
Chilling and Heat Accumulation of Fruit and Nut Trees and Flower Bud Vulnerability to Early Spring Low Temperatures in New Mexico: Meteorological Approach





Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Data Collection
2.2. Data Management
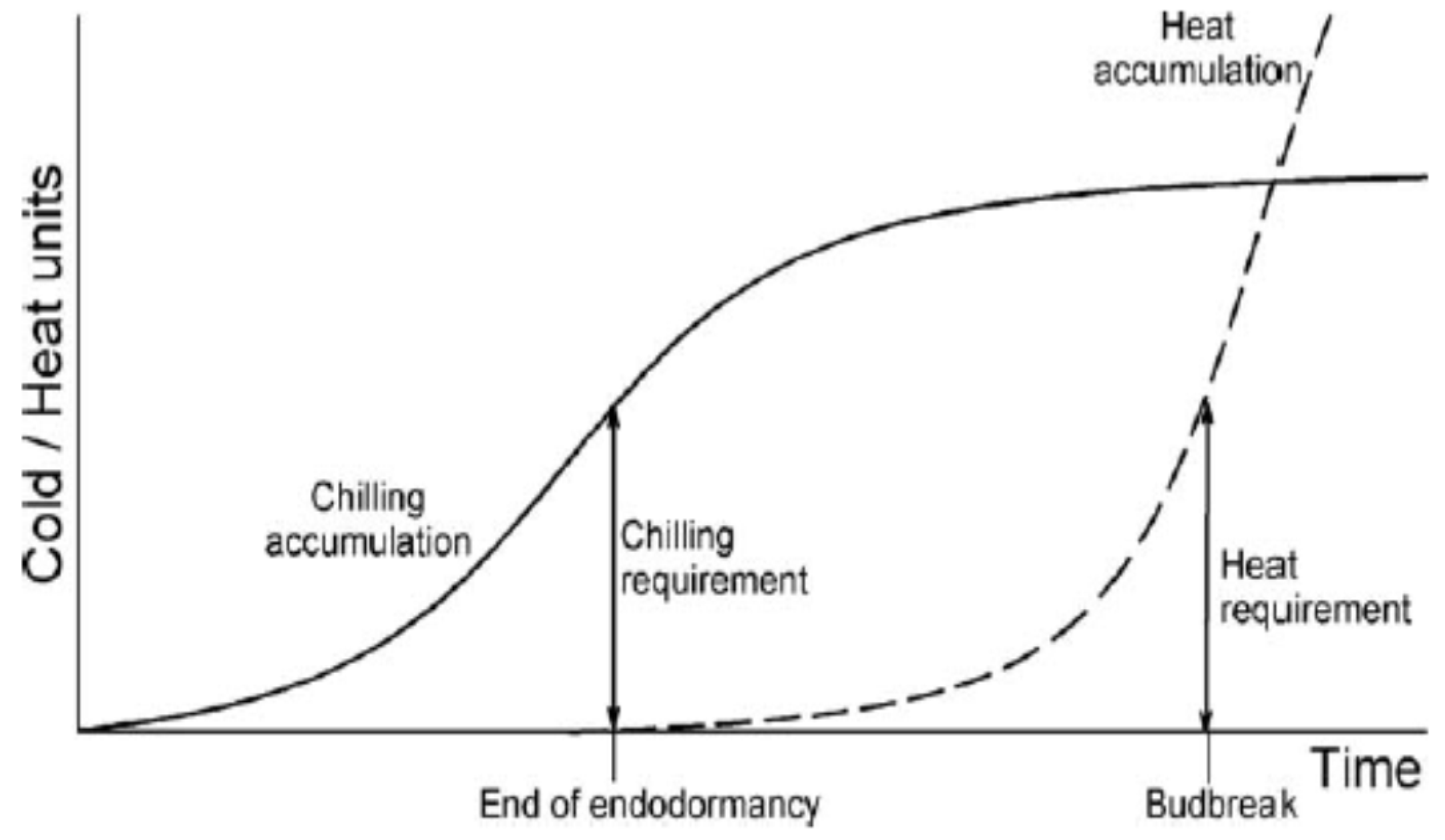
2.3. Estimation of the Heat and Chilling Accumulation: Heat and Chilling Model
- Forcing model: Growing Degree Hour (GDH) Model
- Chilling requirement of fruit trees: The Utah Model [23]
3. Results
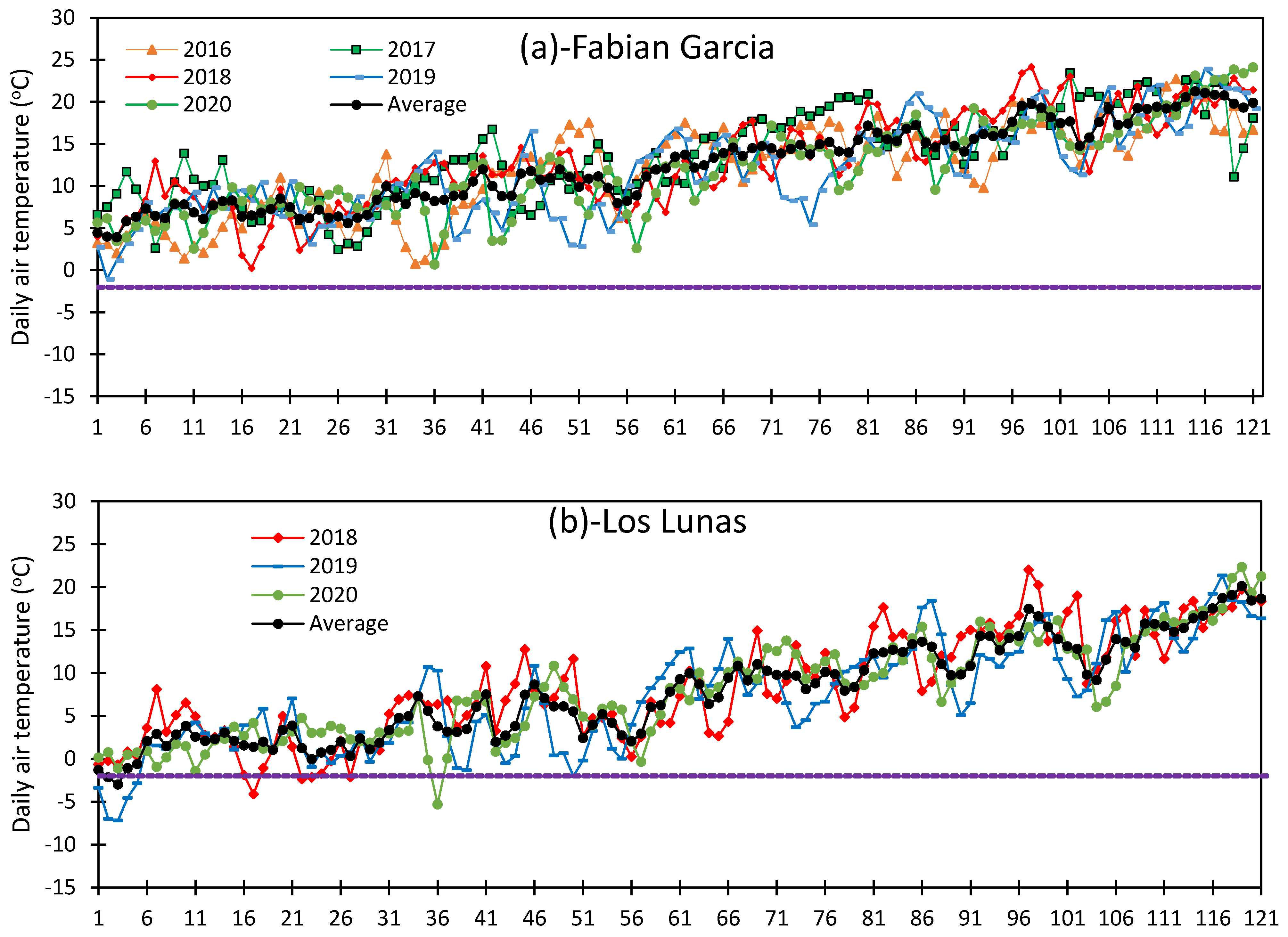
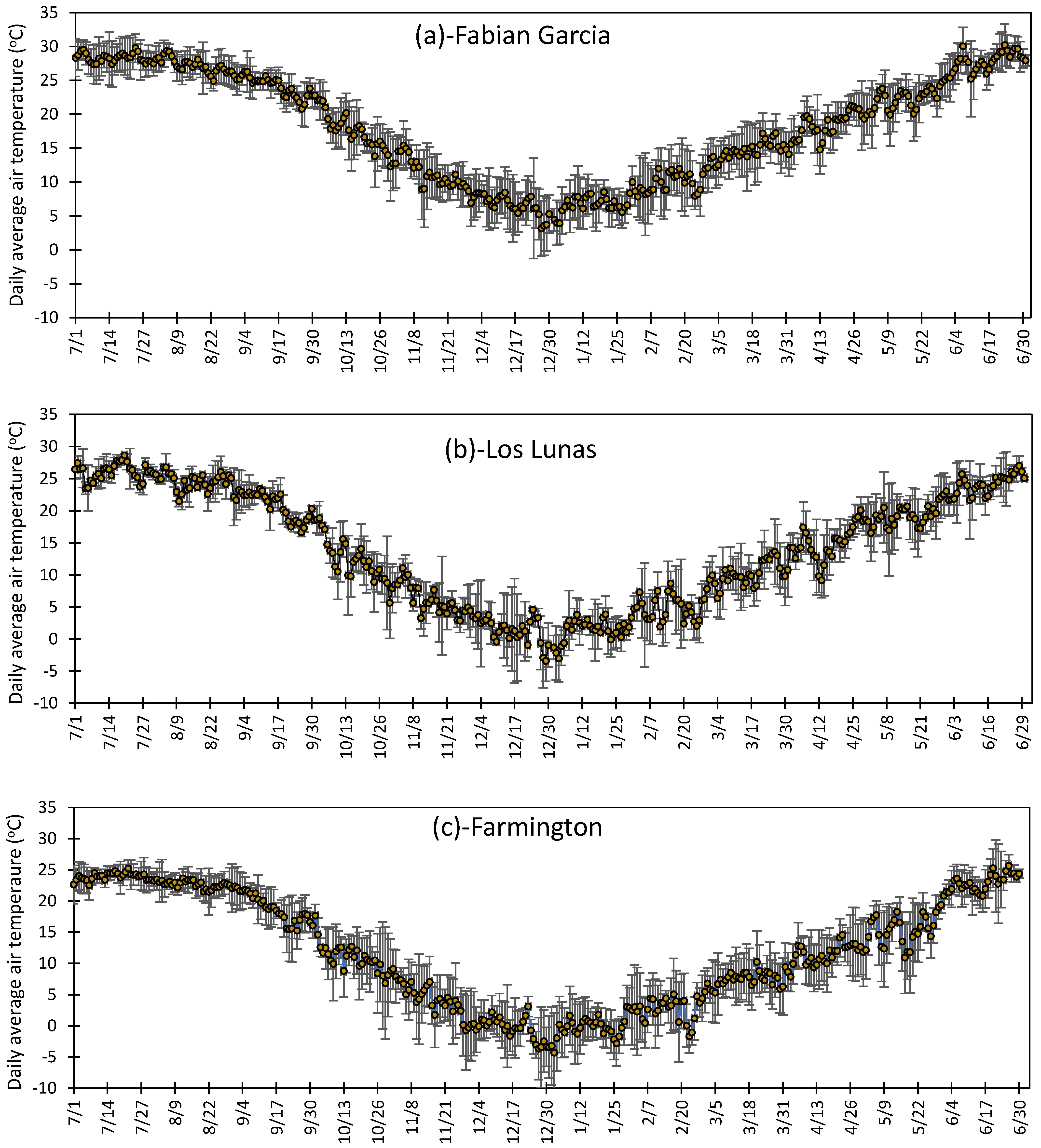
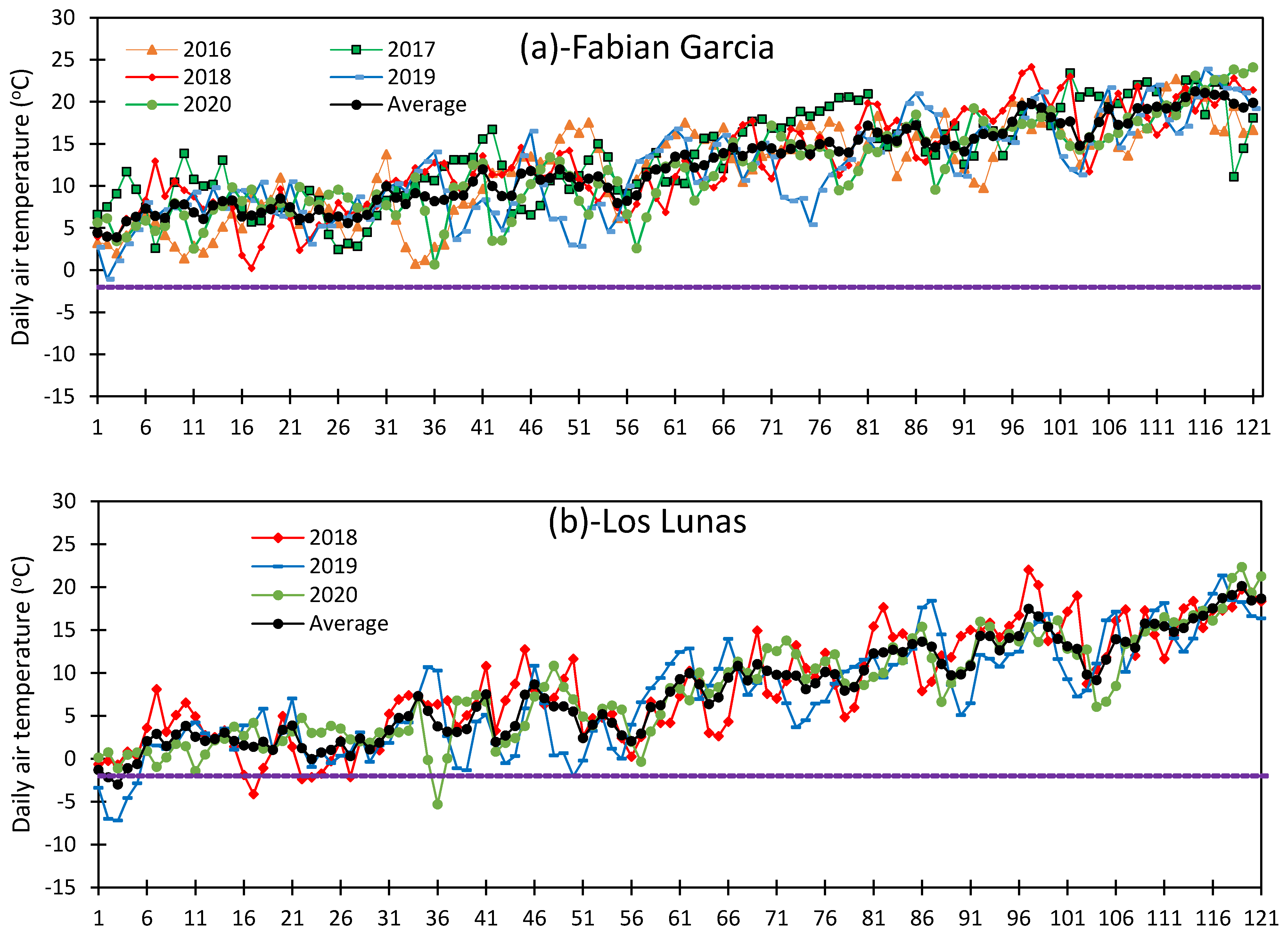
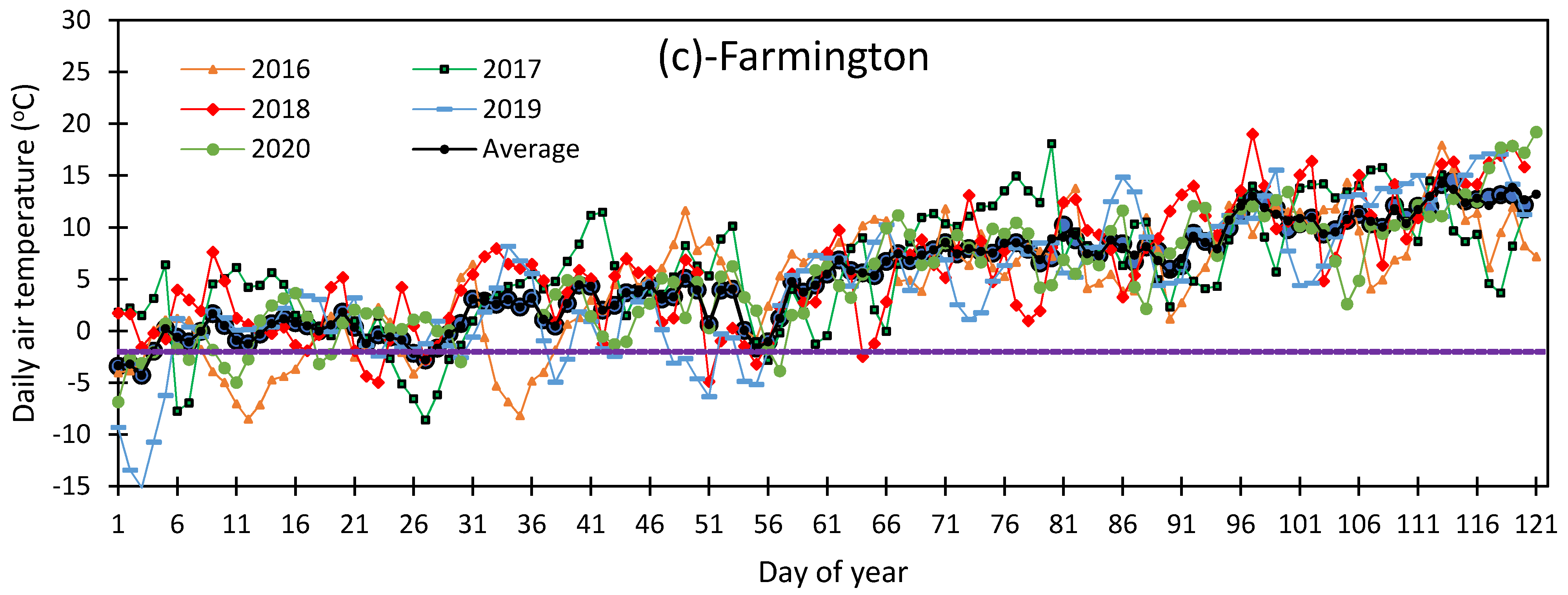
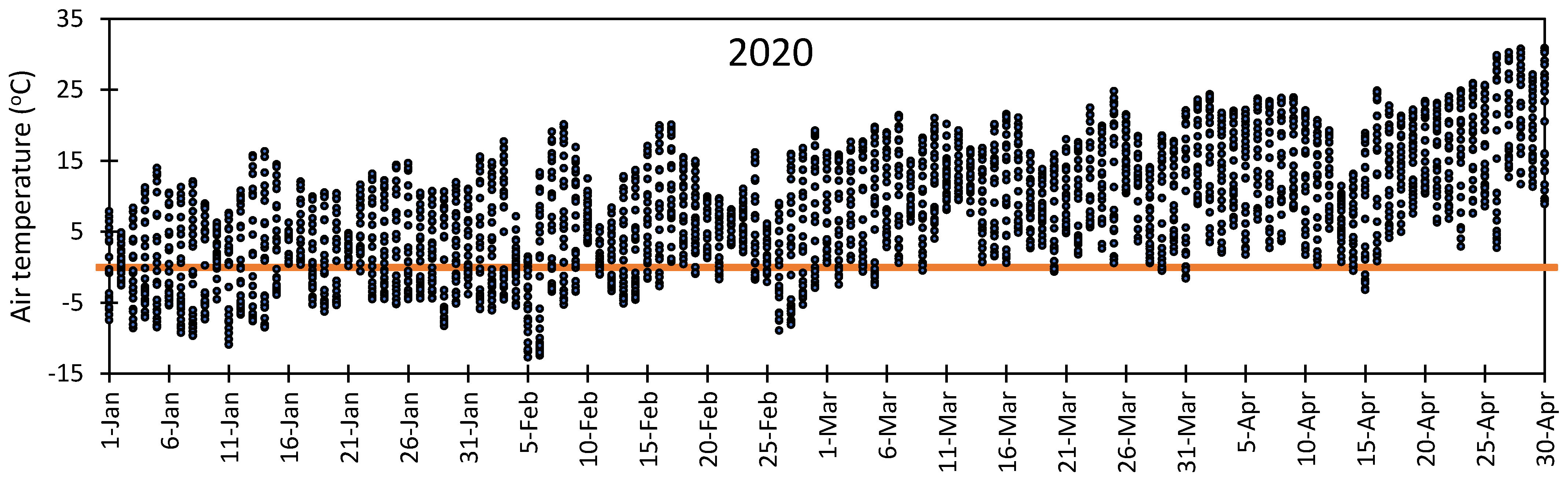
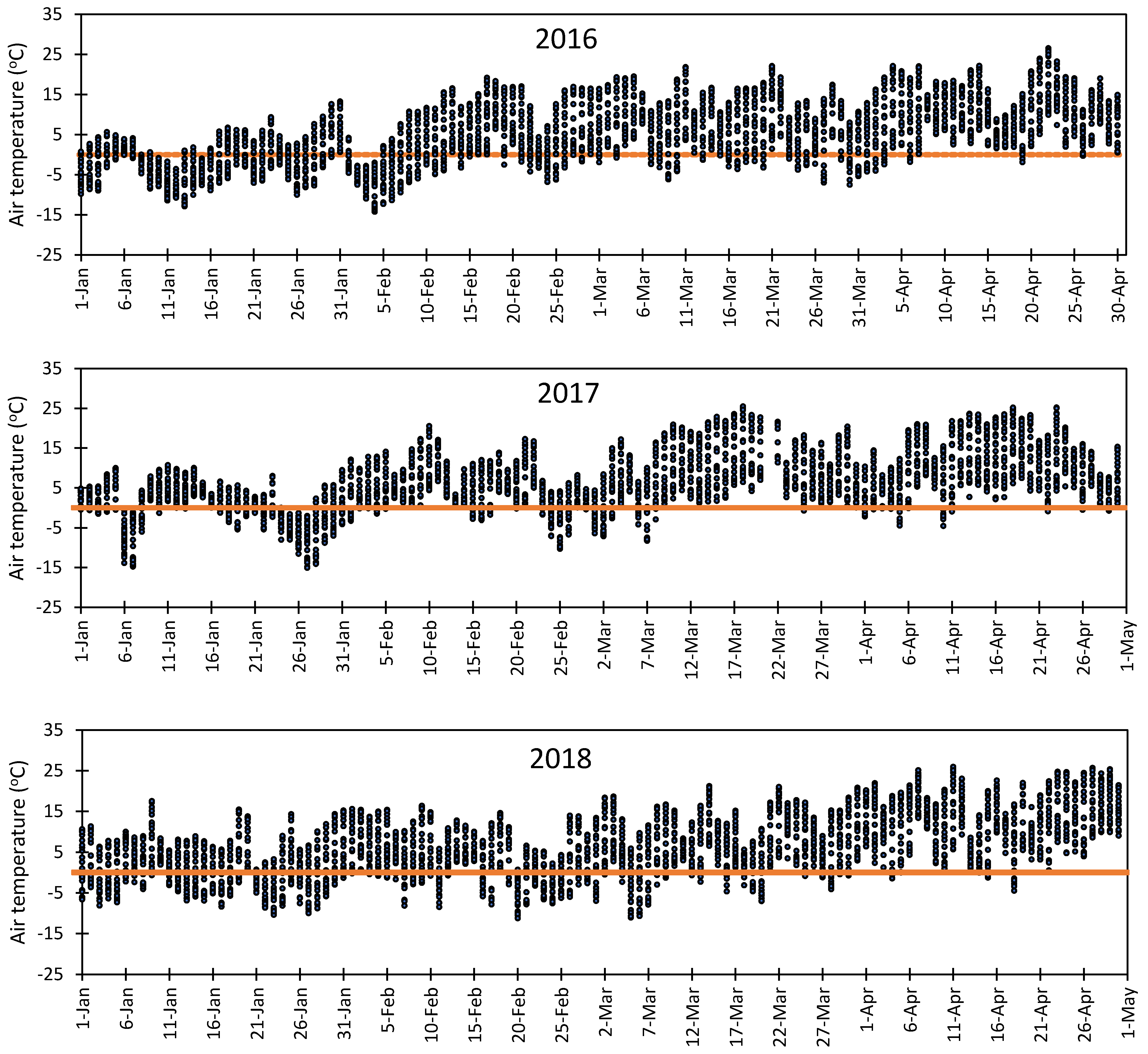
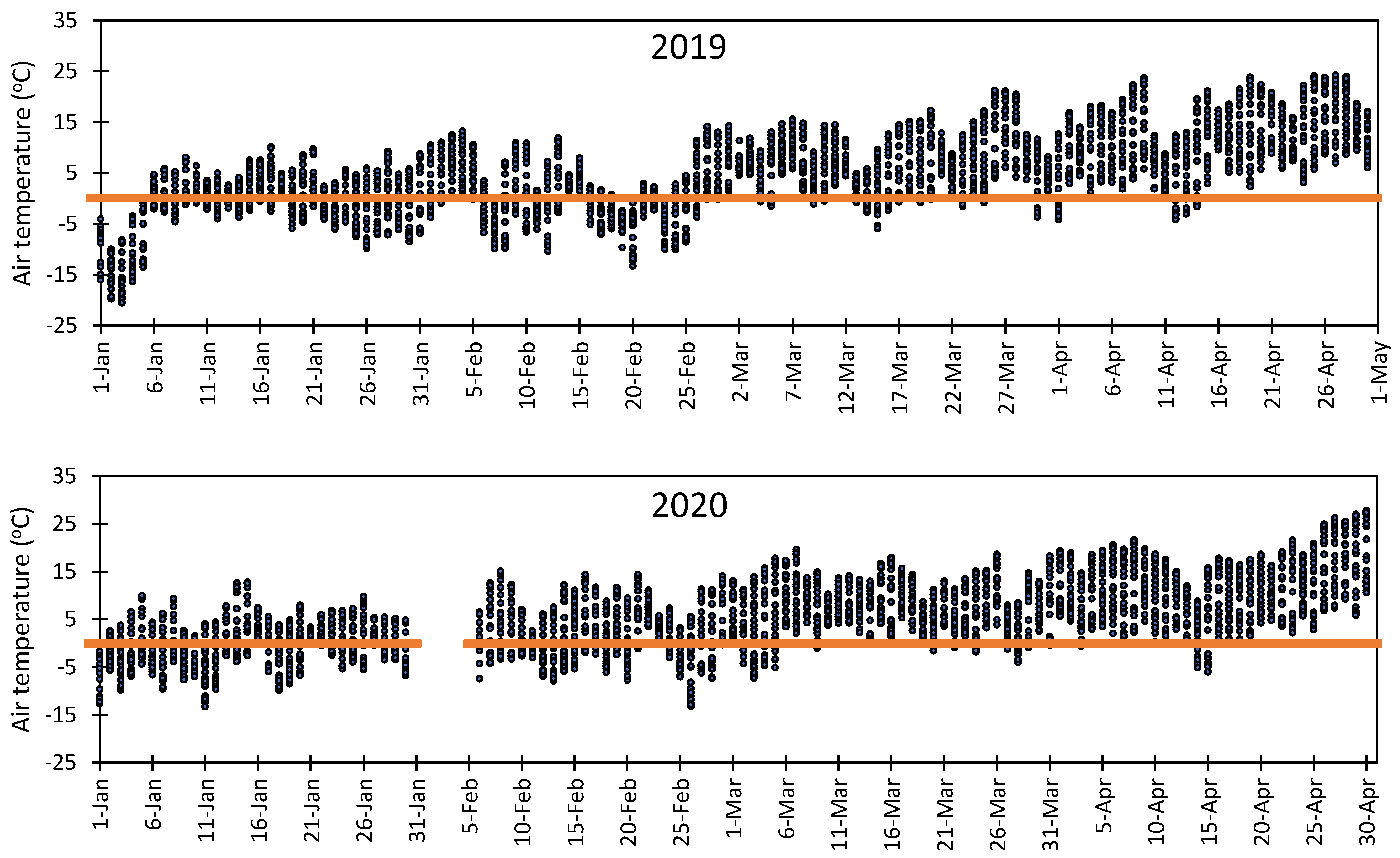
3.1. Variation in Air Temperature at Fabian Garcia, Los Lunas, and Farmington
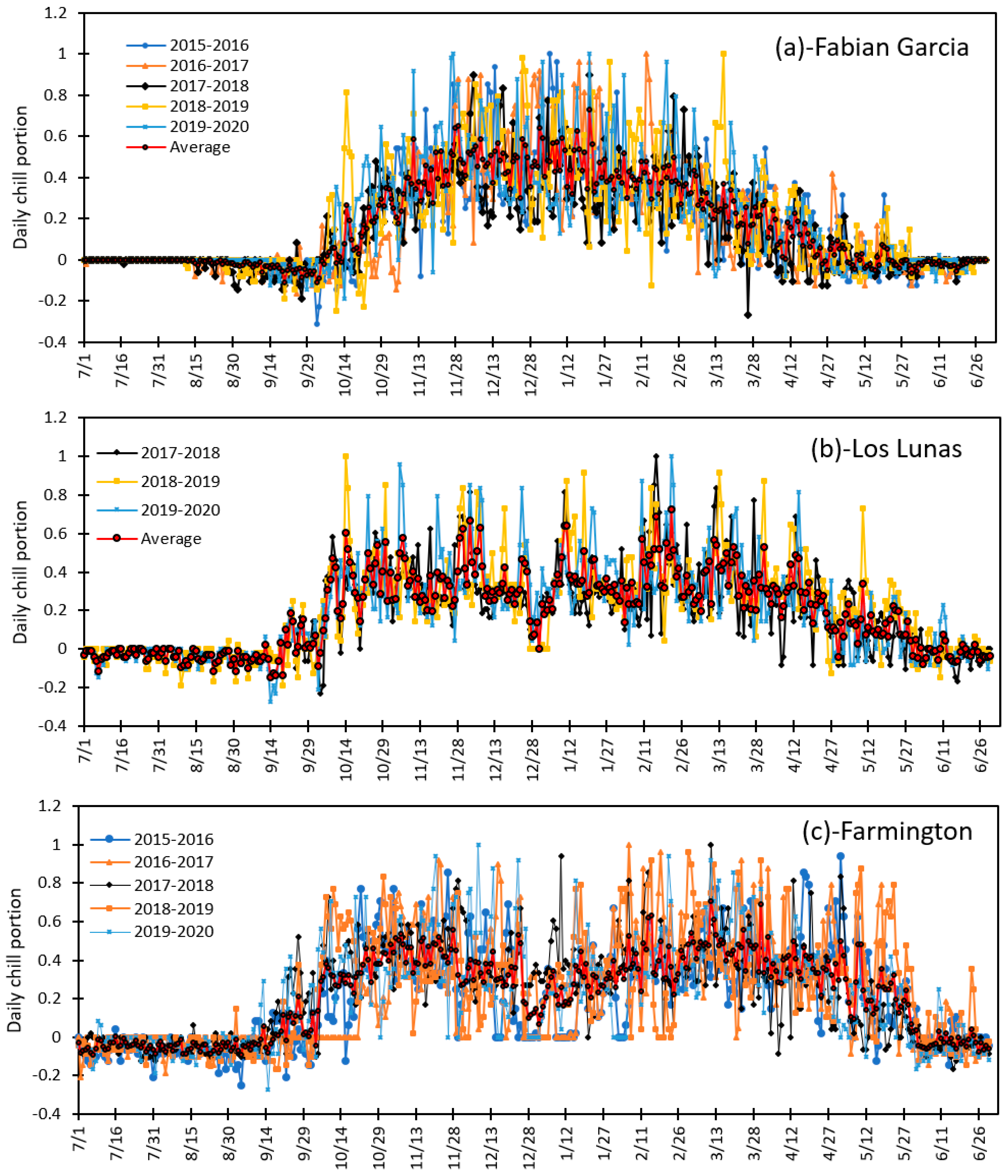
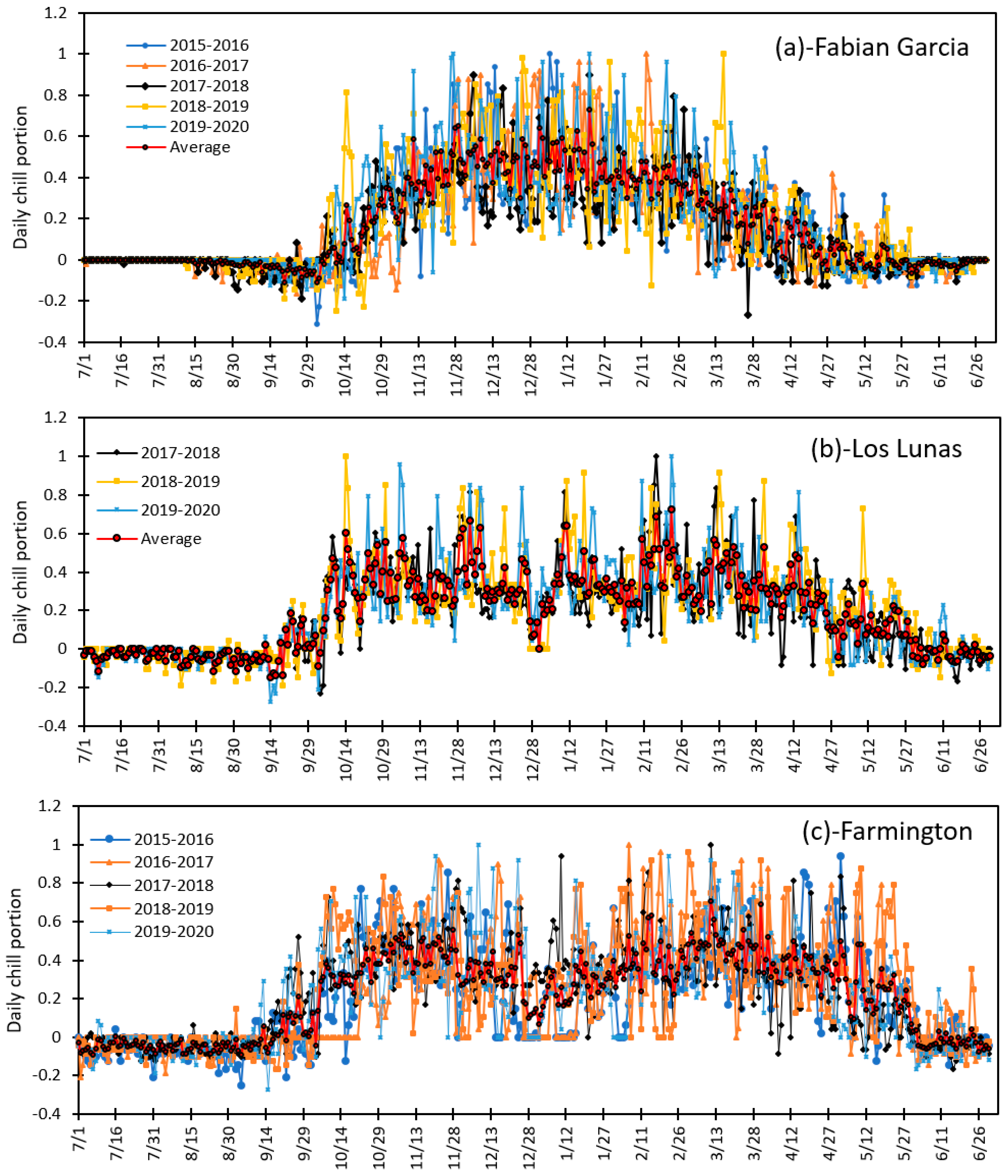
3.2. Variation in Daily Chill Portion and Seasonal Total Chill Hours for the Period of 2015 to 2020 at Fabian Garcia, Los Lunas, and Farmington
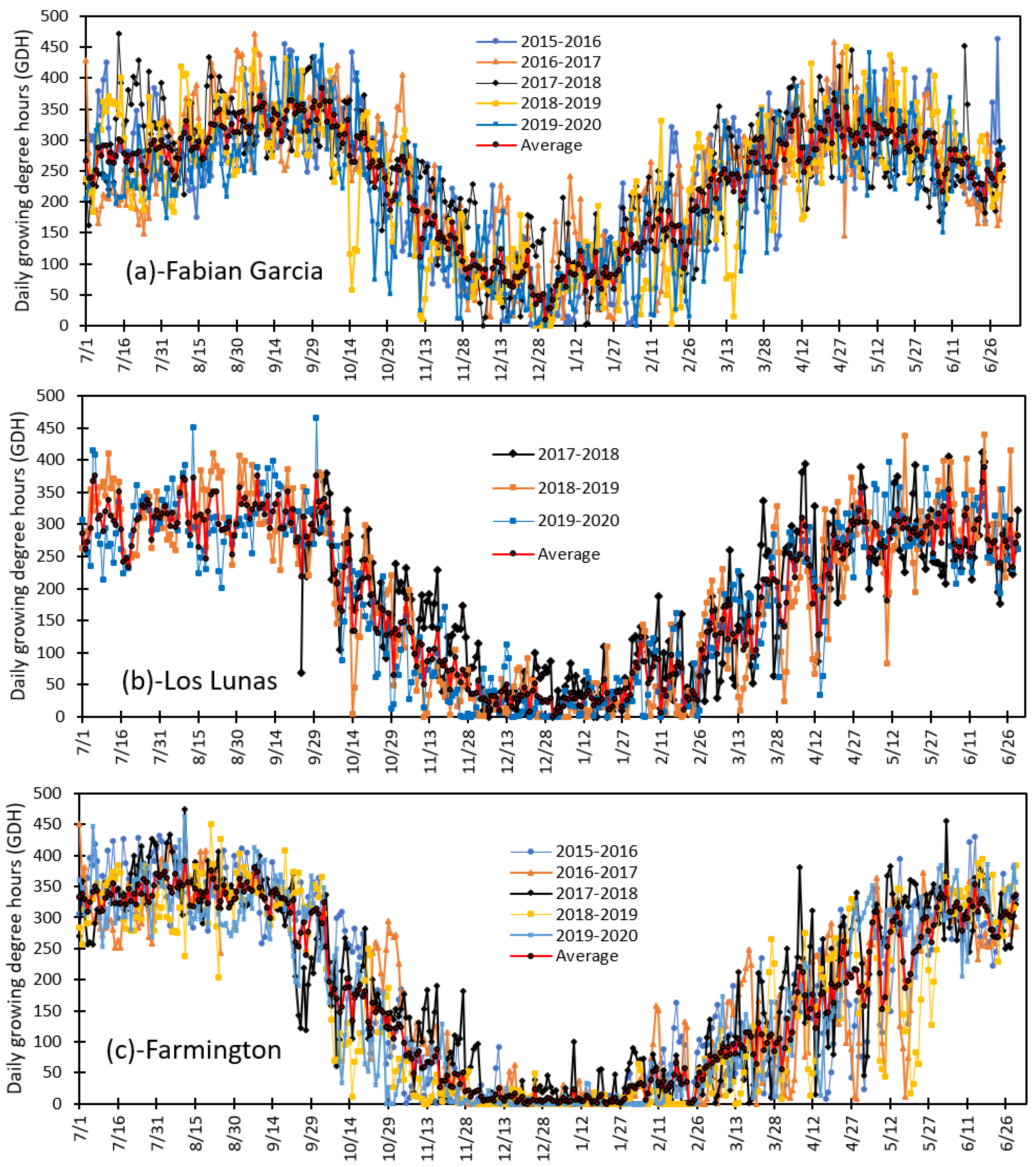
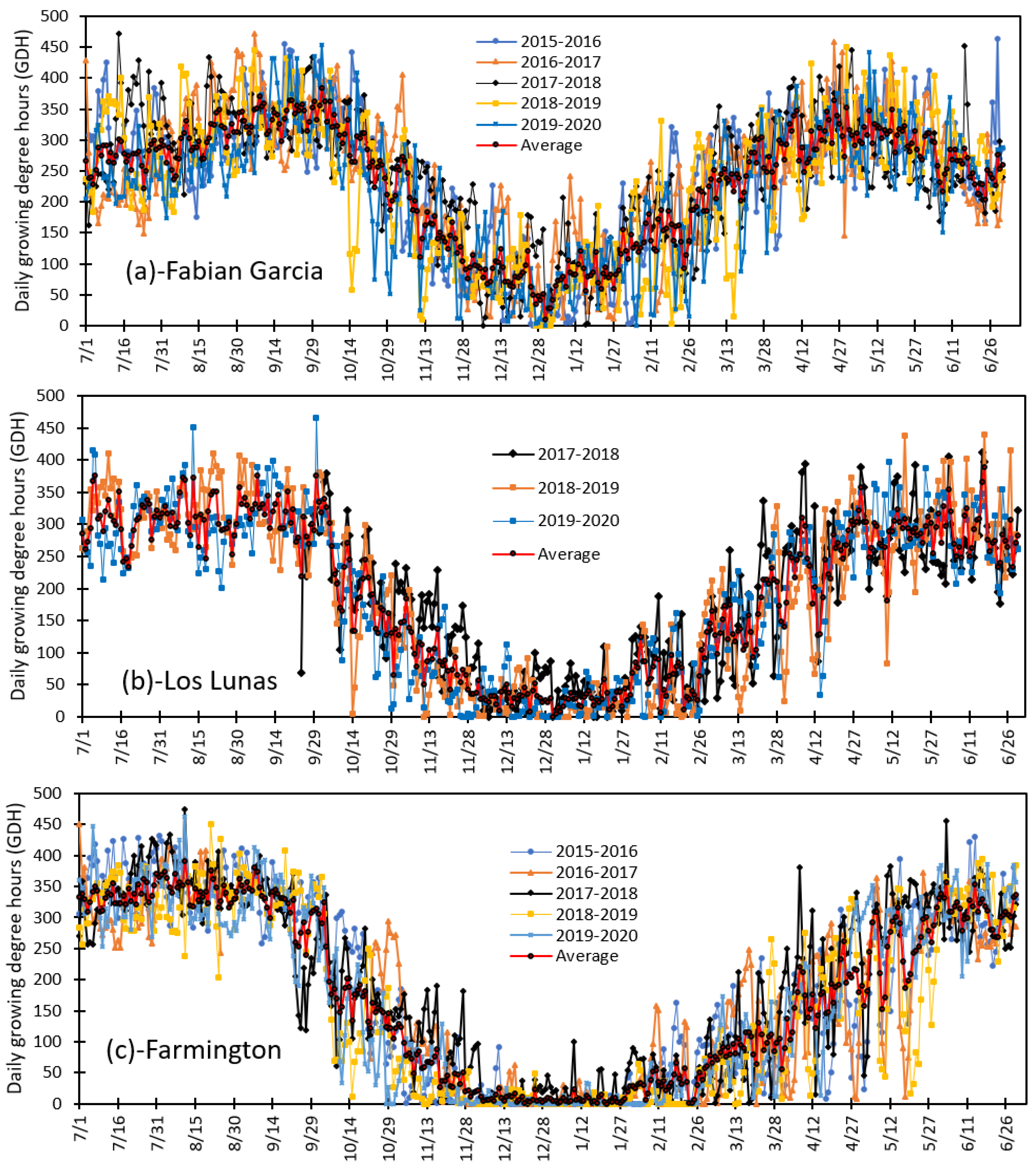
3.3. Variation in Daily Growing Degree Hours and Seasonal Total Heat Accumulated from 2015 to 2020 at Fabian Garcia, Los Lunas, and Farmington
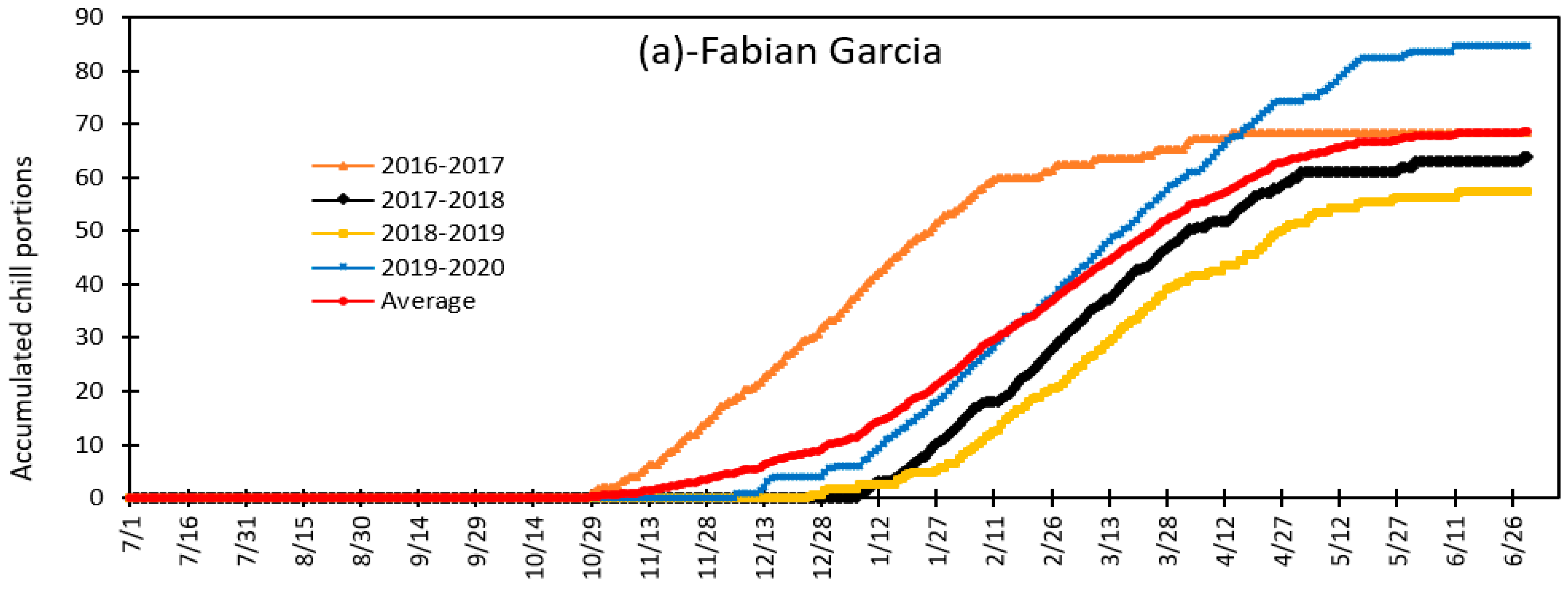
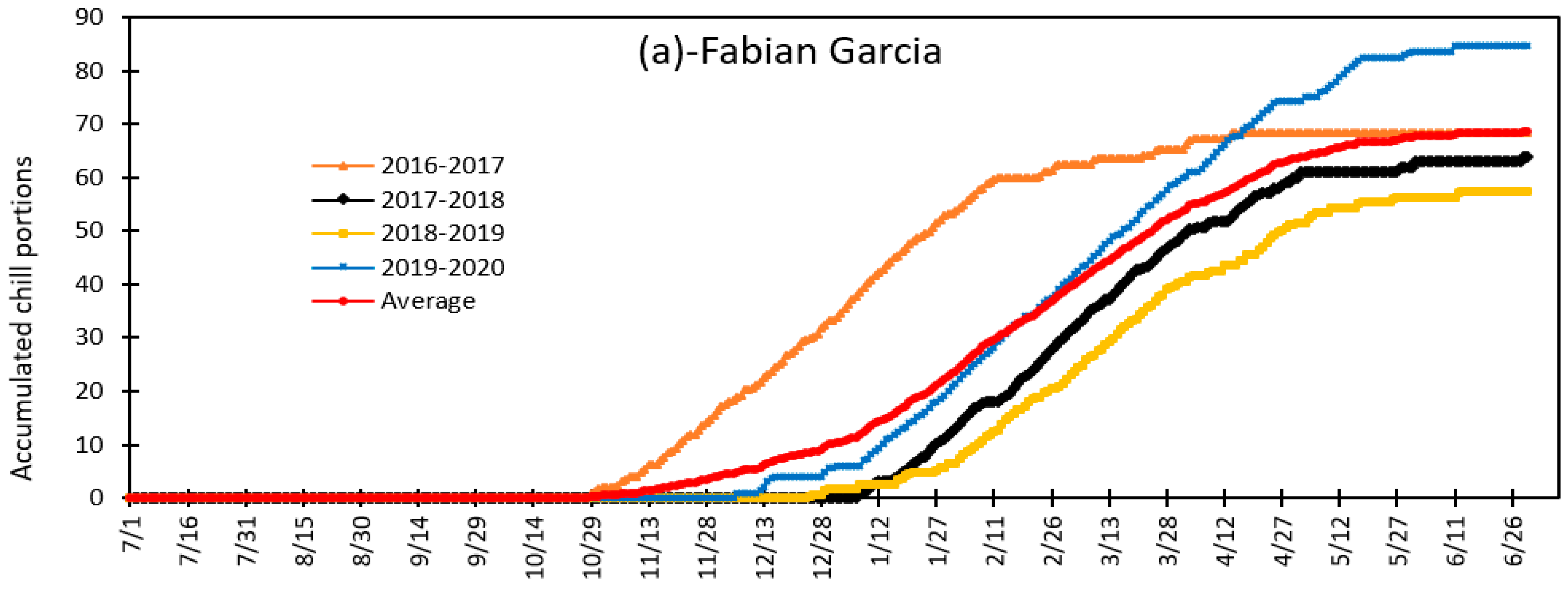
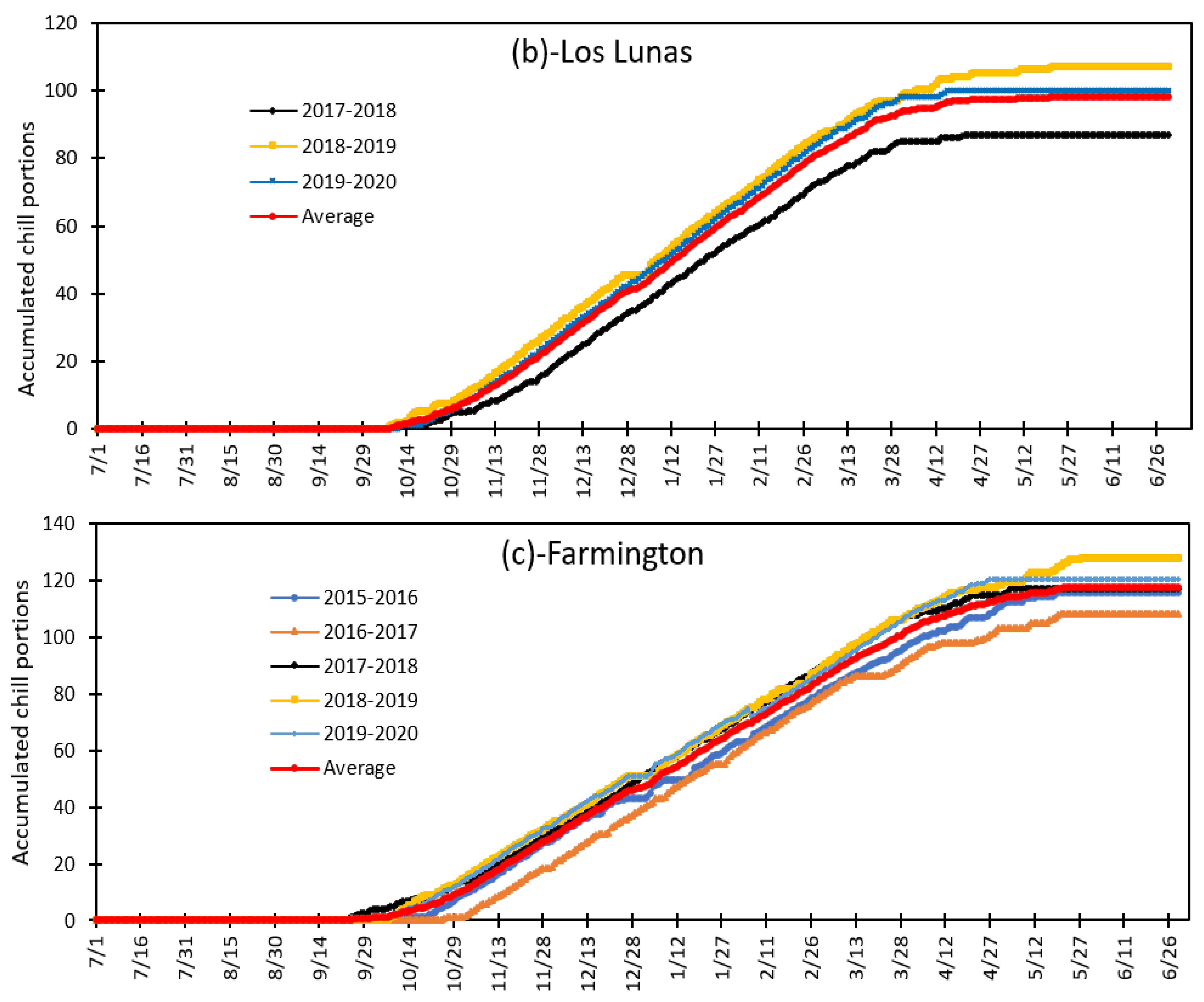
3.4. Variation in Daily Chill Portion and Seasonal Total Chill Portion during Fruit and Nut Trees Endodormancy at Fabian Garcia, Los Lunas and Farmington during the 2015 to 2020 Period
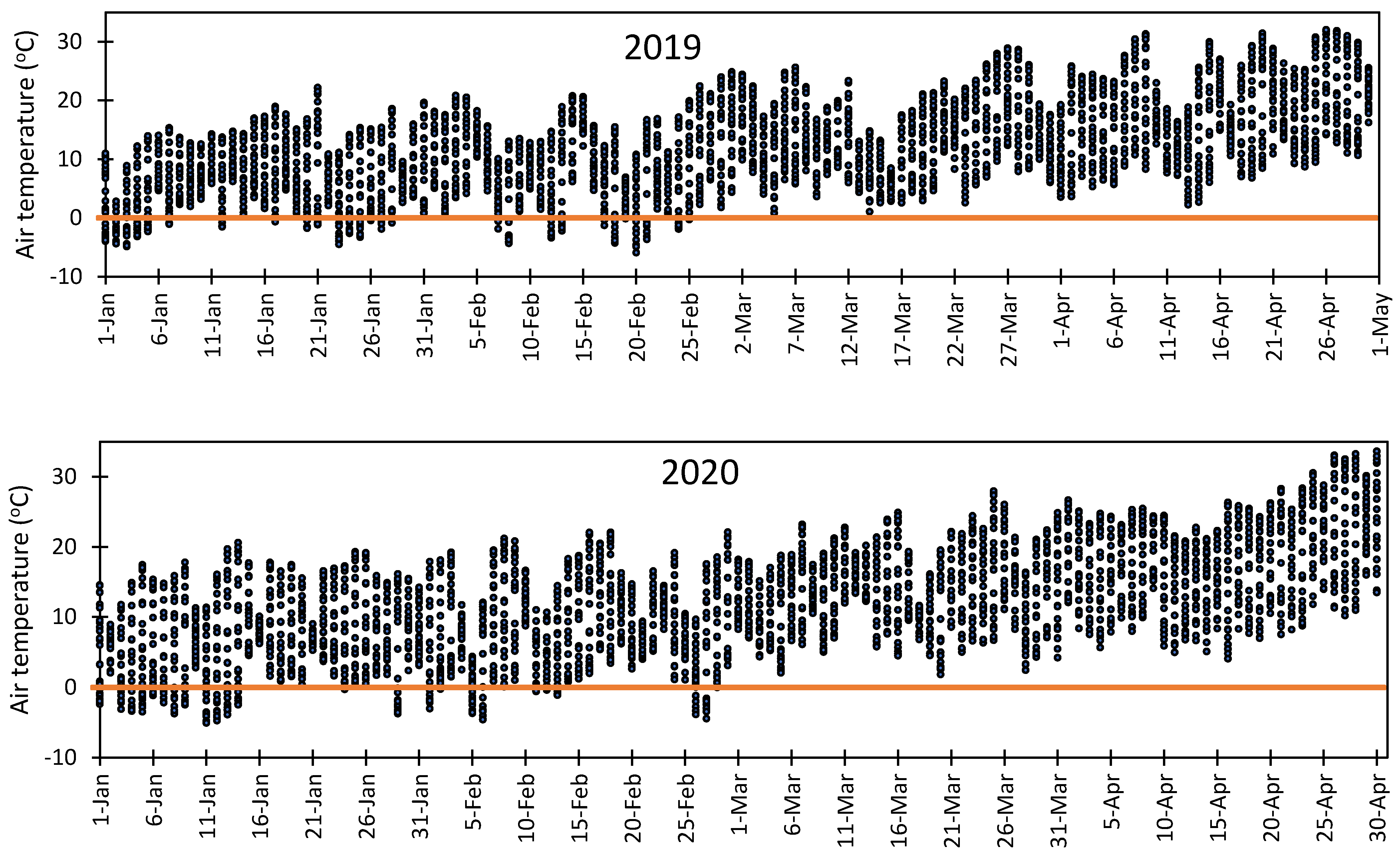
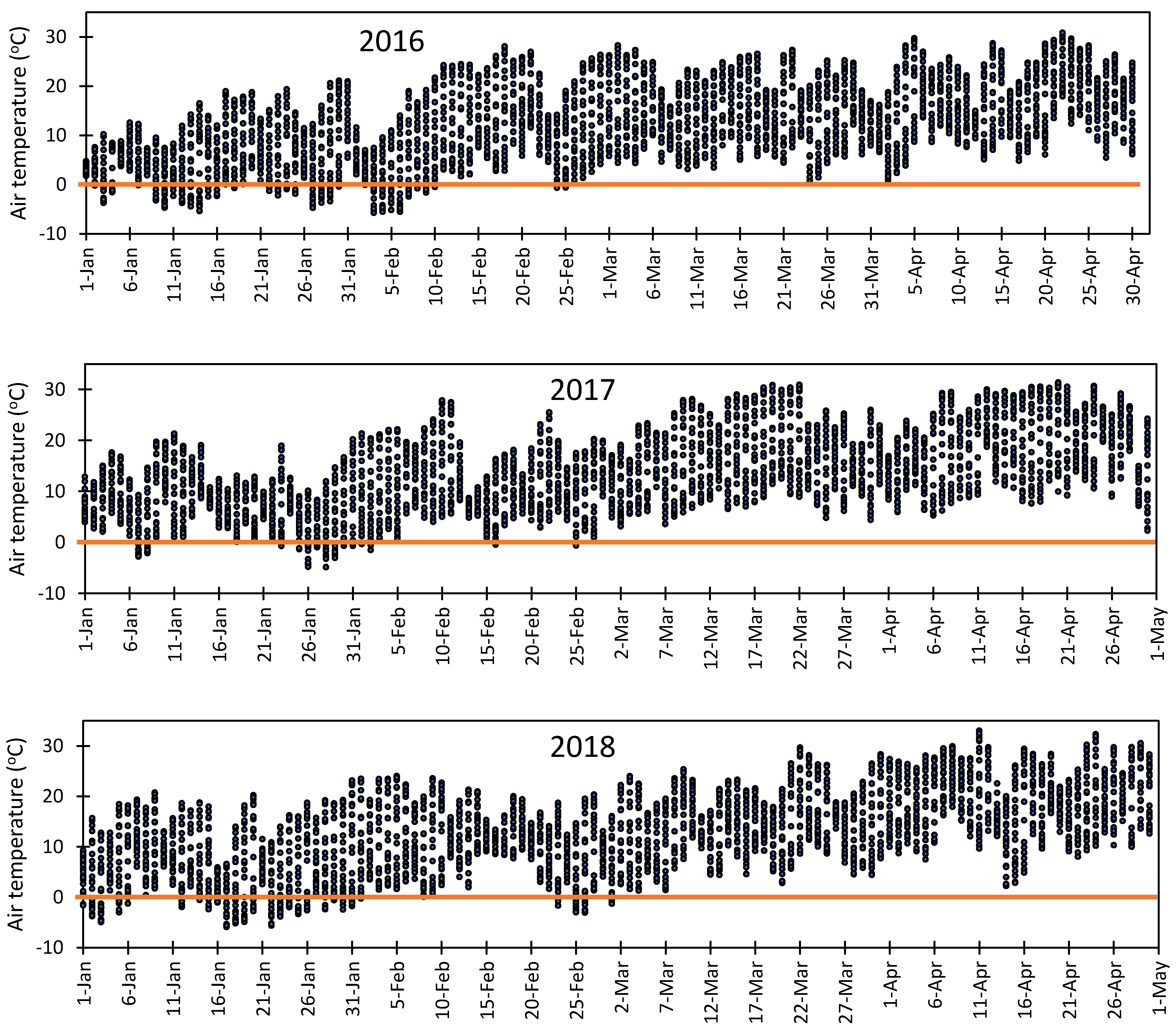
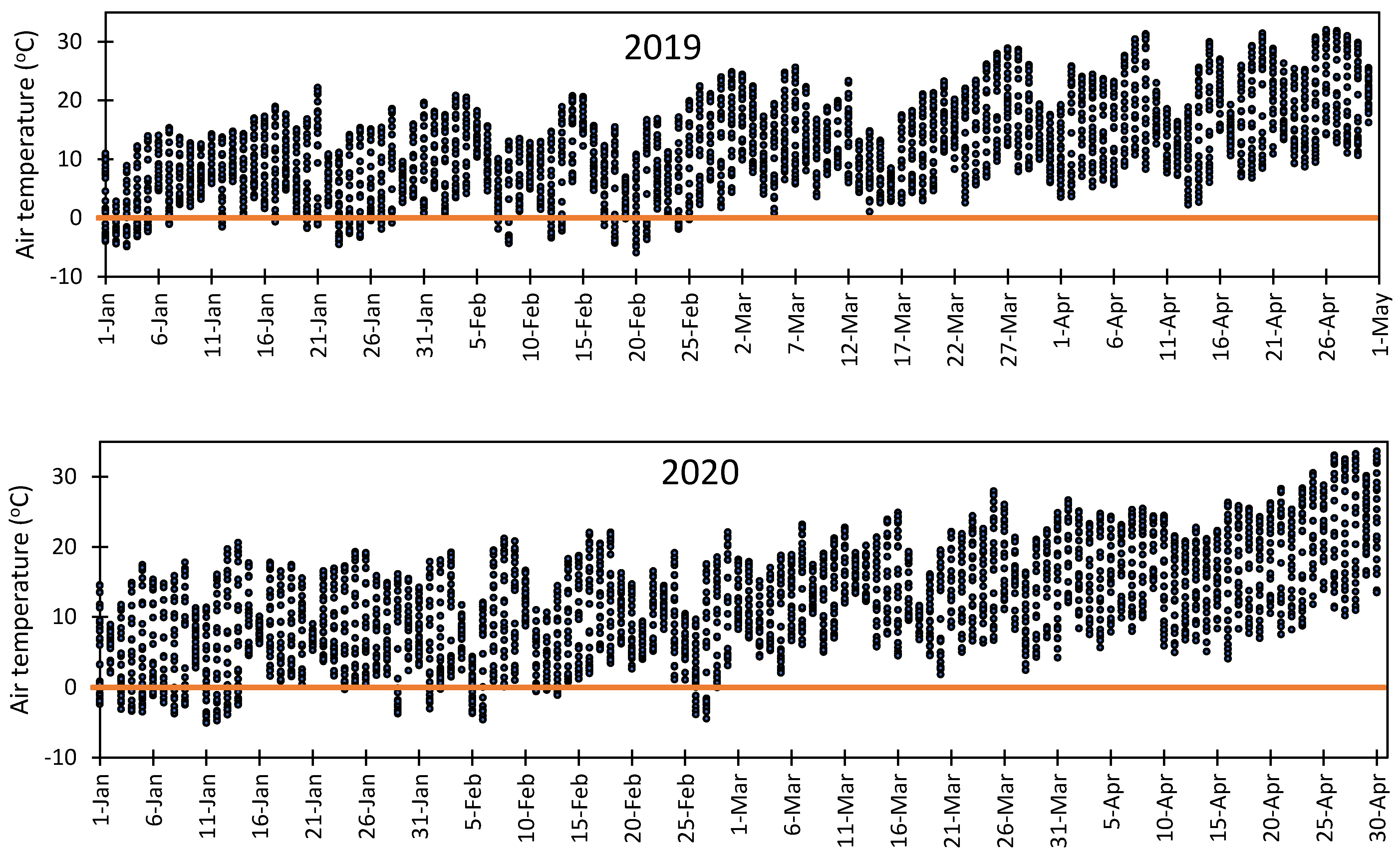
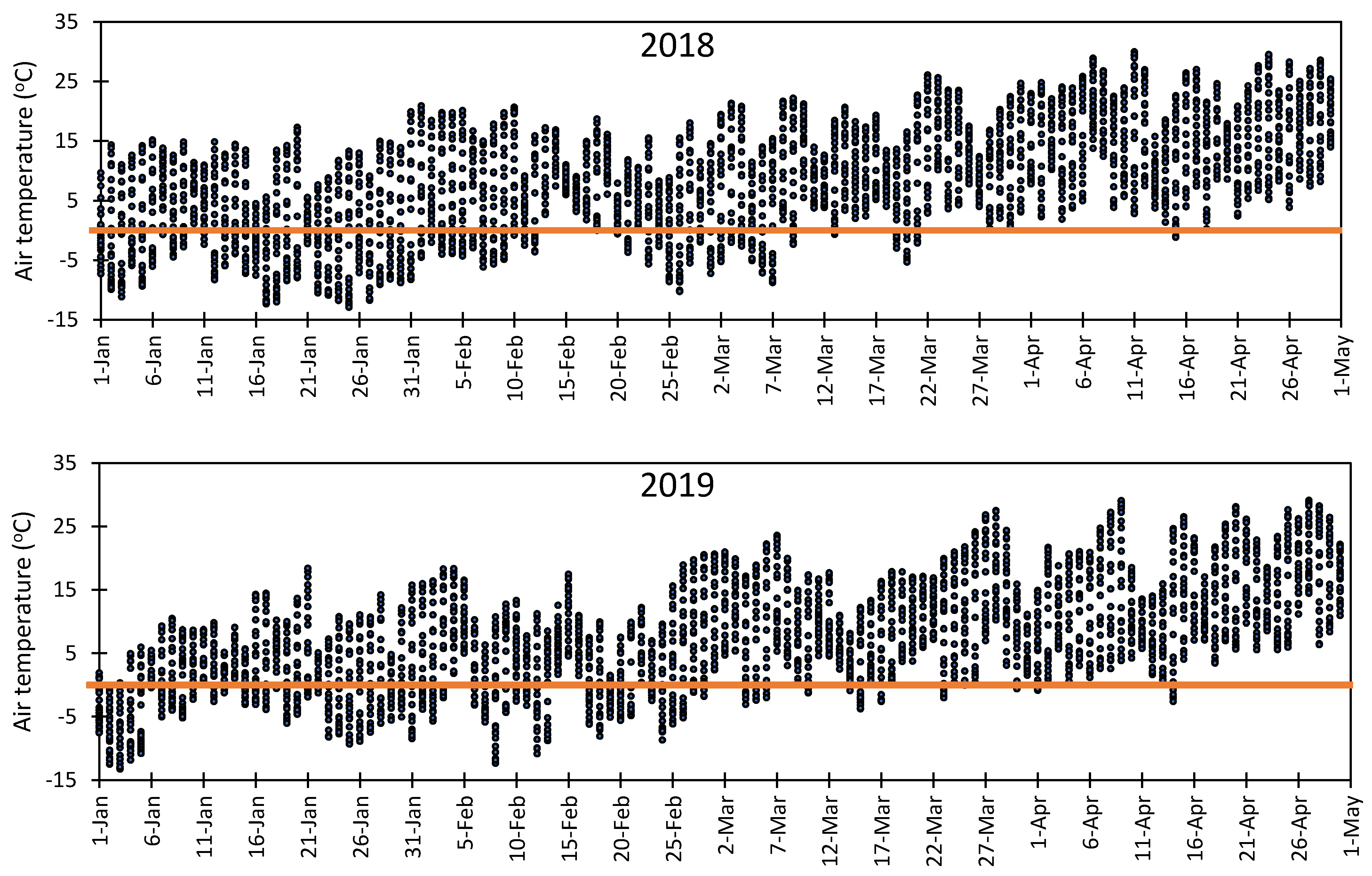
3.5. Vulnerability of Flower Bud to Early Spring Low Temperatures at Fabian Garcia, Los Lunas, and Farmington
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, B.K.C. Temperate and boreal forest tree phenology: From organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Gebauer, J.; Buerkert, A. Climate change effects on winter chill for tree crops with chilling requirements on the Arabian Peninsula. Clim. Chang. 2009, 96, 219–237. [Google Scholar] [CrossRef] [Green Version]
- Campoy, J.A.; Ruiz, D.; Cook, N.; Allderman, L.; Egea, J. Clinal variation of dormancy progression in apricot. S. Afr. J. Bot. 2011, 77, 618–630. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Brown, P.H. A global analysis of the comparability of winter chill models for fruit and nut trees. Int. J. Biometeorol. 2011, 55, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Vitasse, Y.; Signarbieux, C.; Fu, Y.H. Global warming leads to more uniform spring phenology across elevations. Proc. Natl. Acad. Sci. USA 2018, 115, 1004–1008. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.H.; Piao, S.; Vitasse, Y.; Zhao, H.; DeBoeck, H.J.; Liu, Q.; Yang, H.; Weber, U.; Hänninen, H.; Janssens, I.A. Increased heat requirement for leaf flushing in temperate woody species over 1980–2012: Effects of chilling, precipitation and insolation. Glob. Chang. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Piao, S.; Zhou, X.; Geng, X.; Hao, F.; Vitasse, Y.; Janssens, I.A. Short photoperiod reduces the temperature sensitivity of leaf-out in saplings of Fagus sylvatica but not in horse chestnut. Glob. Chang. Biol. 2019, 25, 1696–1703. [Google Scholar] [CrossRef]
- Hart, R.; Salick, J.; Ranjitkar, S.; Xu, J. Herbarium specimens show contrasting phenological responses to Himalayan climate. Proc. Natl. Acad. Sci. USA 2014, 111, 10615–10619. [Google Scholar] [CrossRef] [Green Version]
- Schieber, B.; Kubov, M.; Janík, R. Effects of climate warming on vegetative phenology of the common beech Fagus sylvatica in a submontane Forest of the Western Carpathians: Two decades analysis. Pol. J. Ecol. 2015, 65, 339–351. [Google Scholar] [CrossRef]
- Signarbieux, C.; Toledano, E.; Sanginés de Carcer, P.; Fu, Y.H.; Schlaepfer, R.; Buttler, A.; Vitasse, Y. Asymmetric effects of cooler and warmer winters on beech phenology last beyond spring. Glob. Chang. Biol. 2017, 23, 4569–4580. [Google Scholar] [CrossRef]
- Yu, L.; Han, Y.; Jiang, Y.; Dong, T.; Lei, Y. Sex-specific responses of bud burst and early development to nongrowing season warming and drought in Populus cathayana. Can. J. For. Res. 2018, 48, 68–76. [Google Scholar] [CrossRef]
- Fu, Y.H.; Zhao, H.; Piao, S.; Peaucelle, M.; Peng, S.; Zhou, G.; Ciais, P.; Huang, M.; Menzel, A.; Peñuelas, J.; et al. Declining global warming effects on the phenology of spring leaf unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1957, 22, 371–377. [Google Scholar]
- Basler, D.; Körner, C. Photoperiod and temperature responses of bud swelling and bud burst in four temperate forest tree species. Tree Physiol. 2014, 34, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Harrington, R.; Anton, C.; Dawson, T.P.; De Bello, F.; Feld, C.K.; Haslett, J.R.; Kluvánková, T.; Kontogianni, A.; Lavorel, S.; Luck, G.W.; et al. Ecosystem services and biodiversity conservation: Concepts and a glossary. Biodivers. Conserv. 2010, 19, 2773–2790. [Google Scholar] [CrossRef]
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Xu, J.C.; Dai, J.H.; Cheng, J.M.; Luedeling, E. Statistical identification of chilling and heat requirements for apricot flower buds in Beijing China. Sci. Hortic. 2015, 195, 138–144. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.G.; Ma, Q.; Hänninen, H.; Rossi, S.; Piao, S.; Bergeron, Y. Spring phenology at different altitudes is becoming more uniform under global warming in Europe. Glob. Chang. Biol. 2018, 24, 3969–3975. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Dai, J.; Ranjitkar, S.; Yu, H.; Xu, J.; Luedeling, E. Chilling and heat requirements for flowering in temperate fruit trees. Int. J. Biometeorol. 2014, 58, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Luedeling, E.; Gassner, A. Partial Least Squares regression foranalyzing walnut phenology in California. Agric. For. Meteorol. 2012, 158, 43–52. [Google Scholar]
- Luedeling, E.; Guo, L.; Dai, J.; Leslie, C.; Blanke, M.M. Differential responses of trees to temperature variation during the chilling and forcing phases. Agric. For. Meteorol. 2013, 181, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, J.H. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for Redhaven and Elberta peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Computer simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Mathematical analysis of a two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Shaltout, A.D.; Unrath, C.R. Rest completion prediction model for starkrimson delicious apples. J. Am. Soc. Hortic. Sci. 1983, 108, 957–961. [Google Scholar]
- Melo-Abreu, J.P.; Barranco, D.; Cordeiro, A.M.; Tous, J.; Rogadoe, B.M.; Villalobos, F.J. Modelling olive flowering date using chilling for dormancy release and thermal time. Agric. For. Meteorol. 2004, 125, 117–127. [Google Scholar]
- Linsley-Noakes, G.C.; Allan, P. Comparison of 2 models for the prediction of rest completion in peaches. Sci. Hortic. 1994, 59, 107–113. [Google Scholar] [CrossRef]
- Gilreath, P.R.; Buchanan, D.W. Rest prediction model for low-chilling Sungold nectarine. J. Am. Soc. Hort. Sci. 1981, 106, 426–429. [Google Scholar]
- Linvill, D.E. Calculating chilling hours and chill units from daily maximum and minimum temperature observations. HortScience 1990, 25, 14–16. [Google Scholar] [CrossRef] [Green Version]
- Erez, A.; Fishman, S.; Linsley-Noakes, G.C.; Allan, P. The dynamic model for rest completion in peach buds. Acta Hortic. 1990, 276, 165–174. [Google Scholar] [CrossRef]
- Erez, A. Bud dormancy; phenomenon, problems and solutions in the tropics and subtropics. In Temperate Fruit Crops in Warm Climates; Erez, A., Ed.; Kluwer Academic: Dordrecht, The Netherlands, 2000; pp. 17–48. [Google Scholar]
- Cesaraccio, C.; Spano, D.; Snyder, R.L.; Duce, P. Chilling and Forcing model to predict bud-burst of crop and forest species. Agric. For. Meteorol. 2004, 126, 1–13. [Google Scholar] [CrossRef]
- Ruiz, D.; Campoy, J.A.; Egea, J. Chilling and heat requirements of apricot cultivars for flowering. Environ. Exp. Bot. 2007, 61, 254–263. [Google Scholar] [CrossRef]
- Perez, F.J.; Vergara, R.; Or, E. On the mechanism of dormancy release in grapevine buds: A comparative study between hydrogen cyanamide and sodium azide. Plant Growth Regul. 2009, 59, 145–152. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.H.; Girvetz, E.H. Climatic changes lead to declining winter chill for fruit and nut trees in California during 1950–2099. PLoS ONE 2009, 4, e6166. [Google Scholar] [CrossRef]
- Luedeling, E.; Hale, A.; Zhang, M.; Bentley, W.J.; Dharmasri, L.C. Remote sensing of spider mite damage in California peachorchards. Int. J. Appl. Earth Obs. 2009, 11, 244–255. [Google Scholar] [CrossRef]
- Luedeling, E.; Blanke, M.; Gebauer, J. Climate change effects on winter chill for fruit crops in Germany-Auswirkungen des Klimawandels auf die Verfügbarkeit von Kältewirkung (Chilling) für Obstgehölze in Deutschland. Erwerbs-Obstbau 2009, 51, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Zhang, M.H.; Luedeling, V.; Girvetz, E.H. Sensitivity of winter chill models for fruit and nut trees to climatic changes expected in California’s central Valley. Agric. Ecosyst. Environ. 2009, 133, 23–31. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.; McGranahan, G.; Leslie, C. Validation of winter chill models using historic records of walnut phenology. Agric. For. Meteorol. 2009, 149, 1854–1864. [Google Scholar] [CrossRef]
- Zhang, J.L.; Taylor, C. The dynamic model provides the best description of the chill process on’ Sirora’ pistachio trees in Australia. HortScience 2011, 46, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Girvetz, H.; Semenov, M.A.; Brown, P.H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 2011, 6, 13. [Google Scholar] [CrossRef]
- Luedeling, E.; Kunz, A.; Blanke, M.M. Identification of chilling and heat requirements of cherry trees a statistical approach. Int. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, E.; Whitney, C.; Cuneo, I.F.; Luedeling, E. Prospects of decreasing winter chill for deciduous fruit production in Chile throughout the 21st century. Clim. Chang. 2020, 159, 423–439. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Anderson, J.L.; Richardson, E.A.; Kesner, C.D. Validation of chill unit and flower bud phenology models for “Montmorency” sour cherry. Acta Hortic. 1986, 184, 71–78. [Google Scholar] [CrossRef]
- Djaman, K.; O’Neill, M.; Owen, C.K.; Smeal, D.; Koudahe, K.; West, M.; Allen, S.; Lombard, K.; Irmak, S. Crop Evapotranspiration, Irrigation Water Requirement and Water Productivity of Maize from Meteorological Data under Semiarid Climate. Water 2018, 10, 405. [Google Scholar] [CrossRef] [Green Version]
- Powel, A.; Dozier, W.; Williams, D.; Himelrick, D. Fruit Culture in Alabama. Winter Cgilling Requirements; The Alabama Cooperative Extension System (Alabama A&M University and Auburn University): Auburn, AL, USA, 2018.
- Alburquerque, N.; García-Montiel, F.; Carrillo, A.; Burgos, L. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ. Exp. Bot. 2008, 64, 162–170. [Google Scholar] [CrossRef]
- Kwon, J.H.; Nam, E.Y.; Yun, S.K.; Kim, S.J.; Song, S.Y.; Lee, J.H.; Hwang, K.D. Chilling and heat requirement of peach cultivars and changes in chilling accumulation spectrums based on 100-year records in Republic of Korea. Agric. For. Meteorol. 2020, 288, 108009. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Mimoun, M.B.; Luedeling, E. Chilling and heat requirement for local and foreign almond (Prunus dulcis Mill.) cultivars in a Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- USDA-NASS. Census of Agriculture USDA; National Agricultural Statistics Service: Washington, DC, USA, 2019; p. 820.
- Yao, S.; Heerema, R. Fruits and Nuts for New Mexico Orchards; NMSU, Cooperative Extension Service, College of Agricultural, Consumer and Environmental Sciences: Las Cruces, NM, USA, 2014; Available online: http://aces.nmsu.edu/pubs/_h/H310.pdf (accessed on 15 December 2020).
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration: Guide-lines for computing crop water requirements–FAO Irrigation and Drainage Paper 56. FAO Rome 1998, 300, D05109. [Google Scholar]
- Darbyshire, R.; Webb, L.; Goodwin, I.; Barlow, S. Winter chilling trends for deciduous fruit trees in Australia. Agric. For. Meteorol. 2011, 151, 1074–1085. [Google Scholar] [CrossRef]
- Djaman, K.; Owen, C.; West, M.M.; Allen, S.; Koudahe, K.; Darapuneni, M.; O’Neill, M. Relationship between Relative Maturity and Grain Yield of Maize (Zea mays L.) Hybrids in Northwest New Mexico for the 2003–2019 Period. Agriculture 2020, 10, 290. [Google Scholar] [CrossRef]
- Studer, S.; Stöckli, R.; Appenzeller, C. A comparative study of satellite and ground-based phenology. Int. J. Biometeorol. 2007, 51, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Shi, J. Evaluation and comparison of growing season metrics in arid and semi-arid areas of northern China under climate change. Ecol. Indic. 2021, 121, 107055. [Google Scholar] [CrossRef]
- Fan, D.; Zhao, X.; Zhu, W.; Sun, W.; Qiu, Y. An improved phenology model for monitoring green-up date variation in Leymus chinensis steppe in Inner Mongolia during 1962–2017. Agric. For. Meteorol. 2020, 291, 108091. [Google Scholar] [CrossRef]
- White, M.A.; Hoffman, F.; Hargrove, W.W. A global framework formonitoring phenologcal responses to climate change. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.D.; Reed, B.C.; White, M.A. Assessing satellite-derived start-of-season (SOS) measures in the conterminous USA. Int. J. Climatol. 2002, 22, 1793–1805. [Google Scholar] [CrossRef]
- Nezval, O.; Krejza, J.; Světlík, J.; Šigut, L.; Horáček, T. Comparison of traditional ground-based observations and digital remote sensing of phenological transitions in a floodplain forest. Agric. For. Meteorol. 2020, 291, 108079. [Google Scholar] [CrossRef]
- Rodrigo, J. Spring frost in deciduous fruit trees morphological damage and flower hardiness. Sci. Hortic. 2000, 85, 155–173. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Götz, K.P.; Weber, K.C.; Moryson, S. Climate change and spring frost damages for sweet cherries in Germany. Int. J. Biometeorol. 2018, 62, 217–228. [Google Scholar] [CrossRef]
- Cittadini, E.D.; de Ridder, N.; Peri, P.L.; Keulen, H.V. A method for assessing frost damage risk in sweet cherry orchards of South Patagonia. Agric. For. Meteorol. 2006, 141, 235–243. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 2008, 89, 353–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, R.S. Plant freezing and damage. Ann. Bot. 2001, 87, 417–424. [Google Scholar] [CrossRef]
- Salazar-Gutierrez, M.R.; Chaves, B.; Anothai, J.; Whiting, M.; Hoogenboom, G. Variation in cold hardiness of sweet cherry flower buds through different phenological stages. Sci. Hortic. 2014, 172, 161–167. [Google Scholar] [CrossRef]
- Salazar-Gutierrez, M.R.; Chaves, B.; Hoogenboom, G. Freezing tolerance of apple flower buds. Sci. Hortic. 2016, 198, 344–351. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group 1 to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, C., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2016; p. 1535. [Google Scholar]
- Djaman, K.; Koudahe, K.; Bodian, A.; Diop, L.; Ndiaye, P.M. Long-Term Trend Analysis in Annual and Seasonal Precipitation, Maximum and Minimum Temperatures in the Southwest United States. Climate 2020, 8, 142. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Sureshkumar, S.; Lempe, J.; Weigel, D. Potent induction of Arabidopsis thaliana flowering by elevated growth temperature. PLoS Genet. 2006, 2, 0980–0989. [Google Scholar] [CrossRef]
- Santos, J.A.; Costa, R.; Fraga, H. Climate change impacts on thermal growing conditions of main fruit species in Portugal. Clim. Chang. 2017, 140, 273–286. [Google Scholar] [CrossRef]
- Legave, J.M.; Guedon, Y.; Malagi, G.; El Yaacoubi, A.; Bonhomme, M. Differentiated responses of apple tree floral phenology to global warming in contrasting climatic regions front. Plant Sci. 2015, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Dai, J.; Ranjitkar, S.; Xu, J.; Luedeling, E. Response of chestnut phenology in China to climate variation and change. Agric. For. Meteorol. 2013, 180, 164–172. [Google Scholar] [CrossRef]
- Guédon, Y.; Legave, J.M. Analyzing the time-course variation of apple and pear tree dates of flowering stages in the global warming context. Ecol. Model. 2008, 219, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Legave, J.M.; Giovannini, D.; Christen, D.; Oger, R. Global warming in Europe and its impacts on floral bud phenology in fruit species. Acta Hortic. 2009, 838, 21–22. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Miranda, C.; Royo, J.B. Average dates and accumulated thermal requirements for different phenophases of peach as influenced by climate. In VII International Peach Symposium, Acta Horticulturae; Girona, J., Marsal, J., Eds.; International Society for Horticultural Science: Leuven, Belgium, 2012; Volume 962, pp. 277–284. [Google Scholar]
- Fraga, H.; Costa, R.; Moutinho-Pereira, J.; Correia, C.M.; Dinis, L.T.; Gonçalves, I.; Silvestre, J.; Eiras-Dias, J.; Malheiro, A.C.; Santos, J.A. Modeling phenology, water status, and yield components of three Portuguese grapevines using the STICS crop model. Am. J. Enol. Viticult. 2015, 66, 482–491. [Google Scholar] [CrossRef]
- Moriondo, M.; Ferrise, R.; Trombi, G.; Brilli, L.; Dibari, C.; Bindi, M. Modelling olive trees and grapevines in a changing climate. Environ. Model. Softw. 2015, 72, 387–401. [Google Scholar] [CrossRef]
- Park, Y.; Lee, B.; Park, H. Predicted effects of climate change on winter chill accumulation by temperate trees in South Korea. Hortic. J. Preview 2018, 87, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, C.J.; Brennan, R.M.; Jones, H.G. Declining chilling and its impact on temperate perennial crops. Environ. Exp. Bot. 2013, 91, 48–62. [Google Scholar] [CrossRef]
- Baldocchi, D.; Wong, S. Accumulated winter chill is decreasing in the fruit growing regions of California. Clim. Chang. 2008, 87, S153–S166. [Google Scholar] [CrossRef]
- Wang, S.; Huang, C.; Tao, J.; Zhong, M.; Qu, X.; Wu, H.; Xu, X. Evaluation of chilling requirement of kiwifruit (Actinidia spp.) in south China. N. Z. J. Crop Hortic. Sci. 2017, 45, 289–298. [Google Scholar] [CrossRef]
- Benmoussa, H.; Luedeling, E.; Ghrab, M.; Mimoun, M.B. Severe winter chill decline impacts Tunisian fruit and nut orchards. Clim. Chang. 2020, 162, 1249–1267. [Google Scholar] [CrossRef]
- Wang, X.; Li, D.; Zhang, D.; Shi, X.; Wu, Y.; Qi, Z.; Ding, H.; Zhu, K.; Xia, Y.; Zhang, J. Improving crucial details and selecting the optimal model for evaluating the chilling requirement of Paeonia lactiflora Pall. at low latitudes during four winters. Sci. Hortic. 2020, 265, 109175. [Google Scholar] [CrossRef]
- Fernandez, E.; Whitney, C.; Luedeling, E. The importance of chill model selection-a multi-site analysis. Eur. J. Agron. 2020, 119, 126103. [Google Scholar] [CrossRef]
- Bailey, C.H.; Kotowski, S.; Hough, L.F. Estimate of chilling requirements of apricot selections II. Acta Hortic. 1982, 121, 99–102. [Google Scholar] [CrossRef]
- Kuden, A.B.; Kasaka, M.; Tanriver, E. Determining the chilling requirements and growing degree hours of some pistachionut cultivars and regions. Acta Hortic. 1995, 419, 85–90. [Google Scholar] [CrossRef]
- Dokoozlian, N.K. Chilling temperature and duration interact on the budbreak of ‘perlette’ grapevine cutting. HortScience 1999, 34, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Egea, J.; Ortega, E.; Martínez–Gómez, P.; Dicenta, F. Chilling and heat requirements of almond cultivars for flowering. Environ. Exp. Bot. 2003, 50, 79–85. [Google Scholar] [CrossRef]
- Alono, J.M.; Anson, J.M.; Espiau, M.T.; Sociasi Company, R. Determination of endodormancy break in almond flower buds by a correlation model using the average temperature of different day intervals and its application to the estimation of chill and heat requirements and blooming date. J. Am. Soc. Hortic. Sci. 2005, 130, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Okie, W.R.; Hancock, J.F. Plums. Temperate Fruit Crop Breeding; Hancock, J.F., Ed.; Springer: Amsterdam, The Netherlands, 2008; pp. 337–357. [Google Scholar]
- Rahemi, M.; Pakkish, Z. Determination of chilling and heat requirements of pistachio (Pistacia vera L.) cultivars. Agric. Sci. China 2009, 8, 803–807. [Google Scholar] [CrossRef]
- Schalau, J. Deciduous Fruit Tree Pruning. University of Arizona Cooperative Extension. 2009. Available online: https://cals.arizona.edu/yavapai/anr/hort/byg/archive/fruittreepruning2009.html (accessed on 15 June 2020).
- Chaar, J.; Astorga, D. Determination of the requirement of cold and heat in peach [Prunus persica (L.) Batsch.] through a correlation model. RIA Rev. Investig. Agropecu. 2012, 38, 289–298. [Google Scholar]
- Campoy, J.A.; Ruiz, D.; Allderman, L.; Cook, N.; Egea, J. The fulfillment of chilling requirements and the adaptation of apricot (Prunus armeniaca L.) in warm winter climates: An approach in Murcia (Spain) and the Western Cape (South Africa). Eur. J. Agron. 2012, 37, 43–55. [Google Scholar] [CrossRef]
- Prudencio, A.S.; Martínez-Gómez, P.; Dicenta, F. Evaluation of breaking dormancy, flowering and productivity of extra-late and ultra-late flowering almond cultivars during cold and warm seasons in south-east of Spain. Sci. Hortic. 2018, 235, 39–46. [Google Scholar] [CrossRef]
- Ramirez, L.; Sagredo, G.H.; Reginato, G.H. Prediction models for chilling and heat requirement to estimate full bloom of almond cultivars in the Central Valley of Chile. Acta Hortic. 2020, 872, 107–112. [Google Scholar] [CrossRef]
- Elloumi, O.; Ghrab, M.; Mimoun, M.B. Chilling accumulation effects on performance of pistachio trees cv. Mateur in dry and warm area climate. Sci. Hortic. 2013, 159, 80–87. [Google Scholar] [CrossRef]
- Ikinci, A.; Mamay, M.; Unlu, L.; Bolat, I.; Ercisli, S. Determination of heat requirements and effective heat summations of some pomegranate cultivars grown in Southern Anatolia. Erwerbs-Obstbau 2014, 56, 131–138. [Google Scholar] [CrossRef]
- Scott, C. Chill Requirement of Pecans. 2015. Available online: http://www.pecangrowers.org.au/here/wp-content/uploads/Pecan-Growing-Chill-Requirement-of-Pecans.pdf (accessed on 12 October 2020).
- Funes, I.; Aranda, X.; Biel, C.; Carbo, J.; Camps, F.; Molina, A.J.; Herralde, F.D.; Grau, B.R. Save Future climate change impacts on apple flowering date in a Mediterranean subbasin. Agric. Water Manag. 2016, 164, 19–27. [Google Scholar] [CrossRef]
- Yaacoubi, A.E.; Malagi, G.; Oukabli, A.; Citadin, I.; Hafidi, M.; Bonhomme, M.; Legave, J.M. Differentiated dynamics of bud dormancy and growth in temperate fruittrees relating to bud phenology adaption, the case of apple and almond trees. Int. J. Biometeorol. 2016, 60, 1695–1710. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Cai, B.; Gao, Z.; Zhang, Z. Determination of chilling and heat requirements of 69 Japanese apricot cultivars. Eur. J. Agron. 2016, 74, 68–74. [Google Scholar] [CrossRef]
- Measham, P.F.; Darbyshire, R.; Turpin, S.R.; Murphy White, S. Complexity in chill calculations: A case study in cherries. Sci. Hortic. 2017, 216, 134–140. [Google Scholar] [CrossRef]
- Thompson, C. Georgia’s Peach Farmers Hoping for Colder Winter This Year. 2017. Available online: https://newswire.caes.uga.edu/story.html?storyid=6416secure.caes.uga.edu/extension/publications/files/pdf/B%201518_2.PDF (accessed on 21 June 2020).
- Montazeran, A.; Khadivi, A.; Khaleghi, A. The first report: Chilling and heat requirements of seedless barberry (Berberis vulgaris L. var. asperma). Sci. Hortic. 2018, 231, 188–193. [Google Scholar] [CrossRef]
- Gaeta, L.; Stellacci, A.M.; Losciale, P. Evaluation of three modelling approaches for almond blooming in Mediterranean climate conditions. Eur. J. Agron. 2018, 97, 1–10. [Google Scholar] [CrossRef]
- Chavez, D.; Florkowska, M.; Chen, C.; Cook, J. Home Garden Plums. University of Georgia Extension Bulletin 1518. 2019. Available online: https://secure.caes.uga.edu/extension/publications/files/pdf/B%201518_2.PDF (accessed on 12 October 2020).
- Díez-Palet, I.; Funes, I.; Savé, R.; Biel, C.; de Herralde, F.; Miarnau, X.; Vargas, F.; Àvila, G.; Carbó, J.; Aranda, X. Blooming under mediterranean climate: Estimating cultivar-specific chill and heat requirements of almond and apple trees using a statistical approach. Agronomy 2019, 9, 760. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, H.; Blanke, M. Substitution of winter chilling by spring forcing for flowering using sweet cherry as model crop. Sci. Hortic. 2019, 244, 75–81. [Google Scholar] [CrossRef]
- Parkes, H.; Darbyshire, R.; White, N. Chilling requirements of apple cultivars grown in mild Australian winter conditions. Sci. Hortic. 2020, 260, 108858. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Z.F.; Guo, L.; He, H.S.; Ling, Y.H.; Wang, L.; Zong, S.W.; Na, R.; Du, H.B.; Li, M.H. Effects of winter chilling vs. spring forcing on the spring phenology of trees in a cold region and a warmer reference region. Sci. Total Environ. 2020, 725, 138323. [Google Scholar] [CrossRef]
- Nasrabadi, M.; Ramezanian, A.; Eshghi, S.; Sarkhosh, A. Chilling and heat requirement of pomegranate (Punica granatum L.) trees grown under sustained deficit irrigation. Sci. Hortic. 2020, 263, 109–117. [Google Scholar] [CrossRef]
- Camargo-Alvarez, H.; Salazar-gutiérrez, M.; Keller, M.; Hoogenboom, G. Modeling the effect of temperature on bud dormancy of grapevines. Agric. For. Meteorol. 2020, 280, 107782. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Chill Unit Accumulated |
|---|---|
| <1.4 | 0.0 |
| 1.5–2.4 | 0.5 |
| 2.5–9.1 | 1.0 |
| 9.2–12.4 | 0.5 |
| 12.5–15.9 | 0.0 |
| 16–18 | −0.5 |
| >18 | −1.0 |
| Months | Fabian Garcia | Los Lunas | Farmington | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abs. Tmin | Ave. Tmin | Abs. Tmax | Ave. Tmax | Abs. Tmin | Ave. Tmin | Abs. Tmax | Ave. Tmax | Abs. Tmin | Ave. Tmin | Abs. Tmax | Ave. Tmax | |
| January | −7.7 | −0.7 ± 1.0 | 24.0 | 13.1 ± 1.9 | −15.2 | −3.7 ± 1.1 | 21.1 | 9.2 ± 2.0 | −19.4 | −5.7 ± 1.8 | 15.9 | 5.2 ± 1.8 |
| February | −13.9 | 1.2 ± 1.4 | 27.1 | 16.2 ± 1.8 | −18.4 | −2.3 ± 1.1 | 24.1 | 12.3 ± 1.9 | −19.3 | −4.1 ± 1.3 | 20.7 | 7.9 ± 1.8 |
| March | −7.4 | 4.3 ± 1.5 | 30.9 | 20.6 ± 1.7 | −7.7 | 0.4 ± 1.3 | 27.6 | 17.0 ± 1.7 | −11.5 | −1.4 ± 1.2 | 25.5 | 12.9 ± 1.8 |
| April | −2.7 | 8.1 ± 1.5 | 35.3 | 25.4 ± 1.6 | −5.4 | 3.9 ± 1.4 | 31.8 | 21.6 ± 1.7 | −7.4 | 1.7 ± 1.4 | 28.5 | 17.6 ± 1.6 |
| May | 3.1 | 13.2 ± 1.1 | 39.3 | 30.4 ± 1.6 | −1.4 | 9.2 ± 1.3 | 37.9 | 26.8 ± 1.9 | −4.0 | 6.9 ± 1.5 | 36.2 | 23.3 ± 2.0 |
| June | 10.7 | 18.9 ± 1.3 | 43.4 | 35.5 ± 1.6 | 5.4 | 14.9 ± 1.3 | 40.5 | 32.8 ± 1.6 | 2.3 | 12.8 ± 1.4 | 37.9 | 30.1 ± 1.7 |
| July | 14.9 | 21.1 ± 1.0 | 42.5 | 34.5 ± 1.6 | 10.6 | 18.1 ± 1.0 | 40.3 | 32.7 ± 1.5 | 7.9 | 16.5 ± 1.0 | 39 | 31.7 ± 1.5 |
| August | 14.1 | 20.2 ± 1.1 | 40.8 | 33.3 ± 1.7 | 9.8 | 17.1 ± 1.1 | 37.6 | 31.3 ± 1.6 | 5.9 | 15.5 ± 1.1 | 37.0 | 29.9 ± 1.6 |
| September | 6.7 | 16.5 ± 1.4 | 37.9 | 30.5 ± 1.7 | 0.7 | 12.8 ± 1.4 | 36.2 | 28.1 ± 1.5 | −1.1 | 10.8 ± 1.4 | 35.4 | 26.0 ± 1.5 |
| October | −3.5 | 10.2 ± 1.3 | 35.1 | 25.4 ± 1.5 | −7.1 | 6.1 ± 1.4 | 31.9 | 22.1 ± 1.6 | −9.8 | 4.1 ± 1.5 | 29.7 | 19.1 ± 1.7 |
| November | −7.0 | 3.6 ± 1.7 | 28.7 | 18.2 ± 1.9 | −8.4 | −0.2 ± 1.5 | 24.7 | 14.6 ± 2.0 | −12.5 | −1.9 ± 1.4 | 22.6 | 11.2 ± 2.2 |
| December | −8.3 | −0.5 ± 1.1 | 24.3 | 12.8 ± 1.5 | −17.6 | −3.6 ± 1.1 | 20.3 | 8.9 ± 1.6 | −23.2 | −5.5 ± 1.4 | 17 | 5.1 ± 1.5 |
| Sites | Fruit Tree Growing Seasons | Endodormancy Period | Ecodormancy Period | Chill Portions | Chilling Hours | Accumulated Heat (GDH) |
|---|---|---|---|---|---|---|
| Fabian Garcia | 2015–2016 | 20 October–31 December | 1 January–10 March | 33.8 | 689.06 | 8855.81 |
| 2016–2017 | 6 November–30 December | 31 December–27 February | 0 | 660.84 | 6947.58 | |
| 2017–2018 | 21 October–31 December | 1 January–18 March | 1.6 | 617.82 | 11,352.27 | |
| 2018–2019 | 24 October–31 December | 1 January–8 March | 5.9 | 791.1 | 7817.70 | |
| 2019–2020 | 18 October–31 December | 1 January–14 March | 3 | 820.64 | 8702.91 | |
| Los Lunas | 2017–2018 | 6 October–01 January | 2 January–8 April | 36.4 | 686.08 | 10,748.86 |
| 2018–2019 | 5 October–03 January | 4 January–7 April | 45.7 | 733.26 | 7845.97 | |
| 2019–2020 | 4 October–01 January | 2 January–1 April | 44.6 | 760.24 | 7544.88 | |
| Farmington | 2015–2016 | 7 October–17 January | 18 January–13 April | 53.5 | 750.50 | 6316.67 |
| 2016–2017 | 26 October–29 January | 30 January–29 March | 57.9 | 781.46 | 4475.90 | |
| 2017–2018 | 24 October–11 January | 12 January–28 April | 57.9 | 964.04 | 9894.20 | |
| 2018–2019 | 1 October–6 January | 7 January–9 April | 55.1 | 798.50 | 4007.80 | |
| 2019–2020 | 5 October–6 January | 7 January–15 April | 55.7 | 850.24 | 5228.87 |
| References | Locations | Tree Species | Chill Portions | Chilling Hours | Heat Requirement (GDH) |
|---|---|---|---|---|---|
| Bailey et al. [88] | New Jersey, USA | Apricots | 873–1343 | ||
| Anderson et al. [46] | Sour cherry | 954 | 6130 | ||
| Linvill [30] | South Carolina, USA | 65 | |||
| Kuden et al. [89] | California, USA | Pistacho | 600–1050 | ||
| Dokoozlian [90] | California, USA | 50–400 | |||
| Egea et al. [91] | Santomera, Spain | Almond | 266–996 | 5942–7577 | |
| Alonso et al. [92] | Spain | Almond | 400–600 | 5500–9300 | |
| Ruiz et al. [34] | Calasparra, Spain | Apricots | 596–1266 | 4078–5879 | |
| Okie et al. [93] | Plums | 450 | |||
| Alburquerque et al. [49] | Murcia, Spain | Sweet cherry | 30.4–57.6 | 335 ± 38–1323 ± 68 | 7326–9450 |
| Luedeling et al. [40] | Walnuts | 53.3–79.5 | 700 | ||
| Rahemi and Pakkish [94] | Iran | Pistachio | 750–1400 | 8 852–15 420 | |
| Schalau [95] | Yavapai, Arizona | Fruit trees | 500–1000 | ||
| Chaar and Astorga [96] | Junín, Argentina | Peach | 2177–6490 | ||
| Luedeling [16] | Klein-Altendorf, Germany | Cherry | 68.6 ± 5.7 | 1375 ± 178 | 3473 ± 1236 |
| Campoy et al. [97] | Western Cape, SouthAfrica | Apricot | 26.6–57.2 | 312–1022 | |
| Campoy et al. [97] | Murcia, Spain | Apricot | 31–51.8 | 4605–6247 | |
| Luedeling et al. [21] | Beijing, China | Chestnu | 79.8 ± 5.3 | 13466 ± 1918 | |
| Luedeling et al. [21] | Klein-Altendorf, Germany | Cherry | 104.2 ± 8.9 | 2698 ± 1183 | |
| Prudencio et al. [98] | Spain | Almonds | 49–66 | 308–843 | 32,225–36,087 |
| Raminrez et al. [99] | Chile | Almonds | 23–32 | 220–440 | 5814 ± 669–12,341 ± 637 |
| Elloumi et al. [100] | Tunisia | Pistachio | 36 | 206 | |
| Ikinci et al. [101] | Tuekey | Pomegranate | 25,000–88,052 | ||
| Guo et al. [19] | Beijing, China | Jujube | 89 ± 6 | 13,619 ± 2033 | |
| Guo et al. [19] | Beijing, China | Chestnut | 93 ± 6 | 17,418 ± 1983 | |
| Scott Clark [102] | Pecans | 17–83 | 200–1000 | ||
| Funes et al. [103] | Girona, Spain | Apple | 62.5–68.4 | 7416.2 ± 687–10,272.5 ± 1032 | |
| Yaacoubi et al. [104] | Palmas, Brazil | Apple | 20.3–30.8 | 6893 | |
| Yaacoubi et al. [104] | Marsillargues, France | Apple | 39.1–70.8 | 9443 | |
| Yaacoubi et al. [104] | Ain Taoujdate, Morocco | Apple | 64.2–67.2 | 5985 | |
| Yaacoubi et al. [104] | Ain Taoujdate, Morocco | Almonds | 12.4–16.4 | ||
| Zhuang et al. [105] | Nanjing, China | Japanese apricot | 24–82 | 691.9–2634.7 | |
| Benmoussa et al. [51] | SfaxEl-Maou, Tunisia | Local almond | 3.4–15.5 | 3962–8873 | |
| Benmoussa et al. [51] | SfaxEl-Maou, Tunisia | Foreign almond | 6.7–22.6 | 2894–10,504 | |
| Measham et al. [106] | Western Australia | Cherries | 30.4–61.7 | ||
| Thomppson [107] | Georgia, USA | Peach | 800 | ||
| Montazeran et al. [108] | Iran | Barberry | 1400 | 2904–3432 | |
| Gaeta et al. [109] | Italy | Almond | 24–62 | 3263–6699 | |
| Chavez et al. [110] | Georgia, USA | Japanese plums | 500–900 | ||
| Chavez et al. [110] | Georgia, USA | Europian plums | 700–1000 | ||
| Díez-Palet et al. [111] | Egypt | Almond | 8.40 ± 3.8–52.85 ± 6 | 6232 ± 1221–10,201 ± 1834 | |
| Díez-Palet et al. [111] | Egypt | Apple | 37.8 ± 2.7–54.4 ± 3 | 7471 ± 1191–9501 ± 1556 | |
| Kaufmann and Blanke [112] | Sweet cherries | 400–750 | 4000–13,000 | ||
| Parkes et al. [113] | Australia | Apple | 57 ± 2.9–77 ± 1.5 | 662 ± 44.5–908 ± 23.3 | |
| Yang et al. [114] | Heilongjiang province, China | Ulmus pumila | 86 | 5853 | |
| Yang et al. [114] | Heilongjiang province, China | Populus simonii | 86 | 5853 | |
| Yang et al. [114] | Heilongjiang province, China | Syringa oblata | 86 | 5853 | |
| Nasrabadi et al. [115] | Iran | Pomegranate | 605.56–700 | 7750–9000 | |
| Kwon et al. [50] | Republic of Korea | Peach | 21.3–74.8 | 377–1134 | 4824–5149 |
| Camargo-Alvarez et al. [116] | Washington State, USA | Grapevines | 947–1162 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djaman, K.; Koudahe, K.; Darapuneni, M.; Irmak, S. Chilling and Heat Accumulation of Fruit and Nut Trees and Flower Bud Vulnerability to Early Spring Low Temperatures in New Mexico: Meteorological Approach. Sustainability 2021, 13, 2524. https://doi.org/10.3390/su13052524
Djaman K, Koudahe K, Darapuneni M, Irmak S. Chilling and Heat Accumulation of Fruit and Nut Trees and Flower Bud Vulnerability to Early Spring Low Temperatures in New Mexico: Meteorological Approach. Sustainability. 2021; 13(5):2524. https://doi.org/10.3390/su13052524
Chicago/Turabian StyleDjaman, Koffi, Komlan Koudahe, Murali Darapuneni, and Suat Irmak. 2021. "Chilling and Heat Accumulation of Fruit and Nut Trees and Flower Bud Vulnerability to Early Spring Low Temperatures in New Mexico: Meteorological Approach" Sustainability 13, no. 5: 2524. https://doi.org/10.3390/su13052524
APA StyleDjaman, K., Koudahe, K., Darapuneni, M., & Irmak, S. (2021). Chilling and Heat Accumulation of Fruit and Nut Trees and Flower Bud Vulnerability to Early Spring Low Temperatures in New Mexico: Meteorological Approach. Sustainability, 13(5), 2524. https://doi.org/10.3390/su13052524