Abstract
The development of novel antifungal agents and, in particular, the widespread use of these medications over the course of the past two decades, has had a significant impact on the treatment of fungal infectious diseases. This has resulted in a complete transformation of the treatment of fungal infectious diseases. However, the widespread development of antibiotic resistance has masked the significance of such breakthroughs. Antifungal infection treatment with nanoparticles has been shown to be effective. As a result of their unique characteristics, these substances, in contrast to antibiotics in their purest form, are able to exhibit an increased anti-proliferative capacity while requiring a lower concentration than traditional drugs do in order to achieve the same effect. Decreased drug effectiveness, minimal tissue penetration throughout tissue, restricted tissue penetration, decreased bioavailability, poor drug pharmacokinetics, and low water solubility are some of the major factors contributing to the employment of antifungal medicines in delivery systems. Because of this, one of the primary goals of incorporating antifungal medications into varying sorts of nanoparticles is to reduce the negative effects of the drugs’ inherent qualities. This article provides an overview of the many types of nanoparticles, such as metal, metal oxide, and non-metal oxide nanoparticles, carbon-based nanoparticles, nanostructured lipid carriers, polymeric nanoparticles, solid lipid nanoparticles, nanofibers, antifungal peptides, composites, and ZnO quantum dots, that can be used as antifungal drug delivery systems, as well as the benefits that these nanomaterials have over purified medications.
1. Introduction
From a medical perspective, the relevance of fungi in human infections is clear throughout the world [1]. Fungi that were never infectious in humans before are now included in the list of pathogenic organisms, the number of which is growing at an alarming rate [2]. Because of their capacity to adjust to a wide range of environmental circumstances, these fungi are able to quickly pose a threat to the survival of immunocompromised patients, and have become one of the most common causes of mortality in this patient population [3]. As a result, the accurate diagnosis and prompt treatment of infections of this kind are of the utmost significance, necessitating exhaustive information in this respect. According to recent studies, more than 300 million individuals worldwide are affected by dangerous fungal infections, which are responsible for almost 1.4 million fatalities each year [4]. The management of fungal infectious diseases has been completely transformed as a result of the development of novel antifungals and the widespread usage of these medications over the past two decades [5,6,7,8]. Unfortunately, on the other hand, growing rates of medication resistance throughout the world have obscured such advancements. People living with the human immunodeficiency virus (HIV) have seen a discernible decline in the frequency of fungal infections as a direct result of antiretroviral therapy (ART), which has been implemented in developed countries [9]. Additionally, in underdeveloped nations, where such pharmaceuticals are not readily available, there has been a clear rise in the frequency of fungal infections. Furthermore, the use of invasive therapeutic interventions in healthcare settings, the use of immunosuppressive medications after organ transplantation, the increased need for antifungal prophylaxis with azole derivatives, the treatment of malignancies, and the growing use of amphotericin B during empiric treatment have resulted in considerable shifts in various forms of fungal infection occurrences with different patterns and, particularly, drug resistance emergence [10]. Because of this, researchers have put forth their fullest effort to reduce medication resistance and toxic effects through the use of effective approaches.
Fungi are a varied group of eukaryotic creatures that colonize a variety of ecological niches, each of which requires a specific set of morphological characteristics [11]. It has been projected that there are between 3 and 5 million different types of fungi in the environment and. of those, around 300 different species have the potential to cause diseases in humans [12,13]. Because they most frequently infect people whose immune systems are compromised, the majority of the members of this fungus group are classified as opportunistic pathogens [14]. Meanwhile, only a select few species provide a significant risk to people who are otherwise healthy. The human fungal pathogens are responsible for a wide variety of infectious diseases, including those that affect the mucosa, the skin, and the invasive tissue [15,16,17]. Because of their rapid development, resistance to high temperatures (37 degrees Celsius), ability to exploit the host’s nutrition to their advantage, ability to penetrate tissues, and ability to avoid the host immune system, pathogenic fungi are successful in infecting humans and causing illness [18,19]. The Cryptococcus, Mucor, Aspergillus, and Candida genus are some of the fungal species that are responsible for the majority of the fatal illnesses that are caused by fungi [20,21,22].
It is absolutely necessary for the elimination of the pathogen to have a timely exposure to a suitable antifungal treatment at a dose that is suitably high [23]. The majority of patients diagnosed with fungal infections, on the other hand, have serious underlying conditions in addition to a variety of co-morbidities, which results in an increased sensitivity to adverse medication responses [15,24]. In addition, co-morbidities have the potential to influence the distribution, metabolism, absorption, and removal of antifungals and other crucial medications [25]. Absorption of orally given azoles or flucytosine may be affected, resulting in sub-therapeutic exposure, if the gastrointestinal tract is compromised, such as when chemotherapy is used to treat cancer or when there is decreased perfusion of the gastrointestinal tract [26]. Impairment of renal and hepatic functioning may change metabolism and elimination processes. Typical pathophysiological changes that occur during critical illness, such as changes in tissue perfusion, tissue penetration, hydration, hemodynamics, and plasma protein concentrations, may impact medication distribution [27,28]. Because the great majority of people who have infectious fungal infections also have other medical conditions and use other drugs at the same time, pharmacokinetic and pharmacodynamic drug–drug interactions with antifungals are quite prevalent. The distribution and elimination of a medication may be altered by the use of extracorporeal organ support [29,30]. It is possible that the pharmacokinetics of these unique patient groups are substantially different from those of healthy people or of patients with less severe impairments. Under these unusual circumstances, determining the correct dosage of antifungal medication is difficult because the relevant pharmacokinetic data is scant or perhaps nonexistent [31].
Nanoparticles have gained a lot of attention in recent years due to the diverse range of uses they offer in a variety of biological, pharmaceutical, and medical disciplines [32]. In terms of their structure, their dimensions are only slightly larger than the range of 100 nanometers. These nanomaterials have the ability to exert control over a wide variety of medications, including tiny pharmaceuticals that are either hydrophilic or hydrophobic, as well as biological molecules and vaccinations [33,34]. Nanoparticles have found widespread use in recent years, particularly in the fields of targeted drug delivery, tissue engineering scaffolds, and disease diagnostics [35,36]. The use of nanoparticles in various forms, such as carbon nanotubes, nanoliposomes, nanocapsules, nanofibers, as medication transporters, as well as cellular scaffolds, has seen widespread application [37,38,39,40]. Manufacturing nanoparticles for use as a drug delivery system has several goals, the most important of which is to regulate the size of the particles, surface features, and the efficient distribution of a particular medication at a certain time and location in order to obtain the maximal impact possible from the nanoparticles [41,42]. Not only should the nanoparticles that are utilized for drug administration contain biocompatibility as well as biodegradability features, but their optimal mechanical characteristics, timely release, and simplicity of manufacture are also all important considerations that need to be considered carefully [32]. Because of phagocytosis or systemic circulation, nanoparticles can become trapped inside the body [43]. These nanomaterials can be traced by surface functionalization and, as a result, are preserved in the systemic circulation [44].
Nanostructures can be classified according to a wide number of criteria, including their shape, size, and the components that make them up (Figure 1) [45]. Even the procedures used to prepare nanostructures can result in the production of a wide diversity of nanoparticles, each of which will have a unique load bearing capacity, distribution mechanism, and half-life. When nanoparticles are coupled with medications, they show unique therapeutic potential for treating particular illnesses, such as malignancies. There are two methods in which these nanoparticles are bound to nanocomposites, as follows: (1) The pharmaceuticals are conjugated across the surface of the nanoparticles. (2) The medications are placed inside microcarriers [46]. Numerous polymeric materials have been utilized as drug carriers thus far. These polymers include poly (alkyl-cyanoacrylates), polyamino acid, polyorthoester, polycaprolactone, polyamide, polyurethane, polyester, and polyacrylamide [47]. Among these, aliphatic polyester thermoplastics, such as polyglycolic acid (PGA), polylactic acid (PLA), and as the copolymer of these two, poly lactic-co-glycolic acid (PLGA), have been widely employed in comparison to others thanks to their superb biocompatibility, as well as biodegradability features [48,49]. Langer and Folkman were the first people to discover the controlled release of macromolecules through polymeric materials [50]. This discovery paved the way for further development of the drug release mechanism used in cancer treatments that target angiogenesis. Nanoparticles derived from polymers are an appropriate instrument for the direction of biomolecules, genes, medicines, and vaccinations [51]. By having the medications encapsulated inside nanostructures, it is possible to improve both the pharmaceuticals’ solubility as well as their half-life [32]. By acting in this manner, the primary goals that may be accomplished are focused medication transport using a specific dosage that falls within the therapeutic window and, subsequently, the pleasure of the patient. Polymer transporters, on account of their ability to perform a variety of activities and the possibility that they might be surface modified, have garnered a significant amount of interest over the course of the past several decades [52,53]. Generally, polymeric nanoparticles are miniature colloidal frameworks wherein the medication is either chemically bound to their primary polymer chains or physically disseminated throughout the structure [54]. The higher medication solubility, as well as durability, is one of the advantages of using polymeric nanoparticles as a pharmaceutical nanocarrier. Another benefit is the reduced risk of side effects [55]. Higher permeability, bioavailability, lower treatment cost, improved solubility, longer half-life, greater storage stability, and a sufficient dosage of medicine are some of the advantages that accrue from employing these nanoparticles in conjunction with antifungals [56,57]. In this review, we will provide an overview of the most important studies pertaining to nanomaterials, including carbon-based nanoparticles, metal, metal oxide, and non-metal oxide nanoparticles, solid lipid nanoparticles, antifungal peptides, nanofibers, composites, polymeric nanoparticles, nanostructured lipid carriers, and ZnO quantum dots (QDs), as well as some of the approaches utilizing nanostructures to improve traditional methods of therapy.
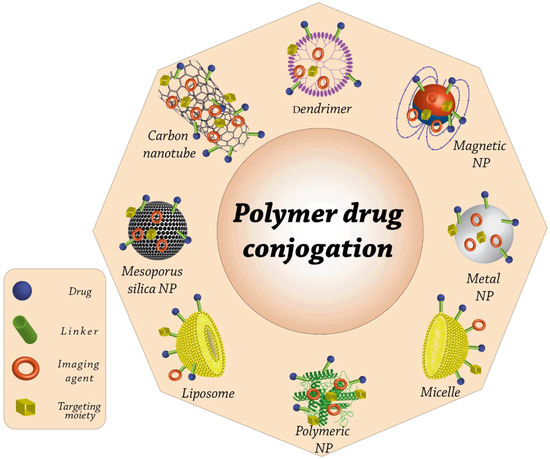
Figure 1.
Cutting-edge drug delivery systems that are mediated by nanocarriers and intended to deliver therapeutic agents to particular locations are now being used in a wide range of applications. In many aspects, nanotechnology can simplify the process of treating chronically sick patients. One method is to ensure that the appropriate medications are delivered at the appropriate times and locations.
2. Fungal Infection and Biofilm Formation
In recent years, several researchers have focused their attention on the process of the formation of biofilms by microorganisms [58,59]. Relative to free planktonic or cells, this kind of proliferation predominates in nature. However, because of its increased vulnerability to environmental variables and antimicrobial drugs, it poses a threat, particularly in clinical settings. This is the case because of the nature of the environment. In recent years, there has been a rise in interest in the biofilms that are produced when pathogenic fungi are allowed to grow, and various species of yeast, filamentous, and dimorphic fungi have been characterized as being able to proliferate into colonies [60,61,62].
Biofilms comprise sessile microbial populations that firmly attach to substrates or surfaces and to each other [63]. They are safeguarded by an extracellular matrix with a polymeric basis that is formed mostly of polysaccharides [64,65]. These cells have a higher resistance to treatment and a distinct phenotype in comparison to planktonic or free cells; they are also related to the continued presence of pathogens.
Infectious fungi are also capable of adhering to inorganic surfaces, including those found on catheters and prostheses [58,66]. Yeasts, in particular, are positioned to take control of this circumstance in order to obtain access to the patient’s circulatory system and, eventually, their internal tissues. This is cause for concern given the high death risk associated with spreading fungal infections [67].
Biofilms may be produced by yeast and also by filamentous fungus; nevertheless, fewer investigations have been conducted on biofilms generated by filamentous fungi relative to those produced by yeast [68,69]. According to Harding and colleagues, this is due to the fact that, for a considerable amount of time, the biofilms created by filamentous fungus did not meet the criteria for biofilms that were associated to bacteria [70]. Therefore, the authors provided a model for the production of biofilm by filamentous fungi and suggested that, despite the unique appearance, this design was comparable to the development of biofilm by yeast and bacteria. Figure 2 provides a representation of the developmental phases of biofilms produced by filamentous fungus. It includes the process of propagule adsorption, which involves the interaction of spores, pieces of hyphae, or sporangia with a surface. After that, active adhesion began, which is characterized by the secretion of adhesins by spores during the germination process and also other reproductive mechanisms. The subsequent step is the formation of the first microcolony, which includes hyphal extension and branching, the formation of a monolayer and, thus, the development of an extracellular matrix. The subsequent stage involves the production of a second microcolony or the initiation of the maturation process. This step sees the formation of dense hyphae networks in three dimensions and also the creation of water channels or an extracellular matrix. The final maturity stage then takes place, which is marked by the formation of fruiting bodies and, maybe, additional structures that help the fungus survive. The dispersion phase, also known as the planktonic phase, has eventually occurred. During this phase, conidia and/or hyphae fragments are expelled, which initiates a new cycle. In addition to this, filamentous fungi are distinguished from other types of organisms by their capacity to secrete tiny proteins referred to as hydrophobins. These proteins are important in the adherence of hyphae to hydrophobic surfaces, which suggests that they may also be important in the creation of biofilms [71].
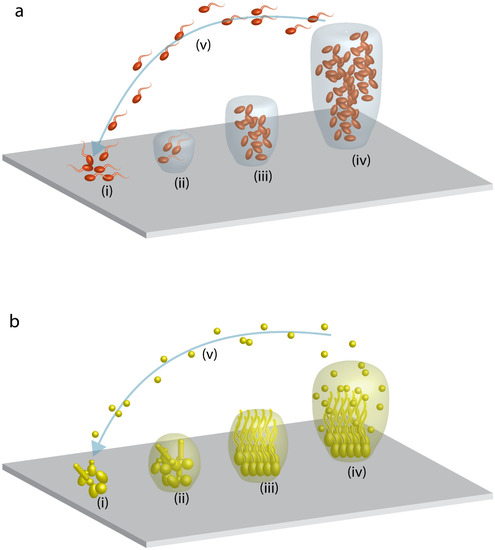
Figure 2.
The five steps that are related to the biofilm formation in bacteria (a) and Candida albicans (b) are as follows: (i) adsorption, (ii) adhesion, (iii) microcolony development, (iv) mature biofilm, and (v) dispersal [70,72,73,74].
The number of studies that are connected to fungal biofilms has significantly risen, and a number of different species have demonstrated the potential to build these populations [75,76]. The systemic fungal disease known as paracoccidioidomycosis is caused by a dimorphic fungus called Paracoccidioides brasiliensis. This disease is indigenous to Latin America. In their study, Sardi and colleagues characterized the biofilms that were produced by this fungus while it was in the yeast phase [77]. They discovered that in vitro community development was related not only to enhanced expression of genes of adhesion molecules but also enzymes including glyceraldehyde 3-phosphate dehydrogenase (GAPDH), enolase, aspartyl proteinase, and glycoprotein gp43, and a lowered expression of phospholipase.
Fungi known as dermatophytes are responsible for the development of dermatophytosis, which is one of the most prevalent dermatomycoses found in humans and other animals [78]. Onychomycosis is a kind of dermatophytosis that frequently recurs and requires protracted therapy that is frequently futile. Given this background, as well as the theory of Burkhart et al. [79], which argues that biofilm development by dermatophytes might characterize dermatophytomas, Costa-Orlandi et al. [73] demonstrated in vitro biofilm development by two of the most prevalent species found all over the world, namely Trichophyton mentagrophytes and T. rubrum.
Pitangui et al. were the first to report the biofilm of Histoplasma capsulatum [80]. In addition to displaying temperature dimorphism, this fungus is the causative agent of histoplasmosis, a systemic and respiratory mycosis whose progression is dependent on the survival of yeast in alveolar macrophages and its ability to replicate. As a result, the authors decided to explore the biofilm development of two strains isolated in vitro, and also their adherence to pneumocytes, as well as their ability to internalize them.
In the cases of Trichophyton, Paracoccidioides, and Histoplasma, additional research is being conducted to characterize their respective biofilms. The purpose of these additional studies is either to associate the populations with an increased resistance to select potential biological markers utilizing “omics” methodologies or to antimicrobial agents [81,82].
Since the middle of the 1990s, research has been carried out on the biofilms that are produced by pathogenic fungus of the Candida species [83,84,85,86]. In vitro investigations are more common than in vivo ones, and they prove the heterogeneity of such biofilm communities [87,88], which are made up of thick layers of pseudohyphae, yeast blastopores, hyphals, and an extracellular matrix [89]. The quorum sensing, formation of an extracellular matrix, morphogenesis, and adherence of biofilms are all controlled by a number of genes, notably in Candida albicans. Furthermore, genetic study indicated that yeasts and hyphae each play a distinct function in the production of biofilm by this species [90,91]. Paramonova and colleagues demonstrated that the majority of filamentation is clearly connected to enhancing the biofilm’s compressive force [92]. This results in biofilms being more resistant to damaging circumstances, such as sonication and vortexing. Silva et al. investigated the variations in the content, development, and shape of the extracellular matrix of biofilms generated by Candida parapsilosis, Candida tropicalis, and Candida glabrata [93]. Regarding the morphology, certain Candida parapsilosis biofilms were composed of yeast cells in addition to pseudohyphae, whereas the biofilms created by other isolates consisted entirely of yeast cells. In conclusion, the majority of the biofilms produced by Candida tropicalis were exclusively made up of yeast cells, with a few exceptions revealing extended hyphal filaments, whereas the biofilms produced by Candida species included only yeast cells. When it came to the biofilms’ matrix composition, the three species that were examined had varying levels of carbohydrates and proteins in their respective biofilms.
3. Preventive and Therapeutic Application of Antifungal Nanomaterials
3.1. Metal Oxide and Non-Metal Oxide Nanoparticles
It is well known that nanoparticles can be effective antifungal agents in certain situations [94,95]. Numerous studies have been conducted to examine the antifungal capability of nanostructures, and the results have shown that these nanoparticles have a major suppressive effect on the vegetative growth of fungal mycelia. For instance, gold nanoparticles have been investigated for their potential antifungal action in Candida albicans biofilms [96,97]. This is due to the fact that, when combined with a photosensitizer, Au nanoparticles have the ability to enhance the efficacy of photodynamic treatment. Through direct contact with the pathogen’s lipids and proteins, Au nanoparticles have the potential to disrupt the cellular membranes of the pathogens. In addition, the combination of photosensitizers with metal nanomaterials might lessen the likelihood that infections will become immune to the effects of photodynamic treatment [96]. Kischkel et al. investigated the effectiveness of Ag nanoparticles in combination with propolis extract (PE) against Candida species as well as other fungus mature biofilms, discovering that the concentration needed for the formulation’s fungicidal action was lower than the cytotoxic concentration [98].
People who are immunocompromised are believed to be at a greater risk of contracting invasive fungal infections since, in most cases, immunosuppression is the cause of invasive fungal infections in humans. Nevertheless, in light of the extremely restricted supply of existing antifungal medications and the growing occurrence of multi-drug resistance, it is imperative that research be conducted into the development of novel and different antifungal agents. Because of their vast surface area and their ability to effectively adhere to the fungal cell surface, biocompatible composites of QDs and metal nanoparticles have drawn a great deal of attention from researchers working on the development of new fungicidal agents [99,100]. The inhibitory mechanism of cadmium telluride QDs on yeast Saccharomyces cerevisiae was found and described by Han et al., which was linked to the accumulation of Cd2+ inside the yeast Saccharomyces cerevisiae cells, which was then followed by cellular dysfunction or deformation [101]. As a result of the cell wall being corroded, the cell wall shrank, which made it possible for Cd2+ to enter the cell through a newly created route. In addition, the growth rate of yeast was slowed down by photoelectron activation of the orange light released by cadmium telluride QDs (17.07 nm/L), whereas the growth rate of yeast cells was slowed down by 18.01 nm/L utilizing the green light emitted. In light of this, the photosensitizing wavelength, as well as the fluorescent color of the light that is emitted, are both regarded as key physiochemical parameters that determine the inhibition rates of yeast employing cadmium telluride QDs. In an attempt to improve the effectiveness of a naturally occurring cytotoxic agent, researchers loaded sesamol onto cadmium sulfide QDs that had been modified by chitosan (10.1016/j.carbpol.2019.03.024). As a result, the cytotoxic activity of sesamol was significantly enhanced, and it has the potential to be an effective agent against disorders caused by fungi or a functioning drug delivery system.
Recent discoveries in the structure and composition of fungal cell walls, in addition to their role in the development of drug resistance, have paved the way for the discovery of new targets for the treatment of fungal pathogens and contributed to a deeper understanding of the mechanisms underlying the evolution of antifungal resistance. Indeed, Ag nanoparticles have the potential to play a significant part in overcoming such resistance. The disintegration of the cellular wall, damage to nucleic acids and surface proteins caused by generation and accumulation of free radicals or reactive oxygen species (ROS), and the obstruction of proton pumps are the results of exposure to Ag nanoparticles. The accumulation of silver ions, which inhibits respiration by preventing the outflow of intracellular ions and, therefore, causes harm to the electron transport system, has been thought to be triggered by Ag nanoparticles [102]. The antifungal action of nanoparticles can be related to their smaller size in comparison to their larger surface area. These Ag nanoparticles of a lower size are able to readily pass through the cell membranes of their targets. To be more specific, silver nanoparticles with sizes ranging from 10 to 15 nm have improved durability, biocompatibility, and antimicrobial activities [103]. The generation of ROS, which ultimately results in apoptosis, is largely responsible for the toxicity of silver nanoparticles. The in vitro toxic effects of silver nanoparticles have been hypothesized to be induced either by the combined action of silver ions and Ag nanoparticles or by each of their individual effects [102,104]. According to the findings of a study, silver nanoparticles induce cellular damage to walls and also the cell membrane. They are capable of penetrating inside the cells, damaging organelles, such as mitochondria and ribosomes, and they are capable of inducing condensation and the margination of chromatin, which is a marker of apoptotic cell death [105]. Additionally, because of their tiny size, the silver nanoparticles have the potential to attach themselves to the cellular surface, enter the cells directly without causing any damage to the cell wall, and ultimately result in the death of the cell. It was demonstrated in a previous study on the bactericidal action of silver nanoparticles that the breakdown of the membranes of Escherichia coli might lead to the death of these bacteria because silver nanoparticles dramatically increase their permeability, which, ultimately, results in aberrant transport through the cellular membranes and cell death [106]. It was previously assumed that silver nanoparticles with high surface areas could readily generate Ag+, which would then attach to the functional groups (−SH) in proteins and cause the proteins to become denaturized. By binding to the surface proteins of fungi, silver nanoparticles have the potential to cause protein denaturation and also the destruction of proton pumps. This occurs as a result of an increase in the permeability of the protein–lipid bilayer or membrane, which ultimately leads to the disruption of the cellular membranes [107,108].
Curcumin possesses extensive antibacterial action and it does not harm living organisms in any way. Curcumin has already had restricted applicability as a treatment owing to its poor solubility in the circulation [109]. This is because of a number of variables, including disintegration and fast systemic clearance. As such, Ag nanomaterials have been developed in order to improve the delivery of curcumin. The amount of suppression caused by the curcumin in conjunction with Ag nanomaterials was proportional to the amount of curcumin that was employed, and it was substantial against fluconazole-resistant Candida glabrata and Candida albicans [110].
Antifungal activity can be found in high concentrations in the nanostructures of noble metals, such as gold and silver. Nasar et al. investigated the widespread antibacterial effectiveness of Ag nanoparticles against pathogenic bacterial strains (Bacillus subtilis, Escherichia coli, and Klebsiella pneumonia), as well as a common fungal pathogen called Aspergillus niger [111]. It has been discovered that Ag nanoparticles are efficient antifungals in the treatment of cutaneous infectious diseases [112]. In addition, Candida albicans and Staphylococcus aureus, which are the most common causes of oral microbial infections in humans, as well as Trichophyton mentagrophytes, can be eliminated with the use of Ag nanoparticles [113,114]. The Ag nanoparticles were able to prevent the development of Fusarium oxysporum at extremely low concentrations (less than 100 ppm), which resulted in a reduction in the generation of mycotoxin [114,115]. Additionally, Kischkel et al. observed the antifungal potential of the Ag nanoparticles against Candida albicans, Fusarium oxysporum, and Microsporum canis [98]. An investigation was carried out with the objective of comprehending the molecular and cellular processes of toxic effects induced by Ag nanoparticles in Candida albicans, which is a prevalent fungal pathogen [116]. Upon tracking the intracellular generation of ROS in the absence and presence of natural antioxidants, changes in surface topography, cellular ultrastructure, membrane microenvironment, membrane fluidity, membrane ergosterol, and fatty acids were used to evaluate the effects of Ag nanoparticles on molecular and cellular targets. At a concentration of 40 g/mL, spherical Ag nanoparticles (10–30 nm) exhibited a minimal inhibitory level. This is the concentration that must be present to stop the development of at least 90% of all organisms. According to the findings of our study, Ag nanoparticles created dose-dependent intracellular ROS, which displayed antifungal effects. Nevertheless, scavenging ROS with antioxidants was unable to provide protection from the death caused by Ag nanoparticles. After treatment with Ag nanoparticles, the surface morphology, cellular ultrastructure, membrane microenvironment, membrane fluidity, ergosterol concentration, and fatty acid composition were all changed. Oleic acid was particularly affected. The Ag nanoparticles had an effect on many cellular targets in the fungal cells, all of which were essential for drug resistance and pathogenicity. The research uncovered additional cellular targets of Ag nanoparticles, some of which are fatty acids, such as oleic acid, which are essential for the formation of hyphae (a pathogenic trait of Candida). Due to the fact that the shift from yeast to hypha is essential for both virulence and the creation of biofilm, it is possible that targeting virulence may emerge as a novel approach in the process of creating nano silver-based treatments for clinical applications in fungal therapies (Figure 3).
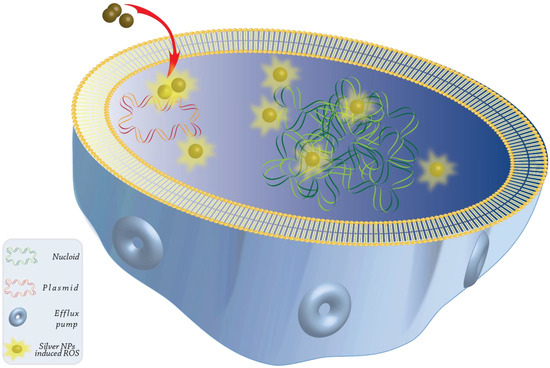
Figure 3.
The proposed model for the mechanism of action of Ag nanoparticles against Candida cells is shown below in the form of a graphical representation. It illustrates various cellular targets, as well as the existence of dependent and independent ROS mechanisms for the fungicidal activity of Ag nanoparticles. It has been indicated that nanoparticles dramatically change drug vulnerabilities in Candida albicans by acting on various cellular targets, which results in changes to the lipid composition, the membrane’s physical state, the cellular ultrastructure, the membrane microenvironment, and the surface morphology. The presence of change-induced Ag nanoparticles in cellular targets has the possibility of having an effect on the pathogenesis, treatment resistance, and virulence of fungal infectious diseases. Among the cellular targets of Ag nanoparticles, oleic acid, and fungal fatty acids, in particular, appear to be fairly significant. As a result, targeting virulence and morphogenesis through fatty acids could be one way to design a strategy for the treatment of Candida infections.
In light of the inert and harmless nature of gold nanoparticles, they are thought to be a comparatively safer alternative to other types of inorganic nanoparticles. In addition, the redox nature of gold is helpful in lowering the amount of ROS formed as a result of exposure to nanoparticles, which is a positive effect [117]. The fact that gold has a high level of biocompatibility is demonstrated by the fact that colloidal gold nanoparticles have been utilized for medicinal purposes for millennia [118,119]. Recent findings from a number of studies have provided conclusive evidence that gold nanoparticles have potent antimicrobial activity against a wide variety of clinical strains of the Candida fungus [120,121]. Extensive research on Candida led to the discovery of the antifungal mechanisms that are associated with Au nanoparticles. One theory proposes that Au nanoparticles are capable of interacting with the H+-ATPase-mediated proton pump of Candida, thus, disturbing the maintenance of the proton gradient. This, in turn, leads to abnormalities in nutrition transfer, which ultimately results in the death of the fungi [121]. Furthermore, gold may interact with phosphorus and sulfur, which are both essential components of fungal DNA and protein, and this can have a direct and deleterious impact on the functioning of those components. According to a different hypothesis, endonuclear Au nanoparticles have the capability to trigger karyopyknosis and DNA fragmentation, whereas cytoplasmic Au nanoparticles interact with mitochondria, which results in a change in bulk, calcium ion level, and the disordering of mitochondrial membrane potential (MMP). It was discovered that the ROS level had remained essentially unaltered, which lends credence to the notion that the fungi-killing function is independent of ROS [122]. The prevention of attachment between substrate surfaces and adhesin is one of the mechanisms that may be linked to the disintegration of fungal biofilms. This results in the disintegration of fungal biofilms as well as an increase in the number of planktonic fungi. Pathogens can also be inhibited or killed by Au nanoparticles because they release ions, rupture cell membranes or cellular walls, produce ROS, and cause damage to DNA or mitochondria, among other mechanisms [123,124]. Furthermore, microscopy using the potassium hydroxide technique, which is the preferred method and most used approach for screening for fungi, allows for direct identification of the organisms that cause fungal infections. Nevertheless, in order to conduct this test, one will need skilled staff who are able to operate specialized equipment, such as a fluorescence microscope and culture facilities. Since the majority of patients with acute infections seek medical attention within the first few days of experiencing symptoms, the best diagnostic test would be one that is both quick and self-diagnostic, which would both simplify treatment and improve its effectiveness. Within two minutes, Aspergillus niger may bring about a transformation in the morphology of gold nanoparticles that leads to a shift in color from red to blue in solutions containing these nanoparticles. The supernatant of human toenail samples that were disseminated in water exhibited a color shift that was analogous to that seen in the samples themselves. The use of scanning electron microscopy, ultraviolet–visible spectroscopy, and Raman spectroscopy is utilized in order to monitor changes in the morphology of the nanoparticles as well as surface plasmon resonance. The absorbance ratio at 520 nm/620 nm is used to conduct the investigation into the connection between the change in color and the fungal infection. The ratio was observed to decrease as the concentration of the fungus grew from 1 to 16 CFU/mL, with a detection limit of 10 CFU/mL. This was discovered by the researchers. This technique for the diagnosis of fungal infections based on plasmonic gold nanoparticles monitors the change in the form of gold nanoparticles and creates colored solutions with different tonality. With just the naked eye, this application might let laboratories and hospitals with limited resources self-diagnose and maintain hygienic conditions [125,126].
Abd-Elsalam et al. have explored the possible suppressive effect that a number of different metal oxide nanostructures have on the development of fungi [127]. Zinc oxide nanoparticles are the most attractive options among various metallic oxide nanoparticles because they suppress the development of Candida albicans at extremely low doses, ranging from 1.013 to 296.0 g/mL [128]. It has been shown that the structure and dimensions of ZnO nanoparticles have a significant role in determining the level of antifungal potential that they exhibit. At levels lower than 5 mM, ZnO nanostructures with flower shapes prevented the growth of Aspergillus flavus, as well as the formation of aflatoxin [129,130]. Iron oxide nanoparticles are the next type of metal oxide nanoparticle that has significant antimicrobial capability. In research involving magnetic (Fe2O3) nanoparticles, the antifungal effectiveness against Aspergillus flavus and the inhibition of aflatoxin formation were documented [130,131,132]. Although Mouhamed et al. established the inhibitory effects of iron oxide nanoparticles on ochratoxigenic Aspergillus sp., other researchers have not been able to replicate these results [133]. In addition, Abd El-Tawab and colleagues found that the growth inhibitory capabilities of Fe2O3 nanoparticles were effective against the microorganisms that are responsible for bovine skin disorders (T.mentagrophytes and Trichophyton verrucosum) [134].
Surface functionalization or coating of metal oxide or metallic nanomaterials might further enhance the antibacterial capabilities of the particulates. Chitosan nanoparticles, which are formed from a deacetylated derivative of chitin, have the ability to inhibit the development of fungi, such as Aspergillus niger, Rhizopus sp., and Fusarium sp., and, as a result, they can be employed in place of chemical pesticides [135]. Chitosan nanoparticles have also been shown to be effective at inhibiting pathogens when tested under in vitro conditions [136]. Chitosan-based polymeric materials have also been used to generate surface coatings for metal oxide nanoparticles, which can enhance the interactions between the biological membranes and nanoparticles, allowing for easier transportation of the nanomaterials. Abd-Elsalam et al. discovered the strong antifungal potential of Cu nanomaterials against mycotoxigenic fungus, both alone and in conjunction with chitosan [137]. This resulted in the suppression of aflatoxin formation. Inhibition of the development of Candida albicans can be achieved by the utilization of an acrylic resin that is fortified with Ag and ZnO nanoparticles [138].
Combining distinct types of nanostructures with favorable biologically active substances is a common practice in modern nanocomposites manufacturing, which results in the creation of pharmaceuticals that have significant uses in animal care [139]. The overlaying or coupling of nanoparticles through other biological macromolecules are connected to the chemical characteristics of the nanoparticles themselves and are exploited in the monitoring of infections inside the body. In this regard, Hassan and colleagues have discovered that the antifungal potential can be greatly increased by conjugating metallic nanoparticles with natural oils [140]. They found that in composites consisting of ZnO nanoparticles or Ag nanoparticles, essential oils were able to successfully suppress the growth of bacterial as well as fungal diseases. In the treatment of bovine skin and udder diseases, as well as carbon nanoparticles, combinations of active carbon nanoparticles with essential oils were used [141,142]. In order to effectively suppress the growth of toxigenic Aspergillus flavus and Escherichia coli, as well as to prevent the production of related toxins, Hassan et al. conjugated ZnO nanoparticles and Ag nanoparticles with cinnamon and olive oil [115,140]. Wang et al. have actually discovered that the hybridization of Au nanoparticles with autoantibodies assist in immune–chromatographic research and detection of toxic AFM1 in milk [143]. It was observed that using QDs to monitor activities and events of body cells was superior to using standard dyes, and this assisted in the release of medication to the desired location of infection. Similar behaviors were achieved using QDs to monitor activities of body cells [144].
Isolates tested of Candida albicans have been used to investigate the antifungal properties of amphotericin B and nystatin that have been coupled to magnetic nanoparticles. According to observations made by Niemirowicz and colleagues, these magnetic nanoparticles had substantial fungicidal activity and inhibited the formation of biofilm [145]. Because a disruption in the redox balance may restrict the development of Candida albicans, the detected effects may well be the result of catalase inactivation (Cat1) in cells that were treated with nanomaterials. After that, magnetic nanoparticles covered with ceragenin CSA-13 and peptide LL-37 also demonstrated fungicidal properties against Candida sp. This was because enhanced production of ROS generation was connected with hole creation in the cellular membranes, which let magnetic nanoparticles penetrate deeper into the yeast cells [146].
Inorganic mesoporous silica nanomaterials, commonly known as MSNs, are capable of functioning as nanocarriers for the transport of drugs to afflicted cells located anywhere in the body [130,147,148,149]. The pharmaceutical molecules can be attached to functionalized silica nanoparticles, or the nanoparticles can sequester or adsorb the medication molecules within the nanopores or on their surface, therefore increasing the delivery of therapeutics to the targeted locations [150]. In order to inhibit the creation of biofilms on medical implants, Kanugala et al. created antimicrobial biomaterial surfaces based on MSNs functionalized with phenazine-1-carboxamide [151]. In addition to their polymicrobial antibiofilm capability, the newly created MSNs demonstrated excellent anti-candidal action. Silica nanoparticles also have the potential to be utilized in the creation of topical cream compositions that cure skin fungal infectious diseases. An MSN-econazole topical cream formulation functionalized with aminopropyl was developed and studied against Candida albicans infections of the skin by Montazeri et al [152]. The researchers found that the formulation increased antifungal effectiveness at decreased concentrations of the packed medication.
3.2. Polymeric Nanoparticles
As shown in Table 1, antimicrobial polymers have cidal effects on both filamentous fungi and yeasts due to their cationic charge, which provides a high binding affinity to the negatively charged microbial membrane surface. These polymers have garnered a growing amount of interest in antifungal research as a result of the fact that they reduce the difficulty of finding novel antifungal targets and broaden the range of possible uses for both newly developed and already existing antifungal medicines. They are not expensive or time-consuming to make, and they can be chemically modified for a wider range of uses or incorporated into nanocomposites for controlled release [153]. As a consequence of this, there has been a renewed interest in the design of novel antimicrobial peptides that have enhanced physicochemical features.

Table 1.
A table representing a summary of synthetic antifungal polymeric materials and the reported targets of these compounds.
The use of amphotericin B deoxycholate as a medication in the treatment of severe fungal infections is still significant. Nevertheless, because the medicine is unable to pass across the blood–brain barrier, its application for the management of cryptococcal meningitis is restricted [164]. In the quest to find a brain medication delivery mechanism, several nanocarriers have been investigated, and some promising findings against Cryptococcus sp. have been revealed [165,166]. Polysorbate 80 is a surfactant as well as an emulsifier that increases nanoparticle absorption in the primary capillary endothelial cells of the brain in human and bovine-related cells. It was employed in the early research of nanocarriers. As a result of their ability to enhance the concentrations of the medication inside the brain by up to 20-fold one hour after injection, polysorbate 80 covered particulates are regarded as an effective “activator” for the brain [167]. Xu and colleagues produced amphotericin B-polybutylcyanoacrylate nanoparticles that were coated with polysorbate for use in a mouse model of systemic delivery. The authors report that nanoparticles with a diameter of about 69 nanometers were found in greater concentrations inside the brain 30 minutes after the administration of liposomal amphotericin B. It is interesting to note that amphotericin B deoxycholate was not found throughout the brain; however, the overall survival rates were 80% for these nanoparticles, 60% for liposomal amphotericin B, and 0% for amphotericin B deoxycholate, respectively. According to Ren et al., polysorbate 80 increases the efficiency of the entrapment of amphotericin B throughout the polymeric matrix when it is in the form of PLA-b-PEG. In experiments conducted in vitro, 100% of the amphotericin B was liberated between the periods of 35 and 40 h. Between 60 and 70 h after mixing, practically all of the amphotericin B contained in the nanoparticles that included the polysorbate was liberated [168]. In vivo and in vitro evaluations of the effectiveness, toxic effects, and oral bioavailability of some of the abovementioned formulations were carried out with the use of amphotericin B-encapsulated PLGA–PEG nanoparticles. The MIC of these nanoparticles against Candida albicans cells was lower in comparison to the MIC of free amphotericin B. When measured using a hemolysis assay, the toxicity of the nano-formulations was significantly lower than that of Fungizone® (APOTHECON®, Ben Venue Laboratories, Inc., Bedford, OH, USA). After one week of oral administration of amphotericin B-encapsulated PLGA–PEG nanoparticles to rats, plasma creatinine as well as blood urea nitrogen levels were found to be normal. This was determined in vivo. Last but not least, the incorporation of glycyrrhizin acid resulted in an increase in the bioavailability of the encapsulated amphotericin B [169]. Synthesized nanoparticles comprising PLGA with chitosan encapsulating amphotericin B attained nanoscale dimension, high encapsulation capacity for amphotericin B, positive surface charge, and reduced polydispersity.
A variety of polymeric nanoparticles have been investigated as potential transporters of amphotericin B for the therapeutic intervention of experimental aspergillosis [170,171]. Italia et al. were one of the research teams that revealed the usefulness of PLGA nanoparticles for the oral delivery of amphotericin B [172]. In neutropenic experimental models of widespread or aggressive aspergillosis, the oral bioavailability of PLGA nanoparticles was found to be more effective than the parenteral injection of Fungizone® and Ambisome® (Gilead Sciences, Inc., Foster City, CA, USA). It is important to note that standard amphotericin B, also known as Fungizone®, does not work in this case. When compared to amphotericin B on its own, the amphotericin B PLGA nanomaterials induced a higher oral absorption of the compound. Because the nanoparticles were capable of protecting the medicine from the degrading effects of digestive enzymes and pH, they were successful in overcoming the effects of entering metabolism, making it possible for additional nanoparticles to be taken up by lymph nodes. Thus, the oral delivery of amphotericin B could be a potential technique for the treatment of widespread fungal infections, or even, at the very least, oral thrush that is resistant to azole medications. Similarly, Van de Ven et al. discovered that PLGA plus nanosuspension nanoparticles carrying amphotericin B delivered intraperitoneally to mice were 3- and 2-fold more efficient than Fungizone® and Ambisome® in lowering fungal burden in widespread aspergillosis studies, respectively [173]. In this instance, the authors posited that the degree of aggregation of amphotericin B inside the delivery mechanism would have an effect on the way in which nanoparticles interacted with the ergosterol that is found within fungal membranes. By studying the UV/VIS absorption spectrum of the assessed formulations, researchers were able to demonstrate that there are distinct changes in the aggregating behaviors of the nanoparticles when they are in solution. In addition, the authors hypothesized that PLGA as well as nanosuspension nanomaterials may have been responsible for transporting the drug in a controlled manner to the tissue compartment. This is due to the fact that the nano-formulation has an optimal dimension for the circulatory system (less than 100 nanometers) and could start promoting accelerated absorption even by the reticuloendothelial system, as seems to be the scenario with formulations, such as Amphocil® (Criticare Laboratories Pvt Ltd., Bangalore, India) and Abelcet® (Leadiant Biosciences, Inc., Pomezia, Italy).
Salama et al. investigated the performance of cross-linked chitosan biguanidine that was packed with silver nanoparticles [170]. As a result of the successful integration of silver, the polymer thermal stability was enhanced and, thus, as a consequence, the nanoparticles produced had relatively lower cytotoxic effects for MCF-7 cells, as well as continuing to improve antibacterial and antifungal activities, in comparison to cross-linked chitosan biguanidine or chitosan. Polymers and silver nanoparticles are typically combined in this scenario with the intention of enhancing antimicrobial activity owing to the inherent characteristics of silver in conjunction with prolonged delivery provided by polymers. After the combination of cross-linked chitosan biguanidine with silver nanoparticles, the nanocomposite began to degrade at increasing temperatures resulting from interactions of these substances, which encouraged an increased stability in the framework but also led to the deterioration of the nanocomposite.
The action potential of antifungal medications can be improved by encapsulating the medications or developing new formulations for them that are in the form of nano-emulsions. As discovered by Deaguero et al., miconazole nano-encapsulated in oleate vesicles of cholesterol or sodium had substantial antifungal efficacy against a variety of fungal infections [174]. As was described by Siopi et al., liposomes encapsulated with amphotericin B have been shown to have great therapeutic effects against mycotic respiratory infectious diseases in mice induced by A. fumigatus [175].
Spadari et al. investigated the efficacy of miltefosine-loaded alginate nanoparticles in combating Candida and Cryptococcus species. Their goal was to minimize the cytotoxicity associated with miltefosine while preserving its antifungal properties. When compared to free miltefosine, miltefosine that was encapsulated in nanoparticles composed of 80% alginate showed a marked reduction in its harmful effects when tested on Galleria mellonella and also in an in vitro system. In addition, the administration of miltefosine–alginate nanoparticles to Galleria mellonella larvae that had been infected by Candida albicans greatly increased the amount of time that the larvae were able to survive. Since alginate-based nanocarriers provide a continual delivery as well as delivery of the medication, this can preserve the drug’s bioavailability and prevent any possible side effects. Another benefit is that the diameter of the nanostructures that were generated in this investigation (the mean diameter was 279.1 ± 56.7 nanometers), which is characterized as being advantageous for both oral and mucosal delivery [176].
Yang et al. devised an antifungal food packaging material produced from charcoal polymers and used it in their research. In order to create the charcoal polymers, charcoal powders were mixed with plastic resin while the process was carried out in a vacuum [177]. In the presence of filamentous fungi, these polymeric materials were examined (C. globosum, T. virens, and P. variotii). After being exposed to the substance, T. virens and P. variotii exhibited a growth inhibition that was 10 and 30%, respectively. Conidia of C. globosum that had been prepared with these polymeric materials likewise failed to germinate after being exposed to them for a period of 5 days in culture. The Ca2+ adsorption into the nanoscale pores of the charcoal is the strategy that these polymers employ in order to inhibit fungal growth. Within five hours, 0.2 g/ml of polymers were able to successfully remove an estimated 15 mM of Ca2+ from a solution containing CaCl2. These polymers lower the quantity of easily accessible calcium that is necessary for conidia to germinate and for the formation of fungal hyphae.
3.3. Carbon Nanoparticles
Antimicrobial properties may also be exhibited by various kinds of nanostructures based on carbon against various fungal and bacterial infections [178]. Nanomaterials that are based on carbon have the ability to thwart the growth of mycotoxigenic fungus and Escherichia coli [140]. Chitosan–carbon nanotube hydrogels, which are used in biomedicine for purposes, such as dressings, as well as medication delivery, were found to limit the growth of Staphylococcus aureus, Escherichia coli, and Candida tropicalis [179]. On a silicon substrate, arrays of branched carbon nanotubes (also known as CNTs) were produced by the process of plasma-enhanced chemical vapor deposition. Furthermore, Ni was employed as the catalyst, and it played a crucial part in the process of realizing branches in nanotubes that were vertically aligned. Under visible light, their antifungal action on Candida albicans biofilms was examined, and the results were compared to the activity of TiO2/CNT arrays and thin films of TiO2. As compared to the TiO2/CNTs and the TiO2 film, the TiO2/branched CNTs exhibited a photocatalytic antifungal activity that increased in significance [180,181]. Carbon dots were created by Parya Ezati et al. by utilizing glucose as a carbon source (GCD). In order to improve the functioning of the carbon dots, they were doped with heteroatoms, such as sulfur, boron, and nitrogen. The GCDs were extremely hydrophilic and durable in an aqueous system; moreover, the nitrogen-doped GCD (NGCD) exhibited the greatest polydispersity index with a value of 0.274. Every one of the GCDs possesses remarkable antioxidant action, with the NGCD having the highest level of activity. Both the sulfur-doped GCD (SGCD) as well as the boron-doped GCD (BGCD) display antibacterial activity that is much more effective against Escherichia coli and Listeria monocytogenes. The NGCD demonstrates the highest level of antibacterial activity and is effective against both types of bacterial strains. The NGCD shows strong antifungal action against Penicillium citrinum and Candida albicans, whereas the SGCD is more effective in inhibiting the growth of Fusarium solani. In addition, even after being exposed to a significant dosage of 500 g/mL for 72 h, more than 80% of the mouse fibroblast L929 cells are still alive, demonstrating that the toxicity level is rather low [182]. Leudjo Taka and colleagues created a nanosponge composite made of polyurethane cyclodextrin co-polymerized with phosphorylated multiwalled carbon nanotube-doped Ag-TiO2 nanoparticles [183]. They then tested the effectiveness of the nanosponge’s antifungal properties against two different strains of Aspergillus. This nanocomposite had a MIC of 437.5 g/mL against both Aspergillus ochraceus and Aspergillus fumigatus, and so this ratio is lower than the nanosponge that did not include doped nanoparticles, which had a value of 1750 g/mL. The antifungal effect is thought to be caused by the functional groups of the fungus membrane’s direct interaction with the nanocomposite, which then causes ROS creation and the interruption of the cell wall membrane. This is thought to be a probable effect of the addition of TiO2 and Ag nanoparticles to the material.
3.4. Nanostructured Lipid Carriers
Nanostructured lipid carriers are biocompatible and biodegradable because they are composed of a blend of solid lipid and a percentage of liquid lipid derived from natural sources [184]. Nanostructured lipid carriers are second-generation nanocarriers that have the potential to overcome the drawbacks of solid lipid nanoparticles (SLNs). These drawbacks include drug loss and a reduced drug load bearing capacity as a result of the formation and reorganization of extremely high crystallinity structures during storage [185]. Because the addition includes the liquid lipid fractions, nanostructured lipid carriers have better properties. These enhanced attributes give increased drug retention capability as well as long-term durability. Given that the majority of medications are lipophilic in nature, this sort of strategy is more successful in drug delivery [186]. Itraconazole that was included in nanostructured lipid carriers demonstrated an encapsulation effectiveness of more than 98% across several trials and maintained its integrity after being stored for a period of six months [187]. Beloqui et al. investigated the biodistribution of nanostructured lipid carriers in rats following intravenous administration of the substance and found that radiolabeled nanostructured lipid carriers continue to circulate in the bloodstream up to 24 h after the delivery of the compound [188]. The size of the particles and electrical charge are further factors that play a role in the biodistribution of nanocarriers. The lung is responsible for capturing larger nanoparticles, while the bone marrow and liver are responsible for capturing microscopic particles. Positive nanoparticles are found throughout the kidney, whereas negative nanoparticles make their way back to the liver. As a result, nanostructured lipid carriers have evolved into useful options in the field of drug delivery research.
It was possible to produce NLCs and SLNs that could be rapidly loaded with amphotericin B. In addition, the nanoparticles demonstrated a decreased level of hemolytic potential in comparison to Fungizone®. When compared to free amphotericin B or Fungizone®, the NLCs and SLNs loaded with amphotericin B demonstrated superior antifungal activity against Candida albicans. According to the findings, these nano-formulations could improve antifungal effectiveness, raise the solubility of amphotericin B, and lessen the treatment’s harmful impact. This impact may well be caused by the continuous release of amphotericin B inside the nano-formulations, as well as by its monomeric nature. Amphotericin B, having a limited degree of agglomeration, is much more specific and attaches predominantly to ergosterol. Therefore, this property of amphotericin B may be responsible for this action [189].
The antifungal activity of SLNs has been explored with Candida strains that are resistant to many drugs. Against the species that were examined, SLNs packed with fluconazole showed superior efficacy in comparison to free fluconazole. These specially constructed SLNs demonstrated rapid drug release during the first half hour, followed by continuous release for the full 24 h [190,191]. The efflux pump’s overexpression is one of the primary mechanisms of resistance in yeasts. This process aids in the reduction in azole levels within the cell. In this particular instance, the enhanced sensitivity to antifungal agents may be connected to the fact that NLCs gave shielding to fluconazole, preventing the medication from being released from the cells and contributing to the heightened vulnerability. Additionally, the hydrophobic surface of engineered NLCs has the potential to enhance the amount of medicine that is able to penetrate yeast. The efficacy of NLCs containing fluconazole as well as lipid core nanoparticles was investigated in C. albicans resistant to fluconazole. Although the NLCs did not seem to be efficient, the lipid nucleus nanoparticles did show some degree of activity at lower fluconazole doses. In addition, the lipid nucleus nanoparticles containing fluconazole were effective in preventing fluconazole from being recognized by the efflux pumps seen in fungal cells [192]. Table 2 presents a number of examples of antifungal medications that have been encapsulated in either NLCs or SLNs. Based on the findings, it was possible to produce drug-loaded NLCs as well as SLNs with a particle size ranging from 50 to 450 nm. An encapsulation efficiency of up to 100% of the drug was attained, proving that the preparation methods that were utilized were effective.
Table 2.
Various NLC and SLN formulations used for the antifungal drug delivery purposes.
3.5. ZnO Quantum Dots
“Quantum dots” (QDs) are spherical nanomaterials that are typically within 10 nanometers in size and have a size that is equivalent to or less than the Bohr radius of the substance [212]. This is the size range in which quantum confinement effects are most noticeable. Quantum confinement in QDs leads to the formation of active sites and surface defects, which, upon interaction with hydroxyl and oxygen ions, result in the formation of highly reactive superoxide and hydroxyl radicals [213]. In addition to this, ZnO nanoparticles facilitate broad spectrum antimicrobial action by targeting several molecular and cellular processes. This has the potential to prevent the future development of drug resistance in fungi. As a result, it is possible that the use of ZnO nanoparticles in conjunction with several other antifungal medications might result in a decrease in the typical dosages of antifungals, as well as the cost of therapy and the toxicity of the treatments [214]. In addition, the size-dependent toxic effects of ZnO nanoparticles toward microorganisms have been well reported, and it has been shown that a decrease in size results in an improvement in antimicrobial activity. Due to their smaller dimensions, ZnO QDs are capable of facilitating a significantly greater dissolution of Zn2+ ions throughout a solution [215,216]. This greater solubility of Zn2+ ions throughout a solution is thought to be accountable for an increase in ROS, which in turn leads to the peroxidation of biomolecules, as well as the death of cells in microbial pathogens [217]. Recent research demonstrated the broad array microbicidal effect of ZnO QDs (5–6 nanometers) against multidrug-resistant pathogens (Candida albicans and Escherichia coli). This finding suggests that the antibacterial activities of nano ZnO may be controlled by modifying the diameter of the particles. Nevertheless, in an investigation, Preeti Chand et al. illustrated the utilization of ZnO QDs in conjunction with antifungal medications of separate classes against drug-resistant and drug-susceptible strains of Candida albicans [218]. This was done in order to develop a biosafe, nano-based multi-functional structure of combination treatment against multi-drug resistant fungal pathogenic organisms, in which the concentration of drug can be reduced to minimize toxic effects while simultaneously benefiting from the benefit of multitargeted activity to restrict the development of fungal drug resistance. An essential theoretical goal for the surface functionalization of tiny (1–3 nm) ZnO quantum dot nanoparticles was conducted by Zahra Fakhroueian et al. in order to hinder the agglomeration and decomposition of nanomaterials in aqueous conditions [219]. The majority of the reagents employed in these processes include polymeric materials, organosilanes, PEG (polyethylene glycol), and oily herbal fatty acids. This is due to the fact that all of these substances are entirely soluble in water and have the potential to be utilized as biological probes in nanomedicine. The vegetable fatty acid-capped ZnO (QD nanoparticles) were produced by dissolving ZnO using the sol–gel technique with the involvement of nonionic surfactants as effective templates at a pH that was appropriate for the process. In the current study, the cellular toxicity of tiny ZnO QD nanoparticles bearing a specific blue fluorescence was the primary focus of our investigation, as it related to the targeted delivery of HT29 and MCF7 cancer cell lines. These studies demonstrated that ZnO QDs had minimal toxic effects on healthy cells (MDBK) and may eventually have a possible application in the field of cancer treatment. Due to the presence of these features, there is a possibility that a potential applicant in the field of nanobiomedicine could be produced. Antifungal and antibacterial capabilities were demonstrated by the robustly designed ZnO QD nanomaterials against Klebsiella pneumonia, Staphylococcus aureus, Staphylococcus epidermidis, and Bacillus anthracis bacteria, as well as various fungi including Trichophyton mentagrophytes, Microsporum canis, Microsporum gypseum, Candida tropicalis, and Candida albicans, in comparison with the standard antibiotic toxicity. The ZnO QDs had a substantial growth-inhibiting effect on Candida albicans cells [220]. Oxidative stress was implicated in the antifungal activity of ZnO QDs against Candida albicans. This effect was mediated by the enhancement of endogenous ROS. In addition, endogenous ROS generation by ZnO QDs, as well as their effect on destroying fungal cells, was researched while an antioxidant known as ascorbic acid was present. The findings demonstrated that the antioxidant did not provide complete protection against the oxidative stress produced by ZnO QDs.
3.6. Peptides That Are Antimicrobial and Have Efficacy against Fungi
Antimicrobial peptides, or AMPs for short, are a component of the innate immune response. They are also known as host defense peptides. Furthermore, AMPs are molecules that can be created by plants, animals, and microbes, and they serve the purpose of defending the host from invasive diseases. These amphiphilic peptides have short sequences that are generally less than one hundred amino acids in length [221]. The AMPs have a cationic charge, and those that are rich in the amino acid histidine have potent antifungal action [222]. Cathelicidins are a good example of this behavior. This group of antifungal AMPs is a component of the human innate defense system. They are typically deposited inside the lysosomes of macrophages and, thus, are responsible for preventing fungal infections [223]. Despite this, there are certain AMPs that have anionic charges and need metallic ions in order to undergo biological activation [224]. Anionic AMPs are responsible for the formation of cationic salt bridges between anionic metallic ions and microbial membranes, which allows for increased membrane permeability. In contrast to cationic AMPs, our understanding of the antibacterial activity of anionic AMPs is still somewhat restricted, despite the fact that this mechanism is ascribed to certain anionic AMPs [225]. The disruption of the cellular membranes that results from the interaction of electrostatic attraction with anionic membranes is thought to be the primary antibacterial mechanism that cationic AMPs possess. In addition to this, some AMPs have the ability to translocate across the membrane as well as impacting on intracellular targets, therefore, inhibiting the production of proteins and DNA [222]. Additionally, AMPs are unstable molecules with a limited half-life, despite the fact that they have antibacterial actions. In the antimicrobial peptides database, there are now 1211 peptides of natural, synthetic, or semi-synthetic origins that have antifungal activities [226].
Synthetic Antimicrobial Peptides Having Antifungal Properties
Due to the fact that synthetic peptide-like polymeric materials are synthesized on the basis of the structural moieties in AMPs, they often impede the growth of microorganisms or destroy them thanks to the cationic surface charge they possess [227]. Polymers that have intrinsic antimicrobial activity are included in this class. Some examples of these polymeric materials are polymers containing N-halamines, quaternary nitrogen groups, and poly-ε-lysine (ε-PL). Because of the potential for toxicity in humans, many synthesized AMPs are rarely utilized in clinical settings, despite the fact that they are very effective antifungal agents. As a result, there is continuing investigation into the synthesis and optimization of novel synthetic AMPs that have high antibacterial activity but lower toxicity. These AMPs are intended to be safer.
Ramamourthy and colleagues synthesized peptides featuring varied numbers of lysine as well as tryptophan repetitions (KWn-NH2) and then studied the antifungal efficacy of these peptides against Candida albicans [155]. The length of these peptides has a direct correlation with the strength of their antifungal and biofilm-removing properties. In addition to this, the shortest peptide, KW2, did not exhibit any antifungal action, while the longest peptide, KW5, exhibited toxic effects on human keratinocyte-treated cells. The fungal cell membranes were not disrupted by the KW4 peptide. On the other hand, laser-scanning confocal imaging revealed that KW4 was localized inside the cytoplasm of Candida albicans, where it was shown to be bound to RNA from the fungus. This leads one to believe that membrane permeabilization is not the sole antifungal mechanism possessed by these peptides. Rather, these artificial AMPs are capable of entering fungal cells and localizing within the cell, where they are capable of preventing biological processes via binding to DNA and RNA, which ultimately results in the death of fungal cells.
In light of the morphological characteristics of antimicrobial peptides (pilosilin and ponericin) collected from Dinoponera quadriceps (giant ant) venom, Dodou Lima and colleagues developed innovative synthesized AMPs featuring antifungal action [154]. Peptides with properties similar to ponericin (Dq-3162), as well as pilosulin (Dq-1503 and Dq-2562), were among the most effective antifungal peptides when used to combat Candida albicans, Candida tropicalis, Candida parapsilosis, and Candida krusei. These synthetic peptides, when combined with antifungal medicines, such as fluconazole, amphotericin B, ciclopirox, nystatin, and miconazole, demonstrated synergistic effects while also exhibiting modest hemolytic activity.
In a similar manner, Kodedová et al. synthesized antimicrobial peptides (hylanines, halictines, and lasioglossins), focusing on the antimicrobial peptides found in bee venom [228]. These peptides were effective against a wide variety of microorganisms. It was shown that these peptides quickly permeabilized the cell membranes of several Candida species, including Candida albicans, Candida parapsilosis, Candida dubliniensis, Candida glabrata, Candida krusei, and Candida tropicalis, as well as Saccharomyces cerevisiae. Nevertheless, the sensitivity of the species to permeabilization was shown to be dependent on the lipid content of the fungal membrane. Candida glabrata was shown to have greater resistance to such synthetic peptides after being pre-treated with terbinafine, which reduced the amount of ergosterol found in the fungal membrane. The cationic peptides demonstrated a powerful electrostatic attraction with the membrane lipids of anionic charges (cardiolipin, phosphatidic acid, and phosphatidylglycerol), as well as a partial interaction with the neutral phosphatidylethanolamine and phosphatidylinositol. This resulted in improved selectivity for the treatment of fungal infections. Therefore, synthesized AMPs that are based on the functional and structural features of AMPs can address the drawbacks of natural AMPs while keeping or increasing their antifungal actions.
3.7. Nanofibers
Due to their adjustable fiber diameter, adequate porosity, high specific surface area, high encapsulation effectiveness, as well as good consistency and flexibility, nanofibers have emerged as a focal point of study in recent years [229]. In a study by Hellen et al., the antibacterial activity of TiO2-CeO2 nanofibers against Candida glabrata, Candida albicans, and Candida krusei was assessed. The findings demonstrated the successful manufacture of mesoporous TiO2-CeO2 fibers with an average diameter ranging from 100 to 282 nanometers. According to the results of the antimicrobial experiment, the TiO2-CeO2 nanofibers considerably impeded the development of Candida glabrata, Candida krusei, and Candida albicans biofilms. The antifungal reaction against Candida glabrata was enhanced with CeO2 levels, but there was no rising trend detected for Candida krusei or Candida albicans. The TiO2-CeO2 nanofibers have intriguing antifungal capabilities and, as a result, they have the potential to be utilized in a variety of antifungal applications [230]. The use of dynamic high-pressure homogenization was explored in order to find a straightforward approach to the manufacturing of chitin nanofibers that is also environmentally friendly [231]. This method has been shown to be an extremely straightforward approach to the production of α-chitin nanofibers from yellow lobster. These nanofibers have a uniform width (below 100 nanometers) as well as a high aspect ratio, both of which could contribute to a significant development in the field of chitin applications. In addition, the resultant alpha-chitin nanofibers were studied and contrasted with natural alpha-chitin in terms of their thermal degradation, chemical and crystalline structures, and antifungal activity. The antifungal efficacy of chitin nanofibers against Aspergillus niger was investigated using biological tests, which revealed that the nanoscale nature of the nanofibers plays a significant role in this activity. Shinsuke Ifuku and colleagues manufactured silver nanoparticles on the surfaces of chitin nanofibers by reducing silver ions with ultraviolet light [232]. Chitin nanofibers have the potential to serve as effective substrates for the immobilization of silver nanomaterials in states of steady dispersion. Because of the action of the silver nanoparticles, the dispersion as well as the nanocomposite film that was made with acrylic resin exhibited the typical absorption behavior in the visible light area. Chitin nanofibers exhibited significant antifungal activity thanks to the presence of silver nanoparticles. Semnani et al. explored the in vitro antifungal effectiveness of eugenol-loaded polyacrylonitrile nanofibers as a result of the desired qualities of polyacrylonitrile [233]. Polyacrylonitrile is the significant phenolic component of clove essential oil used in the medical field. They conducted tests to determine how efficient this combination is against Candida albicans. The findings revealed that, when the ratio of eugenol was increased, there was an improvement in the average diameter of the nanofibers, which varied from 127 to 179 to 218 nanometers. The drug release characteristic of the specimens was progressive, and it was completed after 150 h. According to the findings, these eugenol-loaded nanofiber mats can be used either as a covering on a fabric substrate or even as a temporary wound dressing for cutaneous mucocutaneous candidiasis treatment in high-risk patient populations.
3.8. Antifungal Nanocomposites That Have SMOOTH Surfaces, Which Helps Reduce the Attachment of Microorganisms
In addition to providing other benefits, increasing the surface roughness of the substance can have an antibacterial impact since it reduces the number of locations where microbes can thrive. Polymethyl methacrylate (PMMA) was given antifungal qualities by Fouda et al. so that it could be used as dental filler. This was accomplished by using nanodiamonds to smooth the surface of the resin [234]. This is comparable to the research that was conducted by De Matteis et al., 2019, in which an increase in surface roughness led to a decrease in the attachment of Candida albicans [235]. Fouda et al., 2019, observed alterations in Candida albicans adhesion after adding biocompatible nanodiamonds to PMMA in several concentrations (zero, half, one, and one and a half percent by weight, respectively). A profilometer was used to determine the surface roughness, and a goniometer was utilized to determine the contact angle [234]. The appearance of valleys and peaks on the surface of PMMA was significantly reduced as a result of the incorporation of nanodiamonds into the material. When compared to the control group, this resulted in a reduction in the attachment of Candida albicans cells on the PMMA surface owing to a degradation of settling sites; the level of attachment that was determined to be the lowest was at one percent nanodiamonds.
4. Toxicity Effect of Nanoparticles
Nanoparticles are seeking employment in a wide array of industries, which is a growing trend. The cellular toxicity, immunotoxicity, and genotoxicity of nanoparticles have all been the subject of research. For the purpose of determining whether or not a cell is viable, tetrazolium-based assays, such as MTS, WST-1, and MTT are utilized. Additionally, ELISA is used to measure inflammatory biomarkers, such as tumor necrosis factor, IL-6, as well as IL-8, in order to determine whether or not nanoparticles have triggered an inflammatory response in cells [236,237]. The lactate dehydrogenase test is employed to evaluate the integrity of the cellular membranes [238]. In these in vitro toxicity assays, researchers have utilized a wide variety of cell cultures, including lines derived from cancerous cells. It is commonly accepted that nanoparticles cause interference, either with the substances used in assays or with the detection instruments. Because they have so many uses and may be applied in so many different contexts, human beings are going to be exposed to nanomaterials in one way or another, whether purposefully or not. To determine whether or not a nanoproduct is safe for human consumption, toxicological tests are performed on it first [239]. A number of different experiments have been performed in order to obtain sufficient data for these tests. In order to fulfill this regulatory need, some of the dangerous consequences of nanostructures have been investigated. Nevertheless, the toxicological statistics that have been produced thus far, according to studies, are contradictory and inconsistent [240]. Investigations in toxicology offer a solid foundation upon which to build safeguards not just for people but also for the environment. As a consequence of this, it is possible that, on the premise of the laboratory models that are now accessible, it is erroneous to identify some of the more useful nanoparticles as being harmful to living organisms, and vice versa [236]. In light of the fact that nanoparticles may find use in a wide variety of industries, but in an effort to close the existing knowledge gap, it is imperative that the applicable negative impacts of nanomaterials be evaluated by employing in vivo toxicological modeling techniques that are internationally accepted and objective and that focus on the crucial systems [241,242]. Nevertheless, researchers are of the viewpoint that thoughtful consideration should be given to the adaptation, design, and validation of these unique prototypes in the upcoming years not only for the purpose of the exposure route, the coating substance, and toxicology testing, but also for the sterilization of nanostructures and the kinds of cell culture systems [243,244]. In addition, the Food and Drug Administration (FDA) in the United States, which is a public health agency, has lately taken into consideration the important problem of toxicities related to products that encompass nanoparticles. The FDA does not take into account that these products are either completely dangerous or secure for human use, and instead maintains that every product must be applicable to the same regulations [237].
5. Conclusions and Future Perspectives
Over the course of the last few decades, breakthroughs in nanotechnology have provided progressively unique opportunities to enhance both the health and productivity of animals. In today’s world, fungal infectious diseases are gaining an ever-increasing relevance due to the rising yearly rates of such diseases, as well as the increased death rate which results from them. Despite the abundance of antifungal agents that are efficacious in preventing these fungal infectious diseases, researchers have been looking for ways to improve the available medications due to decreased bioavailability, poor solubility in water, adverse effects, inadequate tissue penetration, drug resistance and drug effectiveness, and inadequate pharmacological dynamics. Additionally, numerous different nanoparticles are utilized as antifungal nanoparticles. These nanostructures also have other applications, including the detection of disease, diagnostic testing, and therapeutics, as well as in the addition of preservatives to animal feed and also the product lines that organisms produce and, eventually, the protection of food. The key therapeutic and preventative properties of antifungal nanoparticles, notably the copper and zinc nanostructures, have been investigated for a wide range of fungal infections and mycotoxicosis in animals. These capabilities have been shown to be effective. In addition, opportunities for initial and precise detection of fungal infectious diseases, followed by thorough diagnosis and treatment, are being found for semiconductor quantum dots, gold nanomaterials, and super paramagnetic iron. Organic and inorganic polymeric nanoparticles have both been exploited for the targeted delivery of a variety of vaccinations, as well as for the on-site and rapid monitoring of fungal infectious diseases or their hallmark proteins and other biological components. The mechanisms of action behind the antifungal activities of nanoparticles are connected to their capacity to break through the cellular membranes and cause significant damage to the cytoplasmic contents, which ultimately results in the cells losing their capacity to perform and eventually dying. In light of this, further research is required to illustrate the cellular toxicity effects of activity that results in ROS damage and contributes to genotoxicity and malignancies in order to conduct an in-depth examination that will allow for the manipulation of the responsibilities that nanomaterials play in animal health. In addition, the toxicological concerns of nanostructures need to be identified prior to the application of nanoparticles in the medical field in order to protect the wellbeing of creatures.
As a result, over the course of the past two decades, one of the most prominent focuses on antifungal drug optimization has been involved in the employment of nanoparticles-based drug-delivery strategies to broaden the spectrum of antifungal activities. The key benefits of these nanoparticles are their tiny dimensions as well as their wide available surface area, which makes them perfect alternatives for a variety of applications. They are also capable of substantially overcoming the restrictions of the medications that are already on the market. However, taking into consideration all of the research that has been carried out in this area, with the exception of the few cases that have been described (such as liposomal amphotericin B, which is sold in nano-formulations), neither of these active ingredients are currently accessible for purchase in the marketplace. This is primarily due to the preclinical challenges that have been associated with the interventional studies and also the drawbacks that are associated with the application of these nanostructures.
Author Contributions
The authors of this manuscript contributed to the study conception and design. Material preparation, data collection and analysis were performed by A.J., Z.Z., A.T. and M.A. The first draft of the manuscript was written by A.V., A.G., L.T., E.V. and H.F. Other authors all the authors including A.M.A., H.K. and S.C. helping during revised versions of the manuscript, Bibliography, Conceptualization and Editing. All authors have read and agreed to the published version of the manuscript.
Funding
Part of the research reported in this paper was supported by National Institute of Dental & Craniofacial Research of the National Institutes of Health under award number R15DE027533, R56 DE029191 and 3R15DE027533-01A1W1.
Institutional Review Board Statement
The manuscript is a review paper and there is no experiment to need ethical approval.
Informed Consent Statement
Not applicable.
Data Availability Statement
The availability data and materials are limited to those presented in this manuscript.
Conflicts of Interest
The authors declare that no conflict of interest would prejudice the impartiality of this scientific work.
Consent to Participate
The authors of this manuscript consent to participate in the manuscript publication and the following has been explained to us: (a) The published manuscript may not be of direct benefit to us. (b) Our participation is completely voluntary.
Consent to Publish
The authors of this manuscript give our consent for the publication of identifiable details, which can include photograph(s) and/or videos and/or case history and/or details within the text (“Material”) to be published in the Sustainability journal.
References
- Vajargah, M.F.; Hashemi, G.; Bibak, M.; Yalsuyi, A.M. The effect of vitamin c-fortified artemia on growth and survival of sepia pharaonis larvae. J. Environ. Treat. Tech. 2021, 9, 815–818. [Google Scholar]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl. Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed]
- Garayemi, S.; Raeisi, F. Graphene Oxide as a Docking Station for Modern Drug Delivery System. by Ulva lactuca species study its antimicrobial, anti-fungal and anti-Blood cancer activity. Adv. Appl. NanoBio-Technol. 2020, 1, 53–62. [Google Scholar]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, rv13–rv165. [Google Scholar] [CrossRef] [PubMed]
- McKeny, P.T.; Nessel, T.A.; Zito, P.M. Antifungal Antibiotics; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar]
- Scorzoni, L.; de Paula e Silva, A.C.A.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.M.A.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.S.; Fusco-Almeida, A.M. Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef]
- Ansarifard, E.; Zareshahrabadi, Z.; Sarafraz, N.; Zomorodian, K. Evaluation of antimicrobial and antibiofilm activities of copper oxide nanoparticles within soft denture liners against oral pathogens. Bioinorg. Chem. Appl. 2021, 2021, 9939275. [Google Scholar] [CrossRef]
- Hejazi, M.; Zareshahrabadi, Z.; Ashayeri, S.; Saharkhiz, M.J.; Iraji, A.; Alishahi, M.; Zomorodian, K. Characterization and Physical and Biological Properties of Tissue Conditioner Incorporated with Carum copticum L. BioMed. Res. Int. 2021, 2021, 5577760. [Google Scholar] [CrossRef]
- Tseng, A.; Seet, J.; Phillips, E.J. The evolution of three decades of antiretroviral therapy: Challenges, triumphs and the promise of the future. Br. J. Clin. Pharmacol. 2015, 79, 182–194. [Google Scholar] [CrossRef]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal evolution: Major ecological adaptations and evolutionary transitions. Biol. Rev. Camb. Philos. Soc. 2019, 94, 1443–1476. [Google Scholar] [CrossRef]
- Köhler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that infect humans. Microbiol. Spectr. 2017, 5, a019273. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Lopez-Ribot, J.L. Current Antimycotics, New Prospects, and Future Approaches to Antifungal Therapy. Antibiotics 2020, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Rokas, A. Evolution of the human pathogenic lifestyle in fungi. Nat. Microbiol. 2022, 7, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, G.; Meng, G. Pathogenic fungal infection in the lung. Front. Immunol. 2019, 10, 1524. [Google Scholar] [CrossRef]
- Kim, J.Y. Human fungal pathogens: Why should we learn? J. Microbiol. 2016, 54, 145–148. [Google Scholar] [CrossRef]
- Marcos, C.M.; de Oliveira, H.C.; de Melo, W.d.C.M.A.; Da Silva, J.D.F.; Assato, P.A.; Scorzoni, L.; Rossi, S.A.; de Paula e Silva, A.C.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Anti-immune strategies of pathogenic fungi. Front. Cell. Infect. Microbiol. 2016, 6, 142. [Google Scholar] [CrossRef]
- Yu, Y.; Wolf, A.K.; Thusek, S.; Heinekamp, T.; Bromley, M.; Krappmann, S.; Beilhack, A. Direct Visualization of Fungal Burden in Filamentous Fungus-Infected Silkworms. J. Fungi 2021, 7, 136. [Google Scholar] [CrossRef]
- Góralska, K.; Blaszkowska, J.; Dzikowiec, M. Neuroinfections caused by fungi. Infection 2018, 46, 443–459. [Google Scholar] [CrossRef]
- Firacative, C. Invasive fungal disease in humans: Are we aware of the real impact? Mem. Inst. Oswaldo Cruz 2020, 115, e200430. [Google Scholar] [CrossRef]
- Prakash, H.; Chakrabarti, A. Global Epidemiology of Mucormycosis. J. Fungi 2019, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Hamdy, R.F.; Zaoutis, T.E.; Seo, S.K. Antifungal stewardship considerations for adults and pediatrics. Virulence 2017, 8, 658–672. [Google Scholar] [CrossRef]
- Bhatt, K.; Agolli, A.; Patel, M.H.; Garimella, R.; Devi, M.; Garcia, E.; Amin, H.; Domingue, C.; Del Castillo, R.G.; Sanchez-Gonzalez, M. High mortality co-infections of COVID-19 patients: Mucormycosis and other fungal infections. Discoveries 2021, 9, e126. [Google Scholar] [CrossRef] [PubMed]
- Bellmann, R.; Smuszkiewicz, P. Pharmacokinetics of antifungal drugs: Practical implications for optimized treatment of patients. Infection 2017, 45, 737–779. [Google Scholar] [CrossRef] [PubMed]
- Ruhnke, M.; Cornely, O.A.; Schmidt-Hieber, M.; Alakel, N.; Boell, B.; Buchheidt, D.; Christopeit, M.; Hasenkamp, J.; Heinz, W.J.; Hentrich, M.; et al. Treatment of invasive fungal diseases in cancer patients—Revised 2019 Recommendations of the Infectious Diseases Working Party (AGIHO) of the German Society of Hematology and Oncology (DGHO). Mycoses 2020, 63, 653–682. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.A.; Roberts, M.S.; Semark, A.; Udy, A.A.; Kirkpatrick, C.M.J.; Paterson, D.L.; Roberts, M.J.; Kruger, P.; Lipman, J. Antibiotic dosing in the ‘at risk’ critically ill patient: Linking pathophysiology with pharmacokinetics/pharmacodynamics in sepsis and trauma patients. BMC Anesthesiol. 2011, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Sanz Codina, M.; Zeitlinger, M. Biomarkers Predicting Tissue Pharmacokinetics of Antimicrobials in Sepsis: A Review. Clin. Pharmacokinet. 2022, 61, 593–617. [Google Scholar] [CrossRef]
- Dzierba, A.L.; Abrams, D.; Brodie, D. Medicating patients during extracorporeal membrane oxygenation: The evidence is building. Crit. Care 2017, 21, 66. [Google Scholar] [CrossRef]
- Wildschut, E.D.; van Saet, A.; Pokorna, P.; Ahsman, M.J.; van den Anker, J.N.; Tibboel, D. The impact of extracorporeal life support and hypothermia on drug disposition in critically ill infants and children. Pediatric Clin. 2012, 59, 1183–1204. [Google Scholar] [CrossRef]
- Pea, F. Plasma Pharmacokinetics of Antimicrobial Agents in Critically Ill Patients. Curr. Clin. Pharmacol. 2013, 8, 5–12. [Google Scholar]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Chenthamara, D.; Subramaniam, S.; Ramakrishnan, S.G.; Krishnaswamy, S.; Essa, M.M.; Lin, F.-H.; Qoronfleh, M.W. Therapeutic efficacy of nanoparticles and routes of administration. Biomater. Res. 2019, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Li, M.; Dey, R.; Chen, Y. Nanomaterials for cancer therapy: Current progress and perspectives. J. Hematol. Oncol. 2021, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Morshed, M.; Memic, A.; Hassan, S.; Webster, T.J.; Marei, H.E.-S. Nanoparticles in tissue engineering: Applications, challenges and prospects. Int. J. Nanomed. 2018, 13, 5637–5655. [Google Scholar] [CrossRef] [PubMed]
- Fathi-Achachelouei, M.; Knopf-Marques, H.; Ribeiro da Silva, C.E.; Barthès, J.; Bat, E.; Tezcaner, A.; Vrana, N.E. Use of Nanoparticles in Tissue Engineering and Regenerative Medicine. Front. Bioeng. Biotechnol. 2019, 7, 113. [Google Scholar] [CrossRef]
- Ding, Q.; Cui, J.; Shen, H.; He, C.; Wang, X.; Shen, S.G.; Lin, K. Advances of nanomaterial applications in oral and maxillofacial tissue regeneration and disease treatment. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2021, 13, e1669. [Google Scholar] [CrossRef]
- Siafaka, P.I.; Üstündağ Okur, N.; Karavas, E.; Bikiaris, D.N. Surface modified multifunctional and stimuli responsive nanoparticles for drug targeting: Current status and uses. Int. J. Mol. Sci. 2016, 17, 1440. [Google Scholar] [CrossRef]
- Aberoumandi, S.M.; Mohammadhosseini, M.; Abasi, E.; Saghati, S.; Nikzamir, N.; Akbarzadeh, A.; Panahi, Y.; Davaran, S. An update on applications of nanostructured drug delivery systems in cancer therapy: A review. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1058–1068. [Google Scholar] [CrossRef]
- Mohammadian, F.; Eatemadi, A. Drug loading and delivery using nanofibers scaffolds. Artif. Cells Nanomed. Biotechnol. 2017, 45, 881–888. [Google Scholar] [CrossRef]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.d.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef]
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-based drug delivery in cancer therapy and its role in overcoming drug resistance. Front. Mol. Biosci. 2020, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, H.H.; Holt-Casper, D.; Grainger, D.W.; Ghandehari, H. Nanoparticle Uptake: The Phagocyte Problem. Nano Today 2015, 10, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Sanità, G.; Carrese, B.; Lamberti, A. Nanoparticle surface functionalization: How to improve biocompatibility and cellular internalization. Front. Mol. Biosci. 2020, 381, 587012. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Review nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Moritz, M.; Geszke-Moritz, M. Recent Developments in the Application of Polymeric Nanoparticles as Drug Carriers. Adv. Clin. Exp. Med. 2015, 24, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Banik, B.L.; Fattahi, P.; Brown, J.L. Polymeric nanoparticles: The future of nanomedicine. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2016, 8, 271–299. [Google Scholar] [CrossRef] [PubMed]
- Elmowafy, E.M.; Tiboni, M.; Soliman, M.E. Biocompatibility, biodegradation and biomedical applications of poly (lactic acid)/poly (lactic-co-glycolic acid) micro and nanoparticles. J. Pharm. Investig. 2019, 49, 347–380. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, J.; Wang, W.; Zhang, X.; Zhang, J. Research progress of biodegradable polymers in repairing Achilles tendon injury. Front. Mater. 2022, 2022, 6. [Google Scholar]
- Langer, R.; Folkman, J. Polymers for the sustained release of proteins and other macromolecules. Nature 1976, 263, 797–800. [Google Scholar] [CrossRef]
- Niculescu, A.-G.; Grumezescu, A.M. Polymer-Based Nanosystems—A Versatile Delivery Approach. Materials 2021, 14, 6812. [Google Scholar] [CrossRef]
- Song, R.; Murphy, M.; Li, C.; Ting, K.; Soo, C.; Zheng, Z. Current development of biodegradable polymeric materials for biomedical applications. Drug Des. Dev. Ther. 2018, 12, 3117–3145. [Google Scholar] [CrossRef] [PubMed]
- Namsheer, K.; Rout, C.S. Conducting polymers: A comprehensive review on recent advances in synthesis, properties and applications. RSC Adv. 2021, 11, 5659–5697. [Google Scholar]
- Zielińska, A.; Carreiró, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesh, D.N.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.W.; Santos, J.L.; Williford, J.M.; Mao, H.Q. Control of polymeric nanoparticle size to improve therapeutic delivery. J. Control. Release 2015, 219, 536–547. [Google Scholar] [CrossRef]
- Ventola, C.L. Progress in Nanomedicine: Approved and Investigational Nanodrugs. Pharm. Ther. 2017, 42, 742–755. [Google Scholar]
- Faustino, C.; Pinheiro, L. Lipid Systems for the Delivery of Amphotericin B in Antifungal Therapy. Pharmaceutics 2020, 12, 29. [Google Scholar] [CrossRef]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef]
- Funari, R.; Shen, A.Q. Detection and Characterization of Bacterial Biofilms and Biofilm-Based Sensors. ACS Sens. 2022, 7, 347–357. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef]
- Tits, J.; Cammue, B.P.A.; Thevissen, K. Combination Therapy to Treat Fungal Biofilm-Based Infections. Int. J. Mol. Sci. 2020, 21, 8873. [Google Scholar] [CrossRef]
- Zara, G.; Budroni, M.; Mannazzu, I.; Fancello, F.; Zara, S. Yeast biofilm in food realms: Occurrence and control. World J. Microbiol. Biotechnol. 2020, 36, 134. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Feng, G.; Moraru, C.I. Micro- and Nanotopography Sensitive Bacterial Attachment Mechanisms: A Review. Front. Microbiol. 2019, 10, 191. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial Extracellular Polysaccharides in Biofilm Formation and Function. Microbiol. Spectr. 2015, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Talaiekhozani, A.; Rezania, S.; Kim, K.-H.; Sanaye, R.; Amani, A.M. Recent advances in photocatalytic removal of organic and inorganic pollutants in air. J. Clean. Prod. 2021, 278, 123895. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Klis, F.M. Flocculation, adhesion and biofilm formation in yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; de Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarcón, K.P.; Melo, W.C.M.A.; Marcelino, M.Y.; Braz, J.D.; et al. Fungal Biofilms and Polymicrobial Diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef]
- Pires, R.H.; Martinez, L.R.; Mendes-Giannini, M.J.S.; Stoianoff, M.A.R. Editorial: Pathogenesis of Fungal Biofilms in Different Environmental Conditions and Clinical Outcomes. Front. Cell. Infect. Microbiol. 2021, 11, 778458. [Google Scholar] [CrossRef]
- Harding, M.W.; Marques, L.L.; Howard, R.J.; Olson, M.E. Can filamentous fungi form biofilms? Trends Microbiol. 2009, 17, 475–480. [Google Scholar] [CrossRef]
- Valsecchi, I.; Dupres, V.; Stephen-Victor, E.; Guijarro, J.I.; Gibbons, J.; Beau, R.; Bayry, J.; Coppee, J.-Y.; Lafont, F.; Latgé, J.-P.; et al. Role of Hydrophobins in Aspergillus fumigatus. J. Fungi 2017, 4, 2. [Google Scholar] [CrossRef]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed]
- Costa-Orlandi, C.B.; Sardi, J.C.; Santos, C.T.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. In vitro characterization of Trichophyton rubrum and T. mentagrophytes biofilms. Biofouling 2014, 30, 719–727. [Google Scholar] [CrossRef]
- Pires, R.H.; Montanari, L.B.; Martins, C.H.; Zaia, J.E.; Almeida, A.M.; Matsumoto, M.T.; Mendes-Giannini, M.J. Anticandidal efficacy of cinnamon oil against planktonic and biofilm cultures of Candida parapsilosis and Candida orthopsilosis. Mycopathologia 2011, 172, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.R.; Fries, B.C. Fungal biofilms: Relevance in the setting of human disease. Curr. Fungal Infect. Rep. 2010, 4, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Borghi, E.; Morace, G.; Borgo, F.; Rajendran, R.; Sherry, L.; Nile, C.; Ramage, G. New strategic insights into managing fungal biofilms. Front. Microbiol. 2015, 6, 1077. [Google Scholar] [CrossRef] [PubMed]
- Sardi Jde, C.; Pitangui Nde, S.; Voltan, A.R.; Braz, J.D.; Machado, M.P.; Fusco Almeida, A.M.; Mendes Giannini, M.J. In vitro Paracoccidioides brasiliensis biofilm and gene expression of adhesins and hydrolytic enzymes. Virulence 2015, 6, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Abd Elmegeed, A.S.M.; Ouf, S.A.; Moussa, T.A.A.; Eltahlawi, S.M.R. Dermatophytes and other associated fungi in patients attending to some hospitals in Egypt. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2015, 46, 799–805. [Google Scholar] [CrossRef]
- Burkhart, C.N.; Burkhart, C.G.; Gupta, A.K. Dermatophytoma: Recalcitrance to treatment because of existence of fungal biofilm. J. Am. Acad. Derm. 2002, 47, 629–631. [Google Scholar] [CrossRef]
- Pitangui, N.S.; Sardi, J.C.; Silva, J.F.; Benaducci, T.; Moraes da Silva, R.A.; Rodríguez-Arellanes, G.; Taylor, M.L.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Adhesion of Histoplasma capsulatum to pneumocytes and biofilm formation on an abiotic surface. Biofouling 2012, 28, 711–718. [Google Scholar] [CrossRef]
- Boolchandani, M.; D’Souza, A.W.; Dantas, G. Sequencing-based methods and resources to study antimicrobial resistance. Nat. Rev. Genet. 2019, 20, 356–370. [Google Scholar] [CrossRef]
- Sandhu, C.; Qureshi, A.; Emili, A. Panomics for Precision Medicine. Trends Mol. Med. 2018, 24, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Henriques, M.; Lopez-Ribot, J.L.; Oliveira, R. Addition of DNase improves the in vitro activity of antifungal drugs against Candida albicans biofilms. Mycoses 2012, 55, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Hawser, S.P.; Douglas, L.J. Biofilm formation by Candida species on the surface of catheter materials in vitro. Infect. Immun. 1994, 62, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.H.G.; Pires, R.H.; Cunha, A.O.; Pereira, C.A.M.; Singulani, J.L.; Abrão, F.; Moraes, T.; Mendes-Giannini, M.J.S. Candida/Candida biofilms. First description of dual-species Candida albicans/C. rugosa biofilm. Fungal Biol. 2016, 120, 530–537. [Google Scholar] [CrossRef]
- Řičicová, M.; Kucharíková, S.; Tournu, H.; Hendrix, J.; Bujdáková, H.; van Eldere, J.; Lagrou, K.; van Dijck, P. Candida albicans biofilm formation in a new in vivo rat model. Microbiology 2010, 156 Pt 3, 909–919. [Google Scholar] [CrossRef]
- Kucharíková, S.; Vande Velde, G.; Himmelreich, U.; van Dijck, P. Candida albicans biofilm development on medically-relevant foreign bodies in a mouse subcutaneous model followed by bioluminescence imaging. J. Vis. Exp. 2015, 95, 52239. [Google Scholar] [CrossRef]
- Pires, R.H.; Santos, J.M.; Zaia, J.E.; Martins, C.H.; Mendes-Giannini, M.J. Candida parapsilosis complex water isolates from a haemodialysis unit: Biofilm production and in vitro evaluation of the use of clinical antifungals. Mem. Inst. Oswaldo Cruz 2011, 106, 646–654. [Google Scholar] [CrossRef]
- Bonhomme, J.; d’Enfert, C. Candida albicans biofilms: Building a heterogeneous, drug-tolerant environment. Curr. Opin. Microbiol. 2013, 16, 398–403. [Google Scholar] [CrossRef]
- Xu, C.; Liang, C.; Wullschleger, S.; Wilson, C.; Mcdowell, N. Nature Reviews. Microbiology. Nat. Reviews Microbiol. 2011, 9, 222. [Google Scholar] [CrossRef]
- Paramonova, E.; Krom, B.P.; van der Mei, H.C.; Busscher, H.J.; Sharma, P.K. Hyphal content determines the compression strength of Candida albicans biofilms. Microbiology 2009, 155 Pt 6, 1997–2003. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.R.; Khorram, M.; Barzegar, S.; Asadian, F.; Zareshahrabadi, Z.; Saharkhiz, M.J.; Ahadian, S.; Zomorodian, K. Antimicrobial core–shell electrospun nanofibers containing Ajwain essential oil for accelerating infected wound healing. Int. J. Pharm. 2021, 603, 120698. [Google Scholar] [CrossRef] [PubMed]
- Giti, R.; Zomorodian, K.; Firouzmandi, M.; Zareshahrabadi, Z.; Rahmannasab, S. Antimicrobial activity of thermocycled polymethyl methacrylate resin reinforced with titanium dioxide and copper oxide nanoparticles. Int. J. Dent. 2021, 2021, 6690806. [Google Scholar] [CrossRef]
- Maliszewska, I.; Lisiak, B.; Popko, K.; Matczyszyn, K. Enhancement of the Efficacy of Photodynamic Inactivation of Candida albicans with the Use of Biogenic Gold Nanoparticles. Photochem. Photobiol. 2017, 93, 1081–1090. [Google Scholar] [CrossRef]
- Sherwani, M.A.; Tufail, S.; Khan, A.A.; Owais, M. Gold Nanoparticle-Photosensitizer Conjugate Based Photodynamic Inactivation of Biofilm Producing Cells: Potential for Treatment of C. albicans Infection in BALB/c Mice. PLoS ONE 2015, 10, e0131684. [Google Scholar] [CrossRef]
- Kischkel, B.; Castilho, P.F.; de Oliveira, K.M.; Rezende, P.S.; Bruschi, M.L.; Svidzinski, T.I.; Negri, M. Silver nanoparticles stabilized with propolis show reduced toxicity and potential activity against fungal infections. Future Microbiol. 2020, 15, 521–539. [Google Scholar] [CrossRef]
- Rajendiran, K.; Zhao, Z.; Pei, D.-S.; Fu, A. Antimicrobial Activity and Mechanism of Functionalized Quantum Dots. Polymers 2019, 11, 1670. [Google Scholar] [CrossRef]
- Tejwan, N.; Saini, A.K.; Sharma, A.; Singh, T.A.; Kumar, N.; Das, J. Metal-doped and hybrid carbon dots: A comprehensive review on their synthesis and biomedical applications. J. Control. Release 2021, 330, 132–150. [Google Scholar] [CrossRef]
- Han, X.; Lai, L.; Tian, F.; Jiang, F.-L.; Xiao, Q.; Li, Y.; Yu, Q.; Li, D.; Wang, J.; Zhang, Q.; et al. Toxicity of CdTe Quantum Dots on Yeast Saccharomyces Cerevisiae. Small 2012, 8, 2680–2689. [Google Scholar] [CrossRef]
- Du, H.; Lo, T.-M.; Sitompul, J.; Chang, M.W. Systems-level analysis of Escherichia coli response to silver nanoparticles: The roles of anaerobic respiration in microbial resistance. Biochem. Biophys. Res. Commun. 2012, 424, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Yacamán, M.J.; Ascencio, J.A.; Liu, H.B.; Gardea-Torresdey, J. Structure shape and stability of nanometric sized particles. J. Vac. Sci. Technol. B Microelectron. Nanometer Struct. Process. Meas. Phenom. 2001, 19, 1091–1103. [Google Scholar] [CrossRef]
- Kim, S.; Ryu, D.-Y. Silver nanoparticle-induced oxidative stress, genotoxicity and apoptosis in cultured cells and animal tissues. J. Appl. Toxicol. 2013, 33, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.-K.; Ma, Q.-H.; Li, S.-Y.; Zhang, D.-Q.; Cong, L.; Tian, Y.-L.; Yang, R.-Y. The antifungal effect of silver nanoparticles on Trichosporon asahii. J. Microbiol. Immunol. Infect. 2016, 49, 182–188. [Google Scholar] [CrossRef]
- Feng, Q.L.; Wu, J.; Chen, G.Q.; Cui, F.Z.; Kim, T.N.; Kim, J.O. A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mater. Res. 2000, 52, 662–668. [Google Scholar] [CrossRef]
- Dibrov, P.; Dzioba, J.; Gosink, K.K.; Häse, C.C. Chemiosmotic mechanism of antimicrobial activity of Ag+ in Vibrio cholerae. Antimicrob. Agents Chemother. 2002, 46, 2668–2670. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.; Kwon, S.; Jeong, S. Preparation of biodegradable polymer/silver nanoparticles composite and its antibacterial efficacy. J. Nanosci. Nanotechnol. 2009, 9, 1098–1102. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Paul, S.; Mohanram, K.; Kannan, I. Antifungal activity of curcumin-silver nanoparticles against fluconazole-resistant clinical isolates of Candida species. Ayu 2018, 39, 182–186. [Google Scholar] [CrossRef]
- Qasim Nasar, M.; Zohra, T.; Khalil, A.T.; Saqib, S.; Ayaz, M.; Ahmad, A.; Shinwari, Z.K. Seripheidium quettense mediated green synthesis of biogenic silver nanoparticles and their theranostic applications. Green Chem. Lett. Rev. 2019, 12, 310–322. [Google Scholar] [CrossRef]
- Hassan, A.; Howayda, M.; Mahmoud, H. Effect of zinc oxide nanoparticles on the growth of mycotoxigenic mould. SCPT 2013, 1, 66–74. [Google Scholar]
- Refai, H.; Badawy, M.; Hassan, A.; Sakr, H.; Baraka, Y. Antimicrobial effect of biologically prepared silver nanoparticles (AgNPs) on two different obturator’s soft linings in maxillectomy patients. Eur. J. Acad. Essays 2017, 4, 15–25. [Google Scholar]
- Atef, A.H.; Mogda, K.M.; Mahmoud, H. Biosynthesis of silver nanoparticles (AgNps)(a model of metals) by Candida albicans and its antifungal activity on Some fungal pathogens (Trichophyton mentagrophytes and Candida albicans). N. Y. Sci. J. 2013, 6, 27–43. [Google Scholar]
- Hassan, A.; Abo-Zaid, K.; Oraby, N. Molecular and conventional detection of antimicrobial activity of zinc oxide nanoparticles and cinnamon oil against Escherichia coli and Aspergillus flavus. Adv. Anim. Vet. Sci. 2020, 8, 839–847. [Google Scholar] [CrossRef]
- Radhakrishnan, V.S.; Mudiam, M.K.R.; Kumar, M.; Dwivedi, S.P.; Singh, S.P.; Prasad, T. Silver nanoparticles induced alterations in multiple cellular targets, which are critical for drug susceptibilities and pathogenicity in fungal pathogen (Candida albicans). Int. J. Nanomed. 2018, 13, 2647. [Google Scholar] [CrossRef]
- Yang, X.; Yang, M.; Pang, B.; Vara, M.; Xia, Y. Gold Nanomaterials at Work in Biomedicine. Chem. Rev. 2015, 115, 10410–10488. [Google Scholar] [CrossRef]
- Connor, E.E.; Mwamuka, J.; Gole, A.; Murphy, C.J.; Wyatt, M.D. Gold Nanoparticles Are Taken Up by Human Cells but Do Not Cause Acute Cytotoxicity. Small 2005, 1, 325–327. [Google Scholar] [CrossRef]
- Huang, W.-C.; Tsai, P.-J.; Chen, Y.-C. Functional gold nanoparticles as photothermal agents for selective-killing of pathogenic bacteria. Nanomedicine 2007, 2, 777–787. [Google Scholar] [CrossRef]
- De Alteriis, E.; Maselli, V.; Falanga, A.; Galdiero, S.; Di Lella, F.M.; Gesuele, R.; Guida, M.; Galdiero, E. Efficiency of gold nanoparticles coated with the antimicrobial peptide indolicidin against biofilm formation and development of Candida spp. clinical isolates. Infect. Drug Resist. 2018, 11, 915. [Google Scholar] [CrossRef]
- Wani, I.A.; Ahmad, T. Size and shape dependant antifungal activity of gold nanoparticles: A case study of Candida. Colloids Surf. B. Biointerfaces 2013, 101, 162–170. [Google Scholar] [CrossRef]
- Seong, M.; Lee, D.G. Reactive oxygen species-independent apoptotic pathway by gold nanoparticles in Candida albicans. Microbiol. Res. 2018, 207, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Li, Q.; Wang, J.; Yu, Y.; Wang, Y.; Zhou, Q.; Li, P. Reactive Oxygen Species-Related Nanoparticle Toxicity in the Biomedical Field. Nanoscale Res. Lett. 2020, 15, 115. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Li, J.; Zhang, Y.; Wang, Y.; Liu, L.; Li, M. Inhibition of gold nanoparticles (AuNPs) on pathogenic biofilm formation and invasion to host cells. Sci. Rep. 2016, 6, 26667. [Google Scholar] [CrossRef] [PubMed]
- Sojinrin, T.; Conde, J.; Liu, K.; Curtin, J.; Byrne, H.J.; Cui, D.; Tian, F. Plasmonic gold nanoparticles for detection of fungi and human cutaneous fungal infections. Anal. Bioanal. Chem. 2017, 409, 4647–4658. [Google Scholar] [CrossRef]
- Chow, T.H.; Li, N.; Bai, X.; Zhuo, X.; Shao, L.; Wang, J. Gold Nanobipyramids: An Emerging and Versatile Type of Plasmonic Nanoparticles. Acc. Chem. Res. 2019, 52, 2136–2146. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Hashim, A.F.; Alghuthaymi, M.A.; Said-Galiev, E. Nanobiotechnological strategies for toxigenic fungi and mycotoxin control. In Food Preservation; Elsevier: Amsterdam, The Netherlands, 2017; pp. 337–364. [Google Scholar]
- Joshaghani, H.; Eskandari, M. Antifungal effect of Sodium Dodecil Sulfate and Nano particle ZnO on growth inhibition of standard strain of Candida albicans. J. Gorgan Univ. Med. Sci. 2011, 12, 64–69. [Google Scholar]
- Hernández-Meléndez, D.; Salas-Téllez, E.; Zavala-Franco, A.; Téllez, G.; Méndez-Albores, A.; Vázquez-Durán, A. Inhibitory effect of flower-shaped zinc oxide nanostructures on the growth and aflatoxin production of a highly toxigenic strain of Aspergillus flavus Link. Materials 2018, 11, 1265. [Google Scholar] [CrossRef]
- Rostamizadeh, S.; Shadjou, N.; Amani, A.M.; Balalaie, S. Silica supported sodium hydrogen sulfate (NaHSO4/SiO2): A mild and efficient reusable catalyst for the synthesis of aryl-14-H-dibenzo [a, j] xanthenes under solvent-free conditions. Chin. Chem. Lett. 2008, 19, 1151–1155. [Google Scholar] [CrossRef]
- Nabawy, G.A.; Hassan, A.A.; Sayed El-Ahl, R.; Refai, M.K. Effect of metal nanoparticles in comparison with commercial antifungal feed additives on the growth of Aspergillus flavus and aflatoxin b1 production. J. Glob. Biosci. 2014, 3, 954–971. [Google Scholar]
- Mousavi, M.; Hashemi, A.; Arjmand, O.; Amani, A.M.; Babapoor, A.; Fateh, M.A.; Fateh, H.; Mojoudi, F.; Esmaeili, H.; Jahandideh, S. Erythrosine Adsorption from Aqueous Solution via Decorated Graphene Oxide with Magnetic Iron Oxide Nano Particles: Kinetic and Equilibrium Studies. Acta Chim. Slov. 2018, 65, 882–894. [Google Scholar] [CrossRef]
- Mouhamed, A.E.; Hassan, A.A.; Hassan, M.; El Hariria, M.; Refai, M. Effect of metal nanoparticles on the growth of ochratoxigenic moulds and ochratoxin A production isolated from food and feed. Int. J. Res. Stud. Biosci 2015, 3, 1–14. [Google Scholar]
- Ashraf, A.; Ahmed, M.; Diasty, E.; Fatma, I.; Ahmed Youssef, M. A comparative study on antifungal activity of FE2O3, and FE3O4 nanoparticles. Int. J. Adv. Res. 2018, 6, 189–194. [Google Scholar]
- Kheiri, A.; Jorf, S.M.; Malihipour, A.; Saremi, H.; Nikkhah, M. Application of chitosan and chitosan nanoparticles for the control of Fusarium head blight of wheat (Fusarium graminearum) in vitro and greenhouse. Int. J. Biol. Macromol. 2016, 93, 1261–1272. [Google Scholar] [CrossRef]
- Ahmed, F.; Soliman, F.M.; Adly, M.A.; Soliman, H.A.; El-Matbouli, M.; Saleh, M. In vitro assessment of the antimicrobial efficacy of chitosan nanoparticles against major fish pathogens and their cytotoxicity to fish cell lines. J. Fish Dis. 2020, 43, 1049–1063. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Alghuthaymi, M.A.; Shami, A.; Rubina, M.S.; Abramchuk, S.S.; Shtykova, E.V.; Yu. Vasil’kov, A. Copper-chitosan nanocomposite hydrogels against aflatoxigenic Aspergillus flavus from dairy cattle feed. J. Fungi 2020, 6, 112. [Google Scholar] [CrossRef]
- Anaraki, M.R.; Jangjoo, A.; Alimoradi, F.; Dizaj, S.M.; Lotfipour, F. Comparison of antifungal properties of acrylic resin reinforced with ZnO and Ag nanoparticles. Pharm. Sci. 2017, 23, 207–214. [Google Scholar] [CrossRef]
- Hassan, A.A.; Sayed-Elahl, R.M.; Oraby, N.H.; El-Hamaky, A.M. Metal nanoparticles for management of mycotoxigenic fungi and mycotoxicosis diseases of animals and poultry. In Nanomycotoxicology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 251–269. [Google Scholar]
- Hassan, A.A.; Mansour, M.K.; El Hamaky, A.M.; El Ahl, R.M.S.; Oraby, N.H. Nanomaterials and nanocomposite applications in veterinary medicine. In Multifunctional Hybrid Nanomaterials for Sustainable Agri-Food and Ecosystems; Elsevier: Amsterdam, The Netherlands, 2020; pp. 583–638. [Google Scholar]
- Atef, H.A.; Mansour, M.K.; Ibrahim, E.M.; Sayed El-Ahl, R.; Al-Kalamawey, N.M.; El Kattan, Y.; Ali, M. Efficacy of zinc oxide nanoparticles and curcumin in amelioration the toxic effects in aflatoxicated rabbits. Int. J. Curr. Microbiol. Appl. Sci 2016, 5, 795–818. [Google Scholar] [CrossRef]
- Sanchez, V.C.; Jachak, A.; Hurt, R.H.; Kane, A.B. Biological interactions of graphene-family nanomaterials: An interdisciplinary review. Chem Res. Toxicol. 2012, 25, 15–34. [Google Scholar] [CrossRef]
- Wang, J.-J.; Liu, B.-H.; Hsu, Y.-T.; Yu, F.-Y. Sensitive competitive direct enzyme-linked immunosorbent assay and gold nanoparticle immunochromatographic strip for detecting aflatoxin M1 in milk. Food Control 2011, 22, 964–969. [Google Scholar] [CrossRef]
- Osama, E.; El-Sheikh, S.M.; Khairy, M.H.; Galal, A.A. Nanoparticles and their potential applications in veterinary medicine. J. Adv. Vet. Res. 2020, 10, 268–273. [Google Scholar]
- Niemirowicz, K.; Durnaś, B.; Tokajuk, G.; Głuszek, K.; Wilczewska, A.Z.; Misztalewska, I.; Mystkowska, J.; Michalak, G.; Sodo, A.; Wątek, M.; et al. Magnetic nanoparticles as a drug delivery system that enhance fungicidal activity of polyene antibiotics. Nanomedicine 2016, 12, 2395–2404. [Google Scholar] [CrossRef] [PubMed]
- Niemirowicz, K.; Durnaś, B.; Tokajuk, G.; Piktel, E.; Michalak, G.; Gu, X.; Kułakowska, A.; Savage, P.B.; Bucki, R. Formulation and candidacidal activity of magnetic nanoparticles coated with cathelicidin LL-37 and ceragenin CSA-13. Sci. Rep. 2017, 7, 4610. [Google Scholar] [CrossRef]
- Carvalho, G.C.; Sábio, R.M.; de Cássia Ribeiro, T.; Monteiro, A.S.; Pereira, D.V.; Ribeiro, S.J.L.; Chorilli, M. Highlights in mesoporous silica nanoparticles as a multifunctional controlled drug delivery nanoplatform for infectious diseases treatment. Pharm. Res. 2020, 37, 191. [Google Scholar] [CrossRef] [PubMed]
- Mahdavinia, G.H.; Rostamizadeh, S.; Amani, A.M.; Sepehrian, H. Fast and efficient method for the synthesis of 2-arylbenzimidazoles using MCM-41-SO3H. Heterocycl. Commun. 2012, 18, 33–37. [Google Scholar] [CrossRef]
- Rostamizadeh, S.; Amani, A.M.; Mahdavinia, G.H.; Shadjou, N. Silica supported ammonium dihydrogen phosphate (NH4H2PO4/SiO2): A mild, reusable and highly efficient heterogeneous catalyst for the synthesis of 14-aryl-14-H-dibenzo [a, j] xanthenes. Chin. Chem. Lett. 2009, 20, 779–783. [Google Scholar] [CrossRef]
- Vallet-Regí, M. Mesoporous silica nanoparticles: Their projection in nanomedicine. Int. Sch. Res. Not. 2012, 2012, 608548. [Google Scholar] [CrossRef]
- Kanugala, S.; Jinka, S.; Puvvada, N.; Banerjee, R.; Kumar, C.G. Phenazine-1-carboxamide functionalized mesoporous silica nanoparticles as antimicrobial coatings on silicone urethral catheters. Sci. Rep. 2019, 9, 6198. [Google Scholar] [CrossRef]
- Montazeri, M.; Razzaghi-Abyaneh, M.; Nasrollahi, S.A.; Maibach, H.; Nafisi, S. Enhanced topical econazole antifungal efficacy by amine-functionalized silica nanoparticles. Bull. Mater. Sci. 2019, 43, 13. [Google Scholar] [CrossRef]
- Gibney, K.A.; Sovadinova, I.; Lopez, A.I.; Urban, M.; Ridgway, Z.; Caputo, G.A.; Kuroda, K. Poly (ethylene imine) s as antimicrobial agents with selective activity. Macromol. Biosci. 2012, 12, 1279–1289. [Google Scholar] [CrossRef]
- Dodou Lima, H.V.; de Paula Cavalcante, C.S.; Rádis-Baptista, G. Antifungal in vitro activity of pilosulin-and ponericin-like peptides from the giant ant Dinoponera quadriceps and synergistic effects with antimycotic drugs. Antibiotics 2020, 9, 354. [Google Scholar] [CrossRef]
- Ramamourthy, G.; Park, J.; Seo, C.J.; Vogel, H.; Park, Y. Antifungal and antibiofilm activities and the mechanism of action of repeating lysine-tryptophan peptides against candida albicans. Microorganisms 2020, 8, 758. [Google Scholar] [CrossRef] [PubMed]
- Rank, L.A.; Walsh, N.M.; Lim, F.Y.; Gellman, S.H.; Keller, N.P.; Hull, C.M. Peptide-like nylon-3 polymers with activity against phylogenetically diverse, intrinsically drug-resistant pathogenic fungi. Msphere 2018, 3, e00223-18. [Google Scholar] [CrossRef] [PubMed]
- Codling, C.E.; Maillard, J.-Y.; Russell, A.D. Aspects of the antimicrobial mechanisms of action of a polyquaternium and an amidoamine. J. Antimicrob. Chemother. 2003, 51, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Jiang, Z.; Zhang, N.; Yang, Z.; Zhou, Z. Interactions of biocidal polyhexamethylene guanidine hydrochloride and its analogs with POPC model membranes. Polymers 2017, 9, 517. [Google Scholar] [CrossRef]
- Azevedo, M.; Ramalho, P.; Silva, A.; Teixeira-Santos, R.; Pina-Vaz, C.; Rodrigues, A. Polyethyleneimine and polyethyleneimine-based nanoparticles: Novel bacterial and yeast biofilm inhibitors. J. Med. Microbiol. 2014, 63, 1167–1173. [Google Scholar] [CrossRef]
- Jung, J.; Bae, Y.; Cho, Y.K.; Ren, X.; Sun, Y. Structural insights into conformation of amphiphilic quaternary ammonium chitosans to control fungicidal and anti-biofilm functions. Carbohydr. Polym. 2020, 228, 115391. [Google Scholar] [CrossRef]
- Hoque, J.; Adhikary, U.; Yadav, V.; Samaddar, S.; Konai, M.M.; Prakash, R.G.; Paramanandham, K.; Shome, B.R.; Sanyal, K.; Haldar, J. Chitosan derivatives active against multidrug-resistant bacteria and pathogenic fungi: In vivo evaluation as topical antimicrobials. Mol. Pharm. 2016, 13, 3578–3589. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Lopez-Jimenez, J.; Pérez-Berná, A.; Huang, I.C.; Jansson, H.B.; Salinas, J.; Villalaín, J.; Read, N.; Lopez-Llorca, L. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol. Microbiol. 2010, 75, 1021–1032. [Google Scholar] [CrossRef]
- Olmedo, G.M.; Cerioni, L.; Sepulveda, M.; Ramallo, J.; Rapisarda, V.A.; Volentini, S.I. Polyhexamethylene guanidine as a fungicide, disinfectant and wound protector in lemons challenged with Penicillium digitatum. Food Microbiol. 2018, 76, 128–134. [Google Scholar] [CrossRef]
- Spadari, C.d.C.; Wirth, F.; Lopes, L.B.; Ishida, K. New Approaches for Cryptococcosis Treatment. Microorganisms 2020, 8, 613. [Google Scholar] [CrossRef]
- Pedroso, L.S.; Khalil, N.M.; Mainardes, R.M. Preparation and In vitro Evaluation of Efficacy and Toxicity of Polysorbate 80-coated Bovine Serum Albumin Nanoparticles containing Amphotericin B. Curr. Drug Deliv. 2018, 15, 1055–1063. [Google Scholar] [CrossRef]
- Xu, N.; Gu, J.; Zhu, Y.; Wen, H.; Ren, Q.; Chen, J. Efficacy of intravenous amphotericin B-polybutylcyanoacrylate nanoparticles against cryptococcal meningitis in mice. Int. J. Nanomed. 2011, 6, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Ramge, P.; Unger, R.E.; Oltrogge, J.B.; Zenker, D.; Begley, D.; Kreuter, J.; Von Briesen, H. Polysorbate-80 coating enhances uptake of polybutylcyanoacrylate (PBCA)-nanoparticles by human and bovine primary brain capillary endothelial cells. Eur. J. Neurosci. 2000, 12, 1931–1940. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Xu, N.; Cao, C.; Yuan, W.; Yu, X.; Chen, J.; Ren, J. Preparation and therapeutic efficacy of polysorbate-80-coated amphotericin B/PLA-b-PEG nanoparticles. J. Biomater. Sci. Polym. Ed. 2009, 20, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Radwan, M.A.; AlQuadeib, B.T.; Šiller, L.; Wright, M.C.; Horrocks, B. Oral administration of amphotericin B nanoparticles: Antifungal activity, bioavailability and toxicity in rats. Drug Deliv. 2017, 24, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.E.; Saad, G.R.; Sabaa, M.W. Synthesis, characterization, and biological activity of cross-linked chitosan biguanidine loaded with silver nanoparticles. J. Biomater. Sci. Polym. Ed. 2016, 27, 1880–1898. [Google Scholar] [CrossRef]
- Yang, C.; Xue, B.; Song, W.; Kan, B.; Zhang, D.; Yu, H.; Shen, N.; Li, X.; Tang, Z.; Chen, X. Reducing the toxicity of amphotericin B by encapsulation using methoxy poly(ethylene glycol)-b-poly(l-glutamic acid-co-l-phenylalanine). Biomater. Sci. 2018, 6, 2189–2196. [Google Scholar] [CrossRef] [PubMed]
- Italia, J.L.; Sharp, A.; Carter, K.C.; Warn, P.; Kumar, M.N. Peroral amphotericin B polymer nanoparticles lead to comparable or superior in vivo antifungal activity to that of intravenous Ambisome® or Fungizone™. PLoS ONE 2011, 6, e25744. [Google Scholar]
- Van de Ven, H.; Paulussen, C.; Feijens, P.B.; Matheeussen, A.; Rombaut, P.; Kayaert, P.; van den Mooter, G.; Weyenberg, W.; Cos, P.; Maes, L.; et al. PLGA nanoparticles and nanosuspensions with amphotericin B: Potent in vitro and in vivo alternatives to Fungizone and AmBisome. J. Control. Release 2012, 161, 795–803. [Google Scholar] [CrossRef]
- Deaguero, I.G.; Huda, M.N.; Rodriguez, V.; Zicari, J.; Al-Hilal, T.A.; Badruddoza, A.Z.M.; Nurunnabi, M. Nano-vesicle based anti-fungal formulation shows higher stability, skin diffusion, biosafety and anti-fungal efficacy in vitro. Pharmaceutics 2020, 12, 516. [Google Scholar] [CrossRef]
- Siopi, M.; Mouton, J.W.; Pournaras, S.; Meletiadis, J. In vitro and in vivo exposure-effect relationship of liposomal amphotericin b against aspergillus fumigatus. Antimicrob. Agents Chemother. 2019, 63, e02673-18. [Google Scholar] [CrossRef] [PubMed]
- Spadari, C.C.; de Bastiani, F.; Lopes, L.B.; Ishida, K. Alginate nanoparticles as non-toxic delivery system for miltefosine in the treatment of candidiasis and cryptococcosis. Int. J. Nanomed. 2019, 14, 5187–5199. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Cha, Y.J.; Kim, H.; Choi, S.S. Antifungal activity of nano and micro charcoal particle polymers against Paecilomyces variotii, Trichoderma virens and Chaetomium globosum. New Biotechnol. 2016, 33, 55–60. [Google Scholar] [CrossRef]
- Hassan, A.A.; Mansour, M.K.; El Ahl, R.M.S.; El Hamaky, A.M.; Oraby, N.H. Toxic and beneficial effects of carbon nanomaterials on human and animal health. In Carbon Nanomaterials for Agri-Food and Environmental Applications; Elsevier: Amsterdam, The Netherlands, 2020; pp. 535–555. [Google Scholar]
- Venkatesan, J.; Jayakumar, R.; Mohandas, A.; Bhatnagar, I.; Kim, S.K. Antimicrobial Activity of Chitosan-Carbon Nanotube Hydrogels. Materials 2014, 7, 3946–3955. [Google Scholar] [CrossRef] [PubMed]
- Darbari, S.; Abdi, Y.; Haghighi, F.; Mohajerzadeh, S.; Haghighi, N. Investigating the antifungal activity of TiO2 nanoparticles deposited on branched carbon nanotube arrays. J. Phys. D Appl. Phys. 2011, 44, 245401. [Google Scholar] [CrossRef]
- Vafa, E.; Bazargan-Lari, R. Bovine serum albumin protected gold nanozymes as a novel anti-cancer nanodrug for acute T-type lymphoblastic leukemia treatment via effect on the expression of anti-apoptotic genes. Appl. Biol. Chem. 2021, 64, 86. [Google Scholar] [CrossRef]
- Ezati, P.; Rhim, J.-W.; Molaei, R.; Priyadarshi, R.; Roy, S.; Min, S.; Kim, Y.H.; Lee, S.-G.; Han, S. Preparation and characterization of B, S, and N-doped glucose carbon dots: Antibacterial, antifungal, and antioxidant activity. Sustain. Mater. Technol. 2022, 32, e00397. [Google Scholar] [CrossRef]
- Leudjo Taka, A.; Doyle, B.P.; Carleschi, E.; Youmbi Fonkui, T.; Erasmus, R.; Fosso-Kankeu, E.; Pillay, K.; Mbianda, X.Y. Spectroscopic characterization and antimicrobial activity of nanoparticle doped cyclodextrin polyurethane bionanosponge. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 115, 111092. [Google Scholar] [CrossRef]
- Khan, S.; Baboota, S.; Ali, J.; Khan, S.; Narang, R.S.; Narang, J.K. Nanostructured lipid carriers: An emerging platform for improving oral bioavailability of lipophilic drugs. Int. J. Pharm. Investig. 2015, 5, 182–191. [Google Scholar]
- Soliman, G.M. Nanoparticles as safe and effective delivery systems of antifungal agents: Achievements and challenges. Int. J. Pharm. 2017, 523, 15–32. [Google Scholar] [CrossRef]
- Salvi, V.R.; Pawar, P. Nanostructured lipid carriers (NLC) system: A novel drug targeting carrier. J. Drug Deliv. Sci. Technol. 2019, 51, 255–267. [Google Scholar] [CrossRef]
- El-Sheridy, N.A.; Ramadan, A.A.; Eid, A.A.; El-Khordagui, L.K. Itraconazole lipid nanocapsules gel for dermatological applications: In vitro characteristics and treatment of induced cutaneous candidiasis. Colloids Surf. B Biointerfaces 2019, 181, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Beloqui, A.; Solinís, M.A.; Delgado, A.; Evora, C.; del Pozo-Rodríguez, A.; Rodríguez-Gascón, A. Biodistribution of Nanostructured Lipid Carriers (NLCs) after intravenous administration to rats: Influence of technological factors. Eur. J. Pharm. Biopharm. 2013, 84, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Jansook, P.; Pichayakorn, W.; Ritthidej, G.C. Amphotericin B-loaded solid lipid nanoparticles (SLNs) and nanostructured lipid carrier (NLCs): Effect of drug loading and biopharmaceutical characterizations. Drug Dev. Ind. Pharm. 2018, 44, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Kelidari, H.R.; Moazeni, M.; Babaei, R.; Saeedi, M.; Akbari, J.; Parkoohi, P.I.; Nabili, M.; Gohar, A.A.; Morteza-Semnani, K.; Nokhodchi, A. Improved yeast delivery of fluconazole with a nanostructured lipid carrier system. Biomed. Pharm. 2017, 89, 83–88. [Google Scholar] [CrossRef]
- Moazeni, M.; Kelidari, H.R.; Saeedi, M.; Morteza-Semnani, K.; Nabili, M.; Gohar, A.A.; Akbari, J.; Lotfali, E.; Nokhodchi, A. Time to overcome fluconazole resistant Candida isolates: Solid lipid nanoparticles as a novel antifungal drug delivery system. Colloids Surf. B. Biointerfaces 2016, 142, 400–407. [Google Scholar] [CrossRef]
- Domingues Bianchin, M.; Borowicz, S.M.; da Rosa Monte Machado, G.; Pippi, B.; Stanisçuaski Guterres, S.; Raffin Pohlmann, A.; Meneghello Fuentefria, A.; Clemes Külkamp-Guerreiro, I. Lipid core nanoparticles as a broad strategy to reverse fluconazole resistance in multiple Candida species. Colloids Surf. B Biointerfaces 2019, 175, 523–529. [Google Scholar] [CrossRef]
- Gupta, M.; Tiwari, S.; Vyas, S.P. Influence of various lipid core on characteristics of SLNs designed for topical delivery of fluconazole against cutaneous candidiasis. Pharm. Dev. Technol. 2013, 18, 550–559. [Google Scholar] [CrossRef]
- Gupta, M.; Vyas, S.P. Development, characterization and in vivo assessment of effective lipidic nanoparticles for dermal delivery of fluconazole against cutaneous candidiasis. Chem. Phys. Lipids 2012, 165, 454–461. [Google Scholar] [CrossRef]
- Kim, J.-K.; Park, J.-S.; Kim, C.-K. Development of a binary lipid nanoparticles formulation of itraconazole for parenteral administration and controlled release. Int. J. Pharm. 2010, 383, 209–215. [Google Scholar] [CrossRef]
- Mirza, M.A.; Panda, A.K.; Asif, S.; Verma, D.; Talegaonkar, S.; Manzoor, N.; Khan, A.; Ahmed, F.J.; Dudeja, M.; Iqbal, Z. A vaginal drug delivery model. Drug Deliv. 2016, 23, 3123–3134. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.; Majumdar, D.K.; Mishra, S.K.; Panda, A.K.; Patnaik, S. Development and characterization of itraconazole-loaded solid lipid nanoparticles for ocular delivery. Pharm. Dev. Technol. 2015, 20, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.M.; Rajinikanth, P.S.; Mallikarjun, C.; Kang, Y.B. Formulation and delivery of itraconazole to the brain using a nanolipid carrier system. Int. J. Nanomed. 2014, 9, 2117. [Google Scholar] [CrossRef] [PubMed]
- Pardeike, J.; Weber, S.; Zarfl, H.P.; Pagitz, M.; Zimmer, A. Itraconazole-loaded nanostructured lipid carriers (NLC) for pulmonary treatment of aspergillosis in falcons. Eur. J. Pharm. Biopharm. 2016, 108, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Cassano, R.; Ferrarelli, T.; Mauro, M.V.; Cavalcanti, P.; Picci, N.; Trombino, S. Preparation, characterization and in vitro activities evaluation of solid lipid nanoparticles based on PEG-40 stearate for antifungal drugs vaginal delivery. Drug Deliv. 2016, 23, 1037–1046. [Google Scholar] [CrossRef]
- Das, S.; Ng, W.K.; Tan, R.B. Sucrose ester stabilized solid lipid nanoparticles and nanostructured lipid carriers: I. Effect of formulation variables on the physicochemical properties, drug release and stability of clotrimazole-loaded nanoparticles. Nanotechnology 2014, 25, 105101. [Google Scholar] [CrossRef]
- Das, S.; Ng, W.K.; Tan, R.B. Are nanostructured lipid carriers (NLCs) better than solid lipid nanoparticles (SLNs): Development, characterizations and comparative evaluations of clotrimazole-loaded SLNs and NLCs? Eur. J. Pharm. Sci. 2012, 47, 139–151. [Google Scholar] [CrossRef]
- Esposito, E.; Ravani, L.; Contado, C.; Costenaro, A.; Drechsler, M.; Rossi, D.; Menegatti, E.; Grandini, A.; Cortesi, R. Clotrimazole nanoparticle gel for mucosal administration. Mater. Sci. Eng. C 2013, 33, 411–418. [Google Scholar] [CrossRef]
- Vaghasiya, H.; Kumar, A.; Sawant, K. Development of solid lipid nanoparticles based controlled release system for topical delivery of terbinafine hydrochloride. Eur. J. Pharm. Sci. 2013, 49, 311–322. [Google Scholar] [CrossRef]
- Kumar, R.; Sinha, V.R. Solid lipid nanoparticle: An efficient carrier for improved ocular permeation of voriconazole. Drug Dev. Ind. Pharm. 2016, 42, 1956–1967. [Google Scholar] [CrossRef]
- Khare, A.; Singh, I.; Pawar, P.; Grover, K. Design and evaluation of voriconazole loaded solid lipid nanoparticles for ophthalmic application. J. Drug Deliv. 2016, 2016, 6590361. [Google Scholar] [CrossRef] [PubMed]
- Song, S.H.; Lee, K.M.; Kang, J.B.; Lee, S.G.; Kang, M.J.; Choi, Y.W. Improved skin delivery of voriconazole with a nanostructured lipid carrier-based hydrogel formulation. Chem. Pharm. Bull. 2014, 62, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Bhalekar, M.R.; Pokharkar, V.; Madgulkar, A.; Patil, N.; Patil, N. Preparation and evaluation of miconazole nitrate-loaded solid lipid nanoparticles for topical delivery. AAPS Pharmscitech 2009, 10, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Aljaeid, B.M.; Hosny, K.M. Miconazole-loaded solid lipid nanoparticles: Formulation and evaluation of a novel formula with high bioavailability and antifungal activity. Int. J. Nanomed. 2016, 11, 441. [Google Scholar] [CrossRef]
- Jain, S.; Jain, S.; Khare, P.; Gulbake, A.; Bansal, D.; Jain, S.K. Design and development of solid lipid nanoparticles for topical delivery of an anti-fungal agent. Drug Deliv. 2010, 17, 443–451. [Google Scholar] [CrossRef]
- Sanna, V.; Gavini, E.; Cossu, M.; Rassu, G.; Giunchedi, P. Solid lipid nanoparticles (SLN) as carriers for the topical delivery of econazole nitrate: In-vitro characterization, ex-vivo and in-vivo studies. J. Pharm. Pharmacol. 2007, 59, 1057–1064. [Google Scholar] [CrossRef]
- Samanta, P.K. Strong and weak quantum confinement and size dependent optoelectronic properties of zinc oxide. Ann. Univ. Craiova Phys. 2018, 28, 17–23. [Google Scholar]
- Siddiqi, K.S.; Husen, A. Properties of zinc oxide nanoparticles and their activity against microbes. Nanoscale Res. Lett. 2018, 13, 141. [Google Scholar] [CrossRef]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef]
- Manikandan, V.; Balasubramanian, B.; Bharti, B.; Velmurugan, P.; Elango, D.; Baskaran, R.; Jayanthi, P. Development of ZnO/MOGAC nanocomposites for enhanced photocatalytic removal of PO43− and NO3-ions from wastewater under various light irradiations. Biomass Convers. Biorefin. 2022, 1–18. [Google Scholar] [CrossRef]
- Lallo da Silva, B.; Caetano, B.L.; Chiari-Andréo, B.G.; Pietro, R.; Chiavacci, L.A. Increased antibacterial activity of ZnO nanoparticles: Influence of size and surface modification. Colloids Surf. B Biointerfaces 2019, 177, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Asok, A.; Ghosh, S.; More, P.A.; Chopade, B.A.; Gandhi, M.N.; Kulkarni, A.R. Surface defect rich ZnO quantum dots as antioxidants inhibiting α-amylase and α-glucosidase: A potential anti-diabetic nanomedicine. J. Mater. Chem. B 2015, 3, 4597–4606. [Google Scholar] [CrossRef] [PubMed]
- Chand, P.; Kumari, S.; Mondal, N.; Singh, S.P.; Prasad, T. Synergism of Zinc Oxide Quantum Dots with Antifungal Drugs: Potential Approach for Combination Therapy against Drug Resistant Candida albicans. Front. Nanotechnol. 2021, 3, 624564. [Google Scholar] [CrossRef]
- Fakhroueian, Z.; Dehshiri, A.M.; Katouzian, F.; Esmaeilzadeh, P. In vitro cytotoxic effects of modified zinc oxide quantum dots on breast cancer cell lines (MCF7), colon cancer cell lines (HT29) and various fungi. J. Nanopart. Res. 2014, 16, 2483. [Google Scholar] [CrossRef]
- Chand, P.; Singh, S.P.; Prasad, T. Effect of Antioxidant on ROS Mediated Antifungal Action of ZnO Quantum Dots in Candida albicans. ECS Trans. 2022, 107, 6621. [Google Scholar] [CrossRef]
- Fernández de Ullivarri, M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal peptides as therapeutic agents. Front. Cell Infect. Microbiol. 2020, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef]
- Scheenstra, M.R.; van Harten, R.M.; Veldhuizen, E.J.; Haagsman, H.P.; Coorens, M. Cathelicidins modulate TLR-activation and inflammation. Front. Immunol. 2020, 11, 1137. [Google Scholar] [CrossRef]
- Łoboda, D.; Kozłowski, H.; Rowińska-Żyrek, M. Antimicrobial peptide–metal ion interactions—A potential way of activity enhancement. New J. Chem. 2018, 42, 7560–7568. [Google Scholar] [CrossRef]
- Harris, F.; Dennison, R.S.; Phoenix, A.D. Anionic Antimicrobial Peptides from Eukaryotic Organisms. Curr. Protein Pept. Sci. 2009, 10, 585–606. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bonilla, A.; Fernández-García, M. Polymeric materials with antimicrobial activity. Prog. Polym. Sci. 2012, 37, 281–339. [Google Scholar] [CrossRef]
- Kodedová, M.; Valachovič, M.; Csáky, Z.; Sychrová, H. Variations in yeast plasma-membrane lipid composition affect killing activity of three families of insect antifungal peptides. Cell Microbiol. 2019, 21, e13093. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yuan, L.; Han, L.; Li, S.; Zhou, W. The preparation, characterization, anti-ultraviolet and antimicrobial activity of gelatin film incorporated with berberine-HP-β-CD. Colloids Surf. Physicochem. Eng. Asp. 2020, 586, 124273. [Google Scholar] [CrossRef]
- Firmino, H.C.T.; Nascimento, E.P.; Bonan, R.F.; Maciel, P.P.; Castellano, L.R.C.; Santana, L.N.L.; Neves, G.A.; Menezes, R.R. Antifungal activity of TiO2-CeO2 nanofibers against Candida fungi. Mater. Lett. 2021, 283, 128709. [Google Scholar] [CrossRef]
- Salaberria, A.M.; Fernandes, S.C.M.; Diaz, R.H.; Labidi, J. Processing of α-chitin nanofibers by dynamic high pressure homogenization: Characterization and antifungal activity against A. niger. Carbohydr. Polym. 2015, 116, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Ifuku, S.; Tsukiyama, Y.; Yukawa, T.; Egusa, M.; Kaminaka, H.; Izawa, H.; Morimoto, M.; Saimoto, H. Facile preparation of silver nanoparticles immobilized on chitin nanofiber surfaces to endow antifungal activities. Carbohydr. Polym. 2015, 117, 813–817. [Google Scholar] [CrossRef]
- Semnani, K.; Shams-Ghahfarokhi, M.; Afrashi, M.; Fakhrali, A.; Semnani, D. Antifungal activity of eugenol loaded electrospun PAN nanofiber mats against Candida albicans. Curr. Drug Del. 2018, 15, 860–866. [Google Scholar] [CrossRef]
- Fouda, S.M.; Gad, M.M.; Ellakany, P.; Al-Thobity, A.M.; Al-Harbi, F.A.; Virtanen, J.I.; Raustia, A. The effect of nanodiamonds on candida albicans adhesion and surface characteristics of PMMA denture base material-an in vitro study. J. Appl. Oral Sci. 2019, 27, 1–10. [Google Scholar] [CrossRef]
- De Matteis, V.; Cascione, M.; Toma, C.C.; Albanese, G.; de Giorgi, M.L.; Corsalini, M.; Rinaldi, R. Silver nanoparticles addition in poly (methyl methacrylate) dental matrix: Topographic and antimycotic studies. Int. J. Mol. Sci. 2019, 20, 4691. [Google Scholar] [CrossRef]
- Bahadar, H.; Maqbool, F.; Niaz, K.; Abdollahi, M. Toxicity of Nanoparticles and an Overview of Current Experimental Models. Iran. Biomed. J. 2016, 20, 1–11. [Google Scholar] [PubMed]
- Kumar, V.; Sharma, N.; Maitra, S.S. In vitro and in vivo toxicity assessment of nanoparticles. Int. Nano Lett. 2017, 7, 243–256. [Google Scholar] [CrossRef]
- Klein, R.; Nagy, O.; Tóthová, C.; Chovanová, F. Clinical and Diagnostic Significance of Lactate Dehydrogenase and Its Isoenzymes in Animals. Vet. Med. Int. 2020, 2020, 5346483. [Google Scholar] [CrossRef] [PubMed]
- Accomasso, L.; Cristallini, C.; Giachino, C. Risk Assessment and Risk Minimization in Nanomedicine: A Need for Predictive, Alternative, and 3Rs Strategies. Front. Pharmacol. 2018, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Badman, R.P.; Moore, S.L.; Killian, J.L.; Feng, T.; Cleland, T.A.; Hu, F.; Wang, M.D. Dextran-coated iron oxide nanoparticle-induced nanotoxicity in neuron cultures. Sci. Rep. 2020, 10, 11239. [Google Scholar] [CrossRef]
- Wiesner, M.R.; Lowry, G.V.; Jones, K.L.; Hochella, J.; Michael, F.; Di Giulio, R.T.; Casman, E.; Bernhardt, E.S. Decreasing uncertainties in assessing environmental exposure, risk, and ecological implications of nanomaterials. ACS Publ. 2009, 2009, 6458–6462. [Google Scholar] [CrossRef]
- Drasler, B.; Sayre, P.; Steinhäuser, K.G.; Petri-Fink, A.; Rothen-Rutishauser, B. In vitro approaches to assess the hazard of nanomaterials. NanoImpact 2017, 8, 99–116. [Google Scholar] [CrossRef]
- Chen, F.-M.; Liu, X. Advancing biomaterials of human origin for tissue engineering. Prog. Polym. Sci. 2016, 53, 86–168. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.-D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).