
Characteristics of Soil Macrofauna and Its Coupling Relationship with Environmental Factors in the Loess Area of Northern Shaanxi

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Collection and Identification of Soil Fauna
2.4. Determination of Litter Mass and Soil Physical and Chemical Properties
2.5. Statistical Analysis
3. Results
3.1. Litter and Soil Variables
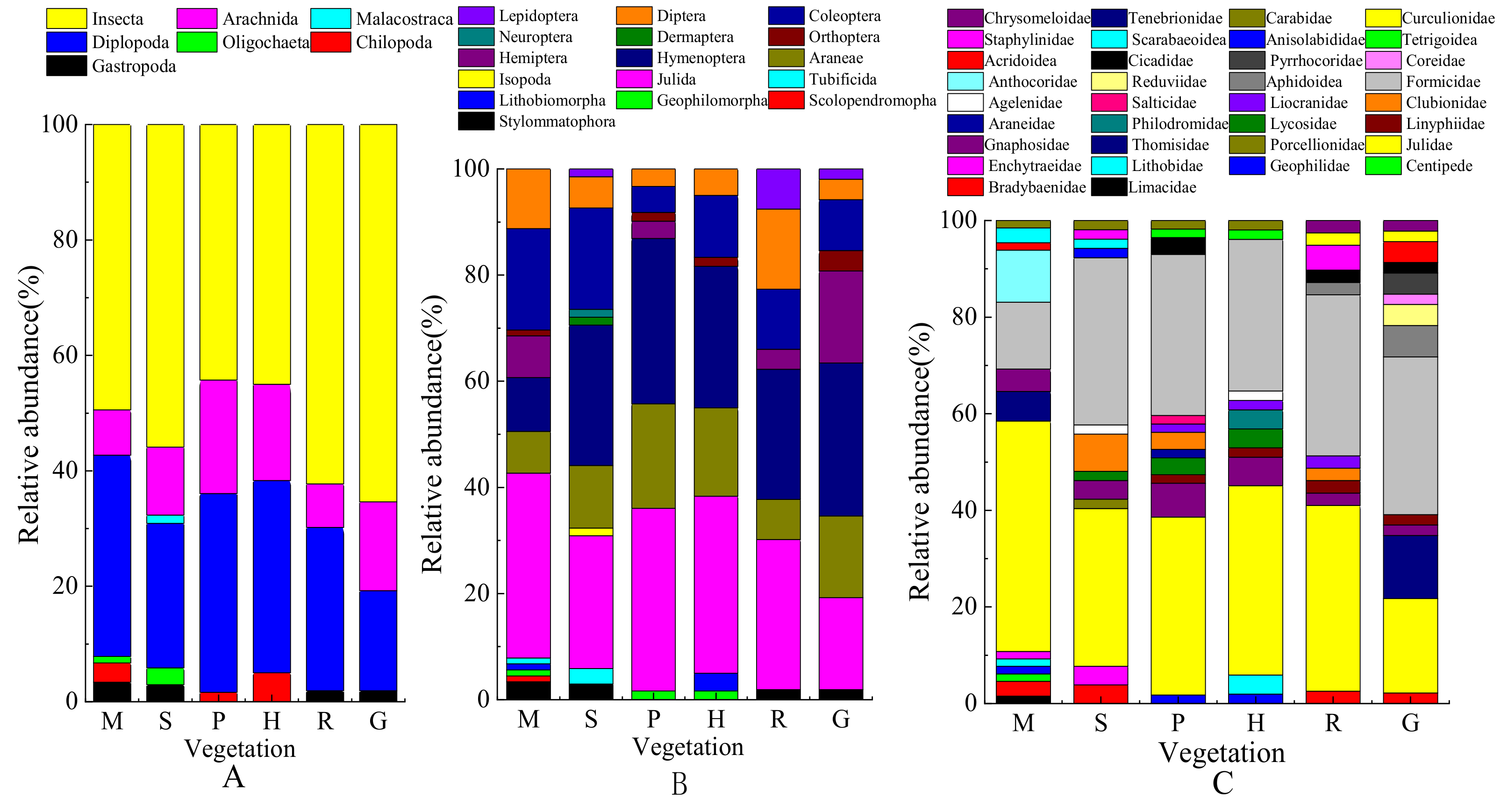
3.2. Composition of Soil Macrofaunal Community and Abundance of Dominate Taxonomical Groups
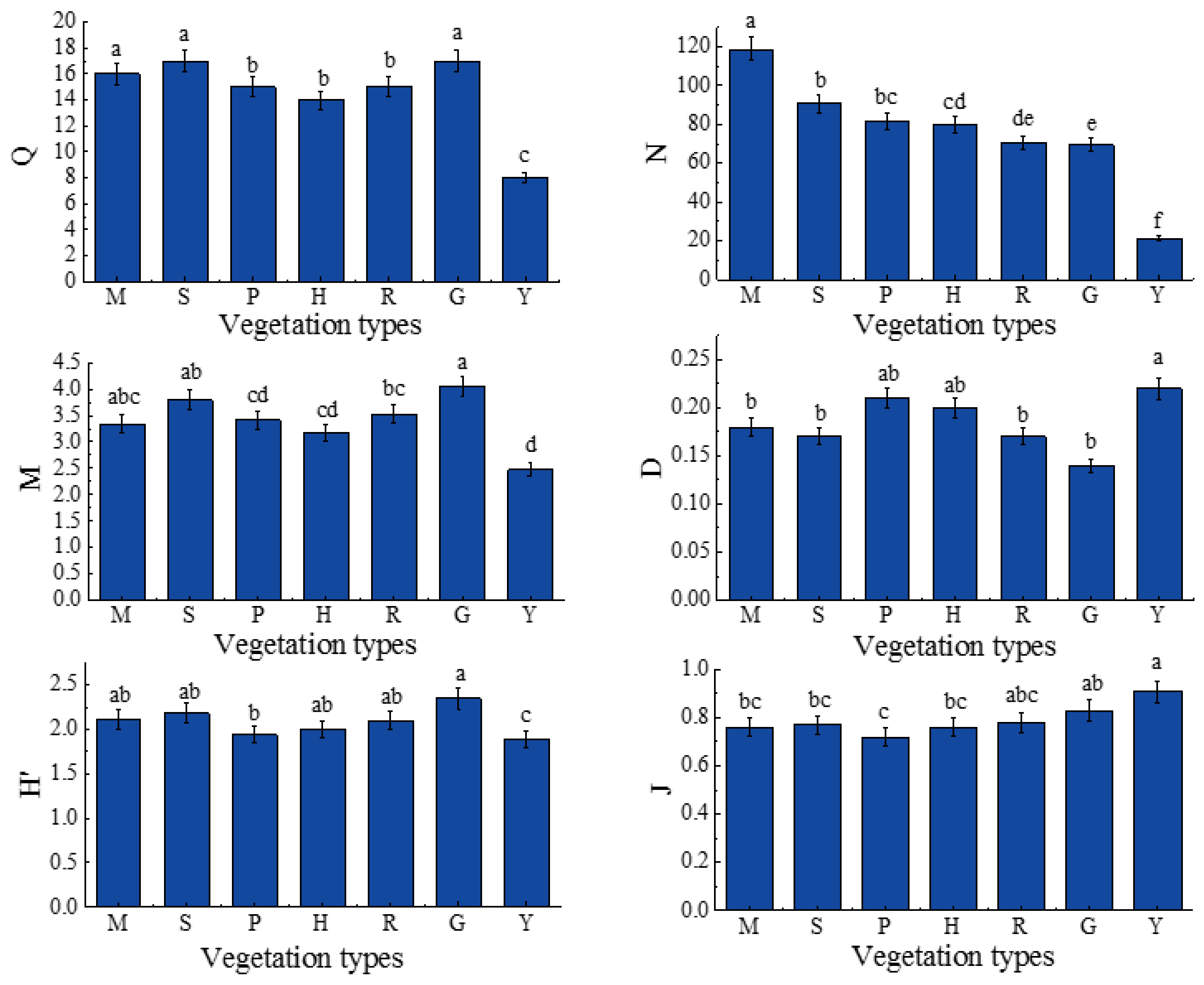
3.3. The Abundance, Group Richness and Diversity Index of Soil Macrofaunal Communites
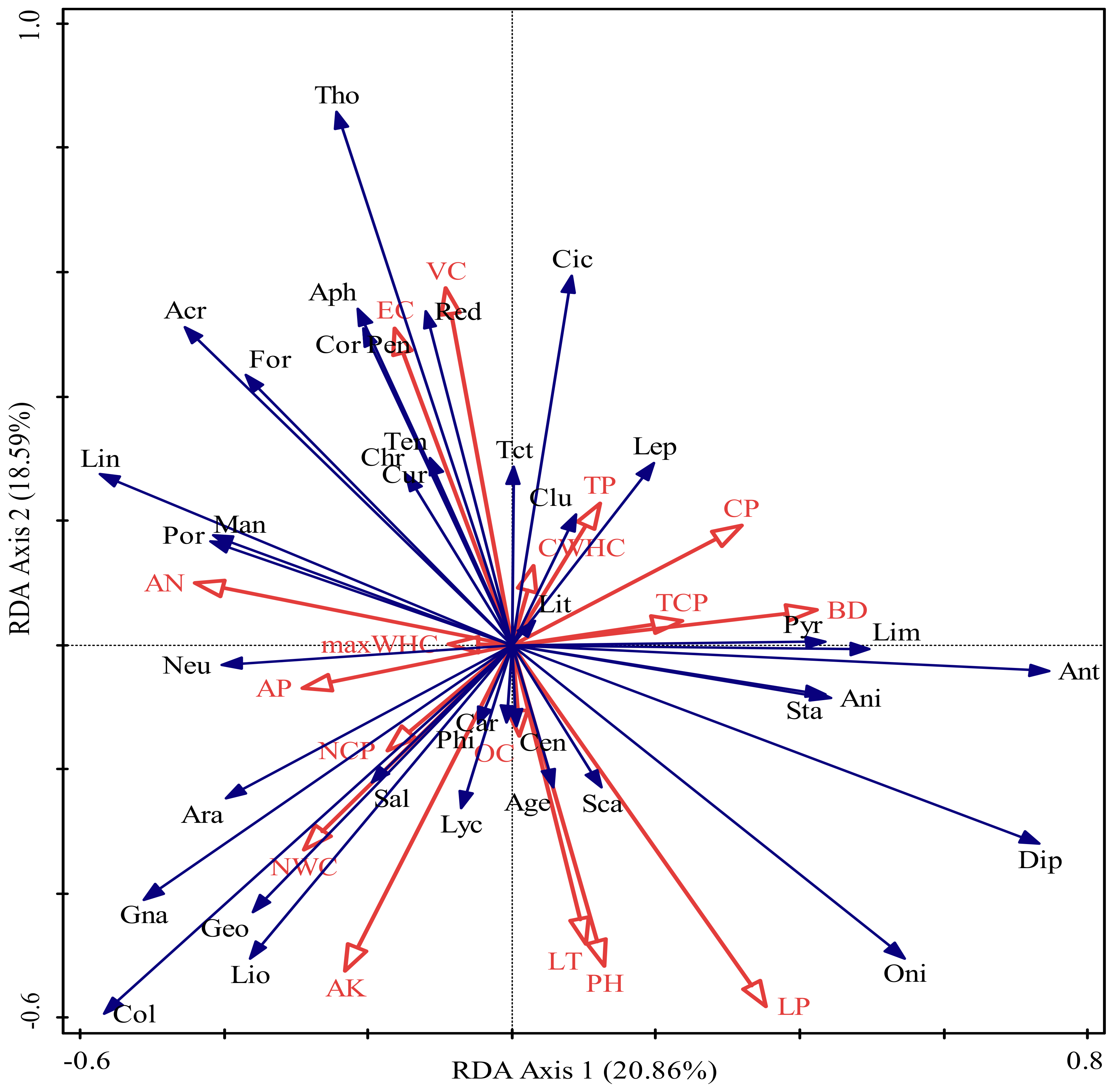
3.4. The Contribution of Litter and Soil Variables to Soil Macrofaunal Communities
4. Discussion
4.1. Soil Physical and Chemical Factors of Different Vegetation Types
4.2. Community Composition and Diversity of Soil Macrofauna in Different Vegetations
4.3. The Relationship between the Community Composition of Soil Macrofauna and Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mahmoud, S.H.; Gan, T.Y. Impact of anthropogenic climate change and human activities on environment and ecosystem services in arid regions. Sci. Total Environ. 2018, 633, 1329–1344. [Google Scholar] [CrossRef]
- Feng, J.; Wang, J.; Yao, S.; Ding, L. Dynamic assessment of forest resources quality at the provincial level using AHP and cluster analysis. Comput. Electron. Agric. 2016, 124, 184–193. [Google Scholar] [CrossRef]
- Luo, Y.; Lü, Y.; Fu, B.; Zhang, Q.; Li, T.; Hu, W.; Comber, A. Half century change of interactions among ecosystem services driven by ecological restoration: Quantification and policy implications at a watershed scale in the Chinese Loess Plateau. Sci. Total Environ. 2019, 651, 2546–2557. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Zhu, G.Y.; Deng, L.; Chen, L.; Shangguan, Z.P. Effects of natural vegetation restoration and afforestation on soil carbon and nitrogen storage in the Loess Plateau, China. Ying Yong Sheng Tai Xue Bao 2018, 29, 2163–2172. [Google Scholar] [CrossRef]
- Wang, B.; Gao, P.; Niu, X.; Sun, J. Policy-driven China’s Grain to Green Program: Implications for ecosystem services. Ecosyst. Serv. 2017, 27, 38–47. [Google Scholar] [CrossRef]
- Sisay, M.; Ketema, H. Variation in abundance and diversity of soil invertebrate macro-fauna and some soil quality indicators under agroforestry based conservation tillage and maize-based conventional tillage in Southern Ethiopia. Int. J. Multidiscip. Res. Dev. 2015, 2, 100–107. [Google Scholar]
- Riutta, T.; Clack, H.; Crockatt, M.; Slade, E. Landscape-Scale Implications of the Edge Effect on Soil Fauna Activity in a Temperate Forest. Ecosystems 2016, 19, 534–544. [Google Scholar] [CrossRef] [Green Version]
- De Deyn, G.B.; Raaijmakers, C.E.; Zoomer, H.R.; Berg, M.P.; de Ruiter, P.C.; Verhoef, H.A.; Bezemer, T.M.; van der Putten, W.H. Soil invertebrate fauna enhances grassland succession and diversity. Nature 2003, 422, 711–713. [Google Scholar] [CrossRef]
- Fujii, S.; Takeda, H. Succession of soil microarthropod communities during the aboveground and belowground litter decomposition processes. Soil Biol. Biochem. 2017, 110, 95–102. [Google Scholar] [CrossRef]
- Xiang, C.-G.; Feng, G.-L.; Pan, G.-X.; Zhang, P.-J. Soil animal diversity and its respondence to ecological restoration and reconstruction in Southwest Karst area-Take the Mile Bailongdong area in Yunnan Province as an example. J. Hunan Agric. Univ. 2007, 33, 324. [Google Scholar]
- Jiayuan, W.; Fucang, Q.; Zhenqi, Y.; Xiaotong, R.; Fei, F.; Ying, Z. Characteristics of Soil Animal Communities Under Different Land Use in Gully Area of Loess Plateau. J. Agric. Sci. Technol. 2021, 23, 156. [Google Scholar]
- Uwizeyimana, H.; Wang, M.; Chen, W.; Khan, K. The eco-toxic effects of pesticide and heavy metal mixtures towards earthworms in soil. Environ. Toxicol. Pharmacol. 2017, 55, 20–29. [Google Scholar] [CrossRef]
- Siebert, J.; Ciobanu, M.; Schädler, M.; Eisenhauer, N. Climate change and land use induce functional shifts in soil nematode communities. Oecologia 2020, 192, 281–294. [Google Scholar] [CrossRef]
- Lan, L.Y.; Yang, W.Q.; Wu, F.Z.; Liu, Y.W.; Yang, F.; Guo, C.H.; Chen, Y.; Tan, B. Effects of soil fauna on microbial community during litter decomposition of Populus simonii and Fargesia spathacea in the subalpine forest of western Sichuan, China. Ying Yong Sheng Tai Xue Bao 2019, 30, 2983–2991. [Google Scholar]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef]
- Sylvain, Z.A.; Wall, D.H. Linking soil biodiversity and vegetation: Implications for a changing planet. Am. J. Bot. 2011, 98, 517–527. [Google Scholar] [CrossRef]
- Yin, X.; Song, B.; Dong, W.; Xin, W.; Wang, Y. A review on the eco-geography of soil fauna in China. J. Geogr. Sci. 2010, 20, 333–346. [Google Scholar] [CrossRef]
- Carrillo, Y.; Ball, B.A.; Bradford, M.A.; Jordan, C.F.; Molina, M. Soil fauna alter the effects of litter composition on nitrogen cycling in a mineral soil. Soil Biol. Biochem. 2011, 43, 1440–1449. [Google Scholar] [CrossRef]
- Eissfeller, V.; Langenbruch, C.; Jacob, A.; Maraun, M.; Scheu, S. Tree identity surpasses tree diversity in affecting the community structure of oribatid mites (Oribatida) of deciduous temperate forests. Soil Biol. Biochem. 2013, 63, 154–162. [Google Scholar] [CrossRef]
- Baobao, H.; Siping, C.; Yang, L.; Jiao, L.; Dahong, Q.; Changhai, L. Emporal and spatial changes of soil animals in returning farmland to forestland in different years in Wuqi County. J. Arid Land Resour. Environ. 2020, 34, 130–136. [Google Scholar]
- Zhu, Y.; Wang, Y.; Chen, L.; Li, Z. Does non-native black locust afforestation affect soil biodiversity at the regional scale? Case study of soil macroinvertebrates across the Chinese Loess Plateau. Catena 2021, 200, 105171. [Google Scholar] [CrossRef]
- Yin, W. Retrieval Atlas of Soil Animals in China; Science Press: Beijing, China, 1998. [Google Scholar]
- Shidan, B. Soil Agrochemical Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Ge, F.; Zhou, Z.; Liu, J.; Wang, N.; Li, J.; Jiang, P. Distribution Characteristics of Root and Their Effect on Soil Separation Rate of Four Typical Plants in Loess Hilly Region. J. Soil Water Conserv. 2017, 31, 164–169. [Google Scholar]
- Zhou, Z.; Wang, C.; Zhang, Q. The effect of land use change on soil carbon, nitrogen, and phosphorus contents and their stoichiometry in temperate sapling stands in northeastern China. Acta Ecol. Sin. 2015, 35, 6694–6702. [Google Scholar]
- Ren-Tao, L.; Ha-Lin, Z.; Xue-Yong, Z. Effects of different afforestation types on soil faunal diversity in Horqin Sand Land. Ying Yong Sheng Tai Xue Bao 2012, 23, 1104–1110. [Google Scholar]
- Zhang, Q.; Yu, X.; Qian, J.; Xiong, T. Distribution characteristics of plant communities and soil organic matter and main nutrients in the Poyang Lake Nanji Wetland. Acta Ecol. Sin. 2012, 32, 3656–3669. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Liu, G.; Xu, M. Effect of vegetation restoration on soil nutrient changes in Zhifanggou watershed of loess hilly region. Bull. Soil Water Conserv. 2001, 22, 1–5. [Google Scholar]
- Xu, H.; Wei-Quan, W.; Jian, Z.; Wan-Qin, Y.; Yang, L.; Yu-Mei, H. Soil faunal diversity under typical alpine vegetations in West Sichuan. Ying Yong Sheng Tai Xue Bao 2010, 21, 181–190. [Google Scholar]
- Siemann, E.; Tilman, D.; Haarstad, J.; Ritchie, M. Experimental tests of the dependence of arthropod diversity on plant diversity. Am. Nat. 1998, 152, 738–750. [Google Scholar] [CrossRef]
- Halin, Z.; Rentao, L.; Ruilian, Z. Effects of changes in land use on soil macro-arthropod communities in Horqin sand land. Acta Pedol. Sin. 2013, 50, 413–418. [Google Scholar]
- Lingjun, M.; Limin, Z.; Limei, Z.; Zhongke, F. Responses of soil macrofaunal communities to tourism trample in the Wudalianchi Yaoquan Mountains. Acta Ecol. Sin. 2016, 36, 6607–6617. [Google Scholar]
- Shi-Ke, I.; Juan-Juan, W.; Sha, Z.; Qi, Z.; Xiang, L.; Wen-Jing, Z.; Wen-Hui, Y. Soil fauna community in the litter of six woodlands in Shanghai urban area. Resour. Environ. Yangtze Basin 2016, 25, 1–8. [Google Scholar]
- Li, F.-R.; Liu, J.-L.; Liu, C.-A.; Liu, Q.-J.; Niu, R.-X. Shrubs and species identity effects on the distribution and diversity of ground-dwelling arthropods in a Gobi desert. J. Insect Conserv. 2013, 17, 319–331. [Google Scholar] [CrossRef]
- Liu, J.-L.; Li, F.-R.; Liu, C.-A.; Liu, Q.-J. Influences of shrub vegetation on distribution and diversity of a ground beetle community in a Gobi desert ecosystem. Biodivers. Conserv. 2012, 21, 2601–2619. [Google Scholar] [CrossRef]
- Yuanyuan, L.; Jiahui, L.; Ziqian, X.; Hanmei, X.; Juanping, N.; Shide, H.; Honghua, R. Effects of organic fertilizer application and understory plant removal on the diversity of soil arthropods in poplar plantations. Acta Ecol. Sin. 2021, 41, 2761–2769. [Google Scholar]
- Yi, W.; Wei, W.; Xing-zhong, Y.; Li-ding, C.; Lei, Y. Interrelationships between soil fauna and soil environmental factors in China: Research advance. Ying Yong Sheng Tai Xue Bao 2010, 21, 2441–2448. [Google Scholar]
- Zhi-Wei, L.; Mei, H.; Hai-Shan, D.; He, Y.; Yu-Chen, Z.; Yan Jin, L.J. Effects of soil meso- and micro-fauna on litter decomposition under nitrogen deposition and rainfall changes. Chin. J. Appl. Ecol. 2021, 32, 4279–4288. [Google Scholar]
- He, Y.; Me, H.; Bayinnamula, Z.; Jing, L.; Jin, Y.; Yuchen, Z.; Zhiwei, L. Effects of water and nitrogen treatments on root decomposition in a Stipa breviflora desert steppe. Chin. J. Appl. Environ. Biol. 2020, 26, 1169–1175. [Google Scholar]
- Guangbin, Y.; Xiaodong, Y. Characteristics of litter and soil arthropod communities at different successional stages of tropical forests. Biodivers. Sci. 2010, 15, 188. [Google Scholar] [CrossRef]
- Jiliang, L.; Wenzhi, Z.; Fengrui, L.; Yibin, B. The relationship of ground spider and beetle assemblage with environmental factors in the natural and artificial fixed-sand shrub forests. Acta Ecol. Sin. 2020, 40, 7987–7996. [Google Scholar]
- Adejuyigbe, C.O.; Tian, G.; Adeoye, G.O. Microcosmic study of soil microarthropod and earthworm interaction in litter decomposition and nutrient turnover. Nutr. Cycl. Agroecosyst. 2006, 75, 47–55. [Google Scholar] [CrossRef]
- Shu-Tang, L.; Pei-Li, L.; Xiao-Ri, H.; Fang-Gong, S.; Rui, C. Effects of Long-Term Located Fertilization on Biological Environment of Non-Calcareous Fluro-Aquic Soil. Bull. Soil Water Conserv. 2006, 1, 26–29. [Google Scholar]
- Wang, M.; Fu, S.; Xu, H.; Wang, M.; Shi, L. Ecological functions of millipedes in the terrestrial ecosystem. Biodivers. Sci. 2018, 26, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Litavský, J.; Majzlan, O.; Stašiov, S.; Svitok, M.; Fedor, P. The associations between ground beetle (Coleoptera: Carabidae) communities and environmental condition in floodplain forests in the Pannonian Basin. Eur. J. Entomol. 2021, 118, 14–23. [Google Scholar] [CrossRef]
- Groner, E.; Ayal, Y. The interaction between bird predation and plant cover in determining habitat occupancy of darkling beetles. Oikos 2001, 93, 22–31. [Google Scholar] [CrossRef]
- Shrewsbury, P.M.; Raupp, M.J. Do top-down or bottom-up forces determine Stephanitis pyrioides abundance in urban landscapes? Ecol. Appl. 2006, 16, 262–272. [Google Scholar] [CrossRef]
- Braschler, B.; Baur, B. Effects of experimental small-scale grassland fragmentation on spatial distribution, density, and persistence of ant nests. Ecol. Èntomol. 2003, 28, 651–658. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Plot | Altitude (m) | Aspect | Slope Position | Latitude-Longitude | Main Herbaceous Vegetation |
|---|---|---|---|---|---|
| Armeniaca sibirica and Populus simonii mixed forest | 1428.33 | Half shady slope | Slope top | 108°12′22″ E, 36°50′44″ N | Artemisia sacrorum, Lespedeza daurica |
| Robinia pseudoacacia | 1309.15 | Half shady slope | Midslop | 108°12′19″ E, 36°52′25″ N | Lespedeza daurica, Setaria viridis |
| Populus simonii | 1327.51 | Half shady slope | Downhill | 108°12′23″ E, 36°52′33″ N | Onopordum acanthium, Artemisia selengensis, Heteropappus altaicus |
| Populus hopeiensis | 1316.40 | Shady slope | Downhill | 108°12′43″ E, 36°52′17″ N | Artemisia sacrorum, Lespedeza bicolor |
| Hippophae rhamnoides | 1354.63 | Half shady slope | Uphill | 108°12′40″ E, 36°52′14″ N | Adenophora stenanthina, Potentilla chinensis, Artemisia sacrorum, Rubia cordifolia |
| Abandoned grassland | 1332.99 | Half shady slope | Uphill | 108°12′51″ E, 36°52′37″ N | Patrinia rupestris, Kochia scoparia, Agropyron cristatum, Lespedeza bicolor |
| Environmental Factors | M | S | P | H | R | G |
|---|---|---|---|---|---|---|
| BD (g/cm3) | 1.23 ± 0.03 ab | 1.26 ± 0.12 ab | 1.23 ± 0.05 ab | 1.16 ± 0.02 ab | 1.3 ± 0.04 a | 1.12 ± 0.16 b |
| NWC (%) | 14.20 ± 0.23 b | 17.47 ± 2.96 a | 10.55 ± 0.16 cd | 17.62 ± 0.77 a | 9.90 ± 1.65 d | 13.35 ± 0.36 bc |
| Max WHC (%) | 45.70 ± 3.29 ab | 35.70 ± 5.70 b | 42.71 ± 5.55 ab | 48.64 ± 2.46 ab | 37.57 ± 1.39 b | 50.5 ± 10.62 a |
| CWHC (%) | 35.47 ± 2.90 ab | 23.94 ± 3.81 b | 31.92 ± 0.19 ab | 40.98 ± 0.90 a | 31.45 ± 0.42 ab | 38.27 ± 5.58 a |
| CP (%) | 43.43 ± 2.95 ab | 29.83 ± 2.12 d | 39.28 ± 1.85 c | 47.40 ± 0.64 a | 40.80 ± 1.58 bc | 42.15 ± 1.78 bc |
| NCP (%) | 12.5 ± 3.77 a | 14.6 ± 1.32 a | 13.1 ± 6.64 a | 8.83 ± 2.22 a | 7.87 ± 1.99 a | 12.9 ± 4.37 a |
| TCP (%) | 55.93 ± 2.75 a | 44.43 ± 2.38 b | 52.38 ± 5.90 ab | 56.23 ± 1.79 a | 48.68 ± 0.44 ab | 55.05 ± 4.77 a |
| OC (g/kg) | 7.37 ± 0.39 b | 7.17 ± 0.49 b | 3.54 ± 0.16 d | 10.15 ± 0.51 a | 8.02 ± 0.45 b | 5.52 ± 0.39 c |
| pH | 8.36 ± 0.03 bc | 8.43 ± 0.04 abc | 8.48 ± 0.03 a | 8.51 ± 0.05 a | 8.44 ± 0.03 ab | 8.33 ± 0.09 c |
| EC (μs/cm3) | 88.5 ± 3.42 b | 85.27 ± 3.21 b | 84.57 ± 1.14 b | 84.47 ± 0.12 b | 84.43 ± 2.16 b | 96.8 ± 3.12 a |
| AP (mg/kg) | 5.37 ± 0.17 c | 9.30 ± 0.94 a | 6.50 ± 0.49 b | 4.17 ± 0.25 d | 5.27 ± 0.31 c | 6.77 ± 0.12 b |
| AN (mg/ kg) | 9.02 ± 0.31 c | 11.6 ± 0.29 b | 7.97 ± 0.48 d | 9.33 ± 0.21 c | 6.15 ± 0.64 e | 28.3 ± 0.29 a |
| AK (mg/kg) | 90.67 ± 8.65 b | 156.67 ± 8.18 a | 79.33 ± 6.13 b | 92 ± 2.16 b | 53.33 ± 2.62 c | 84 ± 5.72 b |
| TP (g/kg) | 0.19 ± 0.06 bc | 0.18 ± 0.07 bc | 0.51 ± 0.03 a | 0.15 ± 0.04 c | 0.55 ± 0.02 a | 0.25 ± 0.01 b |
| VC (%) | 70 ± 3.00 b | 50 ± 5.2 c | 60 ± 4.5 b | 90 ± 5.34 a | 70 ± 4.12 b | 94 ± 3.61 a |
| LT (cm) | 2 ± 0.15 d | 3.43 ± 0.21 c | 5.3 ± 0.2 a | 4.8 ± 0.15 b | 1 ± 0.15 e | 1.2 ± 0.16 e |
| LP (g/m2) | 83.1 ± 1.50 c | 43.75 ± 1.39 d | 99.67 ± 1.94 b | 103.83 ± 2.15 a | 37.92 ± 1.94 e | 37.5 ± 0.91 e |
| Explanatory Variables | Explains % | Contribution % | Pseudo-F | p |
|---|---|---|---|---|
| VC (%) | 11.1 | 12.9 | 2 | 0.03 |
| AN (mg/ kg) | 10.7 | 12.4 | 2 | 0.038 |
| LP (g/m2) | 9.2 | 10.7 | 1.9 | 0.04 |
| BD (g/cm3) | 5.9 | 6.9 | 1.2 | 0.272 |
| Max WHC (%) | 6.3 | 7.3 | 1.3 | 0.208 |
| AK (mg/kg) | 5.4 | 6.3 | 1.2 | 0.32 |
| LT (cm) | 5.2 | 6.1 | 1.1 | 0.368 |
| TP (g/kg) | 4.8 | 5.6 | 1 | 0.392 |
| EC (μs/cm3) | 5.6 | 6.5 | 1.3 | 0.244 |
| TCP (%) | 4.3 | 5 | 1 | 0.528 |
| AP (mg/kg) | 4 | 4.7 | 0.9 | 0.568 |
| OC (g/kg) | 5.8 | 6.7 | 1.3 | 0.264 |
| NCP (%) | 5.2 | 6 | 1.3 | 0.324 |
| CWHC (%) | 4.4 | 5.1 | 1.1 | 0.4 |
| NWC (%) | 4 | 4.7 | 1 | 0.436 |
| PH | 2.3 | 2.7 | 0.4 | 0.694 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Liu, C.; Ai, N.; Tuo, X.; Zhang, Z.; Gao, R.; Qin, J.; Yuan, C. Characteristics of Soil Macrofauna and Its Coupling Relationship with Environmental Factors in the Loess Area of Northern Shaanxi. Sustainability 2022, 14, 2484. https://doi.org/10.3390/su14052484
Zhou Y, Liu C, Ai N, Tuo X, Zhang Z, Gao R, Qin J, Yuan C. Characteristics of Soil Macrofauna and Its Coupling Relationship with Environmental Factors in the Loess Area of Northern Shaanxi. Sustainability. 2022; 14(5):2484. https://doi.org/10.3390/su14052484
Chicago/Turabian StyleZhou, Yongwei, Changhai Liu, Ning Ai, Xianghui Tuo, Zhiyong Zhang, Rui Gao, Jiafeng Qin, and Caixia Yuan. 2022. "Characteristics of Soil Macrofauna and Its Coupling Relationship with Environmental Factors in the Loess Area of Northern Shaanxi" Sustainability 14, no. 5: 2484. https://doi.org/10.3390/su14052484
APA StyleZhou, Y., Liu, C., Ai, N., Tuo, X., Zhang, Z., Gao, R., Qin, J., & Yuan, C. (2022). Characteristics of Soil Macrofauna and Its Coupling Relationship with Environmental Factors in the Loess Area of Northern Shaanxi. Sustainability, 14(5), 2484. https://doi.org/10.3390/su14052484