Sensing under Current and Future Climates
5.5. Exploration of Genetic Diversity and Local Adaptation
5.6. Systematic Germplasm Banking and Development of Genetic Resources
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chomicki, G.; Schaefer, H.; Renner, S.S. Origin and domestication of Cucurbitaceae crops: Insights from phylogenies, genomics and archaeology. N. Phytol. 2020, 226, 1240–1255. [Google Scholar] [CrossRef] [Green Version]
- Akashi, K.; Miyake, C.; Yokota, A. Citrulline, a novel compatible solute in drought-tolerant wild watermelon leaves, is an efficient hydroxyl radical scavenger. FEBS Lett. 2001, 508, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Levi, A.; Jarret, R.; Kousik, S.; Patrick Wechter, W.; Nimmakayala, P.; Reddy, U.K. Genetic Resources of Watermelon. In Genetics and Genomics of Cucurbitaceae; Springer: Berlin/Heidelberg, Germany, 2017; pp. 87–110. [Google Scholar] [CrossRef]
- Ngwepe, R.M.; Mashilo, J.; Shimelis, H. Progress in genetic improvement of citron watermelon (Citrullus lanatus var. citroides): A review. Genet. Resour. Crop Evol. 2019, 66, 735–758. [Google Scholar] [CrossRef]
- Janick, J.; Paris, H.S.; Parrish, D.C. The cucurbits of mediterranean antiquity: Identification of taxa from ancient images and descriptions. Ann. Bot. Lond. 2007, 100, 1441–1457. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Crop Production Data. Available online: http://www.fao.org/faostat/en/#data (accessed on 19 November 2020).
- Pasandide, B.; Khodaiyan, F.; Mousavi, Z.; Hosseini, S.S. Pectin extraction from citron peel: Optimization by Box-Behnken response surface design. Food Sci. Biotechnol. 2018, 27, 997–1005. [Google Scholar] [CrossRef]
- Alam, K.; Rahman, M.H. Women in natural disasters: A case study from southern coastal region of Bangladesh. Int. J. Disaster Risk Reduct. 2014, 8, 68–82. [Google Scholar] [CrossRef]
- Gusmini, G.; Wehner, T.C.; Jarret, R.L. Inheritance of egusi seed type in watermelon. J. Hered. 2004, 95, 268–270. [Google Scholar] [CrossRef] [Green Version]
- Chivenge, P.; Mabhaudhi, T.; Modi, A.T.; Mafongoya, P. The Potential Role of Neglected and Underutilised Crop Species as Future Crops under Water Scarce Conditions in Sub-Saharan Africa. Int. J. Env. Res. Public Health 2015, 12, 5685–5711. [Google Scholar] [CrossRef] [Green Version]
- Gil, J.D.B.; Reidsma, P.; Giller, K.; Todman, L.; Whitmore, A.; van Ittersum, M. Sustainable development goal 2: Improved targets and indicators for agriculture and food security. AMBIO 2019, 48, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Mabhaudhi, T.; Chimonyo, V.G.P.; Hlahla, S.; Massawe, F.; Mayes, S.; Nhamo, L.; Modi, A.T. Prospects of orphan crops in climate change. Planta 2019, 250, 695–708. [Google Scholar] [CrossRef] [Green Version]
- Shepard, D. Africa to push development agenda at upcoming climate summit. Afr. Renew. 2014, 28, 26–29. [Google Scholar] [CrossRef]
- Koutsos, T.M.; Menexes, G.C.; Dordas, C.A. An efficient framework for conducting systematic literature reviews in agricultural sciences. Sci. Total Env. 2019, 682, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Saha, I.; Sarkar, J.P.; Maulik, U. Integrated Rough Fuzzy Clustering for Categorical data Analysis. Fuzzy Sets Syst. 2019, 361, 1–32. [Google Scholar] [CrossRef]
- Akashi, K.; Nishimura, N.; Ishida, Y.; Yokota, A. Potent hydroxyl radical-scavenging activity of drought-induced type-2 metallothionein in wild watermelon. Biochem. Biophys. Res. Commun. 2004, 323, 72–78. [Google Scholar] [CrossRef]
- Baloglu, M.C.; Ulu, F.; Altunoglu, Y.C.; Pekol, S.; Alagoz, G.; Ese, O. Identification, molecular characterization and expression analysis of RPL24 genes in three Cucurbitaceae family members: Cucumber, melon and watermelon. Biotechnol. Biotechnol. Equip. 2015, 29, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Celik Altunoglu, Y.; Baloglu, M.C.; Baloglu, P.; Yer, E.N.; Kara, S. Genome-wide identification and comparative expression analysis of LEA genes in watermelon and melon genomes. Physiol. Mol. Biol. Plants 2017, 23, 5–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.P.; Suh, H.Y.; Kim, J.H.; Ahn, Y.K.; Kim, J.; Lee, E.S.; Kim, D.S.; Lee, H.E. Expression Analysis of Five Arabidopsis PDLP5 Homologous in Watermelon Subjected to Biotic and Abiotic Stresses. Hortic. Environ. Biotechnol. 2017, 58, 367–375. [Google Scholar] [CrossRef]
- Kim, S.J.; Jung, I.L.; Lee, H.E.; Lee, J.H. Abiotic stress and tissue-specific reference genes for quantitative reverse transcription PCR analysis in Korean native watermelons, Citrullus lanatus ‘Black-King’ and ‘Speed-Plus-Honey’. Appl. Biol. Chem. 2018, 61, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, P.; Sun, L.; Wang, Y.; Ji, K.; Sun, Y.; Dai, S.; Chen, P.; Duan, C.; Leng, P. Expression analysis of beta-glucosidase genes that regulate abscisic acid homeostasis during watermelon (Citrullus lanatus) development and under stress conditions. J. Plant Physiol. 2012, 169, 78–85. [Google Scholar] [CrossRef]
- Ram, C.; Verma, M.; Panwar, B.S.; Arya, L. Molecular cloning and in silico analysis of DREB-like gene in watermelon. Indian J. Hortic. 2015, 72, 338–342. [Google Scholar] [CrossRef]
- Si, Y.; Dane, F.; Rashotte, A.; Kang, K.; Singh, N.K. Cloning and expression analysis of the Ccrboh gene encoding respiratory burst oxidase in Citrullus colocynthis and grafting onto Citrullus lanatus (watermelon). J. Exp. Bot. 2010, 61, 1635–1642. [Google Scholar] [CrossRef] [Green Version]
- Unel, N.M.; Cetin, F.; Karaca, Y.; Altunoglu, Y.C.; Baloglu, M.C. Comparative identification, characterization, and expression analysis of bZIP gene family members in watermelon and melon genomes. Plant Growth Regul. 2019, 87, 227–243. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, R.; Yang, X.; Zhu, C.; Li, H.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Comparative Analysis of Calcium-Dependent Protein Kinase in Cucurbitaceae and Expression Studies in Watermelon. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Dong, W.; Zhan, Y.; Huang, Z.A.; Li, Z.; Kim, I.S.; Zhang, C. Genome-wide identification and expression analysis of ClLAX, ClPIN and ClABCB genes families in Citrullus lanatus under various abiotic stresses and grafting. BMC Genet. 2017, 18, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahara, K.; Akashi, K.; Yokota, A. Biochemical and molecular characterization of glutamate N-acetyltransferase involved in citrulline accumulation in wild watermelon during drought/strong-light stresses. Plant Cell Physiol. 2005, 46, S74. [Google Scholar]
- Takahara, K.; Yoshino, A.; Akashi, K.; Yokota, A. Regulation of metabolic pathways for the massive accumulation of citrulline during drought/strong light stress in wild watermelon. Plant Cell Physiol. 2006, 47, S102. [Google Scholar]
- Xu, Z.; Sun, M.; Jiang, X.; Sun, H.; Dang, X.; Cong, H.; Qiao, F. Glycinebetaine Biosynthesis in Response to Osmotic Stress Depends on Jasmonate Signaling in Watermelon Suspension Cells. Front. Plant Sci. 2018, 9, 1469. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhu, J.H.; Yang, Y.X. Genome-Wide Identification and Expression Analysis of NF-Y Transcription Factor Families in Watermelon (Citrullus lanatus). J. Plant Growth Regul. 2017, 36, 590–607. [Google Scholar] [CrossRef]
- Yokota, A.; Kawasaki, S.; Iwano, M.; Nakamura, C.; Miyake, C.; Akashi, K. Citrulline and DRIP-1 protein (ArgE homologue) in drought tolerance of wild watermelon. Ann. Bot. Lond. 2002, 89, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Masuda, A.; Kuwano, M.; Yokota, A.; Akashi, K. Programmed proteome response for drought avoidance/tolerance in the root of a C(3) xerophyte (wild watermelon) under water deficits. Plant Cell Physiol. 2008, 49, 226–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.S.; Yang, Y.G.; Su, Y.Q.; Li, X.F. Antioxidant enzymes activities in leaves and yield analysis of different ecological types watermelon under drought stress. Agric. Res. Arid Areas 2017, 35, 138–143. [Google Scholar] [CrossRef]
- Akashi, K.; Yoshimura, K.; Kajikawa, M.; Hanada, K.; Kosaka, R.; Kato, A.; Katoh, A.; Nanasato, Y.; Tsujimoto, H.; Yokota, A. Potential involvement of drought-induced Ran GTPase CLRan1 in root growth enhancement in a xerophyte wild watermelon. Biosci. Biotechnol. Biochem. 2016, 80, 1907–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alessandro, S.; Mizokami, Y.; Legeret, B.; Havaux, M. The Apocarotenoid beta-Cyclocitric Acid Elicits Drought Tolerance in Plants. iScience 2019, 19, 461–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakki, E.E.; Hamurcu, M.; Gezgin, S.; Demiral, T. Physiological responses of two contrasting watermelon genotypes exposed to drought and nitric oxide. J. Biotechnol. 2016, 231, S25. [Google Scholar] [CrossRef]
- Kohzuma, K.; Akashi, K.; Munekage, Y.; Hisabori, T.; Yokota, A. Preferential decay of the CF1 epsilon subunit induces thylakoid uncoupling in wild watermelon under drought stress. Photosynth. Res. 2007, 91, 191. [Google Scholar] [CrossRef]
- Kajikawa, M.; Morikawa, K.; Abe, Y.; Yokota, A.; Akashi, K. Establishment of a transgenic hairy root system in wild and domesticated watermelon (Citrullus lanatus) for studying root vigor under drought. Plant Cell Rep. 2010, 29, 771–778. [Google Scholar] [CrossRef]
- Liu, A.M.; Latimer, J.G. Water Relations and Abscisic-Acid Levels of Watermelon as Affected by Rooting Volume Restriction. J. Exp. Bot. 1995, 46, 1011–1015. [Google Scholar] [CrossRef]
- Omirou, M.; Ioannides, I.M.; Ehaliotis, C. Mycorrhizal inoculation affects arbuscular mycorrhizal diversity in watermelon roots, but leads to improved colonization and plant response under water stress only. Appl. Soil Ecol. 2013, 63, 112–119. [Google Scholar] [CrossRef]
- Park, M.; Han, J.; Ahn, Y.; Kim, J.; Lee, H.; Jang, Y.; Gaxiola, R.; Hirschi, K.D.; Park, S. Ectopic expression of Arabidopsis H+-pyrophosphatase AVP1 enhances drought resistance in bottle gourd (Lagenaria siceraria Standl.). Plant Cell Tissue Organ Cult. 2014, 118, 383–389. [Google Scholar] [CrossRef]
- Botha, F.C.; Small, J.G. Effect of Water Stress on the Carbohydrate Metabolism of Citrullus lanatus Seeds during Germination. Plant Physiol. 1985, 77, 79–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akashi, K.; Hashizume, T.; Yamashita, A.; Toh, H.; Hattori, M.; Yokota, A. Expressed sequence tag-based gene expression analysis under drought stress in wild watermelon. Plant Cell Physiol. 2004, 45, S94. [Google Scholar]
- Akashi, K.; Ishida, Y.; Nishimura, N.; Yokota, A. Analysis of drought-induced metallothionein in wild watermelon. Plant Cell Physiol. 2003, 44, S1. [Google Scholar]
- Akashi, K.; Morikawa, K.; Yokota, A. Agrobacterium-mediated transformation system for the drought and excess light stress-tolerant wild watermelon (Citrullus lanatus). Plant Biotechnol. 2005, 22, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Akashi, K.; Takahara, K.; Yamada, R.; Miyake, C.; Yokota, A. Functional analysis of DRIP-1, a drought-induced polypeptide in wild watermelon. Plant Cell Physiol. 2002, 43, S112. [Google Scholar]
- Akashi, K.; Yoshida, K.; Kuwano, M.; Kajikawa, M.; Yoshimura, K.; Hoshiyasu, S.; Inagaki, N.; Yokota, A. Dynamic changes in the leaf proteome of a C3 xerophyte, Citrullus lanatus (wild watermelon), in response to water deficit. Planta 2011, 233, 947–960. [Google Scholar] [CrossRef]
- Akashi, K.; Yokota, A. Molecular responses of wild watermelon to drought stress. Plant Cell Physiol. 2003, 44, S111. [Google Scholar]
- Ansari, W.A.; Atri, N.; Ahmad, J.; Qureshi, M.I.; Singh, B.; Kumar, R.; Rai, V.; Pandey, S. Drought mediated physiological and molecular changes in muskmelon (Cucumis melo L.). PLoS ONE 2019, 14, e0222647. [Google Scholar] [CrossRef]
- Baragé, M.; Balla, A.; Adam, T. Identifying sources of water stress tolerance from wild species of the family Cucurbitaceae in vitro culture. Biotechnol. Agron. Soc. Environ. 2006, 10, 3–6. [Google Scholar]
- Cheng, W.S.; Zeng, D.L.; Zhang, N.; Zeng, H.X.; Shi, X.F.; Li, Y.H.; Sun, Y.H. Foliar Application of Abscisic Acid and Sulfonamide Compounds Induced Drought Tolerance in Watermelon. Appl. Mech. Mater. 2015, 723, 705–710. [Google Scholar] [CrossRef]
- Ekbic, E.; Cagiran, C.; Korkmaz, K.; Kose, M.A.; Aras, V. Assessment of watermelon accessions for salt tolerance using stress tolerance indices. Cienc. E Agrotecnologia 2017, 41, 616–625. [Google Scholar] [CrossRef] [Green Version]
- Hamurcu, M.; Hakki, E.E.; Demiral, T.; Gezgin, S. Biochemical effects of drought stress on two Turkish watermelon varieties are different and influenced by nitric oxide. J. Biotechnol. 2016, 231, S26. [Google Scholar] [CrossRef]
- Kim, S.J.; Huh, Y.C.; Ahn, Y.K.; Kim, J.; Kim, D.S.; Lee, H.E. Watermelon (Citrullus lanatus) late-embryogenesis abundant group 3 protein, ClLEA3-1, responds to diverse abiotic stresses. Hortic. Environ. Biotechnol. 2015, 56, 555–560. [Google Scholar] [CrossRef]
- Malambane, G.; Nonaka, S.; Shiba, H.; Ezura, H.; Tsujimoto, H.; Akashi, K. Comparative effects of ethylene inhibitors on Agrobacterium-mediated transformation of drought-tolerant wild watermelon. Biosci. Biotechnol. Biochem. 2018, 82, 433–441. [Google Scholar] [CrossRef]
- Masuda, A.; Yoshimura, K.; Akashi, K.; Yokota, A. Proteomic analysis of avoidance and defense mechanisms to drought stress in the root of wild watermelon. Plant Cell Physiol. 2006, 47, S101. [Google Scholar]
- Nanasato, Y.; Akashi, K.; Yokota, A. Co-expression of cytochrome b561 and ascorbate oxidase in leaves of wild watermelon under drought and high light conditions. Plant Cell Physiol 2005, 46, 1515–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.A.; Chen, C.; Shireen, F.; Zheng, Z.; Sohail, H.; Afzal, M.; Ali, M.A.; Bie, Z.; Huang, Y. Genome-wide expression profiling of leaves and roots of watermelon in response to low nitrogen. BMC Genom. 2018, 19, 456. [Google Scholar] [CrossRef]
- Ramirez, A.H.M.; Jhala, A.J.; Singh, M. Factors Affecting Germination of Citron melon (Citrullus lanatus var. citroides). Weed Sci. 2014, 62, 45–50. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable Grafting: A Toolbox for Securing Yield Stability under Multiple Stress Conditions. Front. Plant Sci. 2017, 8, 2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; Zhang, C.; Meng, S.; Dane, F. Gene expression changes in response to drought stress in Citrullus colocynthis. Plant Cell Rep. 2009, 28, 997–1009. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Yang, Y.; Wang, Y.; Mo, Y.; Zhang, R.; Zhang, Y.; Ma, J.; Wei, C.; Zhang, X. Identification and expression analyses of WRKY genes reveal their involvement in growth and abiotic stress response in watermelon (Citrullus lanatus). PLoS ONE 2018, 13, e0191308. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Akashi, K.; Yokota, A. Proteomic analysis of drought/strong light stress responses in wild watermelon leaves. Plant Cell Physiol. 2005, 46, S72. [Google Scholar]
- Yoshimura, K.; Masuda, A.; Akashi, K.; Yokota, A. Identification of drought-responsible proteins in the root of wild watermelon by proteomic analysis. Plant Cell Physiol. 2005, 46, S74. [Google Scholar]
- Yoshimura, K.; Masuda, A.; Akashi, K.; Yokota, A. Regulation of the root development mechanism involved in Ran GTPase of wild watermelon under drought stress. Plant Cell Physiol. 2006, 47, S101. [Google Scholar]
- Zhou, Y.; Li, J.W.; Wang, J.H.; Yang, W.T.; Yang, Y.X. Identification and Characterization of the Glutathione Peroxidase (GPX) Gene Family in Watermelon and Its Expression under Various Abiotic Stresses. Agronomy 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.J. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot. 2010, 61, 3509–3517. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Mitra, J. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci. 2001, 80, 758–763. [Google Scholar]
- Tardieu, F. Plant response to environmental conditions: Assessing potential production, water demand, and negative effects of water deficit. Front. Physiol. 2013, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Du, H.; Ning, J.; Ye, H.; Xiong, L. Characterization of Glossy1-homologous genes in rice involved in leaf wax accumulation and drought resistance. Plant Mol. Biol. 2009, 70, 443–456. [Google Scholar] [CrossRef]
- Franks, S.J. Plasticity and evolution in drought avoidance and escape in the annual plant Brassica rapa. N. Phytol. 2011, 190, 249–257. [Google Scholar] [CrossRef]
- Sherrard, M.E.; Maherali, H. The adaptive significance of drought escape in Avena barbata, an annual grass. Evolution 2006, 60, 2478–2489. [Google Scholar] [CrossRef] [PubMed]
- Akashi, K.; Yoshimura, K.; Nanasato, Y.; Takahara, K.; Munekage, Y.; Yokota, A. Wild plant resources for studying molecular mechanisms of drought/strong light stress tolerance. Plant Biotechnol. 2008, 25, 257–263. [Google Scholar] [CrossRef]
- Hsiao, T.C.; Acevedo, E. Plant Responses to Water Deficits, Water-Use Efficiency, and Drought Resistance. In Developments in Agricultural and Managed Forest Ecology; Elsevier: Amsterdam, The Netherlands, 1975; Volume 1, pp. 59–84. [Google Scholar]
- Gorai, M.; Laajili, W.; Santiago, L.S.; Neffati, M. Rapid recovery of photosynthesis and water relations following soil drying and re-watering is related to the adaptation of desert shrub Ephedra alata subsp alenda (Ephedraceae) to arid environments. Environ. Exp. Bot. 2015, 109, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.J.; Yang, J.J.; Wei, J.Y.; Li, Y.F.; Xu, J.; Jiang, Y.G. Antioxidant activities of peel, pulp and seed fractions of common fruits as determined by FRAP assay. Nutr. Res. 2003, 23, 1719–1726. [Google Scholar] [CrossRef]
- Paliyath, G.; Bakovic, M.; Shetty, K. Functional Foods, Nutraceuticals, and Degenerative Disease Prevention; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Zhengkang, H.; Wang, G.; Yao, W.; Zhu, W.-Y. Isoflavonic phytoestrogens-new prebiotics for farm animals: A review on research in China. Curr. Issues Intest. Microbiol. 2006, 7, 53–60. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, D.I.; Myer, L.; Klemick, H. A Training Guide for In Situ Conservation on-Farm; Bioversity International, Stanford University Press: Redwood City CA, USA, 2000. [Google Scholar]
- Swislocki, M. Culinary Nostalgia: Regional Food Culture and the Urban Experience in Shanghai; Stanford University Press: Redwood City, CA, USA, 2008. [Google Scholar]
- Mujaju, C.; Zborowska, A.; Werlemark, G.; Garkava-Gustavsson, L.; Andersen, S.B.; Nybom, H. Genetic diversity among and within watermelon (Citrullus lanatus) landraces in Southern Africa. J. Hortic. Sci. Biotech. 2011, 86, 353–358. [Google Scholar] [CrossRef]
- Zadawa, A.N.; Omran, A. Rural Development in Africa: Challenges and Opportunities. In Sustaining our Environment for Better Future; Springer: Berlin/Heidelberg, Germany, 2020; pp. 33–42. [Google Scholar]
- Henriksson Malinga, R.; Jewitt, G.P.W.; Lindborg, R.; Andersson, E.; Gordon, L.J. On the other side of the ditch: Exploring contrasting ecosystem service coproduction between smallholder and commercial agriculture. Ecol. Soc. 2018, 23. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, S.; Goldman, I.; Ortiz, R. Pursuing the Potential of Heirloom Cultivars to Improve Adaptation, Nutritional, and Culinary Features of Food Crops. Agronomy 2019, 9, 441. [Google Scholar] [CrossRef] [Green Version]
- Paris, H.S. Overview of the origins and history of the five major cucurbit crops: Issues for ancient DNA analysis of archaeological specimens. Veg. Hist. Archaeobotany 2016, 25, 405–414. [Google Scholar] [CrossRef]
- Mcgregor, C. Citrullus lanatus germplasm of southern Africa. Isr. J. Plant Sci. 2012, 60, 403–413. [Google Scholar] [CrossRef]
- GBIF. Citrullus lanatus var. citroides (L.H.Bailey) Mansf. In Kulturpflanze, Beih. 2:422. 1959; Wiersema, J.H., Ed.; GBIF: Copenhagen, Denmark, 2019. [Google Scholar] [CrossRef]
- GBIF. Free and Open Access to Biodiversity Data. Available online: https://www.gbif.org/ (accessed on 8 February 2019).
- Sultana, B.; Ashraf, R. Watermelon (Citrullus lanatus) Oil. In Fruit Oils: Chemistry and Functionality; Springer: Berlin/Heidelberg, Germany, 2019; pp. 741–756. [Google Scholar]
- Mashilo, J.; Shimelis, H.; Odindo, A.O.; Amelework, B. Genetic diversity and differentiation in citron watermelon [Citrullus lanatus var. citroides] landraces assessed by simple sequence repeat markers. Sci. Hortic. 2017, 214, 99–106. [Google Scholar] [CrossRef]
- CFF. BamNetWork. Available online: http://bambaragroundnut.org/ (accessed on 15 June 2020).
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- APPF. The Australian Plant Phenomics Facility. Available online: https://www.plantphenomics.org.au/ (accessed on 19 August 2020).
- Arend, D.; Junker, A.; Scholz, U.; Schuler, D.; Wylie, J.; Lange, M. PGP repository: A plant phenomics and genomics data publication infrastructure. Database Oxf. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahla, H.R.; Singh, J.P.; Roy, M.M. Seed Purpose Watermelon in Arid Zone. Available online: http://www.cazri.res.in/publications/HRMahla.pdf (accessed on 6 January 2020).
- Alka, G.; Anamika, S.; Ranu, P. A review on watermelon (Citrullus lanatus) medicinal seeds. J. Pharmacogn. Phytochem. 2018, 7, 2222–2225. [Google Scholar]
- Zabel, F.; Putzenlechner, B.; Mauser, W. Global agricultural land resources--a high resolution suitability evaluation and its perspectives until 2100 under climate change conditions. PLoS ONE 2014, 9, e107522. [Google Scholar] [CrossRef] [Green Version]
- Alcantara, C.; Kuemmerle, T.; Baumann, M.; Bragina, E.V.; Griffiths, P.; Hostert, P.; Knorn, J.; Muller, D.; Prishchepov, A.V.; Schierhorn, F.; et al. Mapping the extent of abandoned farmland in Central and Eastern Europe using MODIS time series satellite data. Environ. Res. Lett. 2013, 8, 035035. [Google Scholar] [CrossRef]
- Ujoh, F.; Igbawua, T.; Paul, M.O. Suitability mapping for rice cultivation in Benue State, Nigeria using satellite data. Geo Spat. Inf. Sci. 2019, 22, 332–344. [Google Scholar] [CrossRef] [Green Version]
- CGIAR_Big_Data. Seed systems in Nepal are going digital. Available online: https://bigdata.cgiar.org/rss-article/seed-systems-in-nepal-are-going-digital/ (accessed on 6 January 2020).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database (s) | Primary Term (s) | Expanded Term (s) |
|---|---|---|
| Scopus Web of Science Science Direct Science.gov Google Scholar | Drought stress | “water stress” OR “moisture stress” OR “water deficit” OR “water shortage” OR “water scarcity” AND |
| C. lanatus spp. | “watermelon” OR “citron watermelon” OR “desert watermelon” OR “wild watermelon” OR “melon” OR “muskmelon” AND | |
| Mechanism(s) | “adaptation strategy” OR “avoidance” OR “escape” OR “tolerance” |
| Mechanisms | |||||
|---|---|---|---|---|---|
| Author | Title | Research summary | DA | DT | DE |
| [44] | Expressed sequence tag-based gene expression analysis under drought stress in wild watermelon | Changes in gene expression in roots within 6 h of water stress. Genes involved in oxidative stress (glutathione peroxidase, glucose-6-phosphate-dehydrogenase, and ascorbate peroxidase) were demonstrated to be regulated by water stress. | ✓ | ||
| [45] | Analysis of drought-induced metallothionein in wild watermelon | Gene (CLMT2) of the same homology with type-2 metallothionein contributed to the survival of wild watermelon under severe drought. | ✓ | ||
| [46] | Agrobacterium-mediated transformation system for the drought and excess light stress-tolerant wild watermelon (Citrullus lanatus) | Experimental basis for molecular studies of wild watermelon genes helps understand their contribution to stress tolerance in this plant. | ✓ | ||
| [16] | Potent hydroxyl radical-scavenging activity of drought-induced type-2 metallothionein in wild watermelon | Gene (CLMT2) of the same homology with type-2 metallothionein contributed to the survival of wild watermelon under severe drought. | ✓ | ||
| [47] | Functional analysis of DRIP-1, a drought-induced polypeptide in wild watermelon | Wild watermelon accumulates high concentrations of citrulline, glutamate, and arginine in its leaves during drought. | ✓ | ||
| [48] | Dynamic changes in the leaf proteome of a C3 xerophyte, Citrullus lanatus (wild watermelon), in response to water deficit | Defense response of wild watermelon involves orchestrated regulation of functional proteins, of which HSPs play a pivotal role in the protection of the plant under water deficit. | ✓ | ||
| [49] | Molecular responses of wild watermelon to drought stress | Rapid accumulation of HSPs in stressed melons. | ✓ | ||
| [35] | Potential involvement of drought-induced Ran GTPase CLRan1 in root growth enhancement in a xerophyte wild watermelon | Ran GTPase (CLRan1), expressed in the roots of drought-resistant wild watermelon, functions as a positive factor for maintaining root growth under osmotic stress. | ✓ | ✓ | |
| [50] | Drought mediated physiological and molecular changes in muskmelon (Cucumis melo L.) | Increased activity of catalase (CAT), superoxide dismutase (SOD), ascorbate peroxidase (APX), and guaiacol (POD). Under drought stress, muskmelon elevates the abundance of defense proteins and suppresses catabolic proteins | ✓ | ✓ | |
| [17] | Identification, molecular characterization, and expression analysis of RPL24 genes in three Cucurbitaceae family members: cucumber, melon, and watermelon | Ribosomal protein L24 (RPL24) is responsible for stabilizing the peptidyl transferase activity—increased expression of CmRPL24-01 genes in melon leaf tissue at 3 h upon polyethylene glycol treatment. | ✓ | ||
| [51] | Identifying sources of water stress tolerance from wild species of the family Cucurbitaceae in vitro culture | Evaluating water stress tolerance capacity using the calus recuperation after dehydration under a laminar flux hot until a loss of 50% of their fresh weight as a basis. | ✓ | ||
| [18] | Genome-wide identification and comparative expression analysis of LEA genes in watermelon and melon genomes | Induction of LEA genes in root and leaf tissues after drought application. | ✓ | ||
| [52] | Foliar application of abscisic acid and sulfonamide compounds induced drought tolerance in watermelon | Sulfacetamide and sulfasalazine improve drought resistance similar to ABA by increasing proline, glycine betaine and malondialdehyde, and ascorbate peroxidase activity. | ✓ | ||
| [36] | The apocarotenoid beta-cyclocitric acid elicits drought tolerance in plants | Volatile compound β-cyclocitral (β-CC) in plant leaves, when converted to β-cyclocitric acid (β-CCA), signals drought tolerance. | ✓ | ✓ | |
| [53] | Assessment of watermelon accessions for salt tolerance using stress tolerance indices | GMP and STI indices indicated that G04, G14, and G21 could be prominent sources to develop drought tolerance. | ✓ | ||
| [43] | Effect of water stress on the carbohydrate metabolism of Citrullus lanatus seeds during germination | Stressed seeds lower the rate of 14CO2 release from [2-14C]acetate, [1-14C]glucose, and [6-14C]glucose. | ✓ | ✓ | |
| [37] | Physiological responses of two contrasting watermelon genotypes exposed to drought and nitric oxide | Drought stress decreased fresh and dry weights of shoots and roots and lengths drought-sensitive genotype KAR. 147 | ✓ | ||
| [54] | Biochemical effects of drought stress on two Turkish watermelon varieties are different and influenced by nitric oxide | MDA (a marker of oxidative damage on lipid membranes) was increased due to drought in watermelon genotypes and NO treatment slightly reduced MDA contents under drought stress. | ✓ | ||
| [19] | Expression analysis of five arabidopsis PDLP5 homologous in watermelon subjected to biotic and abiotic stresses | Plasmodesmata-located protein 5 (PDLP5) controls cell-to-cell communication and defense signaling. | ✓ | ||
| [39] | Establishment of a transgenic hairy root system in wild and domesticated watermelon (Citrullus lanatus) for studying root vigour under drought | A powerful tool for the comparative study of the molecular mechanism underlying drought-induced root growth in desert plants. | ✓ | ✓ | |
| [55] | Watermelon (Citrullus lanatus) late-embryogenesis abundant group 3 protein, ClLEA3-1, responds to diverse abiotic stresses | A representative group of LEA proteins ClLEA3-1 (Cl017745) can be used as an abiotic stress marker gene in watermelon. | ✓ | ||
| [20] | Abiotic stress and tissue-specific reference genes for quantitative reverse transcription PCR analysis in Korean native watermelons, Citrullus lanatus ‘Black-King’ and ‘Speed-Plus-Honey’ | Reference genes (ClACT and ClEF1α) were expressed in flowers, leaves, tendrils, stem, and roots after drought treatment. | ✓ | ||
| [38] | Preferential decay of the CF1 epsilon subunit induces thylakoid uncoupling in wild watermelon under drought stress | Selective decomposition of epsilon subunit induces uncoupling of thylakoid membranes under drought and contributes to the avoidance of over-acidification in the thylakoid lumen under excess light conditions. | ✓ | ||
| [21] | Expression analysis of beta-glucosidase genes that regulate abscisic acid homeostasis during watermelon (Citrullus lanatus) development and under stress conditions | Beta-glucosidase genes regulate ABA content during drought stress. | ✓ | ||
| [40] | Water relations and abscisic-acid levels of watermelon as affected by rooting volume restriction | ABA act as a signal for reduced growth of plants under rooting volume restriction (RVR) conditions. | ✓ | ✓ | |
| [56] | Comparative effects of ethylene inhibitors on Agrobacterium-mediated transformation of drought-tolerant wild watermelon | Controlling ethylene level during co-cultivation and shoot formation using the cad’s-harbouring Agrobacterium enhances drought tolerance. | ✓ | ||
| [57] | Proteomic analysis of avoidance and defence mechanisms to drought stress in the root of wild watermelon | Defense response of wild melons involves the orchestrated regulation of functional proteins. | ✓ | ✓ | |
| [58] | Co-expression of cytochrome b561 and ascorbate oxidase in leaves of wild watermelon under drought and high light conditions | Levels of cDNA (CLb561A) mRNA and protein were elevated in the leaves during drought. | ✓ | ||
| [59] | Genome-wide expression profiling of leaves and roots of watermelon in response to low nitrogen | Under abiotic stress, leaf tissues are more sensitive compared with root tissues. 9598 genes were differentially expressed, out of which 4533 genes were up-regulated, and 5065 genes were down-regulated. | ✓ | ||
| [41] | Mycorrhizal inoculation affects arbuscular mycorrhizal diversity in watermelon roots but leads to improved colonization and plant response under water stress | Plant inoculation with mycorrhizal fungi was related to the response of plants to water stress conditions by improving WUE. | ✓ | ||
| [42] | Ectopic expression of Arabidopsis H+-pyrophosphatase AVP1 enhances drought resistance in bottle gourd (Lagenaria siceraria Standl.) | Wild-type plants showed minimal growth while the AVP-1 expressing plants resumed rapid growth, displaying longer ramified primary roots. | ✓ | ||
| [22] | Molecular cloning and in silico analysis of DREB-like gene in watermelon | DREB genes were identified from watermelon related to drought-tolerant genes. | ✓ | ||
| [60] | Factors Affecting Germination of Citron melon (Citrullus lanatus var. citroides) | Citron melon can grow in a wide range of climatic conditions. | ✓ | ✓ | |
| [61] | Vegetable grafting: a toolbox for securing yield stability under multiple stress conditions | Movement of mRNA through the phloem from rootstock to scion regulate plant growth and adaptation to drought stress. | ✓ | ||
| [27] | Phytohormones regulate accumulation of osmolytes under abiotic stress | Osmolytes (proline, glycine-betaine, polyamines, and sugars) are accumulated to safeguard the cellular machinery. Phytohormones (abscisic acid, brassinosteroids, cytokinins, ethylene, jasmonates, and salicylic acid) modulates the accumulation of osmolytes. | ✓ | ||
| [23] | Cloning and expression analysis of the Ccrboh gene encoding respiratory burst oxidase in Citrullus colocynthis and grafting onto Citrullus lanatus (watermelon) | Drought-responsive gene Ccrboh is functionally important during the acclimation of plants to stress, and it is promising for improving the drought tolerance of other cucurbit species. | ✓ | ||
| [62] | Gene expression changes in response to drought stress in Citrullus colocynthis | During drought, stress-responsive genes and plant hormones are involved in an extensive cross-talk. | ✓ | ||
| [28] | Biochemical and molecular characterization of glutamate N-acetyltransferase involved in citrulline accumulation in wild watermelon during drought/strong-light stresses | Exogenous acetylene triggers the accumulation of citrulline to maintain the plant membrane structure. | ✓ | ||
| [29] | Regulation of metabolic pathways for the massive accumulation of citrulline during drought/strong light stress in wild watermelon | Rapid accumulation of citrulline in leaves and shoot under drought stress. | ✓ | ||
| [24] | Comparative identification, characterization, and expression analysis of bZIP gene family members in watermelon and melon genomes | Genes (ClabZIP and CmbZIP) were expressed in leaf and root tissues after the drought was imposed. | ✓ | ||
| [25] | Comparative analysis of calcium-dependent protein kinase in Cucurbitaceae and expression studies in watermelon | The study provides insights into the evolutionary history of gene families in Cucurbitaceae and indicates a subset of candidate genes for functional characterizations. | ✓ | ||
| [30] | Glycinebetaine biosynthesis in response to osmotic stress depends on jasmonate signalling in watermelon suspension cells | Osmotic stress-induced glycinebetaine biosynthesis occurs via JA signal transduction and contributes to osmotic stress hardening. | ✓ | ||
| [31] | Genome-wide identification and expression analysis of NF-Y transcription factor families in watermelon (Citrullus lanatus) | The study provides a foundation for further functional analysis of NF-Y proteins during watermelon development and responses to drought stress. The results will be valuable for evolutionary analysis of the NF-Y family in Cucurbitaceae species. | ✓ | ||
| [63] | Identification and expression analyses of WRKY genes reveal their involvement in growth and abiotic stress response in watermelon (Citrullus lanatus) | A total of 63 putative WRKY genes in watermelon were reported to regulate respective target genes. | ✓ | ||
| [32] | Citrulline and DRIP-1 protein (ArgE homologue) in drought tolerance of wild watermelon | Wild watermelon accumulates high concentrations of citrulline, glutamate, and arginine in its leaves during drought. | ✓ | ||
| [33] | Programmed proteome response for drought avoidance/tolerance in the root of a C3 xerophyte (wild watermelon) under water deficits | Defense response of wild watermelon involves orchestrated regulation of functional proteins, of which HSPs play a pivotal role in the protection of the plant under water deficit. | ✓ | ||
| [64] | Proteomic analysis of drought/strong light stress responses in wild watermelon leaves | DREB genes were identified from watermelon related to drought-tolerant genes. | ✓ | ||
| [65] | Identification of drought-responsible proteins in the root of wild watermelon by proteomic analysis | DREB genes were identified from watermelon related to drought-tolerant genes. | ✓ | ||
| [66] | Regulation of the root development mechanism involved in Ran GTPase of wild watermelon under drought stress | Ran GTPase genes function in watermelon development, as well as in response to abiotic stress and hormones. | ✓ | ||
| [26] | Genome-wide identification and expression analysis of ClLAX, ClPIN, and ClABCB genes families in Citrullus lanatus under various abiotic stresses and grafting | Expression genes (ClLAX, ClPIN, and ClABCB) under drought helps to understand the roles of auxin transporter genes in watermelon adaptation to environmental stresses. | ✓ | ||
| [34] | Antioxidant enzymes activities in leaves and yield analysis of different ecological types watermelon under drought stress | The content of proline and the activity of SOD, POD, and CAT were genotype related. | ✓ | ✓ | ✓ |
| [67] | Identification and characterization of the glutathione peroxidase (GPX) gene family in watermelon and its expression under various abiotic stresses | ClGPX genes function in watermelon development as well as in response to abiotic stress and hormones. | ✓ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandizvo, T.; Odindo, A.O.; Mashilo, J. Citron Watermelon Potential to Improve Crop Diversification and Reduce Negative Impacts of Climate Change. Sustainability 2021, 13, 2269. https://doi.org/10.3390/su13042269
Mandizvo T, Odindo AO, Mashilo J. Citron Watermelon Potential to Improve Crop Diversification and Reduce Negative Impacts of Climate Change. Sustainability. 2021; 13(4):2269. https://doi.org/10.3390/su13042269
Chicago/Turabian StyleMandizvo, Takudzwa, Alfred Oduor Odindo, and Jacob Mashilo. 2021. "Citron Watermelon Potential to Improve Crop Diversification and Reduce Negative Impacts of Climate Change" Sustainability 13, no. 4: 2269. https://doi.org/10.3390/su13042269
APA StyleMandizvo, T., Odindo, A. O., & Mashilo, J. (2021). Citron Watermelon Potential to Improve Crop Diversification and Reduce Negative Impacts of Climate Change. Sustainability, 13(4), 2269. https://doi.org/10.3390/su13042269