Abstract
Asbestos is an industrially important microfiber present in cement industries and some mining sites and is very toxic to plant growth and development, but it has been neglected over the years. Therefore, this study was conducted to investigate the hazardous effects of asbestos on the growth and development of two important grass species (switchgrass and timothy grass). In order to mitigate the toxic effects of asbestos, a compost (bio-fertilizer) was also used. The asbestos soil samples were collected within a 10 km area of a cement factory. The results revealed that the asbestos-contaminated soils displayed a considerable increment in heavy metal uptake including chromium (Cr), manganese (Mn), vanadium (V), arsenic (As), and barium (Ba), which led to stunted plant growth. Consequently, the activities of antioxidant enzymes such as superoxide dismutase (SOD), glutathione peroxidase (GPX), and catalase (CAT) were stimulated to kept the redox balance under control. Among all the contaminated soils, the soil that was taken within a 0 Km area, closest to the cement factory, was the most toxic one. However, a compost amendment (25%) as a bio-fertilizer substantially reduced the toxic effects of asbestos fiber on the overall growth and development of plants, by reducing the metals’ uptake. Moreover, it was found that the roots of both grass species experienced higher heavy metal accumulation relative to the shoots. Collectively, it can be proposed that the studied grass species can be used for phytoextraction purposes, since both of them absorbed the heavy metals from the asbestos-contaminated soils.
1. Introduction
Asbestos, a highly toxic mining waste, is now categorized as a carcinogenic substance by the International Agency of Cancer Research [1]. This mineral belongs to the hydrous mineral silicates, which are essential components of the Serpentine and Amphibole groups, among a variety of minerals such as amosite, tremolite, actinolite, crocidite, and anthophyllite. Chrysotile is of great importance due to its long fibers and extensive use in the cement industry; on the other hand, it is also considered one of the most toxic microfibers among the other asbestos members [2]. Exposure to chrysotile results in lung cancer, asbestosis, mesothelioma, and cancer of the larynx and gastrointestinal tract in humans, especially asbestos workers [3,4]. Due to its toxic effects, many developed as well as developing countries including Canada, Brazil, and Colombia have already banned its production on all industrial levels [5]. Although, asbestos demand dropped due to its serious health hazards during the late 1970s, the widespread use of asbestos-containing materials for hospitals, gyms, schools, and industrial plants (cement, marble, and ship-breaking industries), during the period of asbestos manufacturing and consumption, and the handling and complete removal of asbestos-containing materials, means it is still a matter of global concern [6]. In Pakistan, asbestos is found in various forms, as some natural resources of this fiber are present in the Himalayan mountainous regions in the north and northwest of KPK Province; ShahKot Qilla, Mohmand Agency, and Malakand are rich natural sources of asbestos; and in Karachi, Hyderabad, and Lahore there are many industries manufacturing asbestos sewerage pipes and construction and roofing materials. Besides this quarrying of marble, the cement-manufacturing and ship-breaking industries are also a source of asbestos production [7,8]. These anthropogenic activities possess serious health hazards and, without taking the necessary preventive measures, can risk human lives [9]. Therefore, the remediation of such polluted sites is essential and can play a vital role in the elimination of toxic substances. From a crop point of view, few studies have investigated the toxic effects of asbestos-contaminated soils on the growth and development of plants including grains (Triticum aestivum), legume (Pisum sativum), and oil seed (Brassica juncea) crops and some grass species (Lemna gibba, C. citratus, C. zizanioides). Sandy-textured asbestos soils are generally rich in chromium and nickel, have a low water-holding capacity, and have a poor nutrient content, which ultimately result in poor plant growth and development [10]. Asbestos soils not only affect the plants’ morphological structure but also disturbs the plants’ physiological and biochemical defense mechanisms, by altering the antioxidants’ activity [11,12]. To reduce these hazardous effects of asbestos soils, different methods have been practiced: organic soil amendments, such as farmyard manure, chicken manure, compost, and sewage sludge, are one of the oldest, cheapest, and environmentally friendly options. Compost, rich in organic and mineral content, with a strong water-holding capacity [13], is considered as a less expensive method ameliorating the toxic effects of metal-contaminated soils as an organic soil amendment [10].
Several remediation approaches including physio-chemical treatments have been employed to purge harmful and noxious pollutants from contaminated sites; however, the restoration of contaminated lands by a layer of grass/plants has always been a cost-effective, non-invasive, and safe alternative to standard soil-cleaning technologies, by utilizing plant species such as trees, grasses, and shrubs to eliminate, degrade, or immobilize toxic elements from the soil [14]. Grasses have shown an effective role in the phytoextraction of toxic substances from the soil surface. Furthermore, grasses can effectively stabilize surface soils by improving the soil’s physical and chemical properties and retard its degradation [15]. Grass species are widely used as phytoremediation in different types of metal toxic soils, due to their fast-growing habit and large biomass production, such as switchgrass, a warm-season perennial species native to North America. Switchgrass (Panicum virgatum) has great adaptability and tolerance to different abiotic stresses including heat, drought, and potentially toxic heavy metals in soils, such as cadmium (Cd), lead (Pb), arsenic (As), copper (Cu), and zinc (Zn) [16,17]. Similarly, timothy grass (Phleum pretense), a perennial grass species native to Central Europe, is cold as well as drought-resistant [18]. The data regarding asbestos toxicity and phytoremediation against this hazardous microfiber are still lacking and need more scientific knowledge. However, it is considered that almost all grass/plant species show a similar response against biotic as well as abiotic stress, by activating their antioxidant defense mechanisms, which help to suppress the ROS generation in plant cells [19,20,21]. The scavenging or detoxification of excessive reactive oxygen species (ROS) by an anti-oxidative defense system consisting of enzymes such as peroxidase (POD), catalase (CAT), Ascorbate peroxidase (APX), glutathione peroxidase (GPX), and non-enzymes such as tocopherol, phenolic compounds, ascorbate (AsA), and carotenoids. Grass plants increase their tolerance against environmental stress by producing high levels of antioxidants to scavenge the ROS toxicity [22,23,24,25].
Keeping in view of the significance of turf grasses as phytoremediation against heavy-metal-contaminated soils, this research was conducted with the following objectives: (i) to investigate the chrysotile-asbestos-induced toxicity in the soil near a cement factory and its effects on plant growth; (ii) to explore the possible extent of damage caused by the asbestos-contaminated soil on growth and biochemical parameters; and (iii) to study the impact of bio-fertilizer (compost) in order to mitigate the hazardous effects of asbestos microfibers, to enhance plant growth and development.
2. Materials and Methods
2.1. Soil Sampling
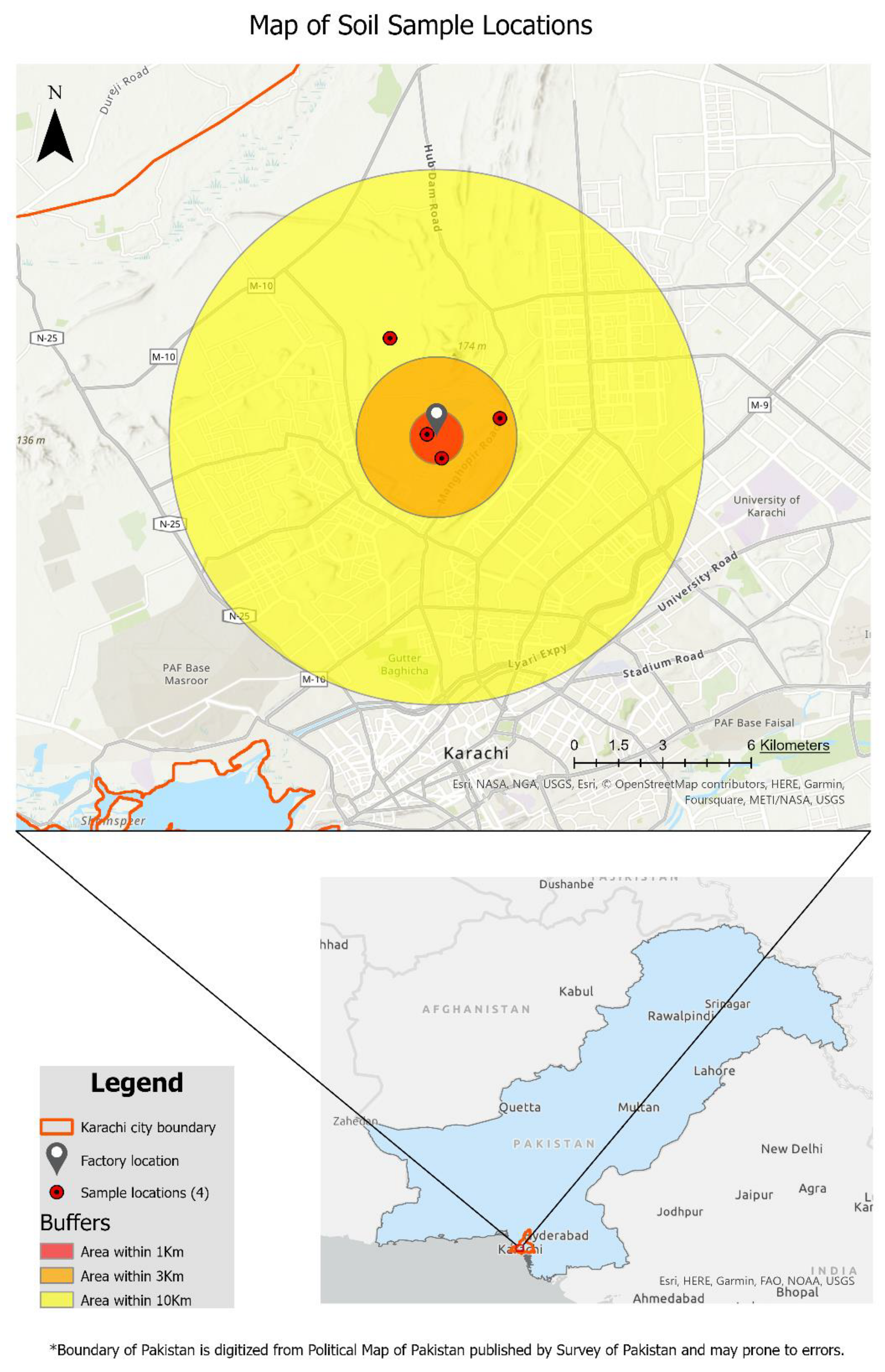
Heavy-metal-contaminated soil samples were collected from an industrial area where the chances of asbestos contamination were high. Soil samples were taken from a cement factory, randomly within the radius of 10 km, and at a depth of 75 cm, at Hyderabad Highway near Karachi, Pakistan, located at 24.962′ latitude and 67.019′ longitude. Soil of the site was sandy loan with poor water-holding capacity and high metal content. Four samples were taken in total and carefully packed in polythene bags, labeled, and transported to the laboratory for further analysis. Map of the sampling location was obtained by using ArcGIS Pro 3.1 (Figure 1). Samples were air-dried for 2 days, and impurities were removed by sieving the samples to ensure a constant particle size of 2 mm. Soil physio-chemical properties including pH, EC, total soluble salts (TSS), and soil salinity level were measured by using pH meter (Kent Eil 7015, Amesbury, MA, USA), conductivity meter (Model 4070, U.S. Salinity Lab, Riverside, CA, USA) by using digital Jenway method, and salinity sensor (for NaCl content) (Soil Moisture Equipment Corporation, Goleta, CA, USA), respectively. Further, 25% compost, obtained from animal manure, a cheaper and environmentally friendly bio-fertilizer, was used as an organic amendment to mitigate the toxic effects of asbestos on the growth and development of turf plants. In addition, the properties of the compost and asbestos-contaminated soil are presented in the Table 1.
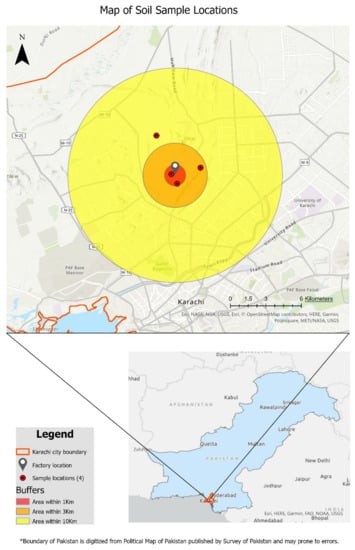
Figure 1.
A GIS map of the cement factory at Hyderabad Highway. The factory is located at the center, and the red-colored dots show the 4 sampling locations within the radius of 10 km.

Table 1.
Physio-chemical properties of compost-amended and chrysotile-asbestos-containing soils.
2.2. Experimental Setup
A pot experiment was conducted in a rain-out shelter with a transparent glass roof with no walls, to ensure ventilation and proper sunlight for plant growth. The two grass species named switchgrass (Panicum virgatum) and timothy grass (Phleum pretense) grass were involved in the experiments. A six-month long experiment was carried out, and 21-day-old grass seedlings (four seedlings per pot) were transplanted into the pots. The plastic pots were filled with asbestos and compost (obtained from animal manure) soils, according to the following treatments used in this experiment: T0 = control, T1 = asbestos soil (within 0 km area = 0A), T2 = asbestos soil (within 1 km area = 1A), T3 = asbestos soil (within 3 km area = 3A), T4 = asbestos soil (within 10 km area = 10A), T5 = 0A + compost (25%), T6 = 1A + compost (25%), T7 = 3A + compost (25%), and T8 = 10A + compost (25%). There were three biological and technical replicates for the experiment. All the pots were watered regularly at the start of the experiment. Afterwards, irrigation was maintained per the plant’s water requirement. Plants were allowed to grow under natural sunlight under covered roof, to avoid any leaching out of the heavy metals due to heavy rain. The position of the pots was changed twice a month, to abrogate the environmental effects due to the asbestos toxicity.
2.3. Parameters Studied
2.3.1. Growth, Biomass, and Heavy Metals
During harvesting, all the plants were carefully removed from the pots (to avoid root damage) and were brought to the laboratory. Harvested plants were thoroughly washed with tap water and then with distilled water (3–4 times) for measuring the total metal accumulation. The roots and leaves of the plants were separated manually (length and biomass of root and shoots were noted) and dried in an oven at 70 °C for 5 days. For the total metal accumulation, dried plant material was digested in HNO3 (70%) using a microwave digestion system (Milestone-Ethos One). As content was estimated by an atomic absorption spectrophotometer (GBC 906 AA), which was coupled to a hybrid generation system. Asbestos is an ally of different heavy metals such as chromium (Cr), nickel (Ni), cobalt (Co), iron (Fe), cadmium (Cd), lead (Pb), arsenic (As), manganese (Mn), etc. In this experiment, we tested the plant for the most toxic and abundantly present heavy metals among the asbestos allies, such as Cr, Mn, As, V, As, and Ba, in both the roots and shoots of two grass species [26].
2.3.2. Enzymatic Activities
In total, 200 mg plant material was homogenized in 100 mM Na-Phosphate buffer (pH 7) using liquid nitrogen in chilled pestle mortar, followed by centrifugation at 12,000× g for 10 min at 4 °C. The supernatant was then used to measure the activities of SOD, ascorbate peroxidase, guaiacol peroxidase, catalase, and glutathione S- transferase enzymes at 25 °C using a spectrophotometer (JASCO- V 530).
The activity of SOD was measured by the earlier published method [27]. The SOD activity was assayed by its ability to inhibit the photochemical reduction of Nitro Blue Tetrazolium (NBT). Inhibition of 50% shows the expression of one unit (1 U) of enzyme at 25 ± 2 °C. A system devoid of enzymes served as control.
The CAT activity was estimated by following the previously documented procedure [28] In brief, the reaction mixture contained 100 mM phosphate buffer (pH 7.0), 6.6 mM H2O2, and 50 μL plant extract. The activity was estimated by monitoring the decrease in absorbance due to H2O2 reduction (extinction coefficient: 39.4 M−1 cm−1) at 240 nm. One unit of catalase activity (U) represents the amount of enzyme catalyzing reduction of 1 μmol of H2O2 in 1 min at 25 ± 2 °C.
The GPX [29], and POD [30] activity was determined by following procedures and measured in U/g protein and U/mg/min FW, respectively.
2.3.3. Heavy Metals Concentration in the Sampling Soils
The amount of toxic heavy metals in the asbestos soil was determined by using the inductively coupled plasma membrane method, and the test was run for metals such as Mn, Cr, V, As, and Ba.
2.3.4. Statistical Analysis
A pot experiment was conducted with 9 treatments in total (3 replications each). All the data were analyzed statistically by using analyses of variance techniques (ANOVA) under complete randomized design (CRD). Treatment means were analyzed statistically by the least significance difference (LSD) test at a 5% level of probability by using SPSS software, and the co-relation was found by using OriginPro2021.
3. Results
3.1. Alleviation of Asbestos-Induced Damage in Growth Indices by Compost Amendment
Asbestos toxicity substantially affected the growth parameters of both the studied grasses as it was observed by the reduction in shoot and root length, and their biomass (Table 2). Among the asbestos treatments, the T1 had the highest effect on growth parameters, since a reduction of 52.8%, 68.2%, 57.6%, and 58.7% in shoot length (SHL), shoot biomass (SB), root length (RL), and root biomass (RB) was observed in switchgrass, respectively, when compared with untreated plants. The timothy grass showed a 49.8%, 57.4%, 22.6%, and 54.8% reduction in SHL, SB, RL, and RB, respectively, after exposure to the T1 treatment. However, the compost amendment in the asbestos-induced toxicity soil, remarkably, improved the growth indices of both the studied species in all the investigated areas. More specifically, relative to T1, T5 improved the SHL, SB, RL, and RB in switchgrass (1.6-, 2.6-, 2-, and 1.6-fold) and in timothy grass (1.5-, 1.9-, 1.1-, and 1.1-fold), respectively. Taken altogether, it was observed that among both grass species, switchgrass exhibited a more sensitive behavior towards the asbestos-inducted toxicity as compared to the timothy grass.
Table 2.
The deleterious effects of chrysotile asbestos on the morphological attributes of the Panicum virgatum and Phleum pretense grass species and its mitigation by organic compost.
3.2. Mitigation of Toxic Metals Uptake by Compost Amendment
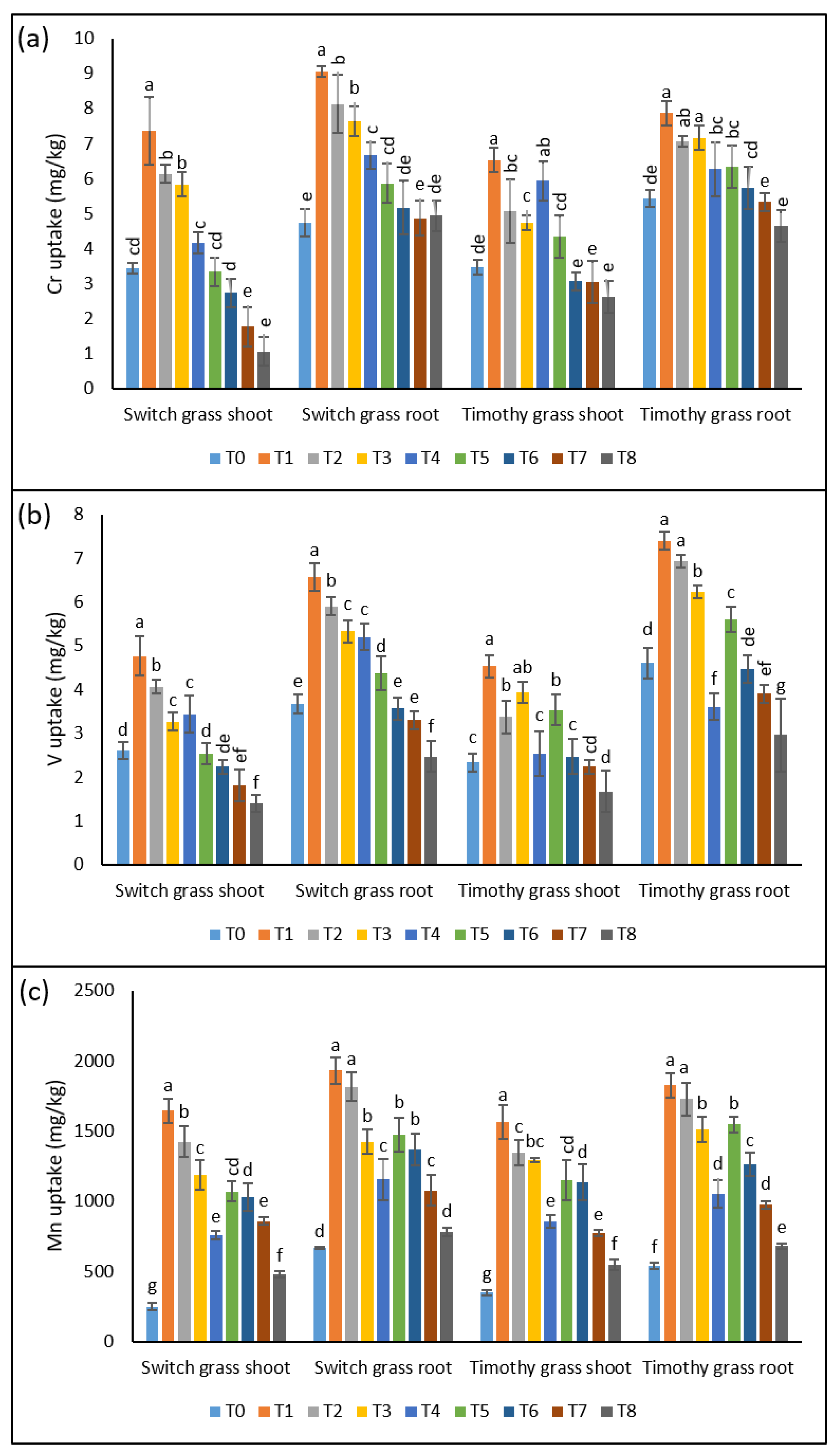
The experiment was conducted to assess the toxic effects of some minor and rare-earth elements such as chromium (Cr), vanadium (V), manganese (Mn), arsenic (As), and barium (Ba), which are highly toxic even in a small amount. The asbestos induced the higher uptake of metals in both the investigated grass species. The soil, which was taken within the 0 km area of the factory (T1), was the most toxic one, since the highest metals uptake was observed in both grass species as compared to the other soils.
3.2.1. Cr Uptake
In switchgrass, the Cr showed the elevation of a 2-, 1.8-, 1.7-, and 1.2-fold increment in the shoots and 1.9-, 1.7-, 1.6-, and 1.4-fold in the roots in T1, T2, T3, and T4, as compared to T0, respectively. However, the 25% supplementation of compost in the asbestos soil remarkably declined the Cr uptake in both organs of the switchgrass. For instance, a 54%, 55%, 70%, and 74% decline in Cr uptake was recorded for the shoots and 35%, 36%, 36%, and 35% for the roots in T5, T6, T7, and T8 over T1, T2, T3, and T4, respectively. While, in the case of timothy grass, T1, T2, T3, and T4 induced a 1.8-, 1.4-, 1.3-, 1.7-fold stimulation in Cr uptake by the shoots, respectively. The uptake of the roots of the timothy grass of the Cr metal was 1.4-, 1.3-, 1.3-, 1.15-fold higher in T1, T2, T3, and T4, over T0, respectively. However, T5, T6, T7, and T8 resulted in the inhibition of Cr uptake in the shoots (33%, 39%, 36%, and 55%) and in roots (19%, 18%, 25%, and 26%) relative to T1, T2, T3, and T4, respectively (Figure 2a).
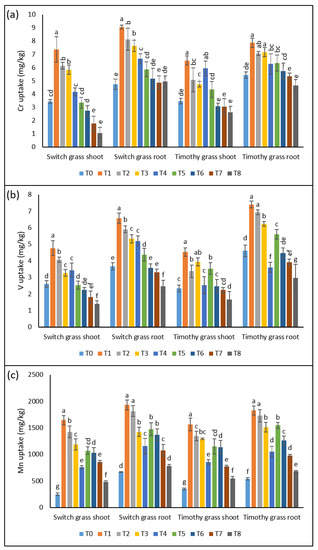
Figure 2.
The lucrative effects of compost on compost on the chrysotile-asbestos-induced damage in roots and shoots of the switchgrass and timothy grass species. (a) Chromium (Cr) uptake by the roots and shoots of the switchgrass and timothy grass species; (b) vanadium (V) uptake by the roots and shoots of the switchgrass and timothy grass species; (c) manganese (Mn) uptake by the roots and shoots of the switchgrass and timothy grass species. T0 = control, T1 = asbestos soil (within 0 km area = 0A), T2 = asbestos soil (within 1 km area = 1A), T3 = asbestos soil (within 3 km area = 3A), T4 = asbestos soil (within 10 km area = 10A), T5 = 0A + compost (25%), T6 = 1A + compost (25%), T7 = 3A + compost (25%), T8 = 10A + compost (25%). Values marked with different letters are significantly different from each other at p ≤ 0.05 levels, in the case of each species (ANOVA followed by least significant difference test; three independent experiments with three parallels each).
3.2.2. V Uptake
Similar to Cr, V accumulation was also noticed in both organs of the assessed grass species, after exposure to toxic soils containing asbestos. In contrast to control plants, T1, T2, T3, and T4 induced an increment of 1.8-, 1.5-, 1.25-, and 1.3-fold in shoots of switchgrass. Interestingly, the V-metal uptake under T4 was lower than the untreated plants. However, the remaining asbestos treatments noticeably stimulated the V-metal uptake in switchgrass roots. The 25% compost amendment resulted in the substantial inhibition of the V-metal uptake in all the sampling pots containing asbestos. Regarding timothy grass, the asbestos immensely stimulated the V-metal uptake by both the organs, while the 25% compost supplementation significantly declined its uptake in all the sampling sites’ soils (Figure 2b).
3.2.3. Mn Uptake
Relative to other metals, Mn concentration was relatively higher in asbestos-contaminated soil since an elevation of 6.6-, 5.7-, 4.7-, and 3-fold in the shoots and 2.8-, 2.7-, 2.1-, and 1.7-fold in the roots of switchgrass was recorded when exposed to T1, T2, T3, and T4, respectively. However, the 25% compost amendment under T5, T6, T7, and T8 inhibited the Mn-metal uptake by 35%, 27%, 27%, and 37% in the leaves and 23%, 24%, 24%, and 32% in the roots of switchgrass, as compared to T1, T2, T3, and T4, respectively. The timothy grass also showed a similar trend in Mn-metal uptake, as the uptake was also significant by the plants grown under asbestos-containing soil. However, the addition of 25% compost declined its uptake, indicating its mitigating effects under asbestos-induced stress (Figure 2c).
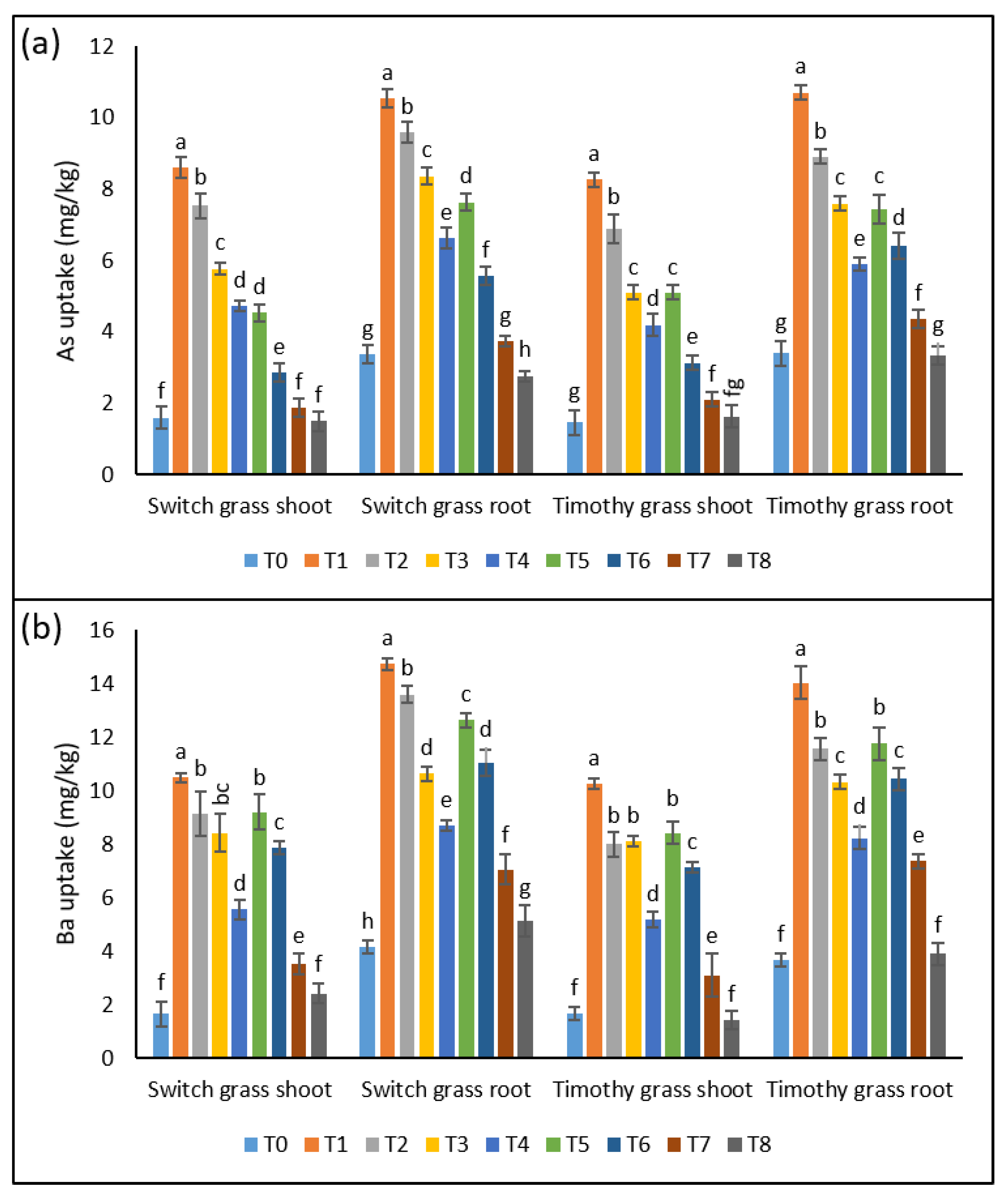
3.2.4. As Uptake
Both plant parts showed a significant rise in As uptake in both the examined species when grown under asbestos-containing soil. An amplification of 5.3-, 4.7-, 3.6-, and 3-fold in the shoots and 3.1-, 2.8-, 2.5-, and 2-fold in the roots of switchgrass was detected after T1, T2, T3, and T4, respectively. Whereas, T5, T6, T7, and T8 induced the inhibition in As uptake by 47%, 62%, 67%, and 68% in the shoots and 67%, 42%, 55%, and 59% in the roots over T1, T2, T3, and T4, respectively. In the case of timothy grass, T1, T2, T3, and T4 resulted in the rise of As uptake by 5.6-, 4.7-, 3.5-, 2.8-fold in the leaves and 3.1-, 2.6-, 2.2-, and 1.7-fold in the roots, respectively. Whereas, the plants showed tremendously lower As uptake when cultivated in 25% compost-amended soil (Figure 3a).
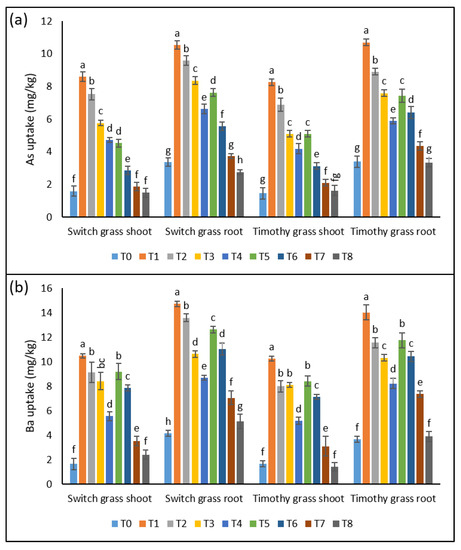
Figure 3.
A constructive effect of compost on compost on the chrysotile-asbestos-induced damage in roots and shoots of the switchgrass and timothy grass species. (a) Arsenic (As) uptake by the roots and shoots of the switchgrass and timothy grass species; (b) barium (Ba) uptake by the roots and shoots of the switchgrass and timothy grass species. T0 = control, T1 = asbestos soil (within 0 km area = 0A), T2 = asbestos soil (within 1 km area = 1A), T3 = asbestos soil (within 3 km area = 3A), T4 = asbestos soil (within 10 km area = 10A), T5 = 0A + compost (25%), T6 = 1A + compost (25%), T7 = 3A + compost (25%), T8 = 10A + compost (25%). Values marked with different letters are significantly different from each other at p ≤ 0.05 levels in the case of each species (ANOVA followed by least significant difference test; three independent experiments with three parallels each).
3.2.5. Ba Uptake
The results revealed the significant up-regulation in Ba uptake when both the studied grass species were exposed to asbestos (T1, T2, T3, and T4), as seen by the 6.3-, 5.5-, 5-, and 3.3-fold rise in the shoots and 3.5-, 3.2-, 2.5-, and 2-fold in the roots of switchgrass, respectively. Nonetheless, the 25% significantly decreased the Ba uptake in both organs in the sampling soils of all investigated sites around the cement factory. Timothy grass showed an increment of 6-, 4.8-, 4.8-, and 3.1-fold in the shoots’ Ba uptake and 3.8-, 3.1-, 3-, and 2.2-fold in the roots’ uptake under T1, T2, T3, and T4, compared to T0, respectively, while the induction of 25% compost in the asbestos-induced toxic soils led to a substantial decline in the Ba uptake in both the leaves and roots (Figure 3b).
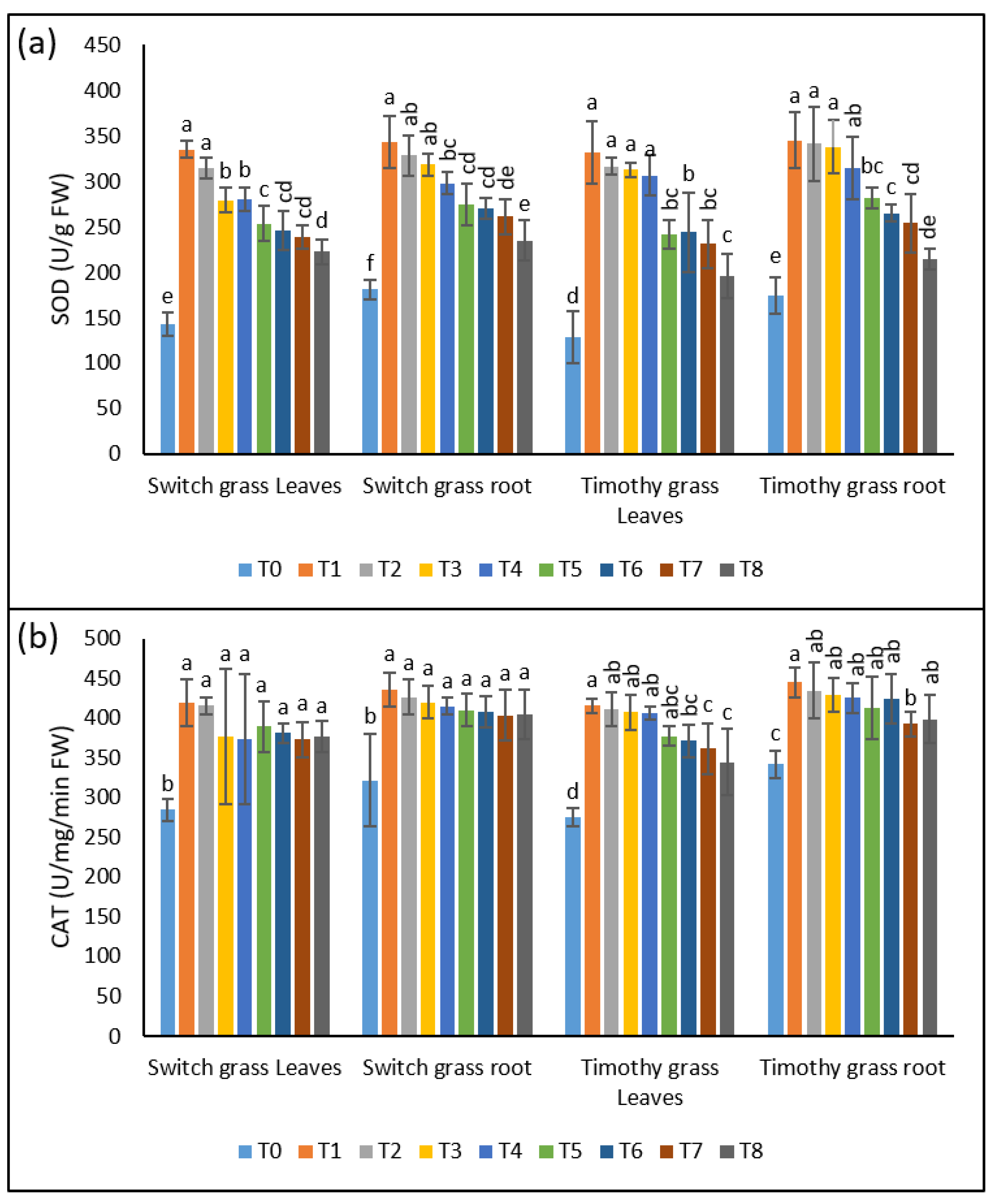
3.3. Compost-Induced Improvement in the Antioxidant Potential of Grasses under Asbestos Toxicity
The stimulation of antioxidant activities is an indication of stress in the plant tissues. Similar changes were exhibited by the leaves and roots of both the investigated grass species, when grown under asbestos-induced toxicity soils. In contrast to control plants, the most significant effect was shown by the T1 treatment among the asbestos treatments. T1, T2, T3, and T4 induced a 2.3-, 2.2-, 2-, 2-fold increment in SOD activities in the shoots and 1.9-, 1.8-, 1.7-, 1.6-fold in the roots of switchgrass, as compared to T0, respectively, indicating the persistence of stress by metals uptake. Nevertheless, T5, T6, T7, and T8 inhibited the SOD activities by 24%, 22%, 15%, and 21% in the leaves and 20%, 17%, 18%, and 21% in the roots over T1, T2, T3, and T4. Similar changes were observed in both organs of timothy grass, as they exhibited higher SOD activities after asbestos stress; however, this up-regulation was remarkably lowered by the supplementation of 25% compost (Figure 4a).
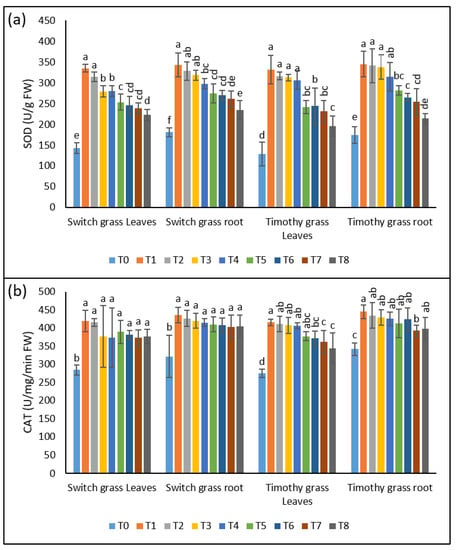
Figure 4.
The remunerative effect of compost on compost on the chrysotile-asbestos-induced damage in roots and leaves of the switchgrass and timothy grass species. (a) SOD activity in roots and leaves of the switchgrass and timothy grass species; (b) CAT activity in roots and leaves of the switchgrass and timothy grass species. SOD (Superoxide Dismutase) and CAT (Catalase). T0 = control, T1 = asbestos soil (within 0 km area = 0A), T2 = asbestos soil (within 1 km area = 1A), T3 = asbestos soil (within 3 km area = 3A), T4 = asbestos soil (within 10 km area = 10A), T5 = 0A + compost (25%), T6 = 1A + compost (25%), T7 = 3A + compost (25%), T8 = 10A + compost (25%). Values marked with different letters are significantly different from each other at p ≤ 0.05 levels in case of each species (ANOVA followed by least significant difference test; three independent experiments with three parallels each).
Likewise, for SOD, the CAT activities were also tremendously affected by asbestos stress, as can be observed by its 1.5-, 1.5-, 1.3-, and 1.3-fold up-regulation in the leaves and 1.3-fold in the roots of switchgrass after T1, T2, T3, and T4, respectively. After a 25% compost addition, the plants displayed a reduction in the CAT activities of both parts as compared to non-composted soils. Regarding timothy grass, the shoots’ CAT activity showed a 1.5-fold enhancement in all the asbestos-containing treatments, and the roots showed a 1.3-fold elevation of CAT activity when grown in asbestos-containing soils, while a noticeable inhibition was seen after compost amendment in both organs (Figure 4b).
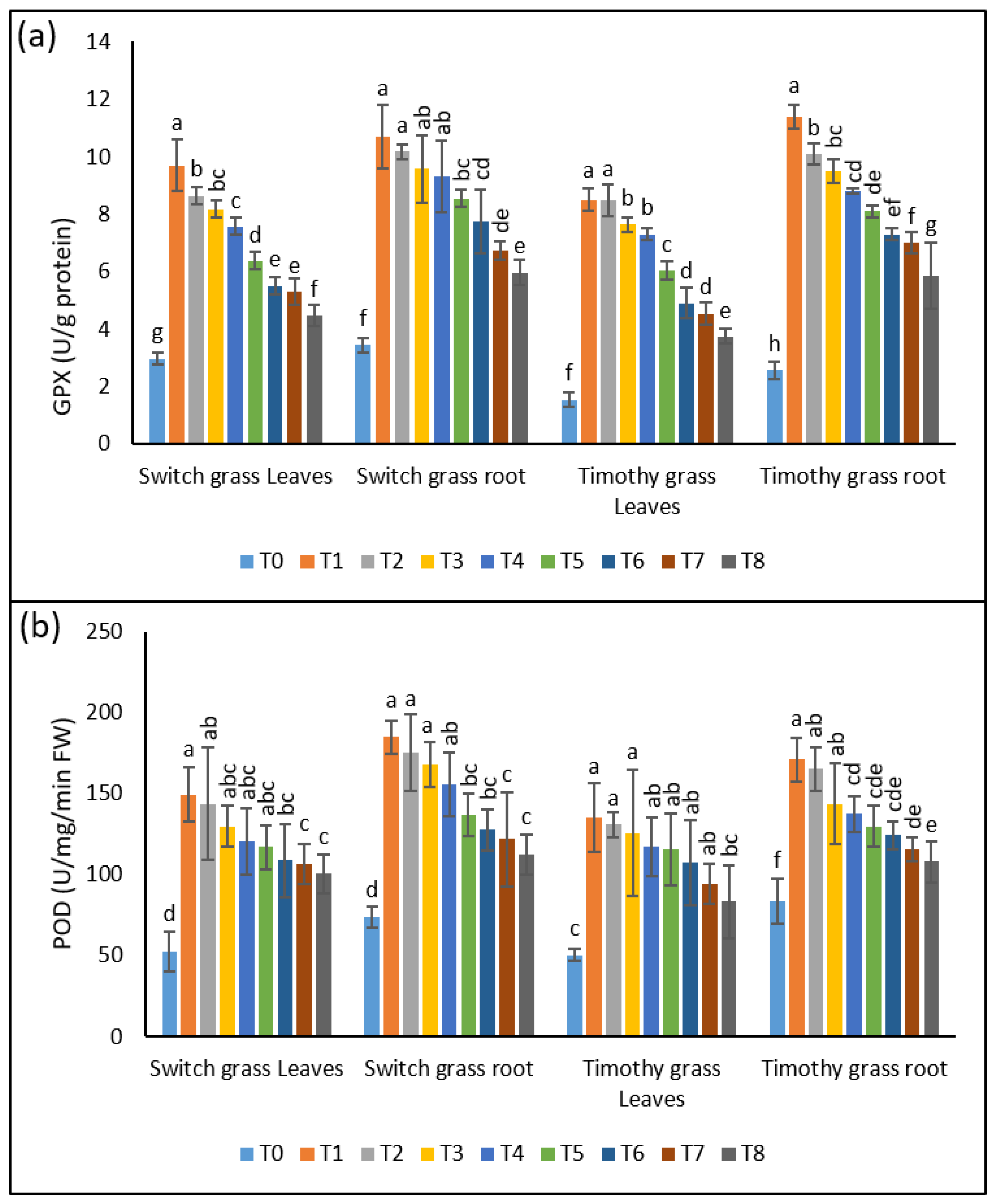
GPX is also an important component of plants’ antioxidant machinery, and it was considerably affected by the asbestos-induced heavy metals accumulation. A 3.2-, 2.9-, 2.7-, and 2.5-fold rise was seen in the shoots’ GPX activities of switchgrass after T1, T2, T3, and T4, respectively. This elevation was 3.1-, 2.9-, 2.8-, and 2.7-fold in the GPX activities of the roots, while a significant inhibition was noticed when the plants were grown under 25% composted soils. In the case of timothy grass, the shoots’ GPX activities were enhanced by 5.5-, 5.5-, 5-, and 4.7-fold after T1, T2, T3, and T4, respectively. In the roots, a 4.4-, 3.9-, 3.7, and 3.4-fold rise was found. Nonetheless, after the addition of 25% compost, a remarkable reduction in GPX activities was presented by both organs of timothy grass (Figure 5a).
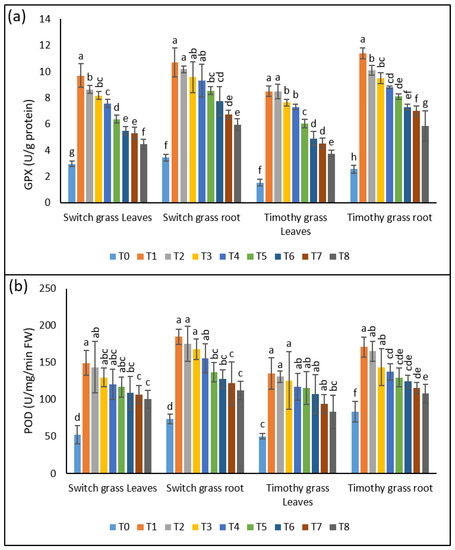
Figure 5.
The ameliorative effect of compost on compost on the chrysotile-asbestos-induced damage in roots and leaves of Panicum virgatum and Phleum pretense species. (a) GPX activity in roots and leaves of the switchgrass and timothy grass species; (b) POD activity in roots and leaves of the switchgrass and timothy grass species. GPX (Glutathione Peroxidase) and POD (Peroxidase). T0 = control, T1 = asbestos soil (within 0 km area = 0A), T2 = asbestos soil (within 1 km area = 1A), T3 = asbestos soil (within 3 km area = 3A), T4 = asbestos soil (within 10 km area = 10A), T5 = 0A + compost (25%), T6 = 1A + compost (25%), T7 = 3A + compost (25%), T8 = 10A + compost (25%). Values marked with different letters are significantly different from each other at p ≤ 0.05 levels in case of each species (ANOVA followed by least significant difference test; three independent experiments with three parallels each).
Similar changes were observed in the POD activities in the shoots and roots of both the studied species, after the interactive effect of asbestos and compost. The POD activities were raised significantly in the shoots (2.8-, 2.7-, 2.5-, 2.3-fold) and the roots (2.5-, 2.3-, 2.2-, 2.1-fold) of switchgrass when the plants were subjected to T1, T2, T3, and T4, respectively. This enhancement in both organs was somewhat reduced by the 25% compost amendment. The timothy grass displayed a considerable up-regulation in the POD activities of the shoots (2.7-, 2.6-, 2.5-, 2.3-fold) and roots (2-, 2-, 1.7-, 1.6-fold) in the plants grown in asbestos-containing soil. Similar to switchgrass, the timothy grass also presented lower POD activities in the roots and shoots when grown under 25% composted soil, indicating its crucial role in stimulating the antioxidant machinery of the plants to avoid the oxidative damage caused by asbestos toxicity (Figure 5b).
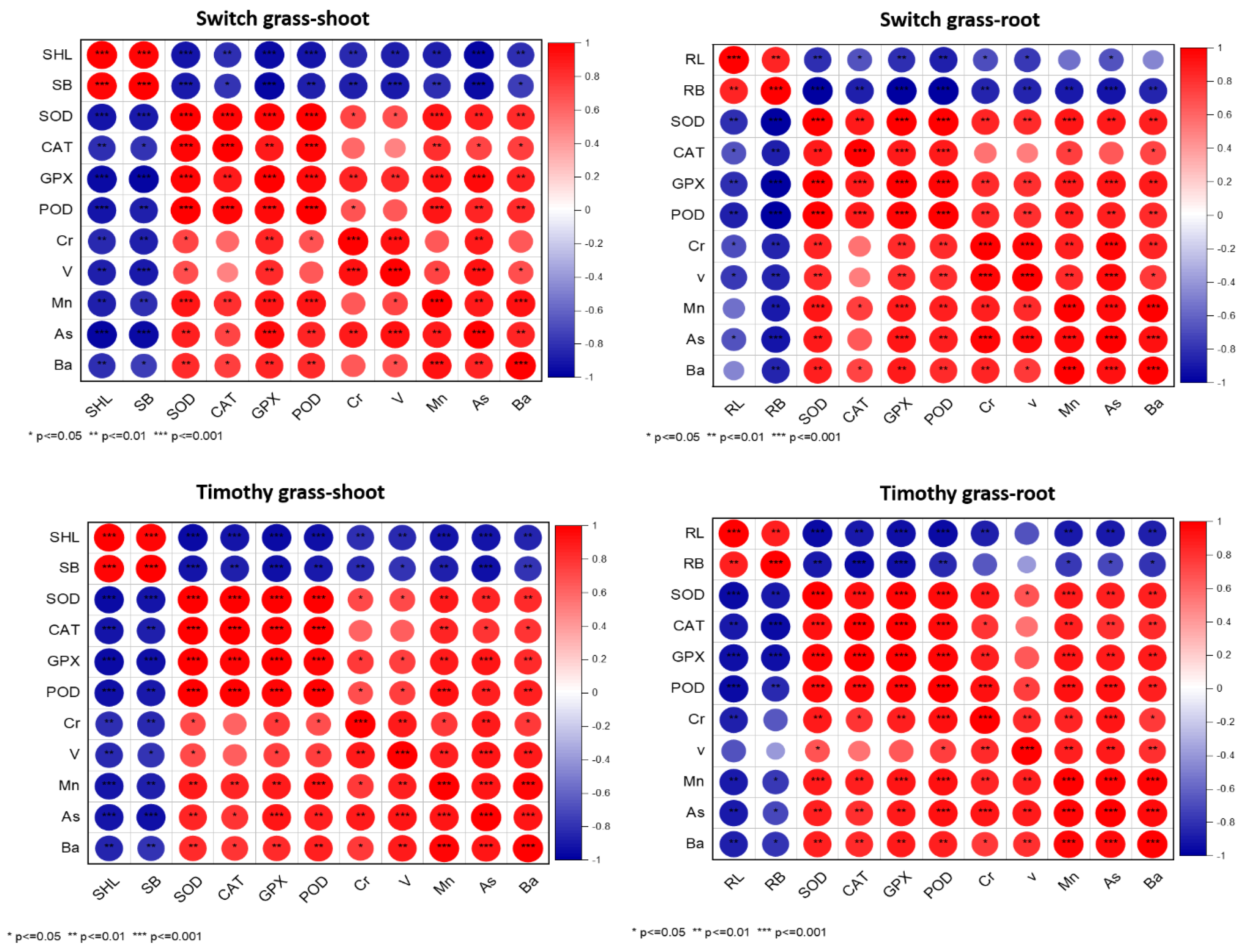
3.4. Correlation among the Investigated Parameters
Pearson’s correlation analysis showed that the SHL had a strong positive correlation with the SB, while it displayed a strong negative relationship with antioxidants and heavy metals in both grass species. Moreover, the RL exhibited a similar trend as SHL, suggesting the persistence of asbestos-induced stress in both the examined grass species (Figure 6). For the switchgrass roots and shoots, it was reported that Cr concentration was positively correlated with V, Mn, As, and Ba concentration and also SOD, POD, CAT, and APX activity, while it was negatively correlated with SHL and SB. Similar results were also obtained from the timothy grass roots and shoots: Cr concentration was positively correlated with V, Mn, As, and Ba concentration and also SOD, POD, CAT, and APX activity, while it was negatively correlated with SHL and SB. These results showed a close concentration between metal uptake and growth in switchgrass and timothy grass.
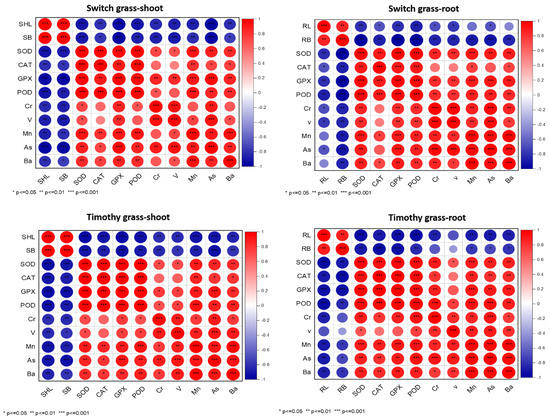
Figure 6.
Relationship between the studied parameters and propitious impact of compost on the chrysotile-asbestos-induced toxicity on Panicum virgatum and Phleum pretense species. Red color represents positive correlation, and blue color represent negative correlation (* p ≤ 0.05). The intensity of color represents the significance of the variables. Root length (RL), shoot length (SHL), root biomass (RB), shoot biomass (SB), SOD (Superoxide Dismutase), CAT (Catalase), GPX (glutathione peroxidase), POD (peroxidase), Cr (chromium), V (vanadium), Mn (manganese), As (arsenic), and Ba (barium) uptake by roots and shoots of the switchgrass and timothy grass species.
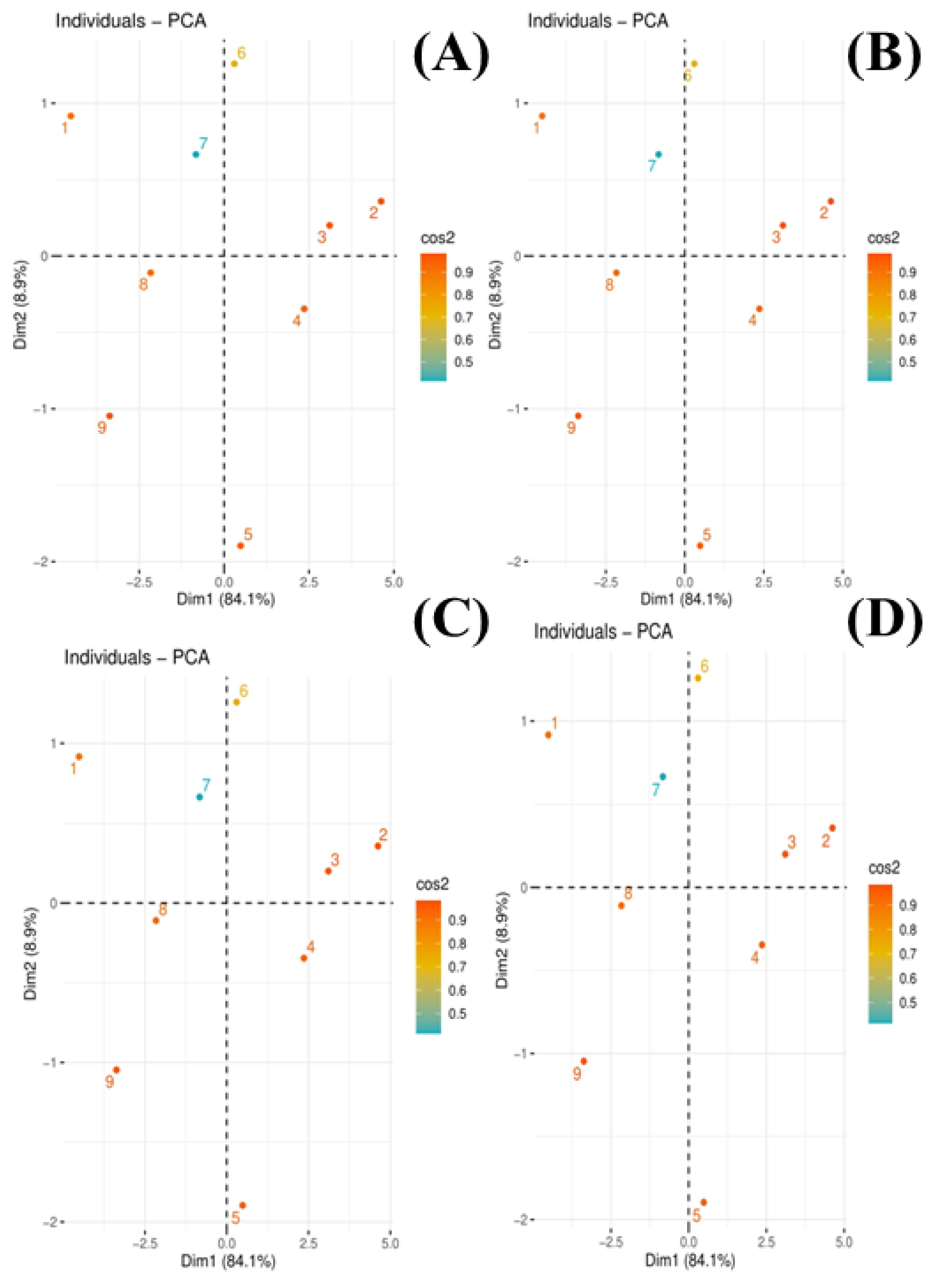
3.5. Principal Component Analysis
The scores of the principal component analysis (PCA) to evaluate the effects of compost on the chrysotile-asbestos-induced toxicity on the Panicum virgatum and Phleum pretense species are given in Figure 6. Among all the principal components, the first two components, i.e., PC1 (Dim 1) and PC2 (Dim 2), exhibited maximum contribution and accounted for 93% of the total variance in the dataset. Of them, PC1 contributed 84.1% while PC2 contributed 8.9%. All of the nine treatments were distributed successfully by the first two principal components (Figure 7). This distribution of treatments gave a clear indication that chrysotile asbestos induced toxicity on Panicum virgatum and Phleum pretense species compared to control (Figure 7). Moreover, (1) is distributed separately from all other scores of PCA, showing that inducing different treatments of chrysotile asbestos induced toxicity on Panicum virgatum and Phleum pretense species. This PCA showed a clear connection between the various studied treatments of chrysotile asbestos in Panicum virgatum and Phleum pretense species.
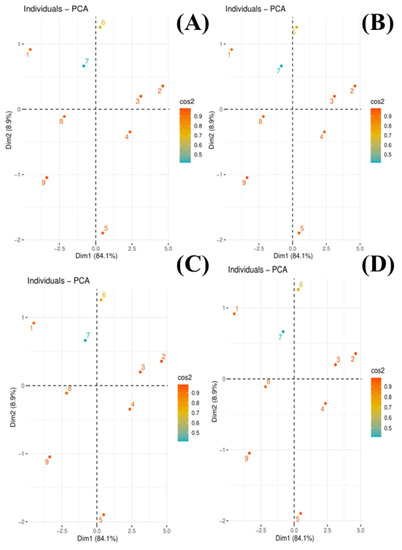
Figure 7.
Scores of principal component analysis (PCA) exhibited the effect of compost on the chrysotile-asbestos-induced toxicity on Panicum virgatum and Phleum pretense species. (A) is repressed as switchgrass shoots, (B) switchgrass roots, timothy grass shoots (C), and timothy grass roots (D). Different scores in the figures are presented as T1 = control, T2 = asbestos soil (within 0 km area = 0A), T3 = asbestos soil (within 1 km area = 1A), T4 = asbestos soil (within 3 km area = 3A), T5 = asbestos soil (within 10 km area = 10A), T6 = 0A + compost (25%), T7 = 1A + compost (25%), T8 = 3A + compost (25%), T9 = 10A + compost (25%).
4. Discussion
The results revealed that the asbestos toxicity adversely affected the plant growth, as was seen by the reduced length of roots and shoots and the reduced biomass in both grass species. Our findings are in line with the previous studies [31,32]. Another study on the hazardous effects of asbestos exhibited inhibited seed germination in wheat, pea, and mustard due to asbestos toxicity [33,34]. There has been little scientific knowledge on the potential use of grasses such as Panicum virgatum and Phleum pretense, as phytoremediation against heavy-metal-contaminated soils [18]; however, data on the use of switchgrass and timothy grass species in the asbestos-contaminated sites are limited. The application of compost as a soil amendment in the polluted soil resulted in a significant increase in the morphological attributes of switchgrass, i.e., the length of the roots and shoots and biomass production. Our results are found to be in favor of this study, where the compost-amended soil enhanced plant growth and biomass by decreasing the concentration of heavy metals in various parts of the plant species [35]. On the basis of the physio-chemical properties of compost, we can assume that compost played a significant role in alleviating the toxic effects of asbestos in plants, however, to better understand this phenomenon, more studies are needed in future on the soil fertility and the extractable and non-extractable metals in the asbestos-contaminated soils.
Cr contamination in both the grass species was higher when the plants were grown under asbestos-containing soil, however, the switchgrass outperformed the timothy grass. Cr is a potentially toxic metal that does not have any essential metabolic function in plants, and its excess concentration in the soil may cause toxic effects in plants and reduce the growth, photosynthesis, mineral nutrients, and quality of the crops. Some timothy grass plants in non-amended soil showed minor toxicity symptoms such as chlorosis, but the compost amendment significantly improved the growth rate of plants; however, these results were in line with previous studies [36], where switchgrass performed well under Cr stress. This contradiction might be due to the presence of highly toxic heavy metals in asbestos soil. Similar results were obtained by [32], where the vativer plants were grown under Cd–Cr stress in a hydroponic system and the plants showed similar results under this Cd–Cr combined toxicity. Our results revealed that the Cr uptake from the non-amended soils was higher, and the amendment of compost significantly reduced Cr uptake; a similar conclusion was reached by [33,34], in which some organic soil amendments were used to reduce the hazardous effects of asbestos in Cymbopogon citratus and Chrysopogon zizanioides. Likewise, V, a potentially toxic, non-essential, rare-earth element accumulated after the asbestos exposure in both grass species. Another study has also confirmed its presence in the asbestos-contaminated soil [1]. A previous study was found in favor of our current research, in which the massive accumulation of V metal caused phyto-toxicity in plant growth and physiological attributes [37]. Regarding the organ-specific accumulation, the V uptake in the underground parts of lettuce, green beans, Glycine max, Phaseolus vulgaris, Setaria viridis, L. sativa, P. virgatum, and E. virginicus plants was higher than that of the aerial parts, and these results are in line with our findings as well [37,38,39,40,41]. However, a contrary statement was also found, where the V accumulation was relatively high in the aerial parts of green beans, forb species (L. americanus and P. vulgaris), and dog’s-tail grass [38,40]. These studies scientifically proved the toxic effects of V on different crops, while there is a paucity of data on V toxicity, especially in asbestos-contaminated soils.
Mn is an essential plant nutrient in relatively low amounts, but it has toxic effects on plant growth and development [33,42]. Similar results were obtained in our research regarding the growth indices of both grass species. Other results were broadly in line with the current findings, except for soybean [43], in which the Mn uptake was higher in the roots of Oryza sativa, Brassica napus, and sugarcane, as compared to the shoots [44,45,46]; whereas, in soybean, the Mn concentration was considerably high in the aerial parts of the plants. As is a highly toxic and carcinogenic element, and the most widespread sources of As in soil and water are natural sources, such as volcanic activities, weathering, the erosion of minerals and rocks, and geothermal waters [47]. It is well-known that As toxicity in crops depends on the bioavailability of As in soils and the concentration of elements, which can compete with As during plant uptake. In the current study, the soil samples taken around the cement factory indicated the presence of As in the asbestos soil. Similar to other heavy metals, the As uptake was relatively higher in the roots compared to the aerial parts of the plants, although a slight difference in the uptake of As by the roots was found in both the tested grass species. These findings are parallel to a previous report, where Vetiveria zizanoides plants showed a similar behavior under As stress [48]. Another study was found in favor of our current findings, in which Solanum tuberosum showed stunted growth under Cd–As combined stress [49]. Moreover, the accumulation of As in Oryza sativa [50,51] and Glycine max [52] plants displayed reduced plant growth. In Festuca rubra [53], Pteris vittate, and Pteris cretica (>99 of As accumulation in roots) [54], as well as Agrostis capillaris and Holcus lanatus [55,56], the As concentration was relatively higher in rthe oots, and our results are broadly in line with these studies. Another study was found on the organic and inorganic As availability in boron rice, and the key findings of this research were in accordance with our current study, where the inorganic As concentration was substantially higher in the roots compared to the leaves and grains [57]. However, some species (Equisetum spp. and C. epigejos) exhibited contrary results to the current findings [55]. The purple guinea and Ruzi grass species showed a significant bioaccumulation of As in the roots of the plants compared to the shoots and leaves [58]. The barnyard grass varieties proved an excellent source of As removal from the soil, irrespective of the As concentration in the soil, while Nipponbare rice species were found to be the best for As remediation [59]; these findings are consistent with the current study.
Ba, a potentially toxic heavy metal associated with asbestos fibers, is considered toxic for plant growth and development [1]. In the current investigation, Ba accumulation was also observed when the plants were cultivated under asbestos toxicity. Cucumis sativus L. showed growth retardation, high Ba accumulation in both the roots and shoots of the plants, low biomass of the roots/shoots, and higher levels of CAT and GPX, when exposed to a high concentration of Ba [60]; these findings were found to be in line with our current results. The data demonstrated that a maximum of Ba was retained in the roots of the plants and a relatively small concentration in non-amended soils moved to the aerial parts of the soils; similar results were obtained by previous studies [61]. A similar pattern was also demonstrated by another report, in which the white clover and tall fescue took up a maximum of Ba from the exchangeable fraction of growth media, and white clover outperformed tall fescue in Ba mobilization than [62]. Another study on cherry orchards was found in accordance with our findings, which demonstrated that the Ba uptake in the roots and leaves of the cherry plants is highly dependent on the soil pH; a significant correlation was observed between the plant leaves and acidic soil fraction [63]. In the present report, the compost amendment substantially reduced its uptake in both plant organs in both the studied grass species. A comprising study was also found in favor of our results on Ba removal from the flooded soils in the studied species (C. papyrus and T. domingensis were found to be potential candidates for phytoremediation of Ba toxicity) [64]. A similar pattern of results was found in another study, where the mono-cropping and inter-cropping patterns were analyzed in Ba-contaminated soil; the results showed that T. domingensis in the inter-cropping pattern absorbed the highest amount of Ba from the soil and facilitated Ba removal, by translocating it to the aerial parts of the plants [65]. A greenhouse study was conducted to check the effects of lime on the toxicity of Ba on nine different types of plant species, including grasses, legumes, and vegetables, and the results were in accordance with our current findings, which highlighted that lime significantly reduced the Ba concentration in aerial parts of the plants [66]. However, a contrary study was found, where rice samples were collected and studied in a Ba mining area; the results demonstrated that the Ba phytoavailability in rice was relatively low, and the main phytoavailable source of Ba could be the water, not the soil or any other chelating agent; moreover, the planned comparison revealed that the risk assessment was significantly low in the local residents, when exposed to the low Ba rice consumption [67].
To deal with the environmental stresses, plants have their own defense mechanism, which includes both enzymatic and non-enzymatic antioxidants. A series of evidence are available for the up-regulation of different antioxidant enzymes, after exposure to various heavy metals, such as Cd, Pb, As, V, Mn, Co, Ni, Hg, Fe, and Cr [31,32,38,68,69,70], while only a few studies have focused on the toxic effects of asbestos on the biochemical characteristics of the studied grass species. In the current research, the asbestos-induced stress considerably up-regulated the activities of SOD, CAT, GPA, and POD in both plant organs in both the studied grass species to reduce the metal uptake. However, a 25% compost amendment in the soil significantly reduced the activities of all the studied enzymes, indicating its stress-alleviating effects. A similar study to the current research was found, in which the different types of organic fertilizer were observed to mitigate the hazardous effects of asbestos in C. citratus and C. zizanioides [33]. Collectively, it is proven that under-stress plants activate antioxidant potential to reduce oxidative stress and phytotoxicity. The limitations of this study include reactive species (OH, O2−, 1O2, and MDA content), reactive carbonyl (RCS), and sulphur species (RSS), as they either play a reactive or signaling role in plants under asbestos stress. Although, the significantly higher concentration of SOD, CAT, POD, and GPA revealed that the ROS toxicity might be high in the plants’ roots and shoots, as a result these antioxidants were produced in response to the stress. Further studies are needed to understand the whole defense mechanism opted by switchgrass and timothy grass under asbestos stress.
5. Conclusions
The results of this study revealed that chrysotile asbestos has toxic effects on the overall growth of both grass species. All the morphological parameters including RL, SHL, RB, and SB were significantly influenced by the chrysotile asbestos’ toxicity. The metal uptake, such as of Cr, Mn, V, As, and Ba, was considerably high in the roots of both grass species, as compared to their aerial parts, suggesting that both grass species can be considered as hyper-accumulator plants in asbestos-prone areas. However, the compost amendment considerably reduced the metal toxicity in both grass species. It can be considered that the higher metal uptake might be due to the low soil fertility or the high toxicity of the asbestos. The physio-chemical properties of asbestos provide evidence that the heavy metal uptake is due to low TSS, acidic pH, low EC, and the high amounts of metals and might be due to the low soil fertility, while future molecular studies are needed to better understand this phenomenon. Another finding of this research is that the antioxidant enzymes, including SOD, CAT, GPX, and POD, were considerably high in asbestos-stressed plants, indicating that the plants activated their defense mechanism to cope with the toxic effects of chrysotile asbestos. An apparent limitation to this research is the limited knowledge previously available. Given this, more scientific knowledge is required to gain deeper insights into the effects of asbestos toxicity on plant growth; its soil–roots and roots–shoots translocation factors and their mechanisms; the soil toxicity and fertility effects on grass production and heavy-metal accumulation; the oxidative stress management in plants; and the cellular stress-management pathway opted for by plants, especially in switchgrass and timothy grass, to avoid the toxic effects of heavy metals.
Author Contributions
Conceptualization, M.A.A. and K.S.; formal analysis, M.H.S., A.R., G.K., B.A. and M.F.A.; funding acquisition, M.H.S., B.A., M.F.A. and E.A.B.; investigation, K.S.; methodology, M.A.A.; resources, E.A.B.; software, A.R., B.A. and E.A.B.; supervision, M.A.A.; validation, G.K. and N.I.; visualization, N.I.; writing—original draft, K.S. and M.A.A.; writing—review and editing, M.A.A., M.H.S., A.R., G.K., N.I., B.A. and M.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Researchers Supporting Project number (RSP-2022R436), King Saud University, Riyadh, Saudi Arabia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge the support provided by the Researchers Supporting Project number (RSP-2022R436), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bloise, A.; Ricchiuti, C.; Punturo, R.; Pereira, D. Potentially Toxic Elements (PTEs) Associated with Asbestos Chrysotile, Tremolite and Actinolite in the Calabria Region (Italy). Chem. Geol. 2020, 558, 119896. [Google Scholar] [CrossRef]
- Qaisar, M.A.; Khan, A.A.H. Mineralogy of Some Asbestos from North-West Pakistan. J. Sci. Ind. Res. 1967, 10, 116–120. [Google Scholar]
- Bloise, A.; Catalano, M.; Barrese, E.; Gualtieri, A.F.; Bursi Gandolfi, N.; Capella, S.; Belluso, E. TG/DSC Study of the Thermal Behaviour of Hazardous Mineral Fibres. J. Therm. Anal. Calorim. 2016, 123, 2225–2239. [Google Scholar] [CrossRef]
- Harper, M. 10th Anniversary Critical Review: Naturally Occurring Asbestos. J. Environ. Monit. 2008, 10, 1394–1408. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Sun, X.-M.; Wu, L. High Time for Complete Ban on Asbestos Use in Developing Countries. JAMA Oncol. 2019, 5, 779–780. [Google Scholar] [CrossRef] [PubMed]
- Gregson, N.; Watkins, H.; Calestani, M. Inextinguishable Fibres: Demolition and the Vital Materialisms of Asbestos. Environ. Plan. A 2010, 42, 1065–1083. [Google Scholar] [CrossRef]
- Jehan, N.; Ahmad, I. Petrochemistry of Asbestos Bearing Rocks from Skhakot-Qila Ultramafic Complex, Northern Pakistan. J. Himal. Earth Sci. 2006, 39, 75–83. [Google Scholar]
- Iftikhar, S.; Ali, M.; Nergis, Y. Risks and Hazards Study of Asbestos in Pakistan. Int. J. Econ. Environ. Geol. 2019, 6, 25–28. [Google Scholar]
- Baumann, F.; Buck, B.J.; Metcalf, R.V.; McLaurin, B.T.; Merkler, D.J.; Carbone, M. The Presence of Asbestos in the Natural Environment Is Likely Related to Mesothelioma in Young Individuals and Women from Southern Nevada. J. Thorac. Oncol. 2015, 10, 731–737. [Google Scholar] [CrossRef]
- Ahmad, M.; Ishaq, M.; Shah, W.A.; Adnan, M.; Fahad, S.; Saleem, M.H.; Khan, F.U.; Mussarat, M.; Khan, S.; Ali, B.; et al. Managing Phosphorus Availability from Organic and Inorganic Sources for Optimum Wheat Production in Calcareous Soils. Sustainability 2022, 14, 7669. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Javed, M.A.; Ahmad, S.; Afridi, M.S.; Sumaira Nadeem, M.; Khan, A.U.R.; Malik, A.; Ullah, A.; Alwahibi, M.S.; et al. Bacterial-mediated salt tolerance in maize: Insights into plant growth promotion, antioxidant defense system, oxidative stress, and surfactant production. Front. Plant Sci. 2022, 13, 978291. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Ahmad, S.; Javed, M.A.; Sumaira Afridi, M.S.; Dawoud, T.M.; Almaary, K.S.; Muresan, C.C.; Marc, R.A.; Alkhalifah, D.H.M.; et al. Bacillus thuringiensis PM25 ameliorates oxidative damage of salinity stress in maize via regulating growth, leaf pigments, antioxidant defense system, and stress responsive gene expression. Front. Plant Sci. 2022, 13, 921668. [Google Scholar] [CrossRef]
- Adnan, M.; Fahad, S.; Saleem, M.H.; Ali, B.; Mussart, M.; Ullah, R.; Arif, M.; Ahmad, M.; Shah, W.A.; Romman, M.; et al. Comparative efficacy of phosphorous supplements with phosphate solubilizing bacteria for optimizing wheat yield in calcareous soils. Sci. Rep. 2022, 12, 11997. [Google Scholar] [CrossRef]
- Rajkumar, M.; Sandhya, S.; Prasad, M.N.V.; Freitas, H. Perspectives of Plant-Associated Microbes in Heavy Metal Phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef]
- Aprill, W.; Sims, R.C. Evaluation of the Use of Prairie Grasses for Stimulating Polycyclic Aromatic Hydrocarbon Treatment in Soil. Chemosphere 1990, 20, 253–265. [Google Scholar] [CrossRef]
- Zhang, Y.; He, L.; Chen, Z.; Zhang, W.; Wang, Q.; Qian, M.; Sheng, X. Characterization of Lead-Resistant and ACC Deaminase-Producing Endophytic Bacteria and Their Potential in Promoting Lead Accumulation of Rape. J. Hazard. Mater. 2011, 186, 1720–1725. [Google Scholar] [CrossRef]
- Deng, H.; Ge, L.; Xu, T.; Wang, X.; Zhang, M.; Zhang, Y. Metabolic Properties of Carbon Sources of the Bacterial Community in Horizontal Subsurface Flow Constructed Wetlands in Eastern China. Fresenius Environ. Bull. 2010, 19, 2657–2664. [Google Scholar]
- Balsamo, R.A.; Kelly, W.J.; Satrio, J.A.; Ruiz-Felix, M.N.; Fetterman, M.; Wynn, R.; Hagel, K. Utilization of Grasses for Potential Biofuel Production and Phytoremediation of Heavy Metal Contaminated Soils. Int. J. Phytoremediation 2015, 17, 448–455. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Azeem, M.A.; Afridi, M.S.; Nadeem, M.; Ghazal, M.; Batool, T.; Qayyum, A.; Alatawi, A.; et al. Bacillus mycoides PM35 Reinforces Photosynthetic Efficiency, Antioxidant Defense, Expression of Stress-Responsive Genes, and Ameliorates the Effects of Salinity Stress in Maize. Life 2022, 12, 219. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Sumaira; Hafeez, A.; Afridi, M.S.; Khan, S.; Zaib-Un-Nisa; Ullah, I.; Amaral Júnior, A.T.; et al. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Amna Ali, B.; Azeem, M.A.; Qayyum, A.; Mustafa, G.; Ahmad, M.A.; Javed, M.T.; Chaudhary, H.J. Bio-Fabricated Silver Nanoparticles: A Sustainable Approach for Augmentation of Plant Growth and Pathogen Control. In Sustainable Agriculture Reviews 53; Springer: Cham, Switzerland, 2021; pp. 345–371. [Google Scholar]
- Dola, D.B.; Mannan, M.A.; Sarker, U.; Mamun, M.A.A.; Islam, T.; Ercisli, S.; Saleem, M.H.; Ali, B.; Pop, O.L.; Marc, R.A. Nano-iron oxide accelerates growth, yield, and quality of Glycine max seed in water deficits. Front. Plant Sci. 2022, 13, 992535. [Google Scholar] [CrossRef]
- Farooq, T.H.; Rafay, M.; Basit, H.; Shakoor, A.; Shabbir, R.; Riaz, M.U.; Ali, B.; Kumar, U.; Qureshi, K.A.; Jaremko, M. Morpho-physiological growth performance and phytoremediation capabilities of selected xerophyte grass species toward Cr and Pb stress. Front. Plant Sci. 2022, 13, 997120. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Ali, B.; Rasheed, R.; Ashraf, M.A.; Aziz, H.; Ercisli, S.; Riaz, S.; Elsharkawy, M.M.; Hussain, I.; et al. Impact of foliar application of syringic acid on tomato (Solanum lycopersicum L.) under heavy metal stress-insights into nutrient uptake, redox homeostasis, oxidative stress, and antioxidant defense. Front. Plant Sci. 2022, 13, 950120. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Yasin, G.; Mumtaz, S.; Qureshi, F.F.; Ali, B.; Ercisli, S.; Alhag, S.K.; Ahmed, A.E.; Vodnar, D.C.; et al. Individual and combinatorial effects of SNP and NaHS on morpho-physio-biochemical attributes and phytoextraction of chromium through Cr-stressed spinach (Spinacia oleracea L.). Front. Plant Sci. 2022, 13, 973740. [Google Scholar] [CrossRef]
- Shojaei, S.; Jafarpour, A.; Shojaei, S.; Gyasi-Agyei, Y.; Rodrigo-Comino, J. Heavy Metal Uptake by Plants from Wastewater of Different Pulp Concentrations and Contaminated Soils. J. Clean. Prod. 2021, 296, 126345. [Google Scholar] [CrossRef]
- Nishikimi, M.; Appaji Rao, N.; Yagi, K. The Occurrence of Superoxide Anion in the Reaction of Reduced Phenazine Methosulfate and Molecular Oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in Vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. ISBN 978-0-12-182005-3. [Google Scholar]
- Flohé, L.; Günzler, W.A. [12] Assays of Glutathione Peroxidase. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 114–120. ISBN 978-0-12-182005-3. [Google Scholar]
- Lobarzewski, J.; Ginalska, G. Industrial Use of Soluble or Immobilized Plant Peroxidases. Plant Peroxidase Newsl. 1995, 6, 3–7. [Google Scholar]
- Rajendran, M.; An, W.; Li, W.; Perumal, V.; Wu, C.; Sahi, S.V.; Sarkar, S.K. Chromium Detoxification Mechanism Induced Growth and Antioxidant Responses in Vetiver (Chrysopogon Zizanioides (L.) Roberty). J. Cent. South Univ. 2019, 26, 489–500. [Google Scholar] [CrossRef]
- Nawaz, H.; Ali, A.; Saleem, M.H.; Ameer, A.; Hafeez, A.; Alharbi, K.; Ezzat, A.; Khan, A.; Jamil, M.; Farid, G. Comparative effectiveness of EDTA and citric acid assisted phytoremediation of Ni contaminated soil by using canola (Brassica napus). Braz. J. Biol. 2022, 82, e261785. [Google Scholar] [CrossRef]
- Kumar, A.; Maiti, S.K. Effect of Organic Manures on the Growth of Cymbopogon Citratus and Chrysopogon Zizanioides for the Phytoremediation of Chromite-Asbestos Mine Waste: A Pot Scale Experiment. Int. J. Phytoremediation 2015, 17, 437–447. [Google Scholar] [CrossRef]
- Trivedi, A.K.; Ahmad, I. Effects of Chrysotile Asbestos Contaminated Soil on Crop Plants. Soil Sediment. Contam. Int. J. 2011, 20, 767–776. [Google Scholar] [CrossRef]
- Shrestha, P.; Bellitürk, K.; Görres, J. Phytoremediation of Heavy Metal-Contaminated Soil by Switchgrass: A Comparative Study Utilizing Different Composts and Coir Fiber on Pollution Remediation, Plant Productivity, and Nutrient Leaching. IJERPH 2019, 16, 1261. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-C.; Lai, H.-Y.; Juang, K.-W. Model Evaluation of Plant Metal Content and Biomass Yield for the Phytoextraction of Heavy Metals by Switchgrass. Ecotoxicol. Environ. Saf. 2012, 80, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Zainab, N.; Amna; Khan, A.A.; Azeem, M.A.; Ali, B.; Wang, T.; Shi, F.; Alghanem, S.M.; Hussain Munis, M.F.; Hashem, M.; et al. PGPR-Mediated Plant Growth Attributes and Metal Extraction Ability of Sesbania sesban L. in Industrially Contaminated Soils. Agronomy 2021, 11, 1820. [Google Scholar] [CrossRef]
- Chen, L.; Liu, J.; Hu, W.; Gao, J.; Yang, J. Vanadium in Soil-Plant System: Source, Fate, Toxicity, and Bioremediation. J. Hazard. Mater. 2021, 405, 124200. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.A.; Baken, S.; Gustafsson, J.P.; Hadialhejazi, G.; Smolders, E. Vanadium Bioavailability and Toxicity to Soil Microorganisms and Plants. Environ. Toxicol. Chem. 2013, 32, 2266–2273. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.G.; Boutin, C.; Knopper, L. Vanadium Pentoxide Phytotoxicity: Effects of Species Selection and Nutrient Concentration. Arch. Environ. Contam. Toxicol. 2013, 64, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, M.; Jia, Y.; Gou, M.; Zeyer, J. Toxicity of Vanadium in Soil on Soybean at Different Growth Stages. Environ. Pollut. 2017, 231, 48–58. [Google Scholar] [CrossRef]
- Guo, Y. Switchgrass Responses to Manganese Availability. Ph.D. Thesis, University of Missouri, Columbia, MO, USA, 2019. [Google Scholar]
- Chen, Z.; Yan, W.; Sun, L.; Tian, J.; Liao, H. Proteomic Analysis Reveals Growth Inhibition of Soybean Roots by Manganese Toxicity Is Associated with Alteration of Cell Wall Structure and Lignification. J. Proteom. 2016, 143, 151–160. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, X.Q.; Hu, Z.M.; Shao, J.F.; Che, J.; Chen, R.F.; Dong, X.Y.; Shen, R.F. Aluminium Alleviates Manganese Toxicity to Rice by Decreasing Root Symplastic Mn Uptake and Reducing Availability to Shoots of Mn Stored in Roots. Ann. Bot. 2015, 116, 237–246. [Google Scholar] [CrossRef]
- Neves, M.I.; Prajapati, D.H.; Parmar, S.; Aghajanzadeh, T.A.; Hawkesford, M.J.; De Kok, L.J. Manganese Toxicity Hardly Affects Sulfur Metabolism in Brassica Rapa. In Sulfur Metabolism in Higher Plants—Fundamental, Environmental and Agricultural Aspects; De Kok, L.J., Hawkesford, M.J., Haneklaus, S.H., Schnug, E., Eds.; Proceedings of the International Plant Sulfur Workshop; Springer International Publishing: Cham, Switzerland, 2017; pp. 155–162. ISBN 978-3-319-56525-5. [Google Scholar]
- Huang, Y.L.; Yang, S.; Long, G.X.; Zhao, Z.K.; Li, X.F.; Gu, M.H. Manganese Toxicity in Sugarcane Plantlets Grown on Acidic Soils of Southern China. PLoS ONE 2016, 11, e0148956. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Islam, F.; Ali, B.; Najeeb, U.; Mao, B.; Gill, R.A.; Yan, G.; Siddique, K.H.M.; Zhou, W. Arsenic Toxicity in Plants: Cellular and Molecular Mechanisms of Its Transport and Metabolism. Environ. Exp. Bot. 2016, 132, 42–52. [Google Scholar] [CrossRef]
- Singh, S.; Sounderajan, S.; Kumar, K.; Fulzele, D.P. Investigation of Arsenic Accumulation and Biochemical Response of in Vitro Developed Vetiveria Zizanoides Plants. Ecotoxicol. Environ. Saf. 2017, 145, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Balal, R.M.; Khan, N.; Zotarelli, L.; Liu, G.D.; Sarkhosh, A.; Fernández-Zapata, J.C.; Martínez Nicolás, J.J.; Garcia-Sanchez, F. Selenium Impedes Cadmium and Arsenic Toxicity in Potato by Modulating Carbohydrate and Nitrogen Metabolism. Ecotoxicol. Environ. Saf. 2019, 180, 588–599. [Google Scholar] [CrossRef]
- Mohd, S.; Shukla, J.; Kushwaha, A.S.; Mandrah, K.; Shankar, J.; Arjaria, N.; Saxena, P.N.; Narayan, R.; Roy, S.K.; Kumar, M. Endophytic Fungi Piriformospora Indica Mediated Protection of Host from Arsenic Toxicity. Front. Microbiol. 2017, 8, 754. [Google Scholar] [CrossRef] [Green Version]
- Mawia, A.M.; Hui, S.; Zhou, L.; Li, H.; Tabassum, J.; Lai, C.; Wang, J.; Shao, G.; Wei, X.; Tang, S.; et al. Inorganic Arsenic Toxicity and Alleviation Strategies in Rice. J. Hazard. Mater. 2021, 408, 124751. [Google Scholar] [CrossRef]
- Armendariz, A.L.; Talano, M.A.; Travaglia, C.; Reinoso, H.; Wevar Oller, A.L.; Agostini, E. Arsenic Toxicity in Soybean Seedlings and Their Attenuation Mechanisms. Plant Physiol. Biochem. 2016, 98, 119–127. [Google Scholar] [CrossRef]
- Dradrach, A.; Karczewska, A.; Szopka, K. Arsenic Accumulation by Red Fescue (Festuca Rubra) Growing in Mine Affected Soils—Findings from the Field and Greenhouse Studies. Chemosphere 2020, 248, 126045. [Google Scholar] [CrossRef]
- Yang, F.; Xie, S.; Wei, C.; Liu, J.; Zhang, H.; Chen, T.; Zhang, J. Arsenic Characteristics in the Terrestrial Environment in the Vicinity of the Shimen Realgar Mine, China. Sci. Total Environ. 2018, 626, 77–86. [Google Scholar] [CrossRef]
- Dradrach, A.; Karczewska, A.; Szopka, K.; Lewińska, K. Accumulation of Arsenic by Plants Growing in the Sites Strongly Contaminated by Historical Mining in the Sudetes Region of Poland. IJERPH 2020, 17, 3342. [Google Scholar] [CrossRef]
- Dradrach, A.; Karczewska, A.; Szopka, K. Arsenic Uptake by Two Tolerant Grass Species: Holcus Lanatus and Agrostis Capillaris Growing in Soils Contaminated by Historical Mining. Plants 2020, 9, 980. [Google Scholar] [CrossRef]
- Sinha, B.; Bhattacharyya, K. Arsenic Toxicity in Rice with Special Reference to Speciation in Indian Grain and Its Implication on Human Health. J. Sci. Food Agric. 2015, 95, 1435–1444. [Google Scholar] [CrossRef]
- Prommarach, T.; Pholsen, S.; Shivaraju, H.P.; Chareonsudjai, P. Growth and Biosorption of Purple Guinea and Ruzi Grasses in Arsenic Contaminated Soils. Environ. Monit. Assess. 2022, 194, 85. [Google Scholar] [CrossRef]
- Sultana, R.; Kobayashi, K.; Kim, K.-H. Comparison of Arsenic Uptake Ability of Barnyard Grass and Rice Species for Arsenic Phytoremediation. Environ. Monit Assess 2015, 187, 4101. [Google Scholar] [CrossRef]
- Sleimi, N.; Kouki, R.; Hadj Ammar, M.; Ferreira, R.; Pérez-Clemente, R. Barium Effect on Germination, Plant Growth, and Antioxidant Enzymes in Cucumis Sativus L. Plants. Food Sci. Nutr. 2021, 9, 2086–2094. [Google Scholar] [CrossRef]
- de Souza Cardoso, A.A.; Monteiro, F.A. Sulfur Supply Reduces Barium Toxicity in Tanzania Guinea Grass (Panicum Maximum) by Inducing Antioxidant Enzymes and Proline Metabolism. Ecotoxicol. Environ. Saf. 2021, 208, 111643. [Google Scholar] [CrossRef]
- Myrvang, M.B.; Gjengedal, E.; Heim, M.; Krogstad, T.; Almås, Å.R. Geochemistry of Barium in Soils Supplied with Carbonatite Rock Powder and Barium Uptake to Plants. Appl. Geochem. 2016, 75, 1–8. [Google Scholar] [CrossRef]
- Sungur, A.; Gur, E.; Everest, T.; Soylak, M.; Ozcan, H. Assessment of Relationship Between Geochemical Fractions of Barium in Soil of Cherry Orchards and Plant Barium Uptake and Determination by Inductively Coupled Plasma Optical Emission Spectrometry. At. Spectrosc. 2019, 40, 173–178. [Google Scholar] [CrossRef]
- de Castro Ribeiro, P.R.C.; Viana, D.G.; Pires, F.R.; Egreja Filho, F.B.; Bonomo, R.; Cargnelutti Filho, A.; Martins, L.F.; Cruz, L.B.S.; Nascimento, M.C.P. Selection of Plants for Phytoremediation of Barium-Polluted Flooded Soils. Chemosphere 2018, 206, 522–530. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.F.M.; Viana, D.G.; Pires, F.R.; Egreja Filho, F.B.; Bonomo, R.; Martins, L.F.; Cruz, L.B.S.; Nascimento, M.C.P.; Cargnelutti Filho, A.; da Rocha Júnior, P.R. Phytoremediation of Barium-Affected Flooded Soils Using Single and Intercropping Cultivation of Aquatic Macrophytes. Chemosphere 2019, 214, 10–16. [Google Scholar] [CrossRef]
- Myrvang, M.B.; Bleken, M.A.; Krogstad, T.; Heim, M.; Gjengedal, E. Can Liming Reduce Barium Uptake by Agricultural Plants Grown on Sandy Soil? J. Plant Nutr. Soil Sci. 2016, 179, 557–565. [Google Scholar] [CrossRef]
- Lu, Q.; Xu, X.; Liang, L.; Xu, Z.; Shang, L.; Guo, J.; Xiao, D.; Qiu, G. Barium Concentration, Phytoavailability, and Risk Assessment in Soil-Rice Systems from an Active Barium Mining Region. Appl. Geochem. 2019, 106, 142–148. [Google Scholar] [CrossRef]
- Mehmood, S.; Khatoon, Z.; Amna Ahmad, I.; Muneer, M.A.; Kamran, M.A.; Ali, J.; Ali, B.; Chaudhary, H.J.; Munis, M.F. Bacillus sp. PM31 harboring various plant growth-promoting activities regulates Fusarium dry rot and wilt tolerance in potato. Arch. Agron. Soil Sci. 2021, 2021, 1971654. [Google Scholar] [CrossRef]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef]
- Afridi, M.S.; Javed, M.A.; Ali, S.; De Medeiros, F.H.V.; Ali, B.; Salam, A.; Sumaira Marc, R.A.; Alkhalifah, D.H.M.; Selim, S.; Santoyo, G. New opportunities in plant microbiome engineering for increasing agricultural sustainability under stressful conditions. Front. Plant Sci. 2022, 13, 899464. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).