
Back-Calculation of Fish Size in Diet Analysis of Piscivorous Predators: A New Index for the Alien Silurus glanis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Laboratory Collection of S. glanis
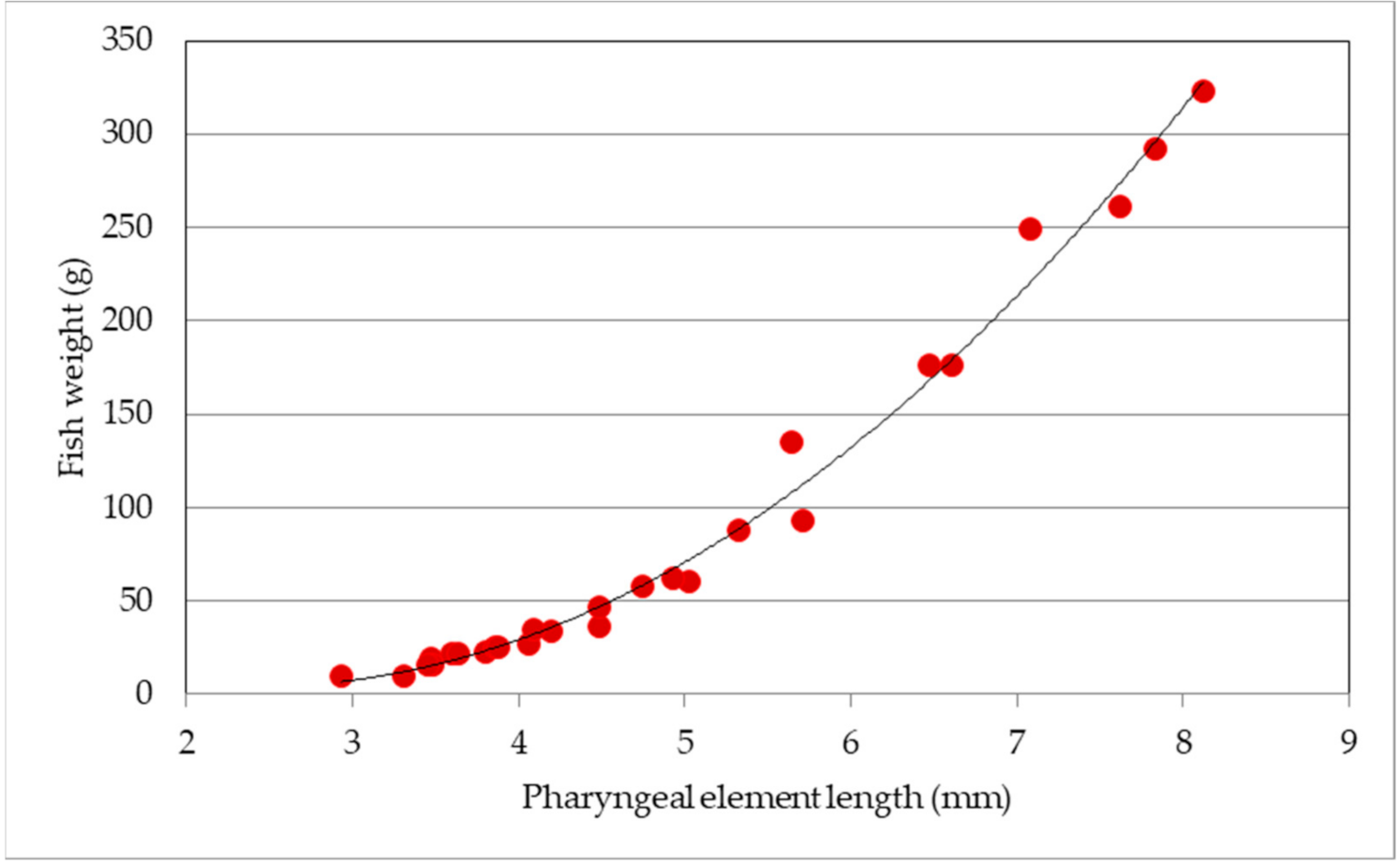
2.2. Bone Measurements for Back-Calculations
2.3. Ethical Statement
2.4. Statistical Analyses
3. Results
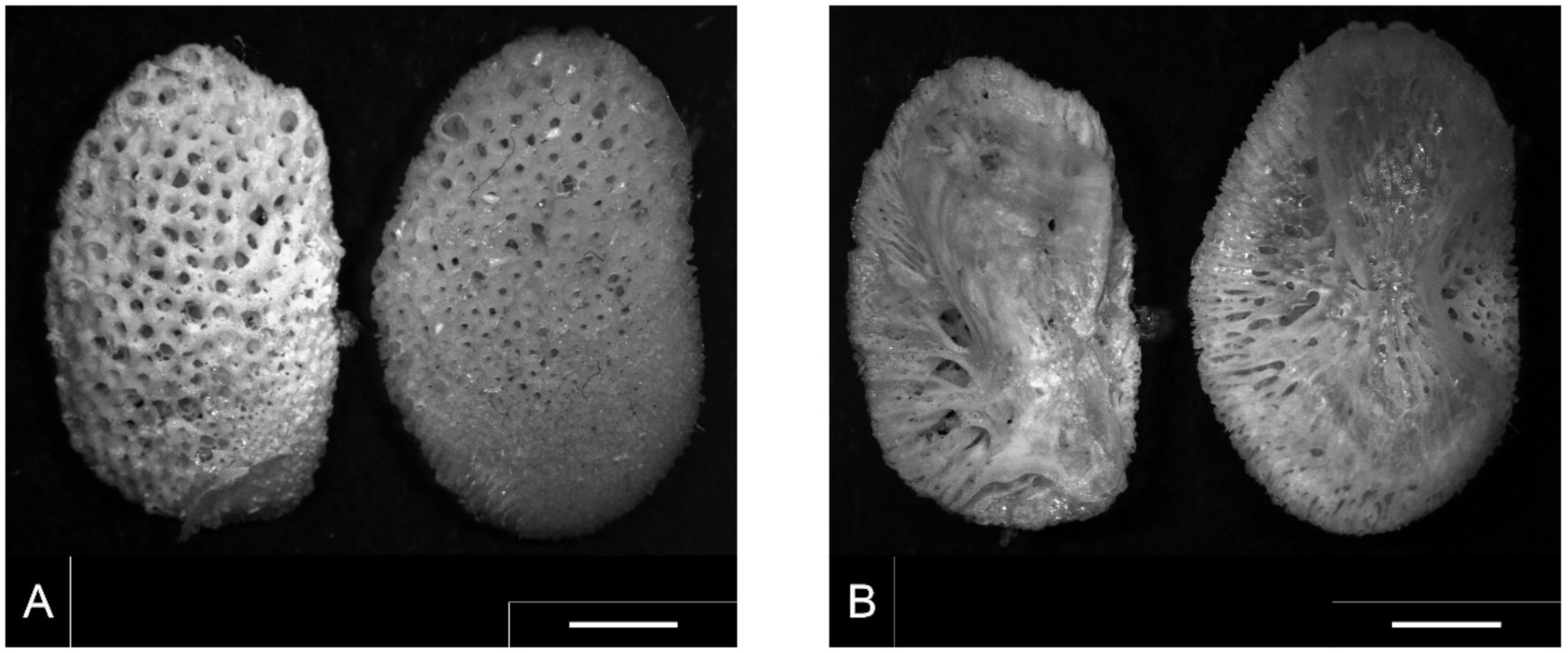
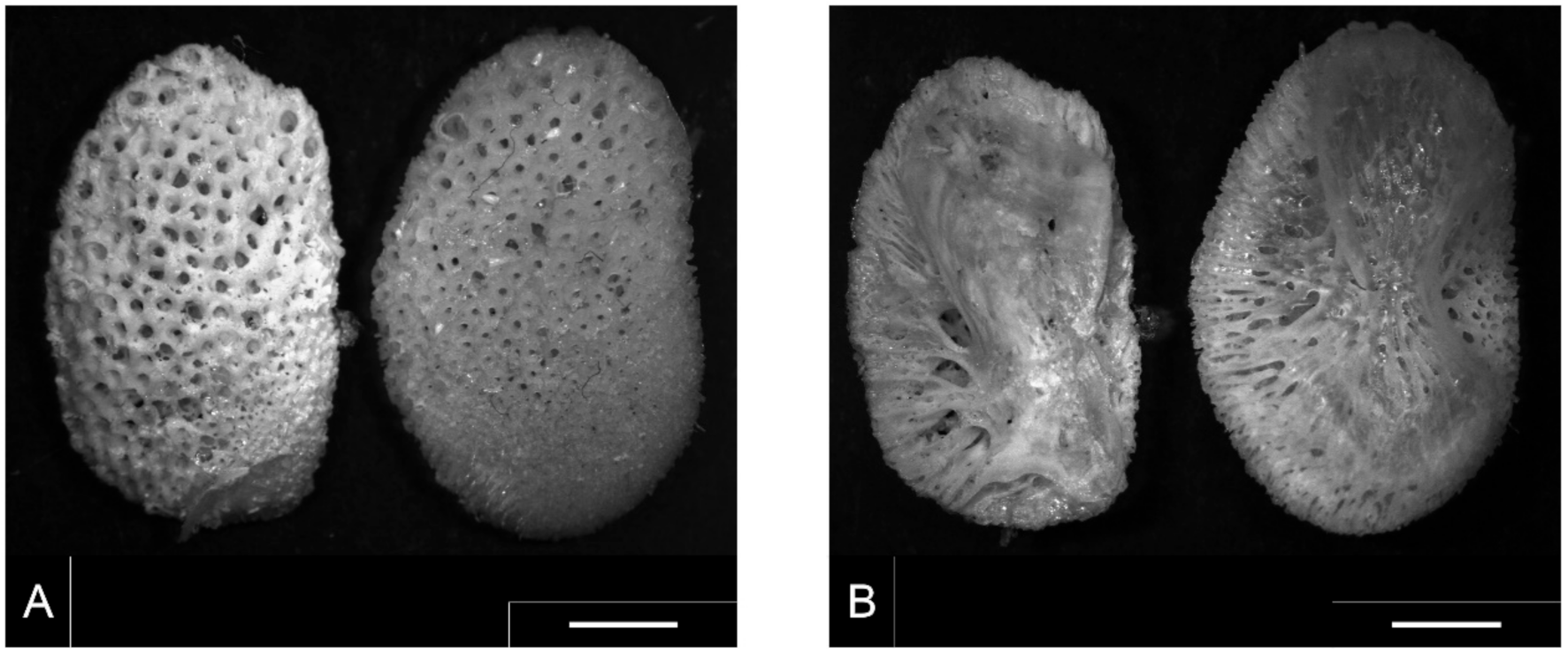
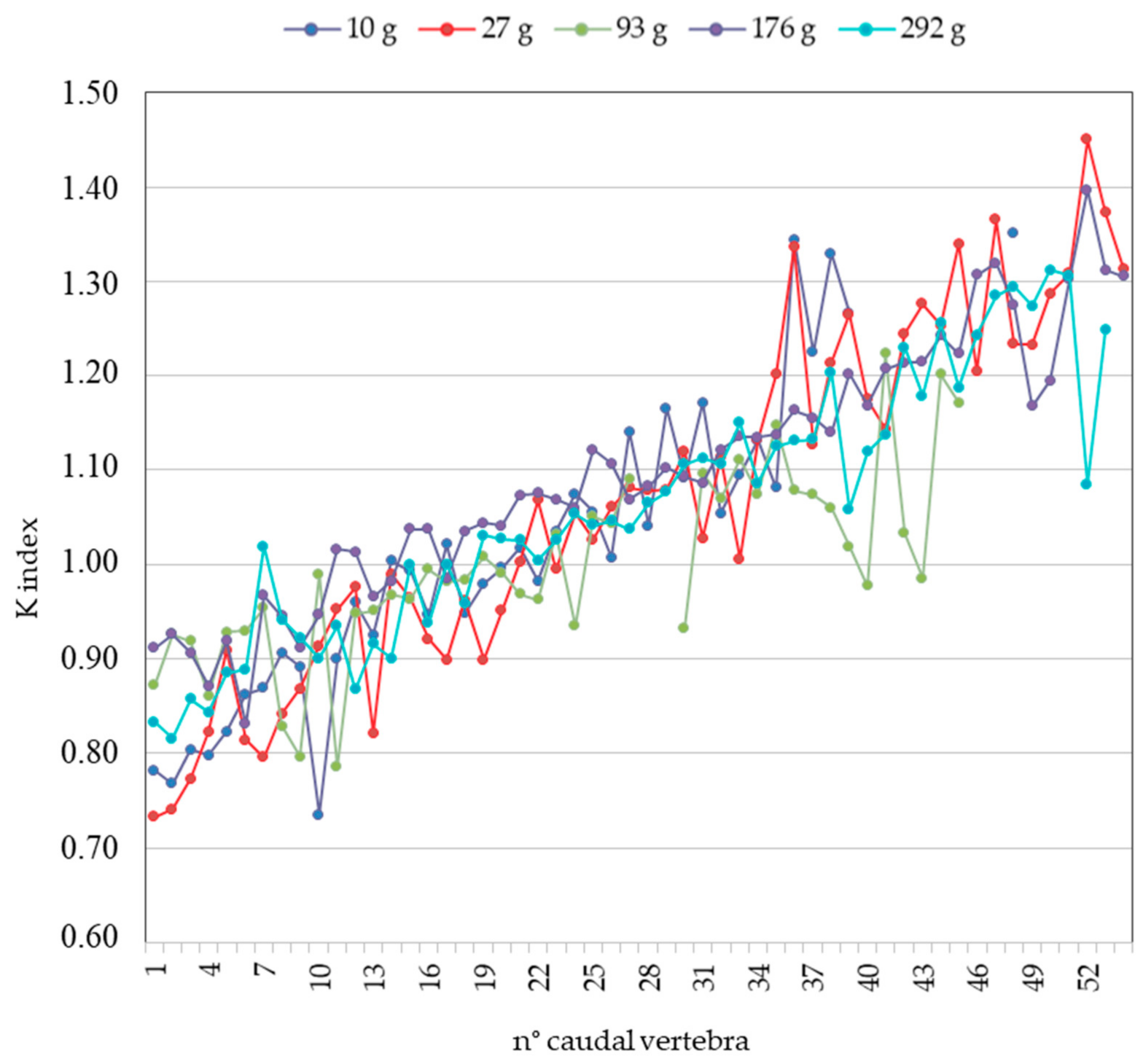
3.1. S. glanis Identification by Pharyngeal Element
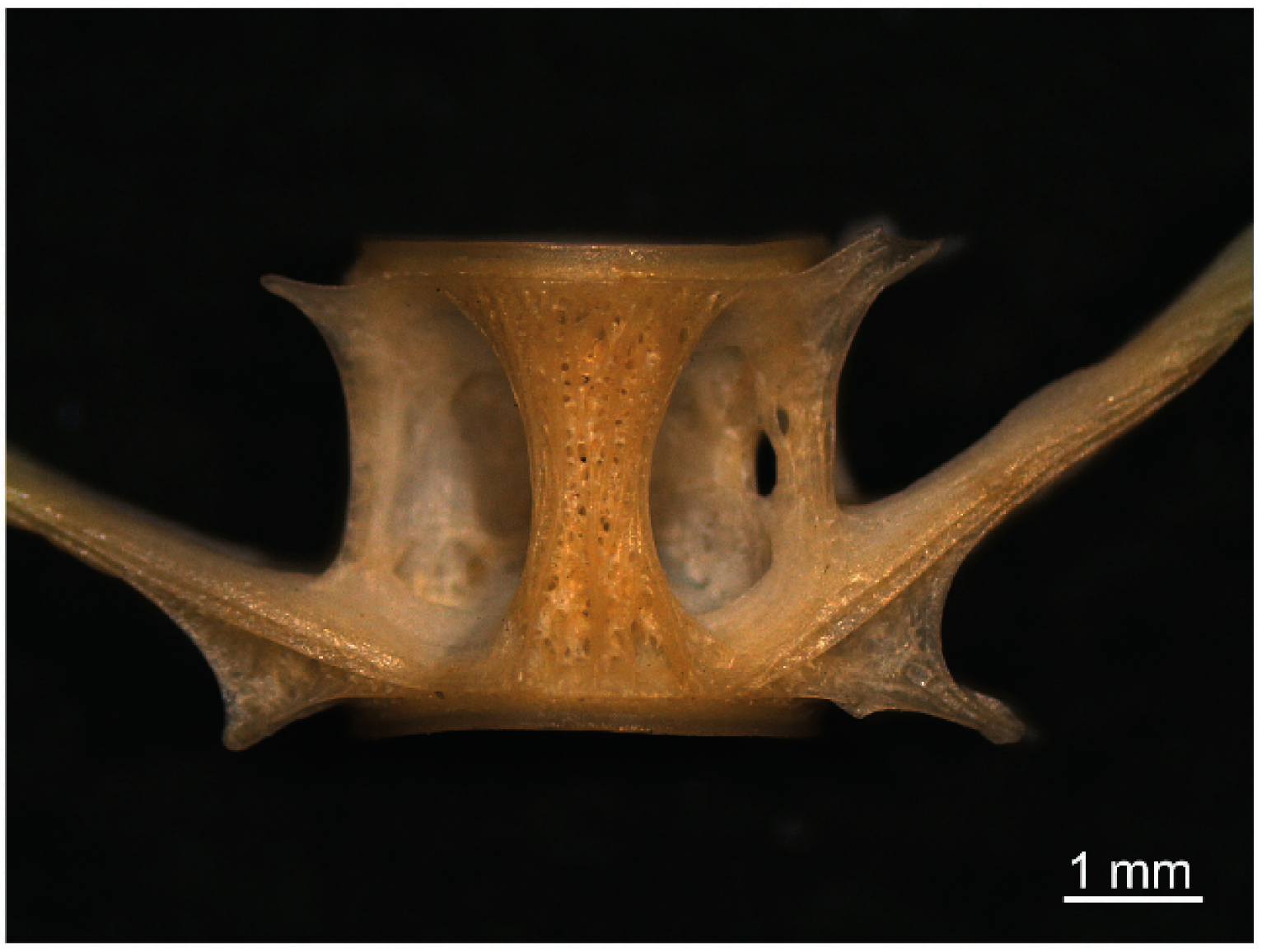
3.2. S. glanis Identification by Vertebra
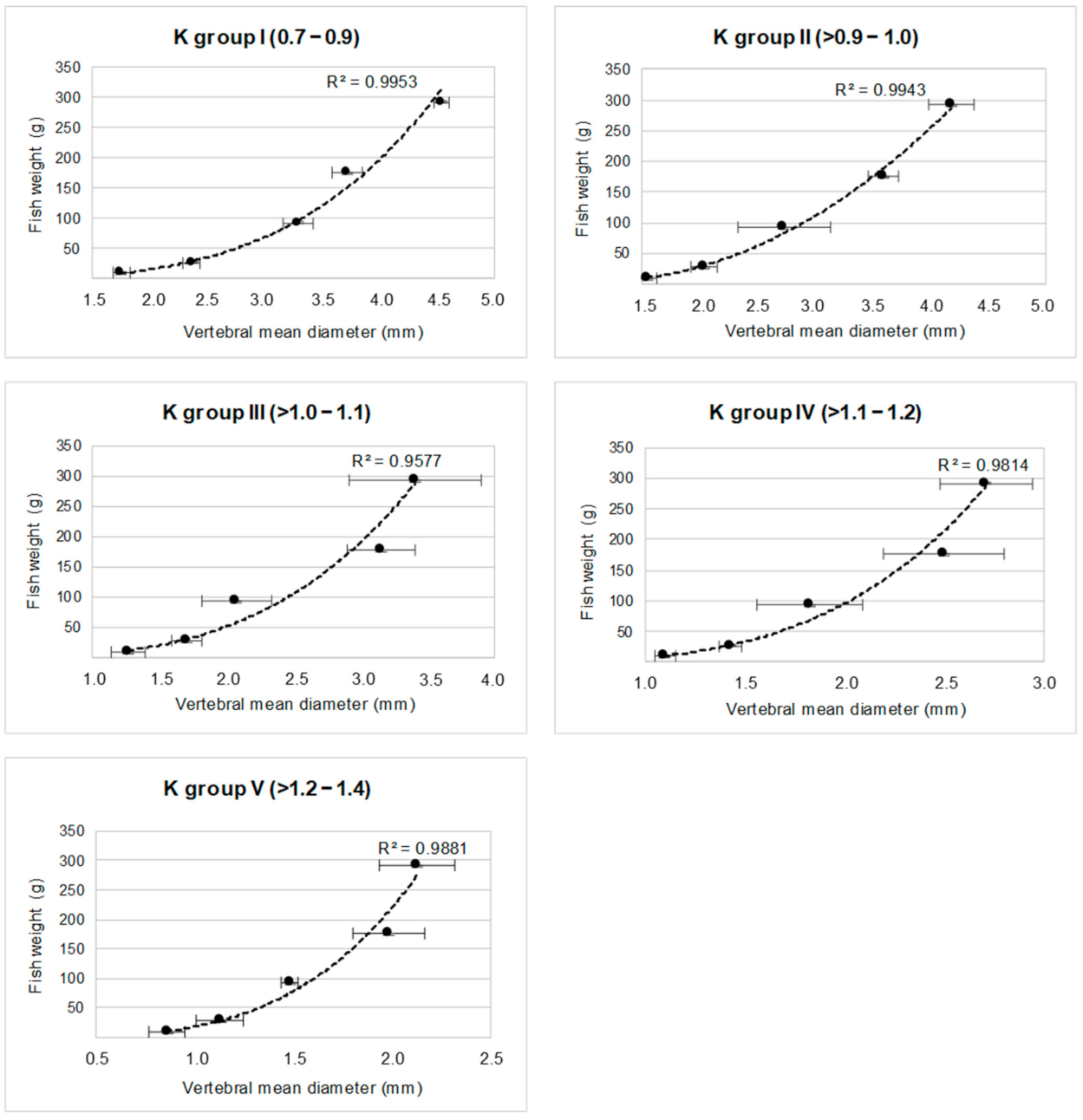
3.3. Model Validation
3.4. Back-Calculation of the S. glanis Size in Otter’s Stools
4. Discussion
5. Conclusions
- The alveolar structure of the hourglass rib on the vertebral body of S. glanis is a diagnostic element for species identification, as are the shape and structure of the pharyngeal dental plate.
- The proposed K-index is able to identify the vertebral position along the column, independently from the fish size, and the set of predictive equations developed and validated in this work can establish the size of the preyed fish with a good level of accuracy.
- The back-calculation of the original size of preyed fish open new perspectives for more detailed studies on the diet of piscivorous predators as well as on control efficacy by the predation to prevent uncontrolled growth of allochthonous species.
- The proposed methodology applied to S. glanis remains in otter stools from the Ticino River revealed the predation of 10 specimens with weights ranging between 5 and 550 g, lengths between 85 and 435 mm, and ages between 0 and 2 years, with more frequent predation on specimens about 1 year old and 250–350 mm in length.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stone, R. The last of the leviathans. Science 2007, 316, 1684–1688. [Google Scholar] [CrossRef]
- Copp, G.H.; Bianco, P.G.; Bogutskaya, N.G.; Eros, T.; Falka, I.; Ferreira, M.T.; Fox, M.G.; Freyhof, J.; Gozlan, R.E.; Grabowska, J.; et al. To be, or not to be, a non-native freshwater fish? J. Appl. Ichthyol. 2005, 21, 242–262. [Google Scholar] [CrossRef]
- Hickley, P.; Chare, S. Fisheries for non-native species in England and Wales: Angling or the environment? Fish. Manag. Ecol. 2004, 11, 203–212. [Google Scholar] [CrossRef]
- Wysujack, K.; Mehner, T. Can feeding of European catfish prevent cyprinids from reaching a size refuge? Ecol. Freshw. Fish 2005, 14, 87–95. [Google Scholar] [CrossRef]
- Valadou, B. Le Silure Glane (Silurus glanis, L.) en France. Évolution de son Aire de Répartition et Prédiction de son Extension; Conseil Supérieur de la Pêche: Fontenay-sous-Bois, France, 2007. [Google Scholar]
- Copp, G.H.; Britton, R.J.; Cucherousset, J.; García-Berthou, E.; Kirk, R.; Peeler, E.; Stakenas, S. Voracious invader or benign feline? A review of the environmental biology of European catfish Silurus glanis in its native and introduced ranges. Fish Fish. 2009, 10, 252–282. [Google Scholar] [CrossRef]
- Prenda, J.; Clavero, M.; Blanco-Garrido, F.; Menor, A.; Hermoso, V. Threats to the conservation of biotic integrity in Iberian fluvial ecosystems. Limnetica 2006, 25, 377–388. [Google Scholar]
- Miranda, R.; Copp, G.H.; Williams, J.; Beyer, K.; Gozlan, R.E. Do Eurasian otters Lutra lutra (L.) in the Somerset Levels prey preferentially on non-native fish species? Fundam. Appl. Limnol. 2008, 266, 255–260. [Google Scholar] [CrossRef]
- Smiroldo, G.; Balestrieri, A.; Pini, E.; Tremolada, P. Anthropogenically altered trophic webs: Alien catfish and microplastics in the diet of Eurasian otters. Mammal Res. 2019, 64, 165–174. [Google Scholar] [CrossRef]
- Carss, D.N.; Kruuk, H.; Conroy, J.W.H. Predation on adult Atlantic salmon, Salmo salar L. by otters, Lutra lutra (L.) within the River Dee system, Aberdeenshire, Scotland. J. Fish Biol. 1990, 37, 935–944. [Google Scholar] [CrossRef]
- Kruuk, H.; Carss, D.N.; Conroy, J.W.H.; Durbin, L. Otter Lutra lutra numbers and fish productivity in rivers of North East Scotland. Symp. Zool. Soc. Lond. 1993, 65, 9–13. [Google Scholar]
- Weinberger, I.C.; Muff, S.; de Jongh, A.; Kranz, A.; Bontadina, F. Flexible habitat selection paves the way for a recovery of otter populations in the European Alps. Biol. Conserv. 2016, 199, 88–95. [Google Scholar] [CrossRef]
- Copp, G.H.; Roche, K. Range and diet of Eurasian otters Lutra lutra (L.) in the catchment of the River Lee (south-east England) since re-introduction. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, 65–76. [Google Scholar] [CrossRef]
- Rutkayová, J.; Biskup, R.; Harant, R.; Šlechta, V.; Koščo, J. Ameiurus melas (black bullhead): Morphological characteristics of new introduced species and its comparison with Ameiurus nebulosus (brown bullhead). Rev. Fish Biol. Fish. 2013, 23, 51–68. [Google Scholar] [CrossRef]
- Rojo, A. Osteological Atlas of the Brown Bullhead (Ameiurus nebulosus) from Nova Scotia Waters: A Morphological and Biometric Study; Curatorial Report Number 100; Nova Scotia Museum: Halifax, NS, Canada, 2013. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Desse, G.; Du Buit, M.-H. Diagnostic des Pièces Rachidiennes des Téléostéens et des Chondrichthyens. III Téléostéens d’eau Douce; Expansion Scientifique Française: Paris, France, 1971. [Google Scholar]
- Moyle, P.; Nichols, R.D. Ecology of Some Native and Introduced Fishes of the Sierra Nevada Foothills in Central California. Copeia 1973, 3, 478–490. [Google Scholar] [CrossRef]
- Puzzi, C.M.; Trasforini, S.; Casoni, A.; Bardazzi, M.A.; Bellani, A. Il siluro (Silurus glanis). Ecologia della Specie nel Fiume Ticino e Risultati Dell’azione di Contrasto alla Sua Espansione Svolta dal Parco Negli Anni 2001–2006; Consorzio del Parco Lombardo della Valle del Ticino; Pontevecchio di Magenta: Milan, Italy, 2007. [Google Scholar]
- Pierce, G.J.; Boyle, P.R. A review of methods for diet analysis in piscivorous marine mammals. Oceanogr. Mar. Biol. Ann. Rev. 1991, 29, 409–486. [Google Scholar]
- Pierce, G.J.; Boyle, P.R.; Watt, J.; Grisley, M. Recent advances in diet analysis of marine mammals. Symp. Zool. Soc. Lond. 1993, 66, 241–261. [Google Scholar]
- Granadeiro, J.P.; Silva, M.A. The use of otoliths and vertebrae in the identification and size-estimation of fish in predator-prey studies. Cybium 2000, 24, 383–393. [Google Scholar]
- Feltham, M.J.; Marquiss, M. The use of first vertebrae in separating, and estimating the size of trout (Salmo trutta) and salmon (Salmo salar) in bone remains. J. Zool. 1989, 219, 113–122. [Google Scholar] [CrossRef]
- Feltham, M.J. The diet of red-breasted mergansers (Mergus serrator) during the smolt run in N.E. Scotland: The importance of salmon (Salmo salar) smolts and parr. J. Zool. 1990, 222, 285–292. [Google Scholar] [CrossRef]
- Desse, J.; Desse-Berset, N. Archaeozoology of groupers (Epinephelinae) identification, osteometry and keys to interpretation. Archaeofauna 1996, 5, 121–127. [Google Scholar]
- Conroy, J.W.H.; Watt, J.; Webb, J.B.; Jones, A. A Guide to the Identification of Prey Remains in Otter Spraints, 3rd ed.; The Mammal Society: London, UK, 2005. [Google Scholar]
- Hermsen, J.; Maarseveen, A.V. A Diet Study of the Eurasian Otter (Lutra lutra) Based on Spraint Analysis; Niewold Wildlife Infocentre: Leeuwarden, The Netherlands, 2011. [Google Scholar]
- Thom, T.J. The Ecology of Otters (Lutra lutra) on the Wansbeck and Blyth River Catchements in Northumberland; Durham E-Theses; Durham University: Durham, UK, 1990. [Google Scholar]
- Hensel, K. Find of a capital wels catfish Silurus glanis (Actinopterygii: Siluridae) in excavations of a Roman military fort in Southern Slovakia. Biologia 2004, 15, 191–203. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Species | IUCN Red List | Ab. |
|---|---|---|---|---|
| Acipenseriformes | Acipenseridae | Acipenser naccarii† | VU | 1 |
| Anguilliformes | Anguillidae | Anguilla anguilla | CR | 2 |
| Cypriniformes | Cobitidae | Cobitis taenia | LC | 3 |
| Misgurnus anguillicaudatus | - | 3 | ||
| Sabanejewia larvata | LC | 1 | ||
| Cyprinidae | Abramis brama | - | 1 | |
| Alburnus alburnus | DD | 4 | ||
| Aspius aspius | - | 1 | ||
| Barbus caninus | EN | 2 | ||
| Barbus plebejus | LC | 4 | ||
| Blicca bjoerkna | - | 3 | ||
| Carassius carassius | - | 3 | ||
| Chondrostoma soetta† | EN | 1 | ||
| Ctenopharyngodon idella | - | 1 | ||
| Cyprinus carpio | - | 2 | ||
| Gobio gobio | LC | 4 | ||
| Leuciscus cephalus | LC | 5 | ||
| Leuciscus souffia | LC | 4 | ||
| Phoxinus phoxinus | LC | 3 | ||
| Protochondrostoma genei | LC | 1 | ||
| Pseudorasbora parva | - | 4 | ||
| Rhodeus sericeus | - | 5 | ||
| Rutilus erythrophthalmus | LC | 3 | ||
| Rutilus pigus | DD | 2 | ||
| Rutilus rutilus | - | 5 | ||
| Scardinus erythrophtalmus | LC | 2 | ||
| Tinca tinca | LC | 2 | ||
| Cyprinodontiformes | Poecilidae | Gambusia holbrooki | - | 2 |
| Esociformes | Esociformi | Esox lucius† | LC | 2 |
| Gadiformes | Lotidae | Lota lota | LC | 1 |
| Gasterosteiformes | Gasterosteidae | Gasterosteus aculeatus | LC | 1 |
| Perciformes | Blennidae | Salaria fluviatilis | LC | 2 |
| Centrarchidae | Lepomis gibbosus | - | 3 | |
| Micropterus salmoides | - | 3 | ||
| Gobidae | Knipowitschia punctatissima | NT | 2 | |
| Padogobius martensii | LC | 3 | ||
| Percidae | Perca fluviatilis | LC | 2 | |
| Stizostedion lucioperca | - | 2 | ||
| Petromyzontiformes | Petromyzontidae | Lethenteron zanandreai | LC | 1 |
| Salmoniformes | Salmonidae | Oncorhynchus mykiss | - | 2 |
| Salmo (trutta) marmoratus† | LC | 1 | ||
| Thymallus thymallus | LC | 1 | ||
| Scorpaeniformes | Cottidae | Cottus gobio | LC | 1 |
| Siluriformes | Ictaluridae | Ictalurus melas | - | 1 |
| Ictalurus punctatus | - | 1 | ||
| Siluridae | Silurus glanis‡ | - | 4 |
| K Group | Central Value | Mean | 95% CI |
|---|---|---|---|
| 0.7–0.9 | 0.8 | 0.84 | 0.821–0.852 |
| >0.9–1.0 | 0.95 | 0.95 | 0.942–0.962 |
| >1.0–1.1 | 1.05 | 1.05 | 1.041–1.060 |
| >1.1–1.2 | 1.15 | 1.14 | 1.127–1.156 |
| >1.2–1.4 | 1.3 | 1.27 | 1.253–1.283 |
| K Group | Equation | R2 | p |
|---|---|---|---|
| I | y = 1.254 x3.6517 | 0.99 | <0.001 |
| II | y = 30.359 x2 − 69.158 x + 45.018 | 0.99 | <0.001 |
| III | y = 5.434 x3.2412 | 0.96 | <0.001 |
| IV | y = 7.842 x3.6157 | 0.98 | <0.001 |
| V | y = 18.056 x3.6038 | 0.99 | <0.001 |
| S. glanis Weight (g) | |||
|---|---|---|---|
| Fresh weight | 20 | 60 | 323 |
| Calculated Group I | 18.4 ± 1.7 | 54.9 ± 1.2 | 366 ± 26 |
| Calculated Group II | 22.2 ± 3.5 | 69.8 ± 8.6 | 309 ± 35 |
| Calculated Group III | 24.0 ± 4.3 | 70.8 ± 13.8 | 355 ± 84 |
| Calculated Group IV | 19.4 ± 5.0 | 71.2 ± 11.5 | 371 ± 126 |
| Calculated Group V | 18.6 ± 6.3 | 67.1 ± 26.8 | 338 ± 98 |
| Mean weight | 20.4 ± 4.9 | 67.5 ± 17.7 | 347 ± 88 |
| Coefficient of variation (%) | 24 | 26 | 25 |
| Stool ID | Sampling Date (DDMMYYYY) | Bone Remain | Mean Diameter (mm) | Height (mm) | K Index | K Group | Estimated Weight (g) |
|---|---|---|---|---|---|---|---|
| Ticino 25 | 22/10/2016 | c v | 4.31 | 3.43 | 0.8 | I | 260 |
| Ticino 26 | 22/10/2016 | c v | 0.99 | 1.03 | 1.04 | III | 5 |
| Ticino 26 | 22/10/2016 | c v | 1.98 | 1.69 | 0.86 | I | 15 |
| Ticino 27 | 10/09/2016 | c v | 3.25 | 3.6 | 1.11 | IV | 556 |
| Ticino 28 | 10/09/2016 | c v | 3.22 | 3.25 | 1.01 | III | 240 |
| Ticino 57 | 28/01/2017 | c v | 2.58 | 2.87 | 1.11 | IV | 241 |
| Ticino 58 | 28/01/2017 | c v | 0.86 | 1.19 | 1.38 | V | 10 |
| Ticino 58 | 28/01/2017 | c v | 0.73 | 1.13 | 1.55 | V | 9 |
| Ticino 61 | 28/01/2017 | c v | 2.26 | 1.7 | 0.75 | I | 25 |
| Ticino 61 | 28/01/2017 | c v | 1.4 | 1.44 | 1.03 | III | 16 |
| Ticino 61 | 28/01/2017 | c v | 1.54 | 1.58 | 1.03 | III | 22 |
| Ticino 95 | 18/03/2017 | c v | 1.8 | 2.2 | 1.22 | V | 150 |
| Ticino 58 | 28/01/2017 | p e | 2.75 a | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacchetta, R.; Marotta, A.; Nessi, A.; Tremolada, P. Back-Calculation of Fish Size in Diet Analysis of Piscivorous Predators: A New Index for the Alien Silurus glanis. Sustainability 2021, 13, 4322. https://doi.org/10.3390/su13084322
Bacchetta R, Marotta A, Nessi A, Tremolada P. Back-Calculation of Fish Size in Diet Analysis of Piscivorous Predators: A New Index for the Alien Silurus glanis. Sustainability. 2021; 13(8):4322. https://doi.org/10.3390/su13084322
Chicago/Turabian StyleBacchetta, Renato, Andrea Marotta, Alessandro Nessi, and Paolo Tremolada. 2021. "Back-Calculation of Fish Size in Diet Analysis of Piscivorous Predators: A New Index for the Alien Silurus glanis" Sustainability 13, no. 8: 4322. https://doi.org/10.3390/su13084322
APA StyleBacchetta, R., Marotta, A., Nessi, A., & Tremolada, P. (2021). Back-Calculation of Fish Size in Diet Analysis of Piscivorous Predators: A New Index for the Alien Silurus glanis. Sustainability, 13(8), 4322. https://doi.org/10.3390/su13084322