
Effect of Land-Use Change on the Changes in Human Lyme Risk in the United States



 and
and
Abstract
:1. Introduction
2. Materials and Methods
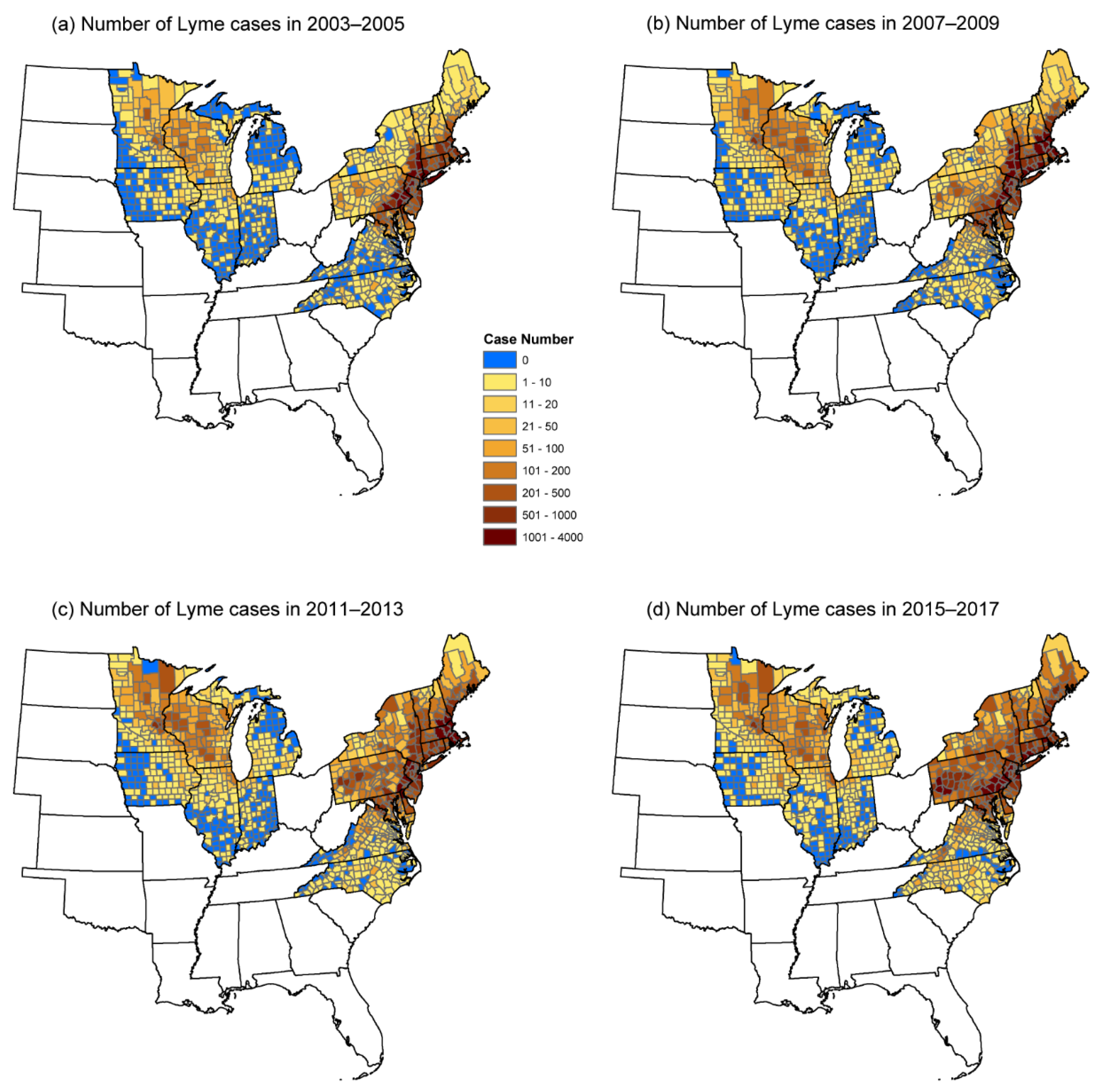
2.1. Lyme Disease Data
2.2. Data of Predictors
2.3. Statistical Analyses
3. Results
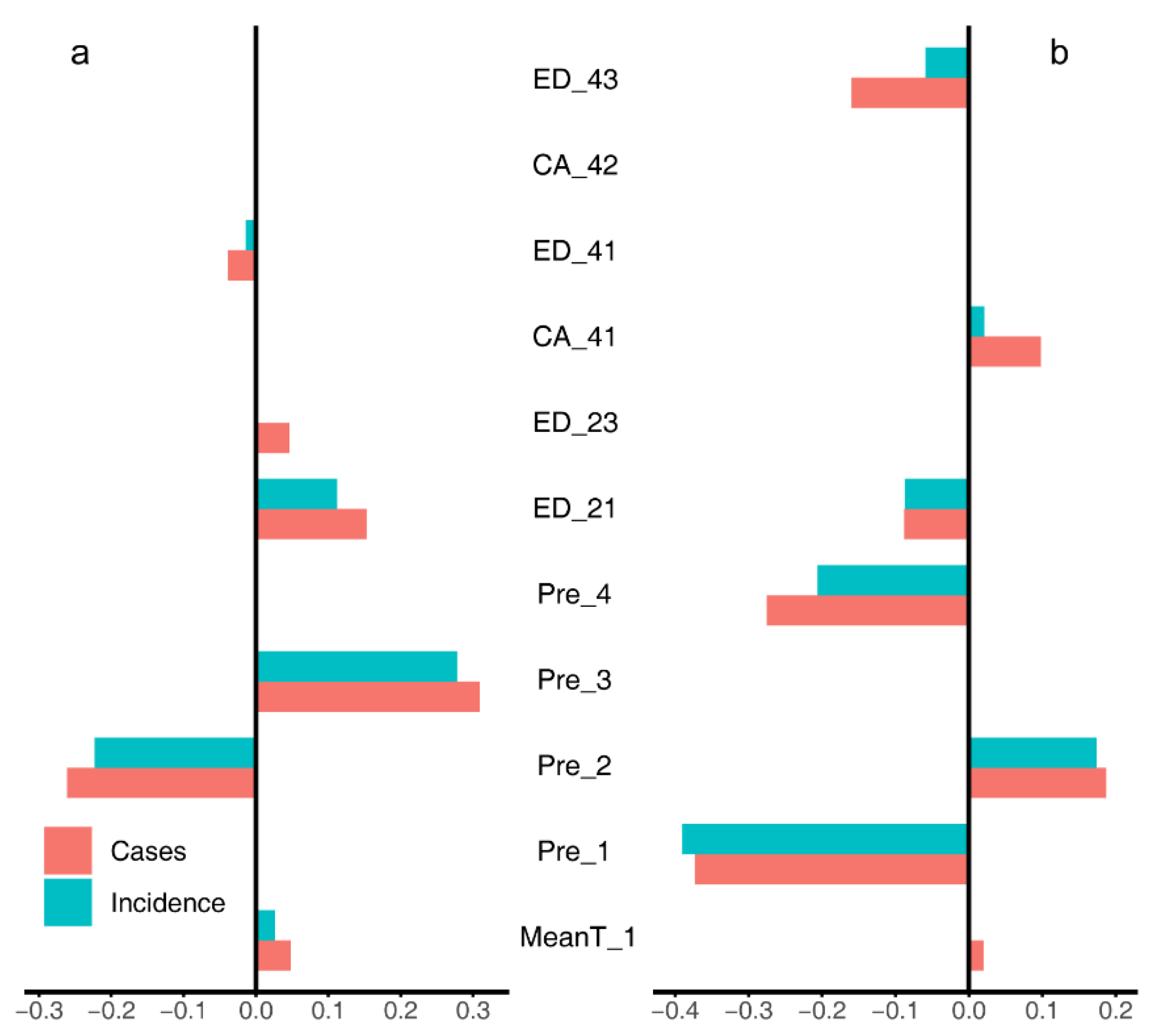
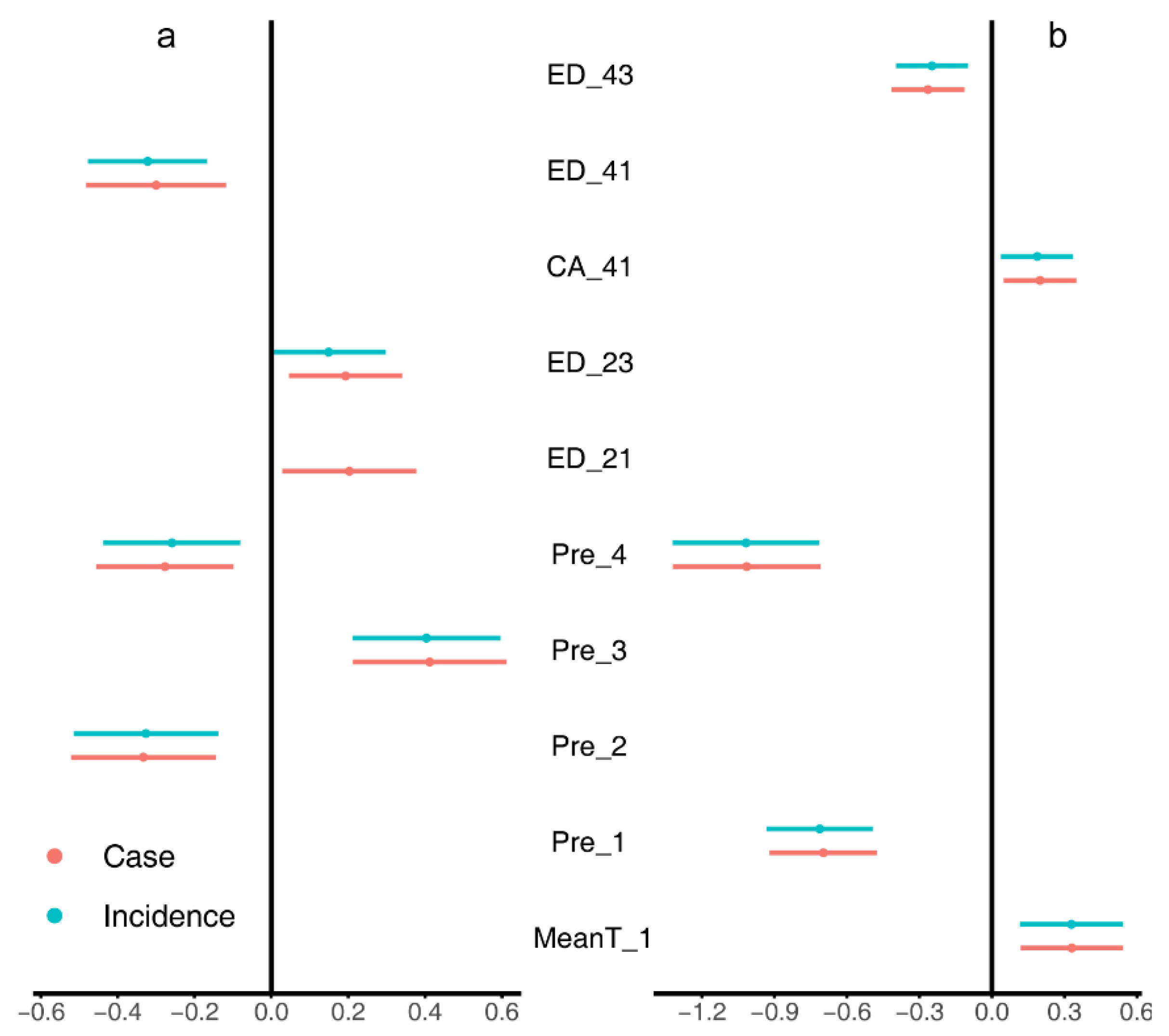
3.1. Univariate Regression Analyses
3.2. Multiple Regression Analyses
4. Discussion
4.1. Effects of Climatic Factors
4.2. Effects of Landscape Factors
4.3. LASSO and Model Averaging
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turney, S.; Gonzalez, A.; Millien, V. The negative relationship between mammal host diversity and Lyme disease incidence strengthens through time. Ecology 2014, 95, 3244–3250. [Google Scholar] [CrossRef]
- Wang, Y.X.; Matson, K.D.; Xu, Y.; Prins, H.H.; Huang, Z.Y.; de Boer, W.F. Forest connectivity, host assemblage characteristics of local and neighboring counties, and temperature jointly shape the spatial expansion of lyme disease in United States. Remote. Sens. 2019, 11, 2354. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A.M.; Pawlikowski, N.C.; Hamer, S.A.; Hickling, G.J.; Miller, J.R.; Schotthoefer, A.M.; Tsao, J.I.; Allan, B.F. Landscape features predict the current and forecast the future geographic spread of Lyme disease. Proc. R. Soc. B 2020, 287, 20202278. [Google Scholar] [CrossRef] [PubMed]
- Dumic, I.; Severnini, E. “Ticking bomb”: The impact of climate change on the incidence of Lyme disease. Can. J. Infect. Dis. Med. 2018, 2018, 5719081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Huang, Z.; Zhang, Y.; Wang, Y.X.; La, Y. Comparing the Climatic and Landscape Risk Factors for Lyme Disease Cases in the Upper Midwest and Northeast United States. Int. J. Environ. Res. Public Health 2020, 17, 1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmeester, T.R.; Jansen, P.A.; Wijnen, H.J.; Coipan, E.C.; Fonville, M.; Prins, H.H.; Sprong, H.; van Wieren, S.E. Cascading effects of predator activity on tick-borne disease risk. Proc. R. Soc. B 2017, 284, 20170453. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Piesman, J.; Zielinski-Gutierrez, E.; Eisen, L. What do we need to know about disease ecology to prevent Lyme disease in the northeastern United States? J. Med. Entomol. 2012, 49, 11–22. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Dobson, A.D.; Levi, T.; Salkeld, D.J.; Swei, A.; Ginsberg, H.S.; Kjemtrup, A.; Padgett, K.A.; Jensen, P.M.; Fish, D. Lyme disease ecology in a changing world: Consensus, uncertainty and critical gaps for improving control. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160117. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Canham, C.D.; Oggenfuss, K.; Winchcombe, R.J.; Keesing, F. Climate, Deer, Rodents, and Acorns as Determinants of Variation in Lyme-Disease Risk. PLoS Biol. 2006, 4, e145. [Google Scholar] [CrossRef]
- Lindgren, E.; Tälleklint, L.; Polfeldt, T. Impact of climatic change on the northern latitude limit and population density of the disease-transmitting European tick Ixodes ricinus. Environ. Health Perspect. 2000, 108, 119–123. [Google Scholar] [CrossRef]
- McPherson, M.; García-García, A.; Cuesta-Valero, F.J.; Beltrami, H.; Hansen-Ketchum, P.; MacDougall, D.; Ogden, N.H. Expansion of the Lyme disease vector Ixodes scapularis in Canada inferred from CMIP5 climate projections. Environ. Health Perspect. 2017, 125, 057008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, R.J.; Eisen, L.; Ogden, N.H.; Beard, C.B. Linkages of weather and climate with Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae), enzootic transmission of Borrelia burgdorferi, and Lyme disease in North America. J. Med. Entomol. 2016, 53, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werden, L.; Barker, I.K.; Bowman, J.; Gonzales, E.K.; Leighton, P.A.; Lindsay, L.R.; Jardine, C.M. Geography, deer, and host biodiversity shape the pattern of Lyme disease emergence in the Thousand Islands archipelago of Ontario, Canada. PLoS ONE 2014, 9, e85640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostfeld, R.S.; Brunner, J.L. Climate change and Ixodes tick-borne diseases of humans. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, D.D. Lower temperature limits for activity of several Ixodid ticks (Acari: Ixodidae): Effects of body size and rate of temperature change. J. Med. Entomol. 1995, 4, 449–452. [Google Scholar] [CrossRef]
- Ogden, N.H.; Lindsay, L.R.; Beauchamp, G.; Charron, D.; Maarouf, A.; O’Callaghan, C.J.; Waltner-Toews, D.; Barker, I.K. Investigation of Relationships Between Temperature and Developmental Rates of Tick Ixodes scapularis (Acari: Ixodidae) in the Laboratory and Field. J. Med. Entomol. 2004, 41, 622–633. [Google Scholar] [CrossRef] [Green Version]
- McCabe, G.J.; Bunnell, J.E. Precipitation and the occurrence of Lyme disease in the northeastern United States. Vector. Borne. Zoonotic. Dis. 2004, 4, 143–148. [Google Scholar] [CrossRef]
- Ballard, K.; Bone, C. Exploring spatially varying relationships between Lyme disease and land cover with geographically weighted regression. Appl. Geogr. 2021, 127, 102383. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; VanAcker, M.C.; Fernandez, M.P. Impact of Land Use Changes and Habitat Fragmentation on the Eco-epidemiology of Tick-Borne Diseases. J. Med. Entomol. 2020, 58, 1546–1564. [Google Scholar] [CrossRef]
- Killilea, M.E.; Swei, A.; Lane, R.S.; Briggs, C.J.; Ostfeld, R.S. Spatial dynamics of lyme disease: A review. Ecohealth 2008, 5, 167–195. [Google Scholar] [CrossRef] [Green Version]
- Wood, C.L.; Lafferty, K.D. Biodiversity and disease: A synthesis of ecological perspectives on Lyme disease transmission. Trends Ecol. Evol. 2013, 28, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, M.R.; Wilson, M.L. Microclimate-dependent survival of unfed adult Ixodes scapularis (Acari: Ixodidae) in nature: Life cycle and study design implications. J. Med. Entomol. 1996, 33, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.L.; Jordan, R.A.; Hung, R.W. Suppression of subadult Ixodes scapularis (Acari: Ixodidae) following removal of leaf litter. J. Med. Entomol. 1995, 32, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Brownstein, J.S.; Skelly, D.K.; Holford, T.R.; Fish, D. Forest fragmentation predicts local scale heterogeneity of Lyme disease risk. Oecologia 2005, 146, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Horobik, V.; Keesing, F.; Ostfeld, R.S. Abundance and Borrelia burgdorferi-infection prevalence of nymphal Ixodes scapularis ticks along forest–field edges. EcoHealth 2006, 3, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.M.; Waller, L. Effects of landscape fragmentation and climate on Lyme disease incidence in the northeastern United States. Ecohealth 2013, 10, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hartemink, N.; Speybroeck, N.; Vanwambeke, S.O. Consequences of landscape fragmentation on Lyme disease risk: A cellular automata approach. PLoS ONE 2012, 7, e39612. [Google Scholar] [CrossRef] [Green Version]
- Millins, C.; Dickinson, E.R.; Isakovic, P.; Gilbert, L.; Wojciechowska, A.; Paterson, V.; Tao, F.; Jahn, M.; Kilbride, E.; Birtles, R.; et al. Landscape structure affects the prevalence and distribution of a tick-borne zoonotic pathogen. Parasit Vector 2018, 11, 621. [Google Scholar] [CrossRef]
- Schauber, E.M.; Ostfeld, R.S.; Evans, J.; Andrew, S. What is the best predictor of annual Lyme disease incidence: Weather, mice, or acorns? Ecol. Appl. 2005, 15, 575–586. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3. 10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Wickham, J.; Homer, C.; Vogelmann, J.; McKerrow, A.; Mueller, R.; Herold, N.; Coulston, J. The multi-resolution land characteristics (MRLC) consortium—20 years of development and integration of USA national land cover data. Remote. Sens. 2014, 6, 7424–7441. [Google Scholar] [CrossRef] [Green Version]
- Tredennick, A.T.; Hooker, G.; Ellner, S.P.; Adler, P.B. A practical guide to selecting models for exploration, inference, and prediction in ecology. Ecology 2021, 102, e03336. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer: New York, NY, USA, 2009. [Google Scholar]
- Randin, C.F.; Dirnböck, T.; Dullinger, S.; Zimmermann, N.E.; Zappa, M.; Guisan, A. Are niche-based species distribution models transferable in space? J. Biogeogr. 2006, 33, 1689–1703. [Google Scholar] [CrossRef]
- Dhingra, M.S.; Artois, J.; Robinson, T.P.; Linard, C.; Chaiban, C.; Xenarios, I.; Engler, R.; Liechti, R.; Kuznetsov, D.; Xiao, X. Global mapping of highly pathogenic avian influenza H5N1 and H5Nx clade 2.3. 4.4 viruses with spatial cross-validation. eLife 2016, 5, e19571. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods. Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Bates, D.; Sarkar, D.; Bates, M.D.; Matrix, L. The lme4 package. R Package Version 2007, 2, 74. [Google Scholar]
- Barton, K. MuMIn: Multi-Model Inference. 2009. Available online: http://r-forge.r-project.org/projects/mumin/ (accessed on 23 April 2022).
- Hastie, T.; Qian, J.; Tay, K. An Introduction to Glmnet. CRAN R Repositary. 2021. Available online: https://cloud.r-project.org/web/packages/glmnet/vignettes/glmnet.pdf (accessed on 23 April 2022).
- Ostfeld, R.S.; Glass, G.E.; Keesing, F. Spatial epidemiology: An emerging (or re-emerging) discipline. Trends Ecol. Evol. 2005, 20, 328–336. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Ostfeld, R.S.; Peterson, A.T.; Poulin, R.; de la Fuente, J. Effects of environmental change on zoonotic disease risk: An ecological primer. Trends Parasitol. 2014, 30, 205–214. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Predictors | Descriptions | Notes |
|---|---|---|
| Pre_X | Seasonal mean precipitation in previous year | X = (1. spring; 2. summer; 3. autumn; 4. Winter) |
| MeanT_X | Seasonal mean temperature in previous year | |
| CA_Y | Total area of a land cover class | Y = (21–24,41–43) |
| ED_Y | Edge density of a land cover at the region |
| Variables | Upper Midwest | Northeast | ||
|---|---|---|---|---|
| b | t | b | t | |
| Mean spring temperature | −0.35 | −2.15 * | 0.37 | 3.42 *** |
| Mean summer temperature | −0.12 | −1.39 | 0.28 | 2.64 ** |
| Mean autumn temperature | −0.21 | −1.48 | 0.42 | 2.77 ** |
| Mean winter temperature | 0.07 | 0.75 | 0.13 | 0.86 |
| Mean spring precipitation | −0.07 | −0.59 | −0.89 | −8.36 *** |
| Mean summer precipitation | −0.30 | −2.99 ** | 0.30 | 3.21 ** |
| Mean autumn precipitation | 0.47 | 4.76 *** | 0.062 | 0.49 |
| Mean winter precipitation | −0.22 | −2.18 * | −1.18 | −7.18 *** |
| Cover area of open space | 0.29 | 3.71 *** | −0.08 | −0.97 |
| Edge density of open space | 0.37 | 4.93 *** | −0.17 | −2.22 * |
| Cover area of low-intensity space | 0.25 | 3.21 ** | −0.19 | −2.45 * |
| Edge density of low-intensity space | 0.27 | 3.54 *** | −0.18 | −2.20 * |
| Cover area of medium-intensity space | 0.18 | 2.18 * | −0.14 | −1.62 |
| Edge density of medium-intensity space | 0.22 | 2.76 ** | −0.13 | −1.50 |
| Cover area of high-intensity space | 0.02 | 0.23 | −0.04 | −0.47 |
| Edge density of high-intensity space | 0.14 | 1.78 | −0.06 | −0.68 |
| Cover area of deciduous forest | −0.20 | −2.75 ** | 0.24 | 3.01 ** |
| Edge density of deciduous forest | −0.38 | −4.87 *** | 0.01 | 0.14 |
| Cover area of evergreen forest | 0.04 | 0.49 | −0.08 | −1.05 |
| Edge density of evergreen forest | −0.04 | −0.49 | −0.19 | −2.45 * |
| Cover area of mixed forest | −0.18 | −2.20 * | −0.11 | −1.45 |
| Edge density of mixed forest | −0.18 | −2.25 * | −0.19 | −2.45 * |
| Predictors | Upper Midwest | Northeast | ||
|---|---|---|---|---|
| b | t | b | t | |
| Mean spring temperature | −0.36 | −2.22 * | 0.39 | 3.63 *** |
| Mean summer temperature | −0.13 | −1.53 | 0.30 | 2.83 ** |
| Mean autumn temperature | −0.24 | −1.67 | 0.45 | 2.93 ** |
| Mean winter temperature | 0.06 | 0.64 | 0.13 | 0.84 |
| Mean spring precipitation | −0.08 | −0.67 | −0.92 | −8.63 *** |
| Mean summer precipitation | −0.30 | −2.94 ** | 0.32 | 3.32 ** |
| Mean autumn precipitation | 0.46 | 4.65 *** | 0.08 | 0.61 |
| Mean winter precipitation | −0.20 | −2.07 * | −1.19 | −7.22 *** |
| Cover area of open space | 0.24 | 3.08 ** | −0.12 | −1.48 |
| Edge density of open space | 0.3 | 4.27 *** | −0.22 | −2.76 ** |
| Cover area of low-intensity space | 0.19 | 2.49 * | −0.25 | −3.17 ** |
| Edge density of low-intensity space | 0.22 | 2.81 ** | −0.24 | −2.93 ** |
| Cover area of medium-intensity space | 0.13 | 1.67 | −0.20 | −2.31 * |
| Edge density of medium-intensity space | 0.18 | 2.21 * | −0.19 | −2.21 * |
| Cover area of high-intensity space | −0.01 | −0.12 | −0.07 | −0.89 |
| Edge density of high-intensity space | 0.10 | 1.31 | −0.11 | −1.26 |
| Cover area of deciduous forest | −0.19 | −2.59 ** | 0.25 | 3.02 ** |
| Edge density of deciduous forest | −0.36 | −4.61 *** | 0.02 | 0.20 |
| Cover area of evergreen forest | 0.04 | 0.48 | −0.07 | −0.95 |
| Edge density of evergreen forest | −0.04 | −0.49 | −0.18 | −2.40 * |
| Cover area of mixed forest | −0.16 | −2.04 * | −0.10 | −1.33 |
| Edge density of mixed forest | −0.17 | −2.10 * | −0.17 | −2.27 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; He, G.; Yang, R.; Wang, Y.X.G.; Huang, Z.Y.X.; Dong, Y. Effect of Land-Use Change on the Changes in Human Lyme Risk in the United States. Sustainability 2022, 14, 5802. https://doi.org/10.3390/su14105802
Ma Y, He G, Yang R, Wang YXG, Huang ZYX, Dong Y. Effect of Land-Use Change on the Changes in Human Lyme Risk in the United States. Sustainability. 2022; 14(10):5802. https://doi.org/10.3390/su14105802
Chicago/Turabian StyleMa, Yuying, Ge He, Ruonan Yang, Yingying X. G. Wang, Zheng Y. X. Huang, and Yuting Dong. 2022. "Effect of Land-Use Change on the Changes in Human Lyme Risk in the United States" Sustainability 14, no. 10: 5802. https://doi.org/10.3390/su14105802
APA StyleMa, Y., He, G., Yang, R., Wang, Y. X. G., Huang, Z. Y. X., & Dong, Y. (2022). Effect of Land-Use Change on the Changes in Human Lyme Risk in the United States. Sustainability, 14(10), 5802. https://doi.org/10.3390/su14105802