



Airborne Pollen, Allergens, and Proteins: A Comparative Study of Three Sampling Methods

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Pollen Sampling
2.3. Particulate Matter Sampling
- One low-volume Cyclone Burkard sampler (CS) (Burkard Manufacturing Co, Rickmansworth, UK), which is a continuous, wind-oriented volumetric sampler using single reverse-flow miniature Cyclone technology to collect airborne particles. Air was aspired with a speed of 16.5 L/min, and particles were collected directly into a 1.5 mL vial, which was substituted every 24 h of sampling.
- Two five-stage Berner impactors (BIs) (Hauke GmbH, Wien, Austria), which are volumetric cascade samplers that allow the separation of the particles collected according to their aerodynamic diameter [52,53]. Both BIs had cut-off sizes (d50, the square root value of Stokes number) [54] at 0.14, 0.42, 1.2, 3.5, and 10 μm in aerodynamic diameter and were operated with a constant air flow rate of 80 L/min. One BI (BI1) was employed to collect samples for protein analysis, using Tedlar foils as sampling surfaces on each stage. The other BI (BI2) was loaded with aluminium foils for carbonaceous aerosol analysis, and with Tedlar foils cut in half and placed on top of the aluminium foils, covering 50% of their surface, for ion-chromatographic analysis [55]. Samples were collected every 24 h.
- One high-volume Dichotomous air sampler (DS) (Model 310 Universal Air Samples (UAS), MSP Corporation, Shoreview, MN, USA), that is a volumetric impactor able to separate the collected particles into two groups, “coarse” and “fine”, based on an aerodynamic diameter threshold [56]. In this study, the aerodynamic diameter ranged from 1 to 10 μm for the coarse fraction and from 0.1 to 1 μm for the fine fraction. The sampler was operated with a constant air flow of 300 L/min, and particles were collected on quartz fibre filters. Samples were collected every 24 h.
2.4. Extraction and Quantification of Airborne Proteins
- Samples collected by the CS in 1.5 mL vials were centrifuged for 1 min at 18,000× g. Total proteins were then extracted at room temperature for 2 h with 120 µL of phosphate buffer (50 mM pH 7.0) supplemented with 150 mM NaCl, 3 mM EDTA, 0.005% Tween 20, and 125 mM ammonium bicarbonate [28,40,57,58]. The extract was separated by centrifugation at 2000× g for 10 min, and then the supernatant was lyophilised and resuspended in 100 μL of 50 mM Tris-HCl, pH 7.5.
- Samples collected on Tedlar supports by BI1 were resuspended in sterile bi-distilled water supplemented with a protease inhibitor cocktail (Merck KGaA, Darmstadt, Germania). Tedlar samples were sonicated twice on ice and stirred overnight at 4 °C. Tedlar foils were removed and the extract was lyophilised and re-suspended in 100 μL of 50 mM Tris-HCl, pH 7.5.
- Quartz fibre filters used in the DS were submerged in a solution that allowed the maximum recovery of proteins (2 mM Tris-HCl pH 7.8 + 0.0001% Tween + protease inhibitor cocktail). Samples were sonicated twice on ice and stirred overnight at 4 °C. Afterwards, filter material and debris were removed from the solution by centrifugation for 10 min, 5000× g, at 4 °C. Samples were lyophilised and resuspended in 100 μL of 50 mM Tris-HCl, pH 7.5.
2.5. Meteorological Data
2.6. Chemical analysis of PM
2.7. Statistical Analysis
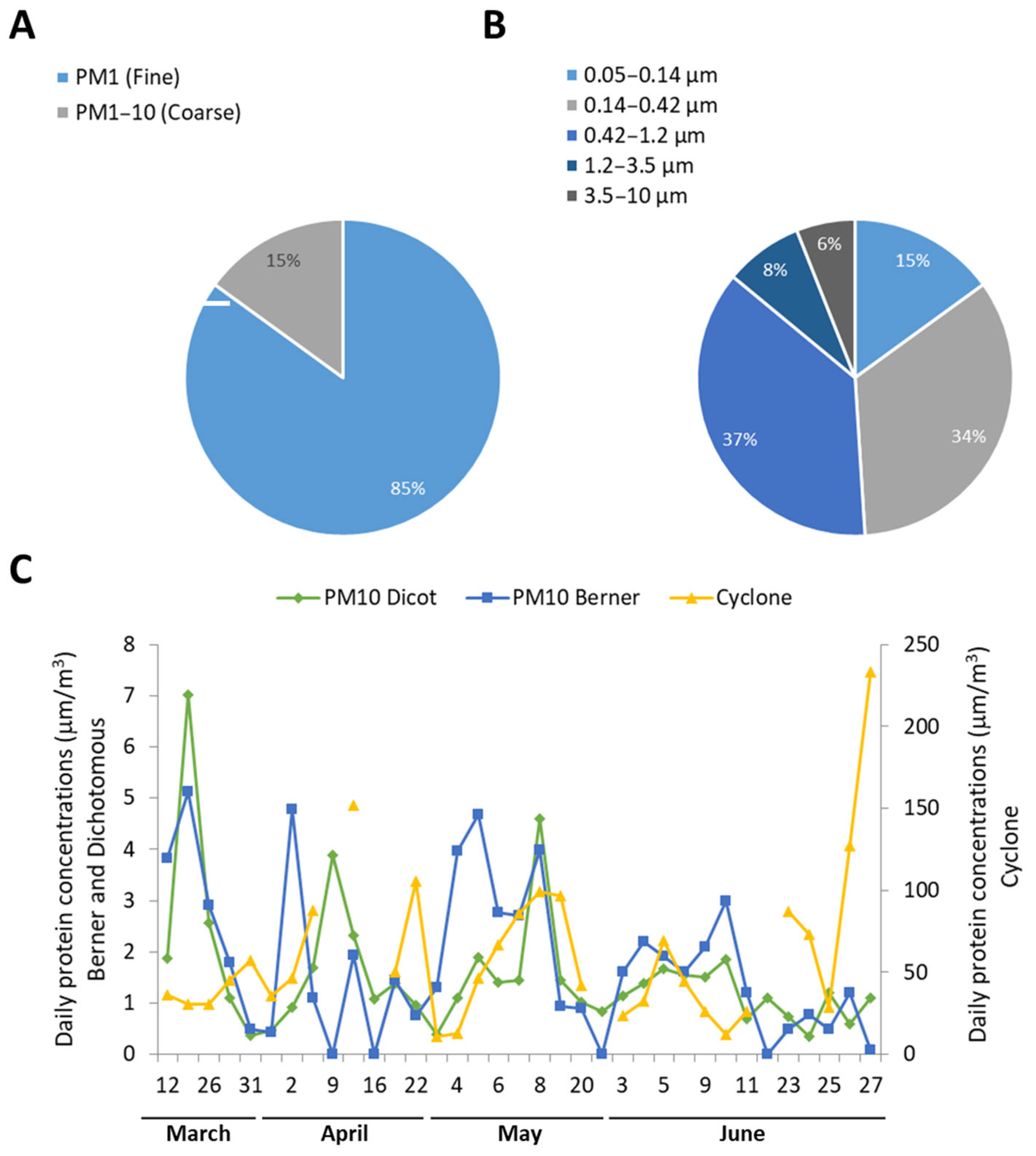
3. Results
3.1. Airborne Proteins and Allergens Concentrations
3.2. Relationships between Biological, Chemical, and Meteorological Parameters
4. Discussion
4.1. Different Samplers Have Different Protein Collection Efficiencies
4.2. Airborne Pollen and Airborne Protein Levels Are not Representative of Aeroallergen Concentrations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- D’Amato, G.; Vitale, C.; Sanduzzi, A.; Molino, A.; Vatrella, A.; D’Amato, M. Allergenic Pollen and Pollen Allergy in Europe. Allergy 2017, 62, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Migueres, M.; Dávila, I.; Frati, F.; Azpeitia, A.; Jeanpetit, Y.; Lhéritier-Barrand, M.; Incorvaia, C.; Ciprandi, G. Types of sensitization to aeroallergens: Definitions, prevalences and impact on the diagnosis and treatment of allergic respiratory disease. Clin. Transl. Allergy 2014, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Ozdoganoglu, T.; Songu, M. The burden of allergic rhinitis and asthma. Ther. Adv. Respir. Dis. 2012, 6, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Grammer, L.C. Chapter 1: An overview of allergens. Allergy Asthma Proc. 2012, 33 (Suppl. S1), 2–5. [Google Scholar] [CrossRef]
- Zuberbier, T.; Lötvall, J.; Simoens, S.; Subramanian, S.V.; Church, M.K. Economic burden of inadequate management of allergic diseases in the European Union: A GA2LEN review. Allergy 2014, 69, 1275–1279. [Google Scholar] [CrossRef]
- Bousquet, J.; Khaltaev, N.; Cruz, A.A.; Denburg, J.; Fokkens, W.J.; Togias, A.; Zuberbier, T.; Baena-Cagnani, C.E.; Canonica, G.W.; Van Weel, C.; et al. Allergic Rhinitis and its Impact on Asthma (ARIA) 2008. Allergy 2008, 63 (Suppl. S86), 8–160. [Google Scholar] [CrossRef]
- Levetin, E. Methods for aeroallergen sampling. Curr. Allergy Asthma Rep. 2004, 4, 376–383. [Google Scholar] [CrossRef]
- Suanno, C.; Aloisi, I.; Fernández-González, D.; Del Duca, S. Monitoring techniques for pollen allergy risk assessment. Environ. Res. 2021, 197, 111109. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.K.; Swanson, M.C.; Reed, C.E.; Yunginger, J.W. Airborne ragweed allergens: Association with various particle sizes and short ragweed plant parts. J. Allergy Clin. Immunol. 1984, 74, 687–693. [Google Scholar] [CrossRef]
- Cecchi, L. From pollen count to pollen potency: The molecular era of aerobiology. Eur. Respir. J. 2013, 42, 898–900. [Google Scholar] [CrossRef]
- Mothes, N.; Horak, F.; Valenta, R. Transition from a Botanical to a Molecular Classification in Tree Pollen Allergy: Implications for Diagnosis and Therapy. Int. Arch. Allergy Immunol. 2004, 135, 357–373. [Google Scholar] [CrossRef]
- Behrendt, H.; Becker, W.M.; Fritzsche, C.; Sliwa-Tomczok, W.; Tomczok, J.; Friedrichs, K.H.; Ring, J. Air Pollution and Allergy: Experimental Studies on Modulation of Allergen Release from Pollen by Air Pollutants. Int. Arch. Allergy Immunol. 1997, 113, 69–74. [Google Scholar] [CrossRef]
- D’Amato, G. Airborne paucimicronic allergen-carrying particles and seasonal respiratory allergy. Allergy 2001, 56, 1109–1111. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Liccardi, G.; Popov, T.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy 2007, 62, 976–990. [Google Scholar] [CrossRef]
- De Linares, C.; de la Guardia, C.D.; Lugilde, D.N.; Alba, F. Airborne Study of Grass Allergen (Lol p 1) in Different-Sized Particles. Int. Arch. Allergy Immunol. 2010, 152, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Grote, M.; Vrtala, S.; Niederberger, V.; Valenta, R.; Reichelt, R. Expulsion of allergen-containing materials from hydrated rye grass (Lolium perenne) pollen revealed by using immunogold field emission scanning and transmission electron microscopy. J. Allergy Clin. Immunol. 2000, 105, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Grote, M.; Valenta, R.; Reichelt, R. Abortive pollen germination: A mechanism of allergen release in birch, alder, and hazel revealed by immunogold electron microscopy. J. Allergy Clin. Immunol. 2003, 111, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Prado, N.; De Linares, C.; Sanz, M.L.; Gamboa, P.; Villalba, M.; Rodríguez, R.; Batanero, E. Pollensomes as Natural Vehicles for Pollen Allergens. J. Immunol. 2015, 195, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Després, V.R.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.S.; Buryak, G.; Fröhlich-Nowoisky, J.; Elbert, W.; Andreae, M.O.; Pöschl, U.; et al. Primary biological aerosol particles in the atmosphere: A review. Tellus B Chem. Phys. Meteorol. 2012, 64, 15598. [Google Scholar] [CrossRef]
- Galan, C.; Antunes, C.; Brandao, R.; Torres, C.; Garcia-Mozo, H.; Caeiro, E.; Ferro, R.; Prank, M.; Sofiev, M.; Albertini, R.; et al. Airborne olive pollen counts are not representative of exposure to the major olive allergen Ole e 1. Allergy 2013, 68, 809–812. [Google Scholar] [CrossRef] [Green Version]
- Harun, N.-S.; Lachapelle, P.; Douglass, J. Thunderstorm-triggered asthma: What we know so far. J. Asthma Allergy 2019, 12, 101–108. [Google Scholar] [CrossRef]
- Kang, I.G.; Ju, Y.H.; Jung, J.H.; Ko, K.P.; Oh, D.K.; Kim, J.H.; Lim, D.H.; Kim, Y.H.; Jang, T.Y.; Kim, S.T. The Effect of PM10 on Allergy Symptoms in Allergic Rhinitis Patients during Spring Season. Int. J. Environ. Res. Public Health 2015, 12, 735–745. [Google Scholar] [CrossRef]
- Taylor, P.E.; Flagan, R.C.; Valenta, R.; Glovsky, M.M. Release of allergens as respirable aerosols: A link between grass pollen and asthma. J. Allergy Clin. Immunol. 2002, 109, 51–56. [Google Scholar] [CrossRef]
- Pöschl, U. Atmospheric Aerosols: Composition, Transformation, Climate and Health Effects. Angew. Chem. Int. Ed. 2005, 44, 7520–7540. [Google Scholar] [CrossRef]
- Shiraiwa, M.; Selzle, K.; Pöschl, U. Hazardous components and health effects of atmospheric aerosol particles: Reactive oxygen species, soot, polycyclic aromatic compounds and allergenic proteins. Free Radic. Res. 2012, 46, 927–939. [Google Scholar] [CrossRef]
- Di Filippo, P.; Pomata, D.; Riccardi, C.; Buiarelli, F.; Gallo, V.; Quaranta, A. Free and combined amino acids in size-segregated atmospheric aerosol samples. Atmos. Environ. 2014, 98, 179–189. [Google Scholar] [CrossRef]
- Poruthoor, S.K.; Dasgupta, A.P.K.; Genfa, Z. Indoor Air Pollution and Sick Building Syndrome. Monitoring Aerosol Protein as a Measure of Bioaerosols. Environ. Sci. Technol. 1998, 32, 1147–1152. [Google Scholar] [CrossRef]
- Aloisi, I.; Del Duca, S.; De Nuntiis, P.; Maray, A.M.V.; Mandrioli, P.; Gutiérrez, P.; Fernández-González, D. Behavior of profilins in the atmosphere and in vitro, and their relationship with the performance of airborne pollen. Atmos. Environ. 2018, 178, 231–241. [Google Scholar] [CrossRef]
- Fernández-González, D.; Maray, A.M.V.; Parrado, Z.G.; Barrera, R.M.V.; Gutiérrez, P.; De Nuntiis, P.; Mandrioli, P. Are the profilins an important component in the atmosphere? Ole e 2-like panallergen. Aerobiologia 2019, 35, 165–175. [Google Scholar] [CrossRef]
- García-Sánchez, J.; Trigo, M.D.M.; Recio, M. Extraction and quantification of Ole e 1 from atmospheric air samples: An optimized protocol. Chemosphere 2019, 225, 490–496. [Google Scholar] [CrossRef]
- Plaza, M.P.; Alcázar, P.; Hernández-Ceballos, M.A.; Galán, C. Mismatch in aeroallergens and airborne grass pollen concentrations. Atmos. Environ. 2016, 144, 361–369. [Google Scholar] [CrossRef]
- Plaza, M.P.; Alcázar, P.; Velasco-Jiménez, M.J.; Galán, C. Aeroallergens: A comparative study of two monitoring methods. Aerobiologia 2017, 33, 363–373. [Google Scholar] [CrossRef]
- Schäppi, G.F.; Suphioglu, C.; Taylor, P.E.; Knox, R.B. Concentrations of the major birch tree allergen Bet v 1 in pollen and respirable fine particles in the atmosphere. J. Allergy Clin. Immunol. 1997, 100, 656–661. [Google Scholar] [CrossRef]
- Spieksma, F.T.M.; Kramps, J.A.; Plomp, A.; Koerten, H.K. Grass-pollen allergen carried by the smaller micronic aerosol fraction. Grana 1991, 30, 98–101. [Google Scholar] [CrossRef]
- Brennan, G.L.; Potter, C.; de Vere, N.; Griffith, G.W.; Skjøth, C.A.; Osborne, N.J.; Wheeler, B.; McInnes, R.N.; Clewlow, Y.; Barber, A.; et al. Temperate airborne grass pollen defined by spatio-temporal shifts in community composition. Nat. Ecol. Evol. 2019, 3, 750–754. [Google Scholar] [CrossRef]
- Cariñanos, P.; Casares-Porcel, M.; de la Guardia, C.D.; Aira, M.J.; Belmonte, J.; Boi, M.; Elvira-Rendueles, B.; De Linares, C.; Fernández-Rodriguez, S.; Maya-Manzano, J.M.; et al. Assessing allergenicity in urban parks: A nature-based solution to reduce the impact on public health. Environ. Res. 2017, 155, 219–227. [Google Scholar] [CrossRef]
- Fernández-González, D.; González-Parrado, Z.; Maray, A.M.V.; Valencia-Barrera, R.M.; Camazón-Izquierdo, B.; De Nuntiis, P.; Mandrioli, P. Platanus pollen allergen, Pla a 1: Quantification in the atmosphere and influence on a sensitizing population. Clin. Exp. Allergy 2010, 40, 1701–1708. [Google Scholar] [CrossRef]
- Fernández-González, D.; Rajo, F.J.R.; Parrado, Z.G.; Barrera, R.M.V.; Jato, V.; Grau, S.M. Differences in atmospheric emissions of Poaceae pollen and Lol p 1 allergen. Aerobiologia 2011, 27, 301–309. [Google Scholar] [CrossRef]
- Fernández-González, M.; Álvarez-López, S.; González-Fernández, E.; Aira, M.J.; Rodríguez-Rajo, F.J. Cross-reactivity between the Betulaceae family and fallout in the real atmospheric aeroallergen load. Sci. Total Environ. 2020, 715, 136861. [Google Scholar] [CrossRef]
- Moreno-Grau, S.; Elvira-Rendueles, B.; Moreno, J.; García-Sánchez, A.; Vergara, N.; Asturias, J.A.; Arilla, M.; Ibarrola, I.; Seoane-Camba, J.A.; Suárez-Cervera, M. Correlation between Olea europaea and Parietaria judaica pollen counts and quantification of their major allergens Ole e 1 and Par j 1-Par j 2. Ann. Allergy Asthma Immunol. 2006, 96, 858–864. [Google Scholar] [CrossRef]
- Rodríguez-Rajo, F.J.; Jato, V.; González-Parrado, Z.; Elvira-Rendueles, B.; Moreno-Grau, S.; Vega-Maray, A.; Fernández-González, D.; Asturias, J.; Suárez-Cervera, M. The combination of airborne pollen and allergen quantification to reliably assess the real pollinosis risk in different bioclimatic areas. Aerobiologia 2011, 27, 1–12. [Google Scholar] [CrossRef]
- De Linares, C.; Plaza, M.P.; Valle, A.M.; Alcázar, P.; de la Guardia, C.D.; Galán, C. Airborne Cupressaceae Pollen and Its Major Allergen, Cup a 1, in Urban Green Areas of Southern Iberian Peninsula. Forests 2021, 12, 254. [Google Scholar] [CrossRef]
- Buters, J.T.M.; Weichenmeier, I.; Ochs, S.; Pusch, G.; Kreyling, W.; Boere, A.J.F.; Schober, W.; Behrendt, H. The allergen Bet v 1 in fractions of ambient air deviates from birch pollen counts. Allergy 2010, 65, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Buters, J.T.M.; Thibaudon, M.; Smith, M.; Kennedy, R.; Rantio-Lehtimäki, A.; Albertini, R.; Reese, G.; Weber, B.; Galan, C.; Brandao, R.; et al. Release of Bet v 1 from birch pollen from 5 European countries. Results from the HIALINE study. Atmos. Environ. 2012, 55, 496–505. [Google Scholar] [CrossRef]
- Buters, J.; Prank, M.; Sofiev, M.; Pusch, G.; Albertini, R.; Annesi-Maesano, I.; Antunes, C.; Behrendt, H.; Berger, U.; Brandao, R.; et al. Variation of the group 5 grass pollen allergen content of airborne pollen in relation to geographic location and time in season. J. Allergy Clin. Immunol. 2015, 136, 87–95.e6. [Google Scholar] [CrossRef]
- Burbach, G.J.; Heinzerling, L.M.; Edenharter, G.; Bachert, C.; Bindslev-Jensen, C.; Bonini, S.; Bousquet, J.; Bousquet-Rouanet, L.; Bousquet, P.J.; Bresciani, M.; et al. GA 2 LEN skin test study II: Clinical relevance of inhalant allergen sensitizations in Europe. Allergy 2009, 64, 1507–1515. [Google Scholar] [CrossRef]
- Singh, M.B.; Bhalla, P.L. Hypoallergenic derivatives of major grass pollen allergens for allergy vaccination. Immunol. Cell Biol. 2003, 81, 86–91. [Google Scholar] [CrossRef]
- Sekerkova, A.; Polackova, M.; Striz, I. Detection of Phl p 1, Phl p 5, Phl p 7 and Phl p 12 Specific IgE Antibodies in the Sera of Children and Adult Patients Allergic to Phleum Pollen. Allergol. Int. 2012, 61, 339–346. [Google Scholar] [CrossRef]
- Schäppi, G.F.; Monn, C.; Wüthrich, B.; Wanner, H.-U. Direct Determination of Allergens in Ambient Aerosols: Methodological Aspects. Int. Arch. Allergy Immunol. 1996, 110, 364–370. [Google Scholar] [CrossRef]
- Plaza, M.P.; Alcázar, P.; Oteros, J.; Galán, C. Atmospheric pollutants and their association with olive and grass aeroallergen concentrations in Córdoba (Spain). Environ. Sci. Pollut. Res. 2020, 27, 45447–45459. [Google Scholar] [CrossRef]
- EN16868 Ambient Air—Sampling and Analysis of Airbone Pollen Grains and Fungal Spores for Networks Related to Allergy—Volumetric Hirst Method. 2019. Vol. CEN/TS 168. Available online: https://standards.iteh.ai/catalog/standards/cen/5f1349aa-f4cc-430a-978e-3044737e3f28/en-16868-2019 (accessed on 7 October 2022).
- Štefancová, L.; Schwarz, J.; Mäkelä, T.; Hillamo, R.; Smolík, J. Comprehensive Characterization of Original 10-Stage and 7-Stage Modified Berner Type Impactors. Aerosol Sci. Technol. 2011, 45, 88–100. [Google Scholar] [CrossRef]
- Berner, A.; Lürzer, C.; Pohl, F.; Preining, O.; Wagner, P. The size distribution of the urban aerosol in Vienna. Sci. Total Environ. 1979, 13, 245–261. [Google Scholar] [CrossRef]
- Hinds, W.C. Aerosol Technology: Properties, Behavior, and Measurement of Airborne Particles, 2nd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1999. [Google Scholar]
- Matta, E.; Facchini, M.C.; Decesari, S.; Mircea, M.; Cavalli, F.; Fuzzi, S.; Putaud, J.-P.; Dell’Acqua, A. Mass closure on the chemical species in size-segregated atmospheric aerosol collected in an urban area of the Po Valley, Italy. Atmos. Chem. Phys. 2003, 3, 623–637. [Google Scholar] [CrossRef]
- Poor, N.; Clark, T.; Nye, L.; Tamanini, T.; Tate, K.; Stevens, R.; Atkeson, T. Field performance of dichotomous sequential PM air samplers. Atmos. Environ. 2002, 36, 3289–3298. [Google Scholar] [CrossRef]
- Jato, V.; Rodríguez-Rajo, F.J.; González-Parrado, Z.; Rendueles, B.E.; Moreno-Grau, S.; Maray, A.M.V.; González, D.F.; Asturias, J.A.; Suárez-Cervera, M. Detection of airborne Par j 1 and Par j 2 allergens in relation to Urticaceae pollen counts in different bioclimatic areas. Ann. Allergy Asthma Immunol. 2010, 105, 50–56. [Google Scholar] [CrossRef]
- Takahashi, Y.; Ohashi, T.; Nagoya, T.; Sakaguchi, M.; Yasueda, H.; Nitta, H. Possibility of real-time measurement of an airborne Cryptomeria japonica pollen allergen based on the principle of surface plasmon resonance. Aerobiologia 2001, 17, 313–318. [Google Scholar] [CrossRef]
- Arilla, M.C.; Ibarrola, I.; Eraso, E.; Aguirre, M.; Martínez, A.; Asturias, J.A. Quantification in mass units of group 1 grass allergens by a monoclonal antibody-based sandwich ELISA. Clin. Exp. Allergy 2001, 31, 1271–1278. [Google Scholar] [CrossRef]
- Sandrini, S.; van Pinxteren, D.; Giulianelli, L.; Herrmann, H.; Poulain, L.; Facchini, M.C.; Gilardoni, S.; Rinaldi, M.; Paglione, M.; Turpin, B.J.; et al. Size-resolved aerosol composition at an urban and a rural site in the Po Valley in summertime: Implications for secondary aerosol formation. Atmos. Chem. Phys. 2016, 16, 10879–10897. [Google Scholar] [CrossRef]
- Gelencsér, A.; Mészáros, T.; Blazsó, M.; Kiss, G.Y.; Krivácsy, Z.; Molnár, A.; Mészáros, E. Structural Characterisation of Organic Matter in Fine Tropospheric Aerosol by Pyrolysis-Gas Chromatography-Mass Spectrometry. J. Atmos. Chem. 2000, 37, 173–183. [Google Scholar] [CrossRef]
- Decesari, S.; Facchini, M.C.; Carbone, C.; Giulianelli, L.; Rinaldi, M.; Finessi, E.; Fuzzi, S.; Marinoni, A.; Cristofanelli, P.; Duchi, R.; et al. Chemical composition of PM10 and PM1 at the high-altitude Himalayan station Nepal Climate Observatory-Pyramid (NCO-P) (5079 m a.s.l.). Atmos. Chem. Phys. 2010, 10, 4583–4596. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 16 June 2022).
- Belis, C.A.; Karagulian, F.; Larsen, B.R.; Hopke, P.K. Critical review and meta-analysis of ambient particulate matter source apportionment using receptor models in Europe. Atmos. Environ. 2013, 69, 94–108. [Google Scholar] [CrossRef]
- Tørseth, K.; Hanssen, J.E.; Semb, A. Temporal and spatial variations of airborne Mg, Cl, Na, Ca and K in rural areas of Norway. Sci. Total Environ. 1999, 234, 75–85. [Google Scholar] [CrossRef]
- Calvo, A.I.; Baumgardner, D.; Castro, A.; Fernández-González, D.; Vega-Maray, A.M.; Valencia-Barrera, R.M.; Oduber, F.; Blanco-Alegre, C.; Fraile, R. Daily behavior of urban Fluorescing Aerosol Particles in northwest Spain. Atmos. Environ. 2018, 184, 262–277. [Google Scholar] [CrossRef]
- Matos, J.T.V.; Duarte, R.M.B.O.; Duarte, A.C. Challenges in the identification and characterization of free amino acids and proteinaceous compounds in atmospheric aerosols: A critical review. TrAC Trends Anal. Chem. 2016, 75, 97–107. [Google Scholar] [CrossRef]
- Kang, H.; Xie, Z.; Hu, Q. Ambient protein concentration in PM10 in Hefei, central China. Atmos. Environ. 2012, 54, 73–79. [Google Scholar] [CrossRef]
- Grewling, Ł.; Bogawski, P.; Kostecki, Ł.; Nowak, M.; Szymańska, A.; Frątczak, A. Atmospheric exposure to the major Artemisia pollen allergen (Art v 1): Seasonality, impact of weather, and clinical implications. Sci. Total Environ. 2020, 713, 136611. [Google Scholar] [CrossRef]
- Taylor, P.E.; Jonsson, H. Thunderstorm asthma. Curr. Allergy Asthma Rep. 2004, 4, 409–413. [Google Scholar] [CrossRef]
- Hughes, D.D.; Mampage, C.B.A.; Jones, L.M.; Liu, Z.; Stone, E.A. Characterization of Atmospheric Pollen Fragments during Springtime Thunderstorms. Environ. Sci. Technol. Lett. 2020, 7, 409–414. [Google Scholar] [CrossRef]
- Ruggiero, F.; Bedini, G. Systematic and morphologic survey of orbicules in allergenic angiosperms. Aerobiologia 2018, 34, 405–422. [Google Scholar] [CrossRef]
- Matthiesen, F.; Løwenstein, H. Group V allergens in grass pollens. II. Investigation of group V allergens in pollens from 10 grasses. Clin. Exp. Allergy 1991, 21, 309–320. [Google Scholar] [CrossRef]
- Namork, E.; Johansen, B.V.; Løvik, M. Detection of allergens adsorbed to ambient air particles collected in four European cities. Toxicol. Lett. 2006, 165, 71–78. [Google Scholar] [CrossRef]
- Knox, R.B.; Suphioglu, C.; Taylor, P.; Desai, R.; Watson, H.C.; Peng, J.L.; Bursill, L.A. Major grass pollen allergen Lol p 1 binds to diesel exhaust particles: Implications for asthma and air pollution. Clin. Exp. Allergy 1997, 27, 246–251. [Google Scholar] [CrossRef]
- Ormstad, H. Suspended particulate matter in indoor air: Adjuvants and allergen carriers. Toxicology 2000, 152, 53–68. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dichotomous Sampler | Berner Impactor | Cyclone Sampler | ||||||
|---|---|---|---|---|---|---|---|---|
| Particle cut-off, μm (Ø) | 0.1–1 (Fine) | 1–10 (Coarse) | 0.14 | 0.42 | 1.2 | 3.5 | 10 | |
| Average protein concentration (μg/m3) | 1.28 | 0.22 | 0.31 | 0.68 | 0.74 | 0.16 | 0.11 | 61.97 |
| Minimum protein concentration (μg/m3) | 0.03 | 0.03 | 0.02 | 0.06 | 0.05 | 0.02 | 0.01 | 10.89 |
| Maximum protein concentration (μg/m3) | 6.72 | 0.72 | 1.27 | 1.88 | 1.88 | 0.60 | 0.47 | 232.95 |
| Median protein concentration (μg/m3) | 0.95 | 0.19 | 0.23 | 0.59 | 0.67 | 0.08 | 0.06 | 46.41 |
| Dichotomous Sampler | Berner Impactor | Cyclone Sampler | |||||
|---|---|---|---|---|---|---|---|
| Fraction | PM1 | PM1–10 | PM10 | PM1.2 | PM1.2–10 | PM10 | |
| Average Phl p 5 concentrations (pg/m3) | 0.13 | 1.05 | 1.08 | <DL | <DL | <DL | 179 |
| Average total protein concentrations (μg/m3) | 1.28 | 0.22 | 1.50 | 1.64 | 0.38 | 2.02 | 61.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suanno, C.; Sandrini, S.; Aloisi, I.; De Nuntiis, P.; Facchini, M.C.; Del Duca, S.; Fernández-González, D. Airborne Pollen, Allergens, and Proteins: A Comparative Study of Three Sampling Methods. Sustainability 2022, 14, 11825. https://doi.org/10.3390/su141911825
Suanno C, Sandrini S, Aloisi I, De Nuntiis P, Facchini MC, Del Duca S, Fernández-González D. Airborne Pollen, Allergens, and Proteins: A Comparative Study of Three Sampling Methods. Sustainability. 2022; 14(19):11825. https://doi.org/10.3390/su141911825
Chicago/Turabian StyleSuanno, Chiara, Silvia Sandrini, Iris Aloisi, Paola De Nuntiis, Maria Cristina Facchini, Stefano Del Duca, and Delia Fernández-González. 2022. "Airborne Pollen, Allergens, and Proteins: A Comparative Study of Three Sampling Methods" Sustainability 14, no. 19: 11825. https://doi.org/10.3390/su141911825
APA StyleSuanno, C., Sandrini, S., Aloisi, I., De Nuntiis, P., Facchini, M. C., Del Duca, S., & Fernández-González, D. (2022). Airborne Pollen, Allergens, and Proteins: A Comparative Study of Three Sampling Methods. Sustainability, 14(19), 11825. https://doi.org/10.3390/su141911825