Abstract
A sufficient and mature vegetative growth is an essential condition for production in the following season and consequently affects the alternate bearing (AB) behavior. However, little is known about its interaction with the crop load and water supply. Herein, we studied the effect of different water regimes and bearing status on the vegetative intensity and flush and its consequence on the flowering parameters of the following season. Rainfed (RF) and fully irrigated (FI) treatments were applied for bearing (ON) and non-bearing (OFF) trees of the ‘Zalmati’ olive orchard in south Tunisia during 2018. The water deficit condition (RF) and the high crop load (ON) have caused a similar decrease in the total node number and shoots length by about 65%. Furthermore, the flowering parameters in 2019, especially the percentage of floral buds (i.e., floral induction intensity), were differentially affected by water supply according to the bearing status of the previous season. FI reduced the percentage of floral buds in 2019 by about 37% for OFF trees, while it increased it by more than four times for the ON trees. Concerning the growth flush, the second (i.e., autumnal) flush seems to respond better to water supply than the first (i.e., spring) one, for both vegetative and flowering parameters. The second flush of growth provided 28% of the total vegetative growth but contributed about 35% to the total number of floral buds for RF OFF trees. Besides, FI makes the first and the second flushes contribute equally to total vegetative growth and to the flowering parameters of the following season. On the other hand, the bearing status of the trees does not affect the contribution of each flush to the total vegetative growth.
1. Introduction
Alternate bearing (AB), which defines the irregularity of the production between years in a wide range of fruit tree species, causes high economic losses not only at the farmer’s level but also for national and international economics [1,2]. Great efforts deployed by researchers for decades have allowed the determination of several factors involved in the regulation of uneven yields. In fact, cultivar (genetic) traits, hormonal balance, and competition for assimilates between reproductive and vegetative growth are the major determinant endogenous factors. However, the cyclicity and the intensity of AB are also deeply influenced by several other exogenous factors such as temperature, water supply and nutritional status [1,3,4].
Olive tree is one of the fruit trees in which culture is mostly affected by the AB behavior (5). The biennial cycle that results in this phenomenon is characterized by the formation of new shoots in the non-bearing year (OFF) when the tree has very few or no fruits. In the following year, called bearing (ON), the buds developed in the previous year (essential condition for the production) form inflorescences that result in a high fruit yield, but a limited or insignificant vegetative growth. For olive trees when studied under rainfed conditions, which is most common in the olive-growing regions, the new shoots are formed mainly during two periods: the spring, or the first period, and the autumn, or the second period. During year OFF, the first growth flush is considered as the most dominant, while the second one seems to represent mainly a small elongation of the shoots formed during the first period [5]. The same phenomenon is repeated during the year ON, but at greatly reduced intensity, in favor of the reproductive development. This pattern in olive tree, like in other fruit crops, was attributed to several factors such as the cultivar, nutritional status, chilling accumulation, crop load and water supply at different degrees [6,7,8,9].
Crop load and water supply are considered as the major endogenous and exogenous determinant factors of the AB behavior in olive tree. An increase in fruits number during ON year inhibits floral induction by hormonal signal and new shoots growth through high competition for assimilates, causing an OFF year in the following season. On the other hand, when there is a water deficiency during OFF years, a reduction in vegetative growth was reported, which can negatively impact the production of the following year [10,11,12]. In practice, these two factors do not act separately on the regulation of the AB behavior, but they are rather in continuous interaction. Besides, the competition between fruits and shoots intensifies when there is a water deficit during the year ON, which can negatively impact both parameters [12,13,14]. Several studies suggest a lesser water stress sensitivity and/or water needs during year OFF in comparison to year ON for olive trees [12,14,15,16,17].
Despite the efforts to decipher the impact of the interaction between water supply and the bearing status of olive trees on AB behavior, some gaps remained in this topic. There is a wide agreement among researchers about the positive effect of water supply on the intensity of the vegetative growth, independently of crop load, which is supposed to increase the productive potential of the following year [10,11,18,19,20,21,22,23]. However, this beneficial effect on the yield of the following year has not always been confirmed, and in case of improvement, it is not clear if this comes from the increase in the potential fructification site (the number of buds) and/or from its floral induction (percentage floral buds). Similarly, there are studies that attribute a greater role to crop load in regulating shoot growth and consequently to AB behavior; while others attribute it to water availability [12,13].
In the Southern region of the Mediterranean, one of the main areas of olive growing in the world, the spring period seems to be rainier and has less evapotranspiration rate than the end of the summer and the start of the autumn [24], which coincide with the two main periods of vegetative growth of the olive tree in rainfed orchards. Furthermore, irregular rains are frequently observed in this same region, resulting in a drier spring and/or autumn, which could be aggravated by global warming and the resulting climate changes [25,26,27,28]. These climatic conditions, in addition to the irrigation which is establishing itself as an increasingly frequent practice using different strategies, can further have a serious effect on the intensity and the dynamics of growth and consequently on the productive potential of the following season [17,21,29,30]. In fact, a high availability of water at the end of summer and the beginning of autumn seems to increase the intensity of the second growth flush, while its reduction during spring can reduce the intensity of the first growth [13,14,31]. All these results make it mandatory to investigate the intensity of each wave of growth and its contribution to the productive potential of the following year under different water availabilities and crop load conditions. This can consequently contribute to the development of agronomic strategies that can be used in the adaptation of olive orchards to the climatic changes and in the improvement of the irrigation management.
From a methodological point of view, the vegetative growth was being evaluated in general by following-up the growth of individual shoots rather than the whole shoots number. This, hence, could be a limiting factor in the interpretation of the real growth intensity. Some studies consider measuring the increase in canopy volume parameter, which could be interesting from a physiological point of view (water relation and photosynthesis), but provides very little information about the productive potential of the tree (i.e., buds number). Tree crown is considered as a collection or population of modules represented by the current-year shoots [32]. The analysis of the population of modules (i.e., unit of ramification) can be more representative and can give a better and complete picture about the tree’s growth and reproductive behavior [6,33]. In addition, in most studies, as part of orchard care, pruning was applied during winter after the application of different water regimes and/or bearing status. This can induce errors in the evaluation of the real contribution of the vegetative growth to the productive potential of the following year and can mask the differences among the different treatments.
The aim of this study was, therefore, to evaluate for the first time the effective contribution of the total vegetative growth and of each flush assessed on the level of ramification unit of unpruned trees, under different water regimes and bearing status, to the flowering intensity of the following season.
2. Materials and Methods
2.1. Experimental Site and Treatments
The experiment was conducted in two consecutive years, 2018 (n) and 2019 (n + 1) in a commercial farm in the region of Medenine (33°20′ N 10°29′ E, at an elevation of 81 m a.s.l.) in southern Tunisia. The region is Mediterranean and the climate is of the ‘hot desert climate’ type according to the Köppen climate classification, with an irregular annual rainfall of ~200 mm/season and with almost no precipitation during the summer. The orchard is established with 25-year-old ‘Zalmati’ olive trees in a square disposition of 15 m × 15 m (i.e., 44 trees ha−1). The site is flat, and the soil of the orchard is sandy loam with about 1.3 m of depth. The trees are formed in a vase shape, and are not pruned after starting the assay in order to avoid an artificial regulation of vegetative growth among treatments, allowing the evaluation of productive potential and the shoot fertility in the following year of their development (i.e., 2019).
Trees grown under two water regimes: rainfed (RF) and fully irrigated (FI) received necessary water to reach 100% of crop evapotranspiration (ETc). The crop evapotranspiration is calculated by the Formula: ETc = ET0 × Kc × Kr [34,35,36]. The ETo is the reference evapotranspiration determined using the Penman-Monteith equation [34,37] and Kc crop coefficient estimated as 0.6 for spring and autumn months and 0.5 for the summer, while Kr, the crop cover coefficients, is set to 0.45 as proposed in the literature [34,35,38].
The experiment was designed as a randomized complete block with four replications, where the water regime (RF, FI) and the bearing status (OFF, ON) are the factors (Figure 1A). For the bearing status factor, two levels were considered: trees without inflorescences (i.e., non-bearing or OFF trees) and with high and similar flowering intensity (i.e., high crop load or ON trees). The block consisted of three adjacent rows, each row composed of twelve trees. The central trees were used as experimental trees (four trees per block and a total of sixteen trees for all the blocks) and the rest as guard trees. All experimental trees had a similar vigor. In each block, four treatments were applied: RF-OFF (water deficit acts as limiting factor), RF-ON (both factors crop load and water deficit act as limiting factors), FI-OFF (neither water nor crop load act as limiting factors) and FI-ON (crop load acts as the limiting factor). At the start of the study (spring 2018) about 23% of ‘Zalmati’ olive trees on the orchard were ON trees. Due to the difficulties faced when using an automated drip irrigation system, an individual manual irrigation system was adopted using a gallon water trailer (water quantity supplied takes into account the irrigation technique efficiency) instead to supply water in the evening (to reduce evaporation), for three days each, for the FI treatment from March to October.
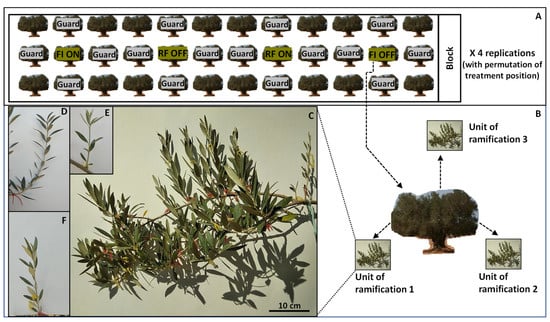
Figure 1.
A representation of the experimental setting. The randomized complete block experimental design (A), the three units of ramification per tree (B), a unit of ramification (C) a spring flush shoot (D), an autumnal flush shoot (E) and a shoot having growth of both flushes (F). Different colored threads used as markers in each unit of ramification (C–F). The sky-blue thread marks the start of the unit of ramification; The red threads mark the start point of spring growth; The yellow threads mark the start point of autumnal growth. RF-ON: non-irrigated trees with high crop load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crop load; FI-OFF: Fully irrigated trees with no fruits.
2.2. Assessed Parameters
The tree water status was assessed monthly between March and June, and then every 10 days until October, by the leaf relative water content (RWC). Fifteen healthy leaves per tree fully expanded were collected each month or ten days in the middle of the canopy and exposed to sunlight. Collected leaves were directly weighted (FW) and then rapidly immersed in distilled water and maintained for 24 h at 5 °C until they reached a constant weight indicating full turgor (TW). Finally, the dry weight was obtained after placing the leaves in an electrical oven at 65 °C during 72 h. Leaf relative water content was calculated as [39]: RWC (%) = 100 × (FW − DW)/(TW − DW).
To assess vegetative and reproductive growth, three units of ramification (group of shoots developed before 2018) with similar volume (≈25 cm × 25 cm × 50 cm), ramification number (≈5 to 6) and leaf density were selected in the middle and regularly distributed around the canopy (Figure 1B). To facilitate the recognition of the growth of each flush, during the first week of July all shoots formed during the first period of the year (considered as spring or first flush) were marked by placing a small dot of paint just before the tip of the shoot. The point of the start of growth of spring flush is recognizable by means of morphological markers (compared to the growth of the previous year). To preserve the painting point, a thread was placed in the same place at the beginning of September until the shoots were sufficiently grown (Figure 1C–F). For the shoots that did not grow at that time, another dot of paint was placed right next to the old one (at the end of October another thread was placed for those that were grown). During 2018, the length and the node number of each shoot and the number of shoots per unit of ramification were recorded. In July 2018 the recording was for spring growth and at the beginning of November for the autumnal flush. The following year (during March 2019 with no irrigation applied) the number of fertile shoots per unit of ramification and the total number of floral buds (developing inflorescences) in the shoots of each flush of 2018 was determined. The number of flowers per inflorescence and flower quality in 2019 were not considered because of their direct dependency on the water availability and the flowering intensity in the same year [40].
2.3. Statistical Analysis
Statistical data analysis was performed by analysis of variance (ANOVA) to evaluate the effect of the water regime and the bearing status of the tree on the vegetative growth of the treatment season and on the flowering parameters of the following season. Tukey’s test was performed to test significance (p ≤ 0.01) of differences among treatments means using Statistix 8.0 (Analytical Software, Tallahassee, FL, USA).
3. Results
3.1. Data Variability
All evaluated parameters were significantly affected by the two studied factors: bearing status and the water regime, but at different degrees (Table 1). The RWC was the most affected parameter by water regime, while it was only influenced by bearing status during a short period (i.e., the end of July and the beginning of August). In addition, two other parameters were more greatly affected by the water regime than by the bearing status: average shoot length and average node number per shoot. In contrast, shoot number per unit of ramification plus all the fertility parameters of the following season were mostly influenced by the bearing status rather than the water regime. Only the total shoot length and the total node number per unit of ramification seem to be similarly affected by the two factors. On other hand, the SS values of the BS∗WR show that the response of all the fertility parameters to one factor was influenced by the other one.

Table 1.
The percentage of sums of squares (SS) attributed to the bearing status (BS), water regime (WR), the interaction among them (BS∗WR) and within treatments (Error) for the morphophysiological parameters.
3.2. Meteorological Data
The yearly reference evapotranspiration is about 1414 mm and varied from 41.9 mm in February to 207.2 mm in July (Table 2). This variation was clearly influenced by the temperature. The daily average temperature during the hottest month of July is about 30 °C. In February, the coolest month, the indicated daily temperature is 13 °C. The water used for irrigation has a pH and electrical conductivity equal to 7.9 and 3.6 ds/m, respectively. The total amount of water supply for FI treatment was about 284 mm/ha. About 55% of this quantity was supplied during the hot summer with very low rainfall.
Table 2.
Rainfall, reference evapotranspiration (ET0) and irrigation amount for the two water regimes during the 2018 season.
3.3. Effect of Bearing Status and Water Treatments on the RWC
The values of RWC, presented in Figure 2, show a clear variation during the season and among treatments. The RWC in the non-irrigated trees (RF) decreased irregularly and progressively starting from May (≈71%) to reach a minimum at the beginning of August (49% to 56%). A slight enhancement in the water status was then observed towards the end of August (≈60%). This enhancement was sustained during the months of September and October. The RWC of the FI trees, however, varies between 83% and 92% and remains obviously higher and more regular through the seasons than those of the RF treatment. The RWC was very similar between the OFF and ON trees, except for a short period, at the end of July and the beginning of August, when the ON trees presented a lower RWC (49%) than the OFF trees (56%) in rainfed conditions.
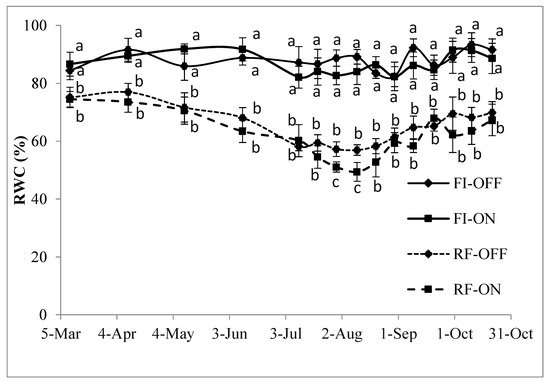
Figure 2.
Leaf relative water content for the bearing status (ON and OFF) and the water regimes (RF and FI) from March to October 2018. RF-ON: non-irrigated trees with high crops load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crops load; FI-OFF: Fully irrigated trees with no fruits. Values (means ± standard error) followed by different letters are significantly different within the same date according to Tukey’s test at p ≤ 0.01.
3.4. Effect of Bearing Status and Water Treatments on the Vegetative Growth Parameters
The vegetative growth during 2018 was clearly affected by water supply and bearing status (Figure 3). The average length and node number of shoots increased significantly by full irrigation for both bearing status, but was more noticeable for the ON (i.e., more than 5-fold) than for the OFF (i.e., 2- to 3-fold) trees. In the presence of fruits (ON trees), shoot growth parameters were reduced for both water regimes (RF and FI), but this reduction was greater under rainfed conditions.
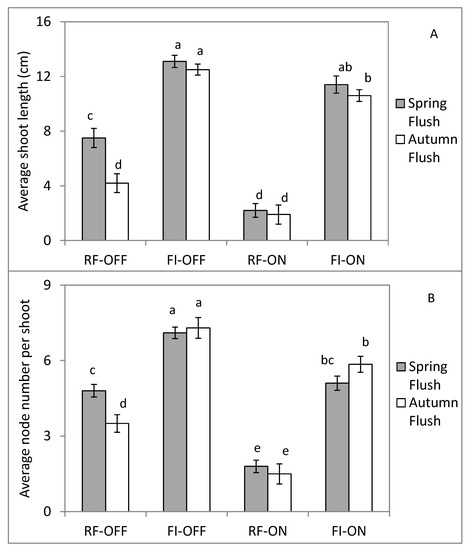
Figure 3.
Average length (A) and node number (B) of shoot for different bearing status (ON and OFF) and the water regimes (RF and FI). RF-ON: non-irrigated trees with high crop load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crop load; FI-OFF: Fully irrigated trees with no fruits. Values (means ± standard error) followed by different letters are significantly different according to Tukey’s test at p ≤ 0.01.
Comparing the flush growth, shoots formed during the spring were longer and had more nodes than those formed during the autumnal growth flush for RF OFF trees (Figure 3). However, the supply of all water needs for the OFF trees resulted in a similar shoot length and node number for both growth flushes. For ON trees, whether under rainfed or fully irrigated conditions, no differences were found between the spring and autumn flushes for these two parameters.
The shoot number per unit of ramification increased by water irrigation (FI treatment) and by the absence of fruits (i.e., OFF trees). A positive effect of water supply on the shoot number during autumn growth was recorded for both bearing status, but only for OFF trees during the spring flush (Figure 4).
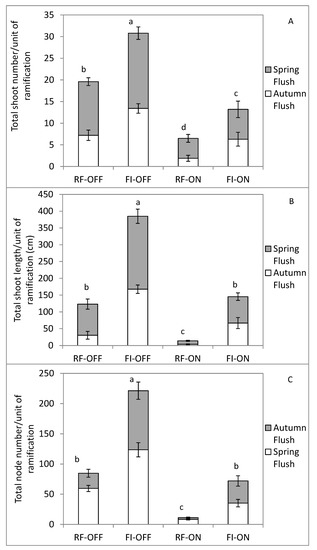
Figure 4.
Total shoot number (A), total shoot length (B) and total node number (C) per ramification unit, for different bearing status (ON and OFF) and the water regimes (RF and FI). RF-ON: non-irrigated trees with high crop load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crop load; FI-OFF: Fully irrigated trees with no fruits. Values (means ± standard error) followed by different letters are significantly different between treatments, according to Tukey’s test at p ≤ 0.01.
The total node number (i.e., the sum of the node number of all shoots formed during 2018 per unit of ramification) and the total shoot length (i.e., the sum of the length of all shoots formed during 2018 per unit of ramification) per unit of ramification increased more than three and six times for FI treatment compared to rainfed regime for OFF and ON trees, respectively (Figure 4). In addition, the fully irrigated ON trees had similar total shoot length and total node number per unit of ramification compared to the non-irrigated OFF trees. The total vegetative growth was significantly greater for FI OFF trees than all the other treatments.
The determination of the contribution of each growth flush to the total node number (i.e., the percentage of the total node number of shoots formed during 2018 per unit of ramification resulting from each flush of growth) and to the total shoot length (i.e., the percentage of the total length of shoots formed during 2018 per unit of ramification resulting from each flush of growth) for the OFF and ON trees showed that the spring flush was dominant under rainfed conditions (RF) by producing about 72% of the total vegetative growth (Figure 5). In contrast, both growth flushes had a similar contribution to the total vegetative growth in the absence (OFF) and presence (ON) of fruits, when they receive all their water needs (i.e., FI treatment).
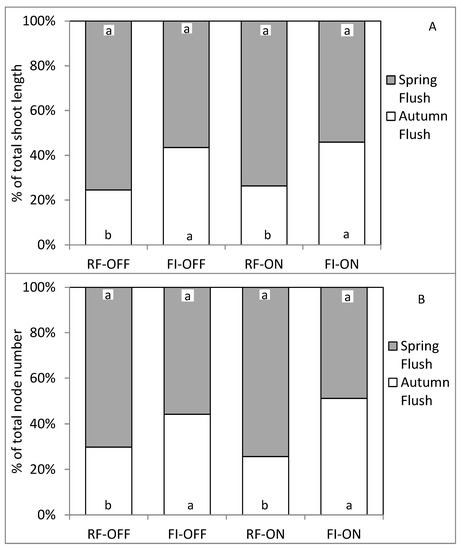
Figure 5.
Contribution in percentage of each growth flush to the total shoot length (A) and node number (B), for different bearing status (ON and OFF) and the water regimes (RF and FI). RF-ON: non-irrigated trees with high crop load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crop load; FI-OFF: Fully irrigated trees with no fruits. Different letters indicate significant difference between growth flush for the same treatment at p ≤ 0.01.
In summary, full irrigation enabled ON trees to achieve total vegetative growth like the RF OFF trees. The contribution of the autumn flush to the total vegetative growth of ON and OFF trees increased from 28% to almost 50% following complete irrigation. The response of vegetative growth at the shoot level to the different treatments was different from that observed at the level of the unit of ramification.
3.5. Effect of Bearing Status and Water Treatments on the Flowering Parameters of the Following Season
The total number of floral buds (i.e., the sum of the floral buds in 2019 of all shoots formed during 2018 per unit of ramification) and fertile shoots per unit of ramification during the spring of 2019 were greatly affected by the tree bearing status in the previous season (Figure 6). The OFF trees produced ten times more total floral buds and fertile shoots per unit of ramification in the following season compared to the ON ones within the same water regime. Additionally, these differences were further accentuated for the total number of floral buds compared to the fertile shoot number and when the trees were grown under rainfed conditions (RF) compared to those that received all their water needs (FI).
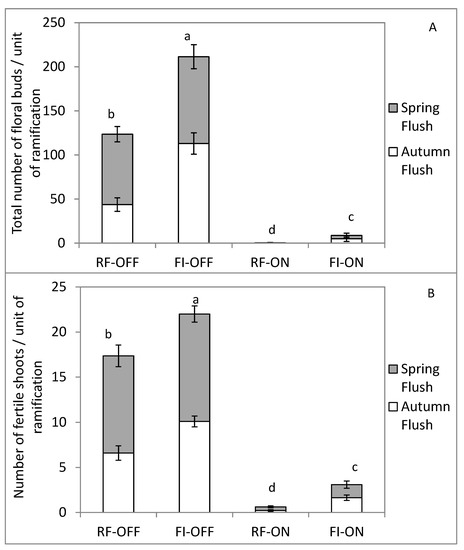
Figure 6.
Total number of floral buds (A) and number of fertile shoots (B) per unit of ramification for different growth flushes, bearing status (ON and OFF) and the water regimes (RF and FI). RF-ON: non-irrigated trees with high crop load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crop load; FI-OFF: Fully irrigated trees with no fruits. Different letters indicate significant difference between treatment according to Tukey’s test at p ≤ 0.01.
Significant differences in flowering parameters were found between the water regimes when the trees had the same reproductive status. FI-OFF trees during 2018 produced 71% more floral buds and 21% more fertile shoots in the following year compared to those grown under rainfed condition (Figure 6). These increases were about 1100% and 400% in the case of ON trees.
As with total growth, for each growth flush the numbers of floral buds and fertile shoots in 2019 was significantly higher in the trees that had OFF status than those that had ON status in 2018, regardless of the water treatment. Furthermore, full irrigation of ON trees caused an increase in these flowering parameters of each growth flush in the following season. The increase was two-fold higher for the autumnal flush compared to the spring flush. However, a full irrigation of OFF trees increased the numbers of floral buds and fertile shoots per unit of ramification during the following season for the autumnal flush but not for the spring flush (Figure 6).
The number of floral buds per node of fertile shoots, the percentage of fertile shoots and the percentage of floral buds (flower induction) per unit of ramification was clearly more affected by the bearing status than the water regime of the previous season (Figure 7). Indeed, regardless of the growth flush and the water regime, the fertility values of the OFF trees were usually higher than those of the ON trees in the following season. The amplitude of these differences depends on the evaluated parameters, which can vary from a few times for the percentage of fertile shoots to ten times for the percentage of floral buds.
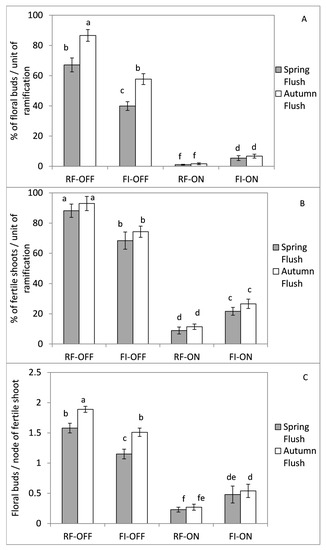
Figure 7.
Percentage of floral buds (A), percentage of fertile shoots (B) and number floral buds per node of fertile shoot (C), for different growth flushes, bearing status (ON and OFF) and the water regimes (RF and FI). RF-ON: non-irrigated trees with high crops load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crops load; FI-OFF: Fully irrigated trees with no fruits. Different letters indicate significant difference between growth flush for the same treatment according to Tukey’s test at p ≤ 0.01.
Water availability had also an effect on the shoot’s fertility and flower induction, but it depended on the bearing status of the tree. For the OFF trees, a full irrigation in 2018 reduced the percentages of floral buds by 37%, fertile shoots by 22% and the number of floral buds per node of fertile shoots 0.8 points during the following year compared to those of the rainfed regime (Figure 7). These reductions have occurred for the spring and the autumnal growth flushes by similar absolute values, but they were higher for the spring one in a relative value. In contrast to the OFF trees, the percentage of fertile shoots and the number of floral buds per node of fertile shoots increased two-fold for the two growth flushes by a full irrigation of ON trees during the previous season, compared to those that were not irrigated (Figure 7). This increase has grown four-fold for the percentage of floral buds parameter.
By comparing the flowering parameters of the spring and autumn flushes for the different treatments, we recorded two responses according to the bearing status of the previous year. The first one was observed for the OFF trees, when the percentage of floral buds and the number of floral buds per node of fertile shoots were approximately 20% and 30% greater in the autumn growth flush compared to the spring one for fully irrigated and rainfed treatments, respectively. However, no difference was recorded between the two flushes of growth for the percentage of fertile shoots. The second response was recorded in the ON trees, which showed similar percentages of floral buds, fertile shoots, and number of floral buds per node of shoot fertility, between the spring and the autumn flushes of growth within the same water regime (Figure 7).
The contribution of each growth flush of 2018 to the total numbers of fertile shoots and floral buds of the following year (i.e., 2019) seems to be affected by the water regime and not by the bearing status factor (Figure 8). Under rainfed condition and for the two bearing status, the spring and the autumn flushes provided approximatively 65% and 35%, respectively, of the total number of fertile shoots and floral buds per unit of ramification. In contrast, in the irrigated trees, the fertile shoots and floral buds were provided in similar proportion from both growth flushes, independently of their bearing status.
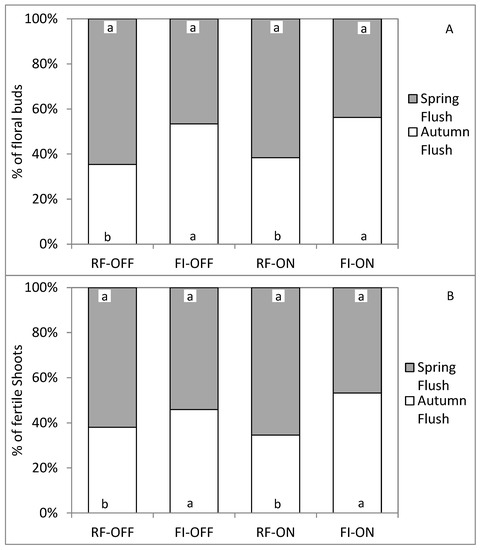
Figure 8.
Contribution in percentage of each growth flush to the total number of floral buds (A) and the total number of fertile shoots (B), for different bearing status (ON and OFF) and water regimes (RF and FI). RF-ON: non-irrigated trees with high crop load; RF-OFF: non-irrigated trees with no fruits; FI-ON: fully irrigated trees with high crop load; FI-OFF: Fully irrigated trees with no fruits. Different letters indicate significant difference between growth flush for the same treatment at p ≤ 0.01.
In brief, the bearing status of the tree seems to have a greater impact on the flowering parameters of the following season than the water regime. On the other hand, water supply reduced shoot’s fertility and flower induction in OFF trees and increased them in ON trees. Furthermore, water supply had a greater positive effect on the flowering intensity of the autumn flush than the one that resulted from the spring flush. The contribution of the autumn flush to the total number of floral buds and the total number of fertile shoots of ON and OFF trees increased from 35% to about 50%, following complete irrigation.
4. Discussion
4.1. Effect of Bearing Status and Water Supply on the Tree Water Status
The RWC was usually used as an indicator of plants water status [41,42] including olive trees [43]. Under rainfed conditions, where the precipitation does not exceed 216 mm (equivalent to about 15% of ET0) and which is mainly centered in the period of low water demand for olive trees (i.e., November to March), OFF and ON trees showed an RWC far less than the fully irrigated ones. The full irrigation treatments showed an RWC that is relatively constant and above 83%, which is similar to other values reported by other studies in olive trees under no water restriction conditions [14]. In contrast, with no irrigation, the RWC decreased gradually from the spring to summer, directly influenced by the reduction in rain and the increase in temperature. The decrease in RWC, which indicates leaf dehydration, coupled with the reduction in water leaf potential is frequently observed in olive trees grown under severe water deficit conditions in arid and semi-arid regions [14,44]. After a very dry summer, we have found that the RWC reached between 49% and 56% in RF trees at the beginning of August, and these values were akin to what was reported in other studies for rainfed olive trees (about 50%) grown in semi-arid regions in Tunisia [45,46]. Rainfall during mid-August resulted in a slight improvement of the water status of the trees. This improvement was sustained in the following months of autumn, where certain precipitations and temperature reduction were recorded. This rapid recuperation of water status with the increase in water availability after a dry period was considered as one of the most efficient features of the olive tree to adapt to water deficit in semi-arid regions [23,47,48]. The improvement in the hydric status, starting from the end of summer, was concomitant with the beginning of the second wave of growth for the RF trees, which started a little earlier in August for the FI trees. This response of fully irrigated olive tree is known and is reported in previous literature [13]. In addition, the availability of water was shown to affect other growth process timings, such as the lignification of the endocarp [49] and olive ripening [50]. Therefore, this proves the elasticity of the penological phases and growth dynamics of the olive trees under irregular precipitations in the arid and semi-arid regions.
The ON and OFF trees showed very similar hydric status when they received the total of their water needs. A similar response was recorded for rainfed trees until the end of July and the beginning of August when ON trees had lower RWC. However, this difference was quickly reduced for the values that were taken after the only rains that had fallen in August. These results could be explained by the presence of an effect of crop load on the olive water relations, a finding that is further corroborated by previous studies [12,16,22,51]. The higher water stress recorded for the ON trees was explained by a major consumption of water caused by the developing fruits [16] which coincides with a lower water availability in the presence of a high evapotranspiration demand for rainfed regime and under semi-arid and arid conditions. The absence of effect of crop load on the fully irrigated trees may be due to a satisfaction of all additional water needs induced by the fruits’ development.
4.2. Effect of Bearing Status and Water Supply on the Vegetative Growth Dynamic and Intensity
Most studies in the literature have evaluated the effect of water supply and/or crop load on the vegetative growth of olive trees based essentially on two parameters: the length and number of nodes of shoot. In fact, these studies have reported that water deficit and crop load had a clear negative effect on both parameters studied which was also corroborated by our results [10,12,14,31]. Despite the wide agreement between most of the studies about the negative effect of these two factors on shoot growth, discrepancy was found concerning the importance of each parameter in determining the intensity of vegetative growth, where sometimes a greater effect was attributed to crop load [13] or to water status [12]. These controversies can be probably attributed to differences in studies conditions (cultivars, age of the tree, crops load and water deficit) or to differences in estimation of the intensity of vegetative growth. This latter could be fully measured using only the cumulative growth of all new shoots which in turn depends on the number of the same shoots [6,52]. In our case for example, if we rely mainly on the length and node number of shoot, more effect should be attributed to water supply, however, this estimation changes completely if we consider the cumulative growth parameters. There are no differences in the values of total (cumulative) shoot length and node number per unit of ramification between RF-OFF (water deficit is the limiting factor) and FI-ON (crop load is the limiting factor) treatments. In addition, if we consider the FI-OFF treatments that offer the maximum potential vegetative growth, we can suggest that the two treatments RF-OFF and FI-ON generate a very similar reduction which reaches nearly 65% of the growth. The present results, hence, suggest a similar impact of both factors (water availability and crop load) on the vegetative growth intensity for the studied cultivar under the arid climatic conditions. This novel finding suggests, from a practical point of view, that the inhibitory effect exerted by crop load on vegetative growth can be canceled by means of full irrigation of olive trees, at least in the case of ‘Zalmati’ cultivar.
The olive tree is characterized by two waves of vegetative growth: the first one in the spring and the second one in autumn. The first wave is considered the most intense and dominant, which begins in spring and continues until the first weeks of summer. The second wave which occurs during the months of August, September and October is considered of low intensity and can even be reduced or avoided in unfavorable conditions of water stress or heavy fruits load [5]. This cycle seems to be dependent on the tree water supply. The period between the two waves of growth seems to be characterized by a slowdown of vegetative growth rather than a total halt [17]. A high availability of water at the end of summer and beginning of autumn seems to increase the intensity of the second growth flush, while its reduction during the spring can reduce the intensity of the first growth [13,14,31]. Despite the plethora of information concerning a possible strong interaction between water supply and the dynamics of vegetative growth, very little is known about the contribution of growth flush to total vegetative growth and how it can be influenced by different water regimes. The results that we have found show that, under rainfed conditions, the spring and autumn flushes contributed to the total vegetative growth by about 72% and 28%, for both OFF and ON trees. They further confirmed that the first flush was the dominant one, but also revealed an important contribution of the second flush in the determination of the total vegetative growth, which is generally neglected under rainfed conditions. By receiving all their water needs, OFF and ON trees showed a similar contribution of the first and the second flushes to the total vegetative growth (total node number and shoot length). One might speculate that this unknown response of the olive trees could be especially specific to the cultivar ‘Zalmati’. Nevertheless, the significant enhancement of shoots growth during the second period of vegetative growth previously reported in several cultivars grown under full irrigation conditions suggests that this response could be observed in other olive trees cultivars [13,14,31,53,54]. From an irrigation management point of view, our results suggest that it could be possible to partially recover the vegetative growth that could be lost during dry springs in rainfed olive groves using late complementary irrigation (that will be tested in further studies).
Heavy crop load is known by its reduction in the vegetative growth as we reported in the present study and which was reported by others [6,54]. However, this does not seem to disturb the contribution of each flush to the total vegetative growth. One explanation of this response could be the continuous presence of active reproductive development throughout the vegetative growth period (from spring to autumn), which emphasizes the influence of the inhibitory effect (i.e., competition for assimilates) during both growth flushes. The evaluation of relative growth rates of shoots under different crop loads reported by [54] showed that shoot growth was limited during most of the season under medium and high crop loads.
The increase in all vegetative growth parameters per unit of ramification that resulted from water supply was greater for the autumn compared to spring flush, for both OFF and ON trees. The better response of vegetative growth to water supply in the second period could be due to major water restrictions during this period compared to the first one. In addition, the increase in the number of growth sites (i.e., number of nodes) formed during the first flush could have probably contributed to this response.
4.3. Effect of Bearing Status and Water Supply in the Productive Potential of the Following Season
The reproductive development in 2019 was significantly affected by the two studied factors: bearing cycle and the water supply status of the previous year, as was also reported in several studies [10,11,12,18,19,20,21,22,23]. The total number of floral buds in 2019 decreased by about 96% due to the high crop load (ON) and by 40% due to water deficit (RF) during the previous season. These results agree with previous studies in which the crop load was suggested as the main cause of alternate bearing in olive tree [13,55,56,57]. However, the total number of floral buds decreased by about 95% for ON trees due to water deficit (RF) in comparison with those that received all their water needs (FI ON trees) during the previous season. Therefore, we can suggest that the impact of water supply on the regulation of the total number of floral buds of the following season depends on the bearing status of the tree.
In olive tree, the presence of growing fruits and seeds was proposed as the most determinant factor of bud induction [6,58,59]. Some studies have suggested a positive effect of water deficit on flower bud induction, but without evaluating its interaction with crops load despite its implication in the induction phenomenon [18,23,60]. The percentage of floral buds was differentially affected by water supply according to the bearing status of the previous season. Full irrigation in 2018 reduced the percentage of floral buds by about 37% for OFF trees, while it increased more than four-fold for the ON trees during the following season. The mechanism that may be behind this response is not clear and needs further investigations, but several hypotheses could be proposed. The higher floral bud induction under rainfed condition could be due to a hormonal effect induced by water stress. In fact, a stimulator effect of abscisic acid on floral induction was reported previously in the olive tree [61], whose biosynthesis seems to increase under water deficit condition [46,62,63]. Another possible explanation could be an increase in the competition between the growing number of buds formed by a better water supply (FI). Indeed, a decrease in assimilates availability was suggested previously to affect negatively floral induction [64,65,66,67]. Similarly, a high competition for assimilates could be induced during ON years by the reproductive development. This negative effect of the competition could be alleviated by increasing water supply, which allows a better floral induction. However, the impact of this improvement on the total number of floral buds, and consequently the production rate, remains limited, despite the enhancement in vegetative growth achieved by FI ON trees. This indicates the great control exerted during ON years by the developing fruits on floral induction, while for OFF years a key role is attributed to water supply. Therefore, these results emphasize the complexity of the control exerted by water availability on flower induction and the need to study it more deeply in other cultivars under different conditions. Nevertheless, some possible consequences on irrigation water management should be investigated. For instance, it could be possible to reduce water supply during the OFF years in irrigated olive groves of cultivars with a high alternate bearing tendency without affecting the productive potential of the following season. Indeed, a recent study suggests that 48% of water supply could be saved by applying a regulated deficit irrigation during OFF years, without affecting the olive yields [17]. However, the mechanism behind this result is not fully understood and the authors recommended investigating more the effects of water deficit on the plant parameters during the following ON year.
To the best of our knowledge, there is no information available about the influence of water and bearing status on the fertility of the two vegetative growth flushes in olive tree. Results show that the total number of floral buds and fertile shoots in each flush increased with the water supply (FI treatment) and decreased with the high crop load (ON trees) of the previous season. However, the autumn flush seems to respond better to water supply than the spring flush. These results could be due to a greater sensitivity of the second period of vegetative growth to water deficit.
Concerning buds fertility, the percentage of floral buds was greater for the second flush growth for OFF trees compared to the spring ones, but not for the ON trees. The period of floral induction is concomitant with the second period of vegetative growth [5,58,67,68,69]. Therefore, buds formed during this period may have better floral induction level. All these suggestions need further studies at different levels (morpho-physiological, biochemical, and molecular) to better understand this behavior.
Despite providing 28% of the vegetative growth, the contribution of the second flush increases to about 35% in total number of floral buds per unit of ramification, for non-irrigated OFF trees. This could be due to their greater fertility compared to the buds developed during the spring flush. Besides, when the olive trees receive all their water needs, the autumn flush has an equal contribution to the spring flush. These results not only challenge what was previously thought about the weak involvement of the second wave of growth in the productive cycle of the olive tree but may also suggest new ideas on the water irrigation management. For instance, it would be interesting to study the possibility of recovery from a negative effect of water deficit during the first growth period in the OFF year (in case of dry spring or low water supply) by improving water supply in the second period.
5. Conclusions
The assessment of the vegetative growth dynamics and intensity at a branching unit on unpruned trees was an effective strategy to explore the influence of the bearing status and water supply on the vegetative and reproductive growth regulation in olive trees, by providing new insights into the theme. The obtained results suggest a similar impact of these two factors on total vegetative growth intensity. RF and ON status decrease the total node number and shoot length by about 65%. The bearing status is the most determinant factor on the flowering parameters of the following season. The water supply affected these parameters differently depending on the bearing status of the tree. For the growth flushes, the second flush had a significant contribution to the total vegetative and flowering intensities of the following season (≈one-third), which can become equal to that of the first flush when the trees are fully irrigated.
Despite the importance of the findings of the present study, we think that additional studies that could evaluate the response of other cultivars that grow under different conditions are mandatory to confirm their broad aspect.
Author Contributions
Conceptualization, S.B.M.H. and B.S.-H.; methodology, S.B.M.H., M.B.L. and N.B.; validation, S.B.M.H., M.B.L. and B.S.-H.; formal analysis, N.B. and M.B.L.; investigation, S.B.M.H.; resources, S.B.M.H., M.B.L., N.B. and B.S.-H.; data curation, S.B.M.H. and M.B.L.; writing—original draft preparation, S.B.M.H. and B.S.-H.; writing—review and editing, S.B.M.H., N.B. and M.B.L.; visualization, S.B.M.H., M.B.L., N.B. and B.S.-H.; supervision, S.B.M.H.; project administration, S.B.M.H. and B.S.-H.; funding acquisition, S.B.M.H., N.B. and B.S.-H. All authors have read and agreed to the published version of the manuscript.
Funding
All authors are funded through the Small Research group project from the Deanship of Scientific Research at King Khalid University under research grant number (RGP.1/295/43) and also by INAT-S.A.C agreement.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University for funding this work through the Small Research group project under grant number (R.G.P.1/295/43) and the INAT-S.A.C agreement.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, H.M.; Samach, A. Constraints to obtaining consistent annual yields in perennial tree crops. I: Heavy fruit load dominates over vegetative growth. Plant Sci. 2013, 207, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Krasniqi, A.L.; Blanke, M.M.; Kunz, A.; Damerow, L.; Lakso, A.N.; Meland, M. Alternate bearing in fruit tree crops: Past, present and future. In International Symposium on Physiological Principles and Their Application to Fruit Production; ISHS: Leuven, Belgium, 2014; Volume 1177, pp. 241–248. [Google Scholar]
- Sharma, N.; Singh, S.K.; Mahato, A.K.; Ravishankar, H.; Dubey, A.K.; Singh, N.K. Physiological and molecular basis of alternate bearing in perennial fruit crops. Sci. Hortic. 2018, 243, 214–225. [Google Scholar] [CrossRef]
- Goldschmidt, E.E.; Sadka, A. Yield Alternation: Horticulture, Physiology, Molecular Biology, and Evolution. Hortic. Rev. 2021, 48, 363–418. [Google Scholar]
- Rallo, L.; Cuevas, J. Fruiting and Production. In Olive Growing; Barranco, D., Fernández-Escobar, R., Rallo, L., Eds.; RIRDC: Wagga Wagga, Australia, 2010; pp. 113–145. [Google Scholar]
- Castillo-Llanque, F.; Rapoport, H.F. Relationship between reproductive behavior and new shoot development in 5-year-old branches of olive trees (Olea europaea L.). Trees 2011, 25, 823–832. [Google Scholar] [CrossRef]
- Turktas, M.; Inal, B.; Okay, S.; Erkilic, E.G.; Dundar, E.; Hernandez, P.; Dorado, G.; Unver, T. Nutrition Metabolism Plays an Important Role in the Alternate Bearing of the Olive Tree (Olea europaea L.). PLoS ONE 2013, 8, e59876. [Google Scholar] [CrossRef]
- Arji, I. Determining of Growth and Yield Performance in Some Olive Cultivars in Warm Conditions. In Biological Forum; Research Trend: Dewas, India, 2015; Volume 7, p. 1865. [Google Scholar]
- Ramos, A.; Rapoport, H.; Cabello, D.; Rallo, L. Chilling accumulation, dormancy release temperature, and the role of leaves in olive reproductive budburst: Evaluation using shoot explants. Sci. Hortic. 2018, 231, 241–252. [Google Scholar] [CrossRef]
- Wahbi, S.; Wakrim, R.; Aganchich, B.; Tahi, H.; Serraj, R. Effects of partial rootzone drying (PRD) on adult olive tree (Olea europaea) in field conditions under arid climate: I. Physiological and agronomic responses. Agric. Ecosyst. Environ. 2005, 106, 289–301. [Google Scholar] [CrossRef]
- Iniesta, F.; Testi, L.; Orgaz, F.; Villalobos, F. The effects of regulated and continuous deficit irrigation on the water use, growth and yield of olive trees. Eur. J. Agron. 2009, 30, 258–265. [Google Scholar] [CrossRef]
- Martín-Vertedor, A.I.; Rodríguez, J.M.P.; Losada, H.P.; Castiel, E.F. Interactive responses to water deficits and crop load in olive (Olea europaea L., cv. Morisca) I.—Growth and water relations. Agric. Water Manag. 2011, 98, 941–949. [Google Scholar] [CrossRef][Green Version]
- Mezghani, M.A.; Charfi, C.M.; Gouiaa, M.; Labidi, F. Vegetative and reproductive behaviour of some olive tree varieties (Olea europaea L.) under deficit irrigation regimes in semi-arid conditions of Central Tunisia. Sci. Hortic. 2012, 146, 143–152. [Google Scholar] [CrossRef]
- Abboud, S.; Dbara, S.; Abidi, W.; Braham, M. Differential agro-physiological responses induced by partial root-zone drying irrigation in olive cultivars grown in semi-arid conditions. Environ. Exp. Bot. 2019, 167, 103663. [Google Scholar] [CrossRef]
- Naor, A.; Schneider, D.; Ben-Gal, A.; Zipori, I.; Dag, A.; Kerem, Z.; Birger, R.; Peres, M.; Gal, Y. The effects of crop load and irrigation rate in the oil accumulation stage on oil yield and water relations of ‘Koroneiki’ olives. Irrig. Sci. 2012, 31, 781–791. [Google Scholar] [CrossRef]
- Bustan, A.; Dag, A.; Yermiyahu, U.; Erel, R.; Presnov, E.; Agam, N.; Kool, D.; Iwema, J.; Zipori, I.; Ben-Gal, A. Fruit load governs transpiration of olive trees. Tree Physiol. 2016, 36, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Siakou, M.; Bruggeman, A.; Eliades, M.; Djuma, H.; Kyriacou, M.C.; Moriana, A. Phenology, Morphology and Physiology Responses of Deficit Irrigated ‘Koroneiki’ Olive Trees as Affected by Environmental Conditions and Alternate Bearing. Agronomy 2022, 12, 879. [Google Scholar] [CrossRef]
- Grattan, S.; Berenguer, M.; Connell, J.; Polito, V.; Vossen, P. Olive oil production as influenced by different quantities of applied water. Agric. Water Manag. 2006, 85, 133–140. [Google Scholar] [CrossRef]
- Gucci, R.; Lodolini, E.; Rapoport, H. Productivity of olive trees with different water status and crop load. J. Hortic. Sci. Biotechnol. 2007, 82, 648–656. [Google Scholar] [CrossRef]
- Greven, M.; Neal, S.; Green, S.; Dichio, B.; Clothier, B. The effects of drought on the water use, fruit development and oil yield from young olive trees. Agric. Water Manag. 2009, 96, 1525–1531. [Google Scholar] [CrossRef]
- Palese, A.M.; Nuzzo, V.; Favati, F.; Pietrafesa, A.; Celano, G.; Xiloyannis, C. Effects of water deficit on the vegetative response, yield and oil quality of olive trees (Olea europaea L., cv Coratina) grown under intensive cultivation. Sci. Hortic. 2010, 125, 222–229. [Google Scholar] [CrossRef]
- Moriana, A.; Perez-Lopez, D.; Prieto, M.H.; Ramírez-Santa-Pau, M.; Pérez Rodríguez, J.M. Midday stem water potential as a useful tool for estimating irrigation requirements in olive trees. Agric. Water Manag. 2012, 112, 43–54. [Google Scholar] [CrossRef]
- Trentacoste, E.R.; Calderón, F.J.; Contreras-Zanessi, O.; Galarza, W.; Banco, A.P.; Puertas, C.M. Effect of regulated deficit irrigation during the vegetative growth period on shoot elongation and oil yield components in olive hedgerows (cv. Arbosana) pruned annually on alternate sides in San Juan, Argentina. Irrig. Sci. 2019, 37, 533–546. [Google Scholar] [CrossRef]
- Masmoudi-Charfi, C.; Habaieb, H. Rainfall Distribution Functions for Irrigation Scheduling: Calculation Procedures Following Site of Olive (Olea europaea L.) Cultivation and Growing Periods. Am. J. Plant Sci. 2014, 5, 2094–2133. [Google Scholar] [CrossRef][Green Version]
- Field, C.B.; Barros, V.R. (Eds.) Climate Change 2014-Impacts, Adaptation and Vulnerability: Regional Aspects; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Sonwa, D.J.; Dieye, A.; El Mzouri, E.-H.; Majule, A.; Mugabe, F.T.; Omolo, N.; Wouapi, H.; Obando, J.; Brooks, N. Drivers of climate risk in African agriculture. Clim. Dev. 2017, 9, 383–398. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mishra, A.; Trenberth, K.E. Climate Change and Drought: A Perspective on Drought Indices. Curr. Clim. Chang. Rep. 2018, 4, 145–163. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skeg, J.; Buendia, E.C.; Masson-Delmotte, V.; Pörtner, H.O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; Van Diemen, S.; et al. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Philpapers: London, UK, 2019. [Google Scholar]
- Arampatzis, G.; Hatzigiannakis, E.; Pisinaras, V.; Kourgialas, N.; Psarras, G.; Kinigopoulou, V.; Panagopoulos, A.; Koubouris, G. Soil water content and olive tree yield responses to soil management, irrigation, and precipitation in a hilly Mediterranean area. J. Water Clim. Chang. 2018, 9, 672–678. [Google Scholar] [CrossRef]
- Fraga, H.; Pinto, J.G.; Santos, J.A. Olive tree irrigation as a climate change adaptation measure in Alentejo, Portugal. Agric. Water Manag. 2020, 237, 106193. [Google Scholar] [CrossRef]
- Pérez, D.; Ribas, F.; Olmedilla, J.N. Influence of irrigation on a traditional rainfed olive orchard (cv Cornicabra). Options Méditerranéennes 2004, 60, 85–89. [Google Scholar]
- Hasegawa, S.; Takeda, H. Functional specialization of current shoots as a reproductive strategy in Japanese alder (Alnus hirsuta var. sibirica). Can. J. Bot. 2001, 79, 38–48. [Google Scholar]
- Suzuki, A. Patterns of vegetative growth and reproduction in relation to branch orders: The plant as a spatially structured population. Trees 2000, 14, 329–333. [Google Scholar] [CrossRef]
- Allen, R.G. Crop Evapotranspiration-Guideline for computing crop water requirements. Irrig. Drain 1998, 56, 300. [Google Scholar]
- Fereres, E.; Goldhamer, D.A.; Sadras, V.O. Yield response to water of fruit trees and vines: Guidelines. FAO Irrig. Drain. Pap. 2012, 66, 246–497. [Google Scholar]
- Romero-Trigueros, C.; Vivaldi, G.A.; Nicolás, E.N.; Paduano, A.; Salcedo, F.P.; Camposeo, S. Ripening indices, olive yield and oil quality in response to irrigation with saline reclaimed water and deficit strategies. Front. Plant Sci. 2019, 10, 1243. [Google Scholar] [CrossRef] [PubMed]
- Jabloun, M.D.; Sahli, A. Evaluation of FAO-56 methodology for estimating reference evapotranspiration using limited climatic data: Application to Tunisia. Agric. Water Manag. 2008, 95, 707–715. [Google Scholar] [CrossRef]
- Trentacoste, E.R.; Puertas, C.M.; Sadras, V.O. Effect of irrigation and tree density on vegetative growth, oil yield and water use efficiency in young olive orchard under arid conditions in Mendoza, Argentina. Irrig. Sci. 2015, 33, 429–440. [Google Scholar] [CrossRef]
- Pietragalla, J.; Mullan, D. Leaf Relative Water Content. In Physiological Breeding II: A Field Guide to Wheat Genotyping; CIMMYT: Mexico City, Mexico, 2012; pp. 25–27. [Google Scholar]
- Rapoport, H.; Hammami, S.B.; Martins, P.; Pérez-Priego, O.; Orgaz, F. Influence of water deficits at different times during olive tree inflorescence and flower development. Environ. Exp. Bot. 2012, 77, 227–233. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.; Khan, I.; Xue, L. Methyl Jasmonate-Induced Alteration in Lipid Peroxidation, Antioxidative Defence System and Yield in Soybean Under Drought. J. Agron. Crop Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Wahid, A.; Ahmad, N.; Saleem, B.A. Improving the drought tolerance in rice (Oryza sativa L.) by exogenous application of salicylic acid. J. Agron. Crop Sci. 2009, 195, 237–246. [Google Scholar] [CrossRef]
- Torres, I.; Sanchez, M.T.; Benlloch-Gonzalez, M.; Perez-Marin, D. Irrigation decision support based on leaf relative water content determination in olive grove using near infrared spectroscopy. Biosyst. Eng. 2019, 180, 50–58. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, B.; Boujnah, D.; Zarrouk, M. Seasonal changes in water relations and gas exchange in leaves of two Tunisian olive (Olea europaea L.) cultivars under water deficit. J. Hortic. Sci. Biotechnol. 2007, 82, 721–726. [Google Scholar] [CrossRef]
- Rouina, B.B.; Trigui, A.; d’Andria, R.; Boukhris, M.; Chaieb, M. Effects of water stress and soil type on photosynthesis, leaf water potential and yield of olive trees (Olea europaea L. cv. Chemlali Sfax). Aust. J. Exp. Agric. 2007, 47, 1484–1490. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaïbi, W.; Zarrouk, M. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. 2009, 119, 257–263. [Google Scholar] [CrossRef]
- Tognetti, R.; d’Andria, R.; Morelli, G.; Calandrelli, D.; Fragnito, F. Irrigation effects on daily and seasonal variations of trunk sap flow and leaf water relations in olive trees. Plant Soil 2004, 263, 249–264. [Google Scholar] [CrossRef]
- Boughalleb, F.; Hajlaoui, H. Physiological and anatomical changes induced by drought in two olive cultivars (cv Zalmati and Chemlali). Acta Physiol. Plant. 2011, 33, 53–65. [Google Scholar] [CrossRef]
- Hammami, S.B.M.; Costagli, G.; Rapoport, H.F. Cell and tissue dynamics of olive endocarp sclerification vary according to water availability. Physiol. Plant. 2013, 149, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, F.; Basile, B.; Morelli, G.; D’Andria, R.; Tonutti, P. Effects of irrigation on fruit ripening behavior and metabolic changes in olive. Sci. Hortic. 2012, 144, 201–207. [Google Scholar] [CrossRef]
- Ben-Gal, A.; Yermiyahu, U.; Zipori, I.; Presnov, E.; Hanoch, E.; Dag, A. The influence of bearing cycles on olive oil production response to irrigation. Irrig. Sci. 2010, 29, 253–263. [Google Scholar] [CrossRef]
- Hammami, S.B.; de la Rosa, R.; Sghaier-Hammami, B.; León, L.; Rapoport, H.F. Reliable and relevant qualitative descriptors for evaluating complex architectural traits in olive progenies. Sci. Hortic. 2012, 143, 157–166. [Google Scholar] [CrossRef]
- Strippoli, G.; Vivaldi, G.A.; Camposeo, S.; Contò, F. Sprouts seasonal elongation of two olive cultivars in a high-density orchard. Agric. Sci. 2013, 04, 376–381. [Google Scholar] [CrossRef]
- Fernández, F.J.; Ladux, J.L.; Searles, P. Dynamics of shoot and fruit growth following fruit thinning in olive trees: Same season and subsequent season responses. Sci. Hortic. 2015, 192, 320–330. [Google Scholar] [CrossRef]
- Ramos, A.M.D.S. Inducción Floral y Latencia de Las Yemas Del Olivo (Olea europaea L.); ETSIAM: Córdoba, Spain, 2000. [Google Scholar]
- Lavee, S. Alternate bearing in olive initiated by abiotic induction leading to biotic responses. Adv. Hortic. Sci. 2015, 29, 213–220. [Google Scholar]
- Dag, A.; Bustan, A.; Avni, A.; Tzipori, I.; Lavee, S.; Riov, J. Timing of fruit removal affects concurrent vegetative growth and subsequent return bloom and yield in olive (Olea europaea L.). Sci. Hortic. 2010, 123, 469–472. [Google Scholar] [CrossRef]
- Fernandez-Escobar, R.; Benlloch, M.; Navarro, C.; Martin, G.C. The time of floral induction in the olive. J. Am. Soc. Hortic. Sci. 1992, 117, 304–307. [Google Scholar] [CrossRef]
- Stutte, G.W.; Martin, G.C. Effect of killing the seed on return bloom of olive. Sci. Hortic. 1986, 29, 107–113. [Google Scholar] [CrossRef]
- Gómez-del-Campo, M. Summer deficit-irrigation strategies in a hedgerow olive orchard cv.’Arbequina’: Effect on fruit characteristics and yield. Irrig. Sci. 2013, 31, 259–269. [Google Scholar] [CrossRef]
- Ulger, S.; Sonmez, S.; Karkacier, M.; Ertoy, N.; Akdesir, O.; Aksu, M. Determination of endogenous hormones, sugars and mineral nutrition levels during the induction, initiation and differentiation stage and their effects on flower formation in olive. Plant Growth Regul. 2004, 42, 89–95. [Google Scholar] [CrossRef]
- Kitsaki, C.K.; Drossopoulos, J.B. Environmental effect on ABA concentration and water potential in olive leaves (Olea europaea L. cv “Koroneiki”) under non-irrigated field conditions. Environ. Exp. Bot. 2005, 54, 77–89. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Diaz-Espejo, A.; Perez-Martin, A.; Hernandez-Santana, V. Role of hydraulic and chemical signals in leaves, stems and roots in the stomatal behaviour of olive trees under water stress and recovery conditions. Tree Physiol. 2015, 35, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Proietti, P. Changes in photosynthesis and fruit characteristics in olive in response to assimilate availability. Photosynthetica 2003, 41, 559–564. [Google Scholar] [CrossRef]
- Tombesi, A. Biologia fiorale e di fruttificazione. Olea. Trattato Di Olivic. 2003, 35–55. [Google Scholar]
- Cherbiy-Hoffmann, S.U.; Hall, A.J.; Searles, P.S.; Rousseaux, M.C. Responses of olive tree yield determinants and components to shading during potentially critical phenological phases. Sci. Hortic. 2015, 184, 70–77. [Google Scholar] [CrossRef]
- Hueso, A.; Trentacoste, E.R.; Ruiz, C.; Meng, L.; Pérez-Pastor, A.; De la Rosa, J.M.; Gómez-del-Campo, M. Effect of deficit irrigation during the oil synthesis period on carbohydrate content in olive ‘Arbequina’ hedgerows. Acta Hortic. 2018, 1199, 75–80. [Google Scholar] [CrossRef]
- Fabbri, A.; Benelli, C. Review Article Flower bud induction and differentiation in olive. J. Hortic. Sci. Biotechnol. 2000, 75, 131–141. [Google Scholar] [CrossRef]
- Andreini, L.; Bartolini, S.; Guivarch, A.; Chriqui, D.; Vitagliano, C. Histological and immunohistochemical studies on flower induction in the olive tree (Olea europaea L.). Plant Biol. 2008, 10, 588–595. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).