Abstract
The widespread use of tetracycline antibiotics in the poultry and cattle sectors endangers both human health and the terrestrial ecosystem. Chlortetracyclines (CTCs), in particular, have been proven to affect soil microorganisms in addition to plants in the terrestrial ecosystem. In order to assess the effects of CTC on soil properties, eggplant growth, and soil microorganisms, a potted experiment was carried out in this study. CTC significantly reduced the levels of ammonium nitrogen (NH4+–N) and nitrite nitrogen (NO2−–N) in soil. Meanwhile, the eggplant’s growth was clearly hampered. CTC dramatically and dose-dependently lowered the fluorescence parameters except the quantum yield of non-regulated energy dissipation (ΦNO). Rhodoplanes and Cupriavidus, which were involved in N cycle, were enriched by 10 mg/kg CTC, according to results about different microorganisms at the genus level. Flavisolibacter was reduced by 10 and 50 mg/kg CTC, while Methylosinus and Actinocorallia were enriched by 250 mg/kg CTC. Redundancy analysis highlighted the profound impact of CTC on the soil microbial community, where strong correlations were observed with soil potential of hydrogen (pH), nitrate nitrogen (NO3−–N), and NO2−–N. These findings demonstrated the interdependence between the microbial community and soil characteristics, with CTC primarily affecting the microbes responsible for nitrogen cycling. Consequently, chlortetracycline poses potential hazards to both eggplant plants and the soil microbes in eggplant cultivation soil.
1. Introduction
Tetracycline (TC) antibiotics, one of the primary antibiotics groups, have been used extensively in human and animal medicine to treat and prevent disease as well as to increase growth rates in livestock and poultry industries [1]. The repeated and widespread use of TCs has led to their accumulation in aquatic and terrestrial environments, subsequently triggering detrimental secondary toxic effects on the non-target organisms [2]. Notably, extensive soil sampling across several provinces in China has revealed alarmingly high concentrations of TCs, with average levels ranging from 102 to 1687 μg/kg [3]. One of the most prevalent TCs and frequently found in agricultural soils is chlortetracycline (CTC), which has a higher adsorption coefficient value (Kd) [4,5]. It has been reported that CTC concentrations ranged from 2.94 to 1590 μg/kg in agriculture soil of China [3,6,7]. In 2001, 4.6–7.3 μg/kg CTC were found in Northern Germany [8]. According to Lee et al., the predicted environmental concentrations of CTC in soil in the world were in a range of 3.42–67.59 μg/kg soil, for a 90% confidence level [9]. Thus, China, in which the CTC concentration is higher, faces more severe CTC pollution.
In terrestrial environments, the accumulation of TCs can produce toxic effects on various components such as soil flora, fauna, and microorganisms [2,10,11]. Ma et al. [12] found soil nitrification potential and dehydrogenase activity was greatly stimulated after 28 days exposure of 30 mg/kg OTC. The soil microbial community serves as the foundation of soil ecological function and plays a vital role in soil nutrient cycling (such as carbon and nitrogen), soil fertility enhancement, and promotion of plant diversity [13,14]. The introduction of TCs can create a selective pressure on bacteria, inducing antibiotic resistance and subsequently altering the soil microbial community structure and function [15,16]. Zheng et al. [17] found tetracycline has a concentration-dependent impact on the structure of soil bacterial community. OTC, one of TCs, also can significantly reduce the abundances of certain genera which is related to plant growth-promoting in soil, such as Arthrobacter, Gemmatimonas, and Sphingomonas [2]. The study by Han et al. [18] which focused on the effects of CTC on soil microbial community discovered that microbial functional diversity followed a suppression-recovery-stimulation pattern in CTC-treated manured soil.
In addition to destroying the microbial community in the soil environment, TCs pollution would also be absorbed and accumulated by edible plants and even affect the growth of vegetables, posing a threat to global food security and soil ecosystem function, which has attracted growing concerns [19,20]. It has been reported that TCs can prevent the growth of crops and vegetables, such as wheat, ginger, and Ipomoea aquatica Forsk [2,19,21]. The influence of TCs on edible plants is closely related to the frequent use of livestock and poultry manure. In vegetable planting regions, livestock and poultry manure are frequently utilized as fertilizer to increase productivity, promote green and low-carbon agriculture sustainable development, improve the rural ecological environment, and achieve the double carbon objective [22]. However, antibiotics are typical pollutants in livestock and poultry manure, and long-term manure application will inevitably introduce a considerable number of exogenous antibiotics into the soil, particularly vegetable-based soil [6,22,23]. TCs which were detected in high concentrations in vegetable-based soil can affect the growth of vegetables, and even be absorbed and accumulated by vegetables, posing a serious threat to vegetable security and soil ecological security [24,25,26,27]. Studies about the effects of CTC on vegetables and crops growth have demonstrated the above results. Cheong et al. [28] found that CTC diminished chlorophyll content in leaves and photosynthetic efficiency, and downregulated the genes involved in the primary root growth of Brassica campestris seedlings. Guo et al. [2] found that antibiotics’ bactericidal capabilities significantly decreased the abundances of certain rhizobacteria, which had a negative influence on biomass and height of wheat seedling. Plant growth, development, and biomass accumulation are all influenced by soil microbes, particularly rhizosphere microorganisms [29,30,31]. Thus, to understand the impact of antibiotics on plant growth, it is critical to investigate the microbial community structure in soil.
Eggplant (Solanum melongena), a vegetable crop distributed worldwide, has considerable nutritional benefits due to its high concentration of vitamins, phenolics, and antioxidants [32]. The top producer and consumer of eggplant worldwide is China. However, little information regarding the responses of eggplant and rhizobacteria to CTC exposure is currently available. This study aimed to investigate the impacts of CTC on eggplant growth, relative chlorophyll content, fluorescence metrics, soil characteristics, and the soil microbial population by conducting a potted experiment with varying concentrations of CTC. This study can provide data support for the risk assessment of antibiotic residues in agriculture, so as to ensure the safety of crops and promote the sustainable development of agriculture.
2. Materials and Methods
2.1. Materials Collection
The eggplant seed (Solanum melongena) was purchased from seed market in Hefei, Anhui Province, China. The clean soil (0–15) cm was obtained from Hefei Normal University, nutritional soil was purchased from gardening market. Chlortetracycline (CTC, ≥97%) was purchased from Sigma-Adrich (St. Louis, MO, USA). Other chemicals were bought from Hefei Meifeng Chemical Co., Ltd. in Hefei, Anhui province, China.
2.2. Potted Experiment
The clean soil was air-dried, sieved to a diameter of 4 mm, and mixed with nutritional soil with 1:1 (v:v). Each pot was filled with 4 kg soils, and the moisture content of the soils was carefully regulated to reach 55% of their maximum water-holding capacity (WHC) with distilled water. CTC was dissolved in distilled water and then poured evenly into each pot to obtain the concentrations of 10.0 (L), 50.0 (M), and 250.0 (H) mg/kg CTC. There were three replicates for each treatment. The same amount of distilled water was added to the pot in 0 (CK) mg/kg CTC treatment.
Hydrogen peroxide (3%, v/v) was used to disinfect the seeds, and they were then thoroughly rinsed with sterile deionized water. In a climate chamber with a temperature of 25 °C and a humidity of 60%, the sterilized seeds were then placed in a tray, covered with gauze, and allowed to germinate. After germination, 12 uniform healthy eggplant seedlings were transplanted to the corresponding pots and planted for 70 days at a temperature of 25 °C with a cycle of 12 h of light and 10 h of darkness. Twice every week, distilled water was used to make up for water loss. After being in culture for 70 days, the eggplant and related rhizosphere soil were harvested. To gauge the growth traits, eggplants were gently removed from the pots. Each plant’s rhizosphere soil was collected using the shake-off method [33,34]. The soil that was weakly holding on was gently shaken off and thrown away. The soil that was firmly clung to the eggplant roots (0–1 mm) was brushed off and recorded as rhizosphere soil. All samples were divided into two parts: one part was immediately stored at −80 °C for soil DNA extraction; the other part was used to measure soil properties as follows.
2.3. Measurement of Soil Properties
The soil was air dried, crushed, and screened through a 2 mm sifter to measure its properties after the visible organic remains had been eliminated. Soil pH (1:2.5 soil/water suspension) was determined using pHS-25 (precision of 0.05, Leici., Shanghai, China) with the People’s Republic of China’s agricultural industry standard NY/T 1377-2007 as a guide [35]. The People’s Republic of China’s agricultural industry standard NY/T 1121.6-2006 was used to measure the content of soil organic matter (OM) by using standard solution titration of ferrous sulfate [36]. Following the National Environmental protection standards of the People’s Republic of China HJ 634-2012, potassium chloride solution extraction-spectrophotometry was used to measure the ammonium nitrogen (NH4+–N), nitrite nitrogen (NO2−–N), and nitrate nitrogen (NO3−–N) in soil [37]. The levels of microbial biomass N in soil were measured by fumigation [38].
2.4. Measurement of Growth and Physiological Indexes
The Soil Plant Analysis Development (SPAD)-502 Plus (Konica Minolta, Tokyo, Japan) chlorophyll meter was used to determine the relative chlorophyll amount of leaf (represented as SPAD value). The measuring point was the center of the first fully unfolded leaf at the top of the plant, avoiding the apparent vein. The three readings’ average values were calculated.
Junior—PAM (Walz, Effeltrich, Germany) based modulation fluorometer was used for the determination of chlorophyll fluorescence parameters. First, after exposing the measuring object to darkness for 30 min, the maximal photochemical efficiency of PS II (Fv/Fm) was determined. The kinetic parameters of chlorophyll fluorescence induction were then automatically measured under 190 μmol/m2/s of light intensity for 15 times using an artificial light source. The main components were electron transfer efficiency (ETR), photochemical quenching coefficient (qP), non-photochemical quenching coefficient (NPQ), quantum efficiency of Phtosynthetic system II (ΦPSII), quantum yield of regulated energy dissipation (ΦNPQ), and quantum yield of non-regulated energy dissipation (ΦNO).
After the above experiment, all plants were carefully dug up and labeled. First, the root soil was cleaned with distilled water, and then the root soil was cleaned with double distilled water three times. The root parts were used to measure the fresh weight, and a steel ruler was used to measure the root’s height from the base.
2.5. Soil Microbial Community Analysis
Total DNA extraction of soil samples from the three replicates in each treatment was carried out as follows: Total DNA were extracted using FastDNA Soil Kit (MP Biomedicals). The concentration and purity of the final soil DNA were measured by Nano-Drop 2000, a UV–vis spectrophotometer from NanoDrop Technologies, Willmington, DE, USA. And the DNA quality was determined by 1% agarose gel electrophoresis. The 16S rRNA gene of bacteria was amplified using primers 515F (5′-GTGYCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACNVGGGTWTCTAAT-3′) [39]. Before sequencing on an Illumina Miseq (Illumina, San Diego, CA, USA), the individual samples of the amplicons were first barcoded and then pooled to create the sequencing library. Mothur software (Version 1.35.1) was used to remove low-quality and chimeric reads from the raw sequencing data generated from Illumina Miseq. After quality filtration, all datasets were rarefied to 21,213 sequences to achieve same sequencing depth. High-quality readings were grouped into OTUs at a 97% identity level using the Quantitative Insights into Microbial Ecology (QIIME, version 1.9.1). The parallel_pick_open_reference_otus workflow script and the Greengenes 13_8 Database were both used to conduct OTU searches. The α-diversity indexes (Chao, ACE, Simpson, and Shannon) and Good’s coverage were calculated by QIIME. To determine the β-diversity of the soil microbiota, principal component analysis (PCA) and unweighted pair group method with arithmetic mean (UPGMA) clustering were carried out. The Ribosomal Database Project (RDP) Classifier (http://rdp.cme.msu.edu/, accessed on 30 September 2016) was used to classify representative sequences from each OUT using an 80% confidence level.
2.6. Statistical Analysis
All data in this study are expressed as the mean ± SD. Differences between the treatment groups and the control group with respect to growth indexes, physiological indexes, soil properties and α-diversity indexes were detected by ANOVA test followed by LSD multiple comparison using SPSS 21.0 software (SPSS Inc., Chicago, IL, USA). One-way PERMANOVA test by PAST was used to analyze the difference of bacterial community structure. Different microorganisms at genus compared with the control were detected by a t-test.
3. Results
3.1. Changes in Soil Properties
The physicochemical properties of eggplant cultivated soil were shown in Table 1. The ammonium nitrogen (NH4+–N) and nitrite nitrogen (NO2−–N) were significantly decreased by CTC (Table 1). The content of NH4+–N and NO2−–N in 250 mg/kg group were 48.33% and 67.27% lower than that in the control group, respectively. All CTC groups in this study showed lower nitrate nitrogen (NO3−–N) levels. In this study, microbial biomass nitrogen was increased by CTC with a dose–effect relationship.

Table 1.
Physicochemical properties of soil samples treated with different CTC concentrations (n = 3).
3.2. Effects of CTC on Eggplant Growth and Fluorescence Parameters
In this study, the growth of eggplant was inhibited by CTC-contaminated soil. As illustrated in Figure 1, the root fresh weight, root length and SPAD value in the CTC treatment groups demonstrated a concentration-dependent decrease with an increase in CTC dosage. Root fresh weight, root length, and SPAD value were all significantly (p < 0.05) decreased with the highest CTC treatment (250 mg/kg), by 38.58%, 63.49%, and 16.35%, respectively.
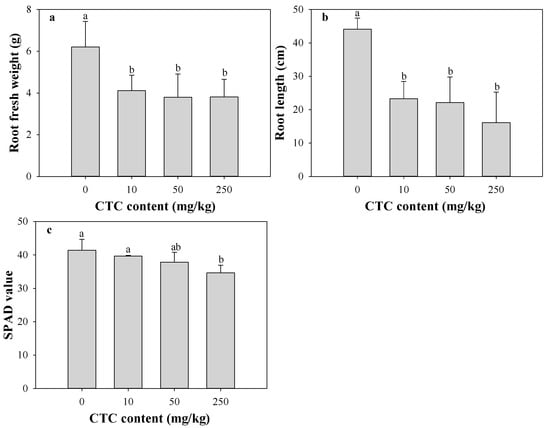
Figure 1.
Effects of CTC on plant root fresh weight biomass (a), root length (b) and SPAD value (c) of eggplant. Data are expressed as mean ± SD (n = 3). Data with different letters means significant difference at p < 0.05.
Table 2 showed that CTC posed a substantial impact on fluorescence parameters of eggplant. Fv/Fm value was decreased by 0.99 to 1.73% in the CTC groups, in which only 50 mg/kg group significantly decreased the Fv/Fm. The qP value was significantly decreased by CTC and decreased by 31.58% in the presence of 250 mg/kg CTC, indicating diminished electron transfer activity at PS II by CTC. The NPQ values rapidly declined with increasing CTC content and fell by 34.71% in the 250 mg/kg group, indicating heat dissipation performance of eggplant decreased. ETR and ΦPS II were dramatically reduced by CTC treatments with a dose–effect relationship. The most important indexes of light protection and light damage are ΦNPQ and ΦNO, respectively. And in this study, the change in ΦNPQ is small without significant difference. ΦNO values increased significantly in the presence of CTC and increased by 42.86% in 250 mg/kg group.
Table 2.
Chlorophyll fluorescence parameters of eggplants exposed to different CTC concentrations (n = 3).
3.3. Effects of CTC on Soil Microorganism Communities
3.3.1. CTC Effects on Soil Microbial Diversity
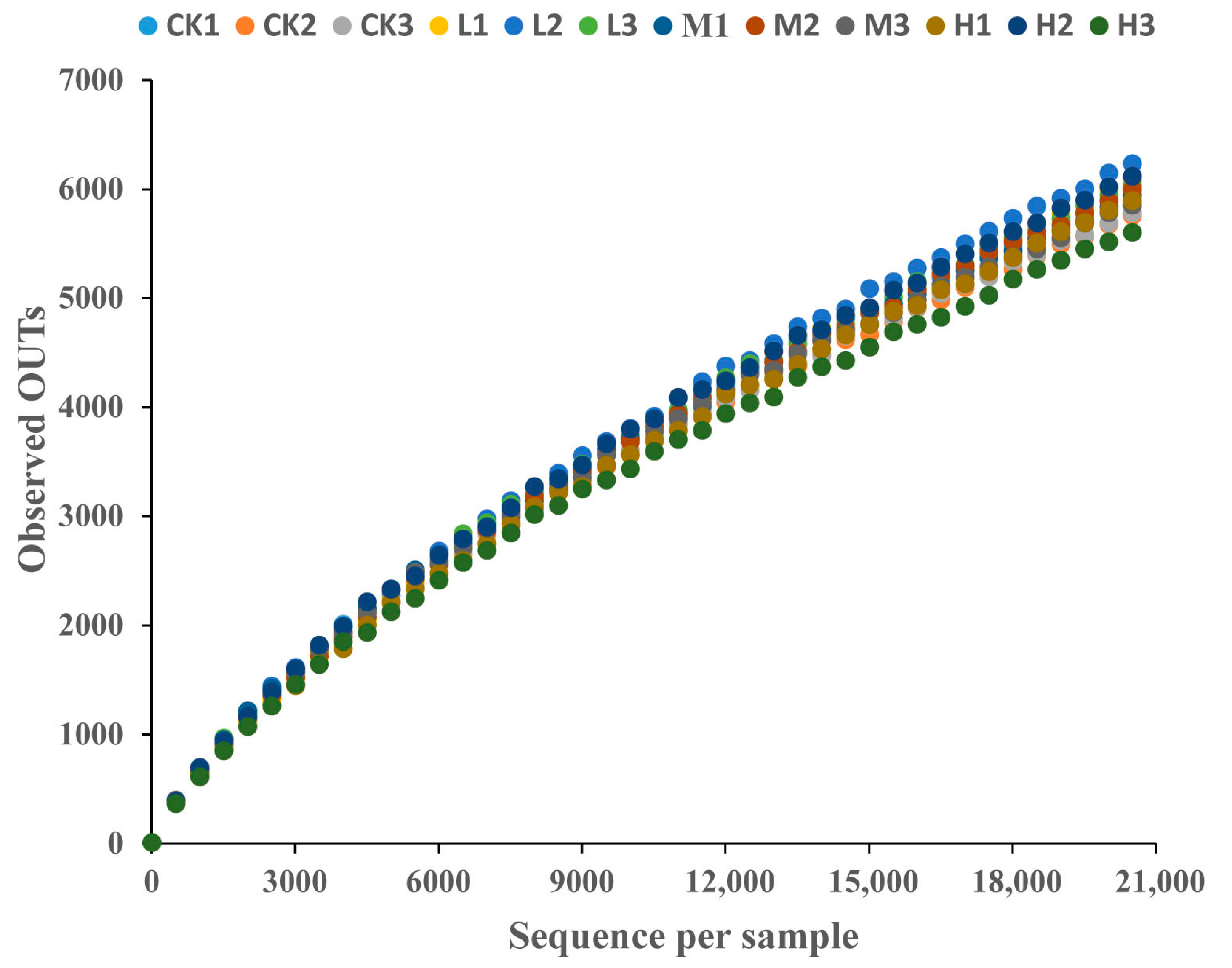
High-throughput sequencing of the 16S rRNA gene amplicons convincingly demonstrated the alterations in the soil microbiota in response to the chronic CTC exposure. A total of 254,556 sequences were obtained from all samples. The sequences’ rarefaction curves (Figure 2) revealed that enough sampling coverage was achieved for the soil samples.
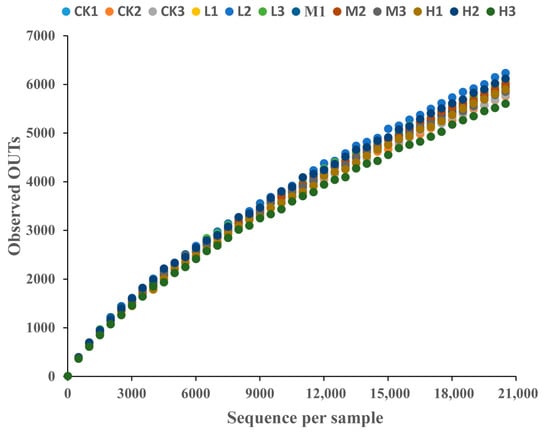
Figure 2.
Rarefaction curve of soil microbial DNA samples under different concentrations of CTC. CK 1, 2 and 3 are the 3 replicated of 0 mg/kg CTC. L1, 2 and 3 are the 3 replicated of 10 mg/kg. M1, 2 and 3 are the 3 replicated of 50 mg/kg CTC. H1, 2 and 3 are the 3 replicated of 250 mg/kg CTC.
As shown in Table 3, the Good’s coverage of each sample was greater than 80%. The observed OTUs in CTC groups were higher than those in control group. The community richness indexes Chao and ACE of soil microbiota were higher in CTC groups than those in the control group. Near parity existed in the community diversity indexes Simpson between the CTC groups and the control group. Table 3 indicated that CTC slightly increased the community richness, but without significant differences and had no effect on the community diversity.
Table 3.
Microbial α-diversities of soil samples treated with different CTC concentrations (n = 3).
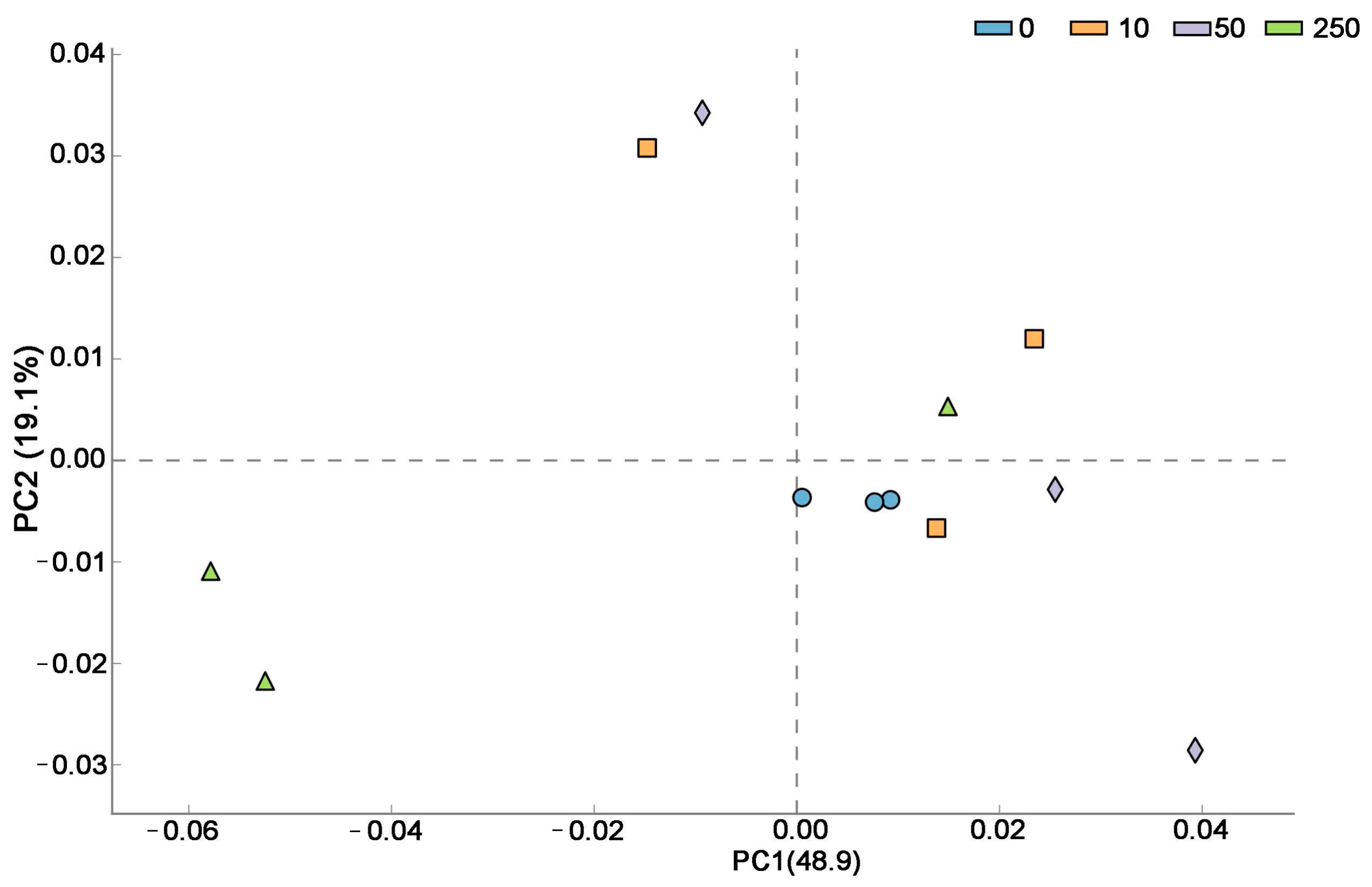
Figure 3 demonstrated that principal component (PC) 1 and 2 explained 68% of the total variance, with PC1 contributing 48.9%, showing that the first principal component was primarily responsible for the variations in microbes between control and CTC-treated soil. The PCA plot revealed that whereas soil samples from the CTC-treated groups displayed a more dispersed distribution, indicating some degree of variance, samples from the control group tended to be closely clustered, indicating a generally similar community makeup. The PCA plot in Figure 3 demonstrated that soil samples from several groups had a large difference in distribution distance, indicating that each group’s microbial community composition varied greatly.
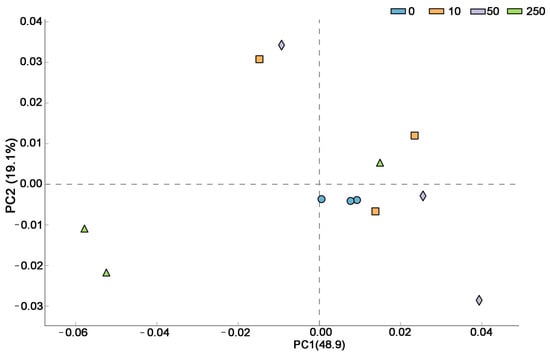
Figure 3.
The soil microorganism community patterns (β-diversity) in the control and CTC-treated soils by PCA.
3.3.2. CTC Effects on Soil Microbial Composition
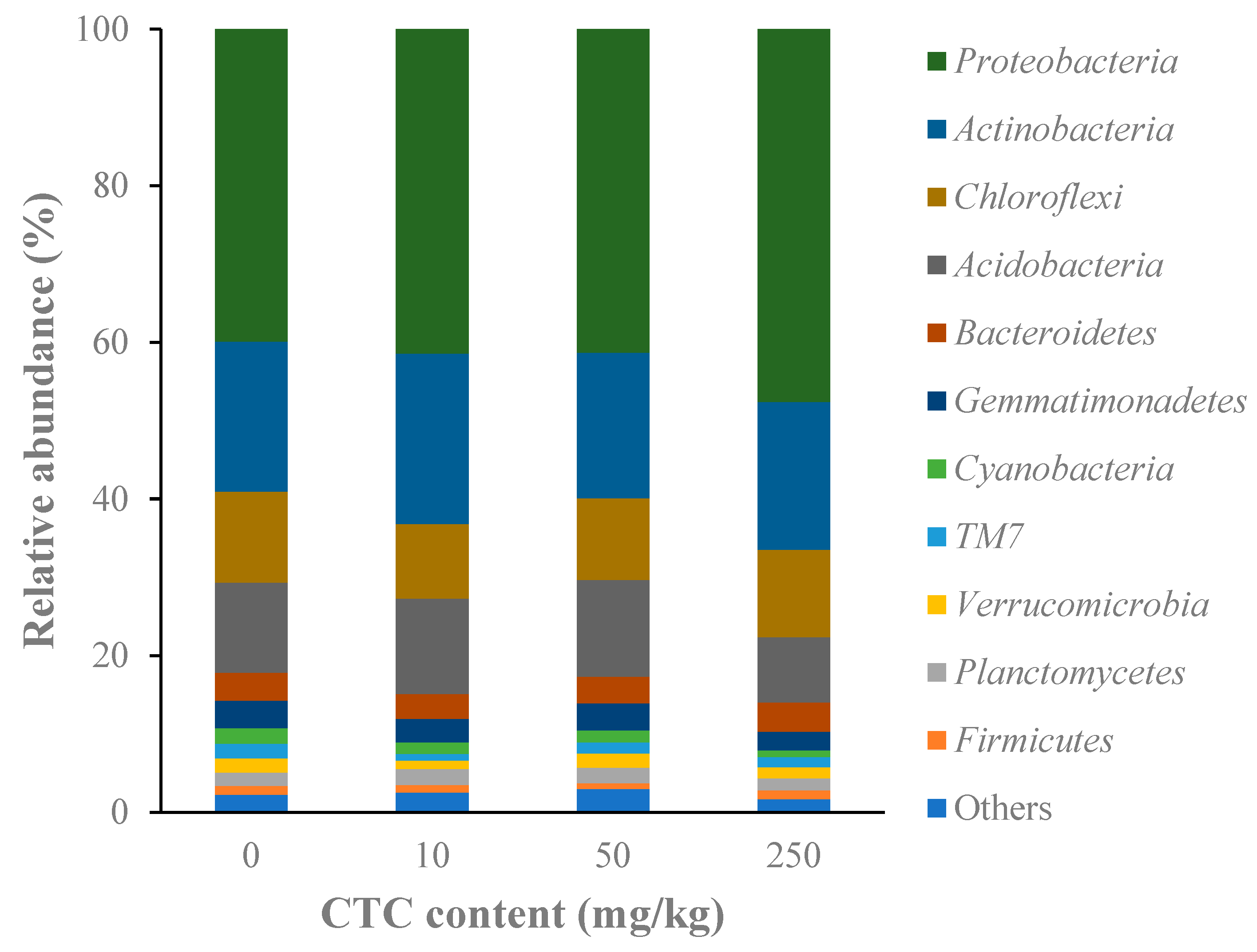
In all soil samples, the microbial composition revealed high variety. The microorganisms in all soil samples belonged to 48 phyla. Proteobacteria (39.9–47.6%) and Actinobacteria (18.6–21.8%) made up more than half of all bacterial phyla, making them the dominant phyla (Figure 4). Although the abundance of the detected phyla between the control and CTC-treated groups did not differ significantly according to the ANOVA test, microbes had a distinct pattern of change. Results demonstrated that all CTC treatments increased the relative abundance of Proteobacteria, but reduced the abundance of Gemmatimonadetes and Cyanobacteria with a dose-dependent relationship. In 250 mg/kg group, the relative abundance of Bacteroidetes was increased.
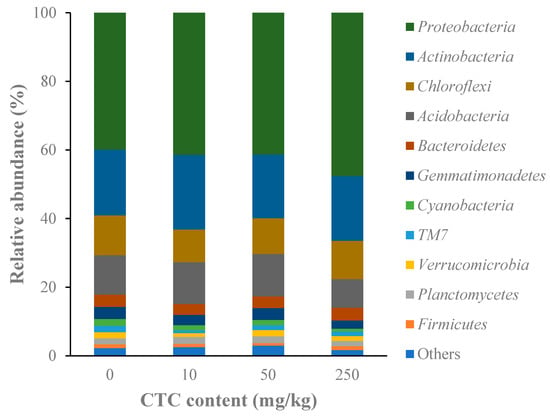
Figure 4.
The relative abundances of microorganisms at the phylum level at different CTC treatments.
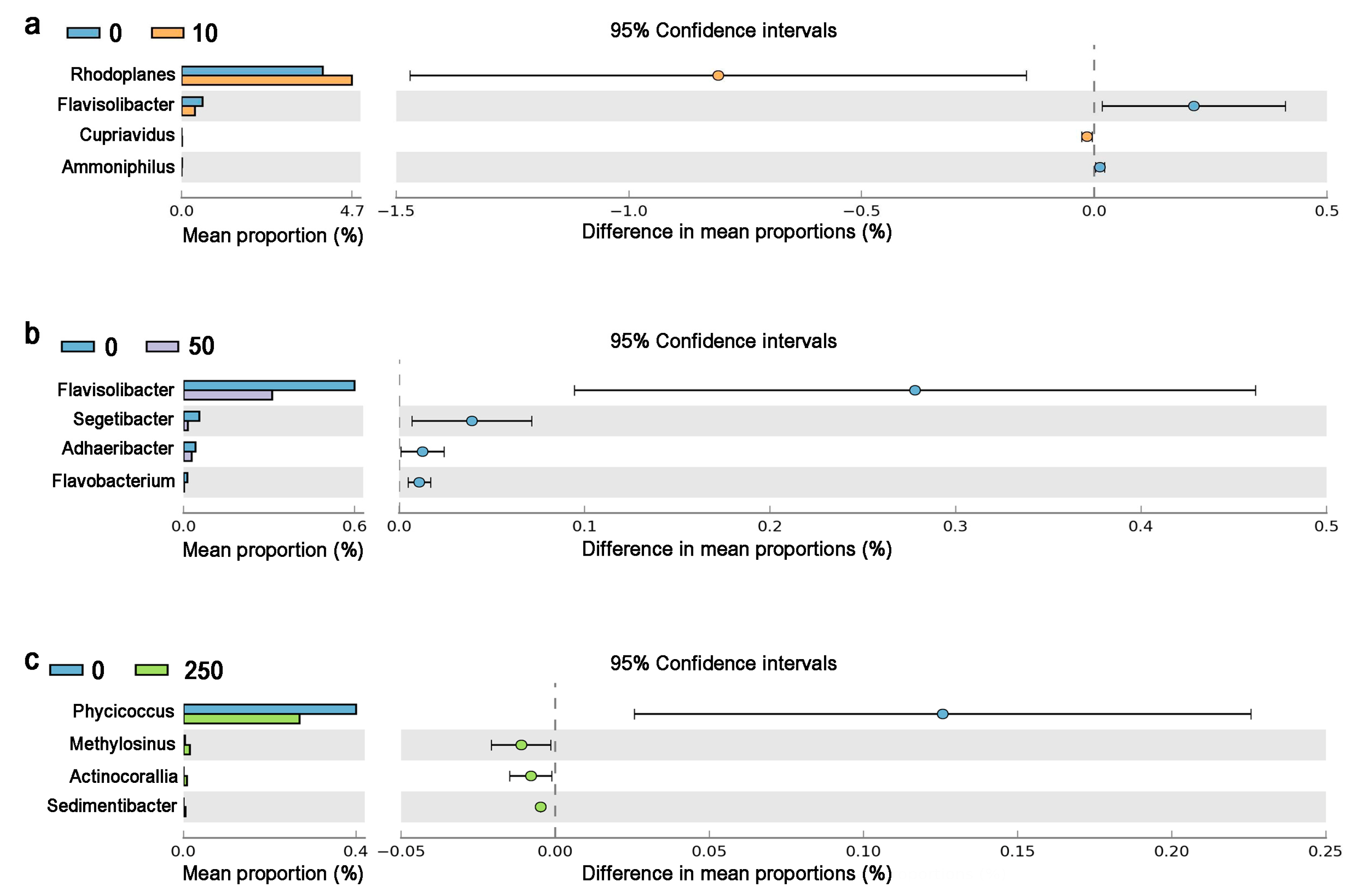
The chronic CTC exposure significantly (p < 0.05) altered the relative abundance of 11 genera (Figure 5). Rhodoplanes and Cupriavidus were enriched in 10 mg/kg CTC group indicating they may prefer low-dose CTC. Methylosinus, Actinocorallia, and Sedimentibacter were enriched in 250 mg/kg CTC group, indicating that high-dose CTC stimulated the growth of these genera. Compared with the control, the relative abundances of Flavisolibacter and Ammoniphilus were significantly reduced by 10 mg/kg CTC. The relative abundances of Flavisolibacter, Segetibacter, Adhaeribacter, and Flavobacterium were significantly reduced by 50 mg/kg CTC. A total of 250 mg/kg CTC significantly inhibited the growth of Phycicoccus.
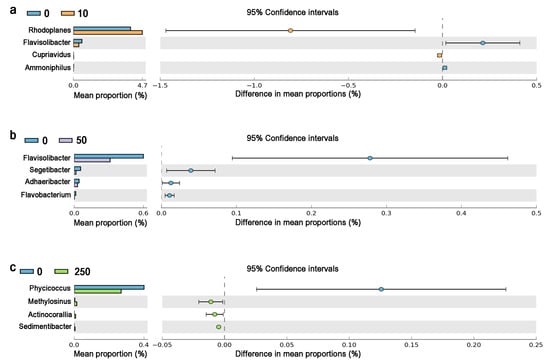
Figure 5.
Relative abundance of significantly changed soil microbes at genus in CTC groups compared with the control. Notes: (a) represented the different genus between 10 mg/kg CTC and the control, (b) represented the different genus between 50 mg/kg CTC and the control, and (c) represented the different genus between 250 mg/kg CTC and the control.
3.4. Correlation between Each Indexes
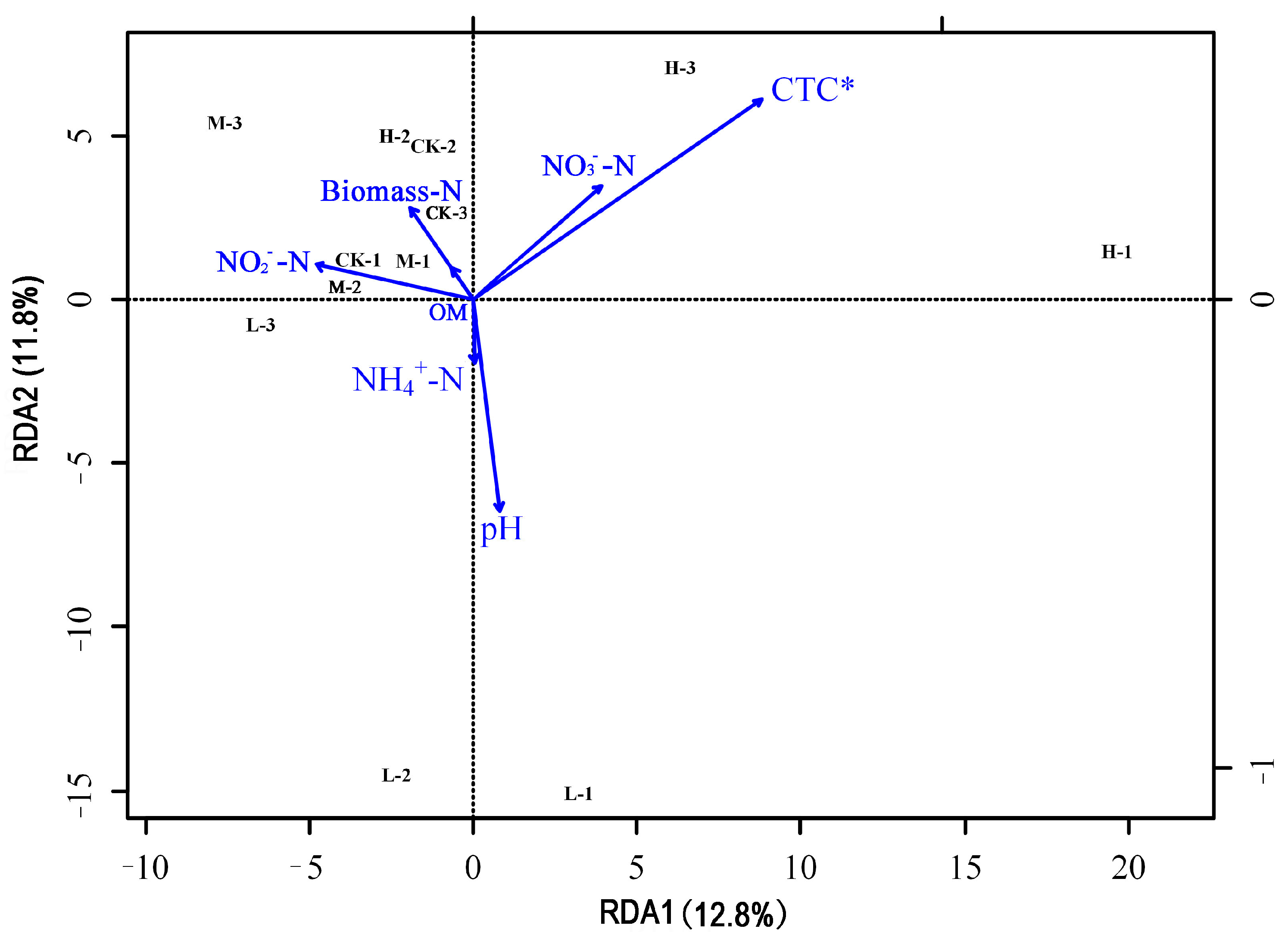
Figure 6 shows the results of redundancy analysis (RDA) between the microbial communities and soil environment factors for the groups. Soil environmental factors had an overall explanation of 81.8% for the microorganism’s community’s differences. RDA1 and RDA2 had an overall explanation of 24.6%, with RDA 1 and RDA2 contributing 12.8% and 11.8%, respectively. As shown in Figure 6, CTC significantly influenced soil microbial community. Soil pH, NO3−–N and NO2−–N had strong correlation with the soil microbial community.
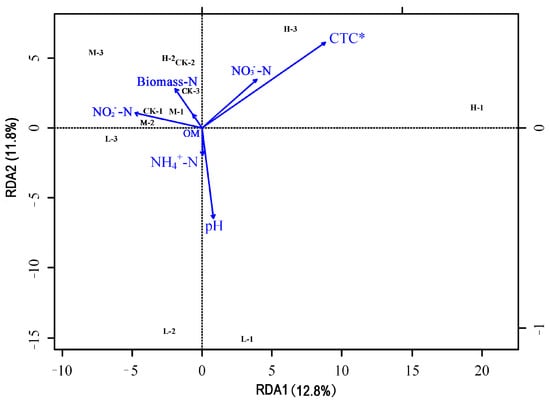
Figure 6.
Redundancy analysis of soil microbial community and soil environmental factors. * represented CTC significantly influenced soil microbial community (p < 0.05).
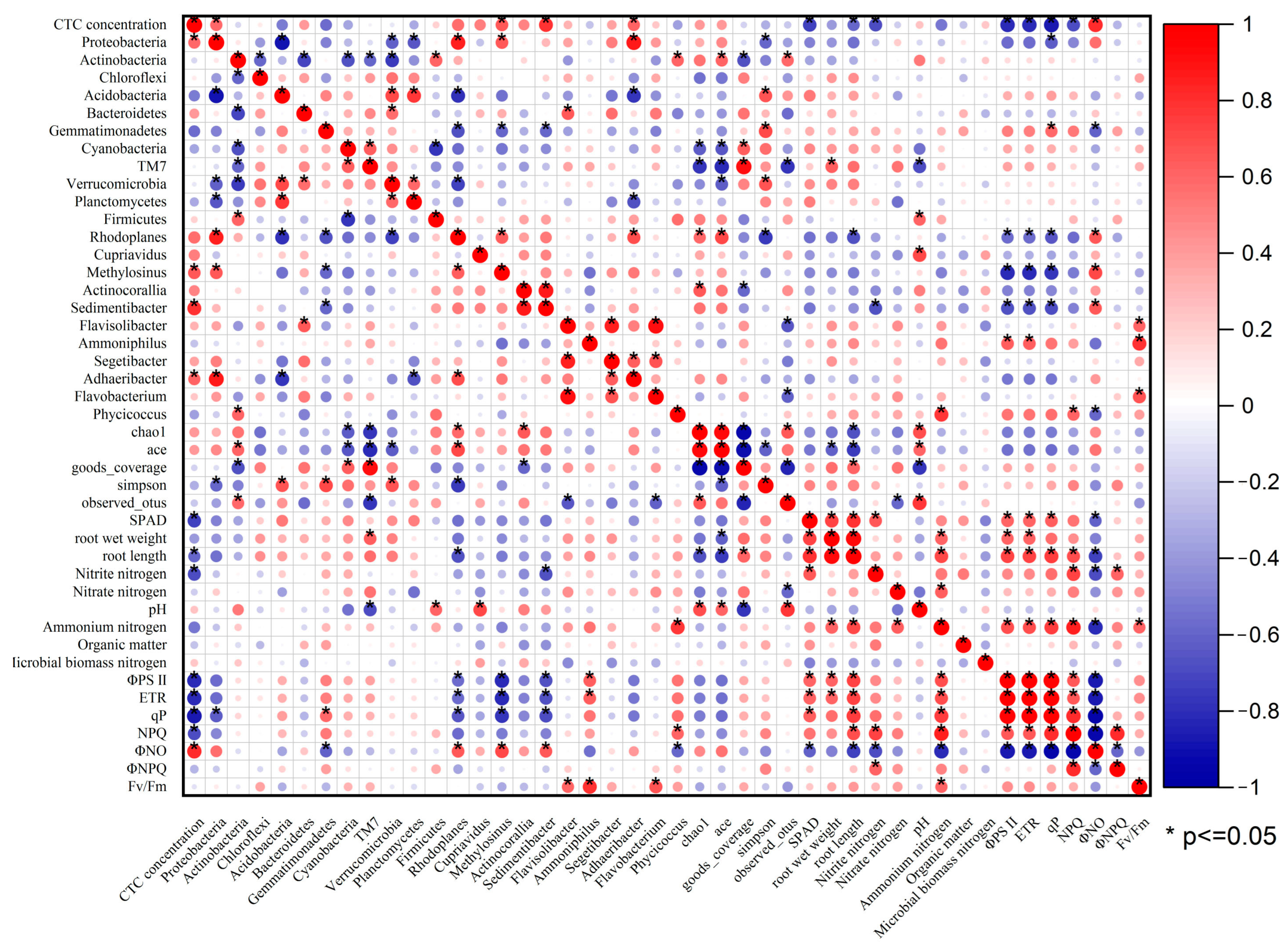
To examine the interrelationships among different indexes, the correlation analysis (Pearson) was carried out in this study (Figure 7). The red and blue circles represented the positive and negative correlation between the two related indexes, respectively. And the size of the circles represented the absolute value of the correlation coefficient. As shown in Figure 7, there was some degree of correlation between CTC, soil characteristics, plant growth, fluorescence parameters, and soil bacteria, demonstrating that these variables were closely correlated. Root length had significant negative correlation with ΦNO, positive correlation with ETR, qP, NPQ, ΦPSII and NH4+–N. In this study, Rhodoplanes demonstrated a positive correlation with Methylosinus and Adhaeribacter. Actinocorallia and Sedimentibacter exhibited a positive correlation. Flavobacterium exhibited a positive correlation with Flavisolibacter and Segetibacter.
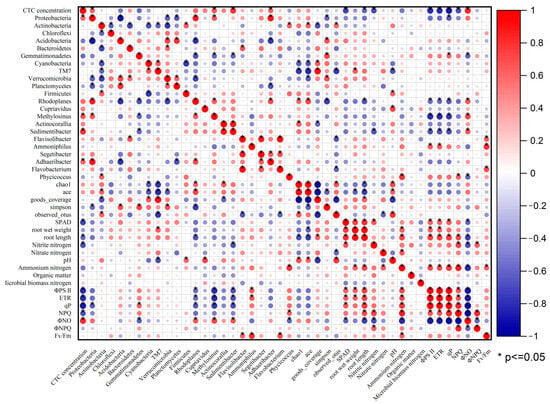
Figure 7.
Heat map of Pearson correlation between all indexes in this experiment.
4. Discussion
Previous studies have demonstrated that NH4+–N and NO3−–N were the main forms of nitrogen absorbed and utilized by plants, and their content can influence plant growth and effectively indicate the nitrogen supply in soil [40]. In this study, the significant reduction in NH4+–N and NO2−–N by CTC may be related to the decline of azotification and nitrite bacteria activity. As a measure of soil nitrification capability, NO3−–N concentration is much more sensitive to the presence of pharmaceuticals [41]. And due to high sensitivity of NO3−–N, all CTC groups showed lower nitrate nitrogen NO3−–N levels. The inhibitory effects of CTC on soil nitrification capacity, which may be brought on by the inhibitory effects on nitrobacteria, are similar to previous studies on effects of pharmaceutical antibiotics on nitrification capacity [42,43,44].
Leaf SPAD observations which show the index of chlorophyll a and chlorophyll b in thylakoid membrane in the leaf mesophyll chloroplasts are collinearly correlated with leaf chlorophyll content for many crops [45]. CTC may be involved in the breakdown of chlorophyll by interacting with Mg2+ in the chlorophyll molecule and thus reduced chlorophyll content [46,47,48]. In this study, the root fresh weight, root length, and SPAD value in the CTC treatment groups all demonstrated a concentration-dependent decrease. This phenomenon of growth inhibition was supported by previous research on the impacts of CTC on the growth of maize and Brassica campestris [49,50]. According to the results in Table 1 and Figure 1, the inhibition of eggplant growth at CTC groups was thought to be related to the disturbance of chlorophyll fluorescence parameters, NH4+–N and NO3−–N.
The maximal photochemical efficiency of PS II (Fv/Fm) was used to measure the efficiency of the primary conversion of light energy at the PS II reaction center [19,51,52]. In this study, the decrease in Fv/Fm in all CTC groups indicated that the eggplant suffered from severe stress of CTC. The photochemical quenching coefficient (qP) reflects the proportion of the light energy absorbed by the PS II antenna chlorophyll that is used in photochemical electron transfer, which can reflect the openness of PS II, and represents the electron transfer activity of PS II [19,53]. The qP value was significantly decreased by CTC, indicating diminished electron transfer activity at PS II by CTC. NPQ reflects the utilization of excitation energy in leaves and represents heat dissipation that defends against the destruction of photo inhibition, and is an important mechanism for plant protection of PS II [54]. The NPQ values rapidly declined with increasing CTC content, indicating heat dissipation performance of eggplant decreased. Both ETR and ΦPS II serve as relative indicators of the electron transfer rate of plant leaves during photosynthesis and can reflect the actual photochemical reaction efficiency of plants in PS II [55]. In this study, ETR and ΦPS II were dramatically reduced by CTC treatments with a dose–effect relationship, indicating that CTC exposure inhibited electron transport during photosynthesis. ΦNPQ and ΦNO are indexes of light protection and light damage, respectively. The values of ΦNPQ were decreased but without significant difference, while the ΦNO values increased significantly. The increase in ΦNO indicated that there was a tendency of excess excitation energy to cause potential damage in CTC groups, and the photochemical energy conversion and protective regulated energy dissipation are not enough to coordinate the light energy absorption of photosynthetic apparatus [56,57]. The aforementioned findings demonstrated that with prolonged exposure to CTC, the photosynthetic system suffered serious damage, heat dissipation performance deteriorated, and reaction centers entered the closed state.
The community richness in this study was somewhat boosted by CTC, but there were no appreciable differences, and CTC had no impact on the community diversity. This phenomenon was also discovered in previous studies, which demonstrated that TCs had no significant effect or promotion effect on soil bacterial diversity, but had significant effect on different species at phyla and genera level [12,58]. In this study, all CTC groups increased the relative abundance of Proteobacteria, but reduced the abundance of Gemmatimonadetes and Cyanobacteria, and the highest CTC group increased the relative abundance of Bacteroidetes. The multiplication of Proteobacteria and Bacteroidetes suggested that they can tolerate the high dose of CTC. Similarly, Zhao et al. [59] indicated that the relative abundance of Bacteroidetes increased with the concentrations of antibiotic residues increased in typical greenhouse vegetable soils. Guo et al. [2] also revealed the reduction in Gemmatimonadetes after 150 mg/kg oxytetracycline treatment. Cyanobacteria, a sizable group of photosynthetic prokaryotes, play a significant role in the global CO2 and N2 fixation process [60]. Based on the results of previous studies, the reduction in Cyanobacteria would result in the reduction of nitrogen fixation and then the reduction in ammonium nitrogen content in this study.
According to a t-test examination of the microorganism differences between the control and CTC groups, Rhodoplanes and Cupriavidus were enriched in low-dose CTC group, and Methylosinus was enriched in the highest CTC group. Rhodoplanes, one of denitrifiers in soil, was affiliated with N cycling and involved in denitrification process [61,62]. Cupriavidus is highly resistant to heavy metals and can degrade toxic organic pollutants in soil to be benefit for plant growth [63,64]. Previous studies have found that Cupriavidus has the ability of heterotrophic nitrification and aerobic denitrification to remove ammonium, and nitrite [61]. These findings can explain the results that NH4+–N and NO3−–N were reduced at 10 mg/kg CTC group in this study. Methylosinus is a type II methane-oxidizing bacteria that reduces methane emissions, and participates in nitrogen oxide metabolism, and benefits the balance between nitrogen and methane cycling [65,66]. Flavisolibacter is one of plant growth promoting rhizobacteria, and its relative abundance was found to be negatively correlated with some stress, such as disease incidence [67]. The relative abundances of Flavisolibacter in this study were significantly reduced by 50 mg/kg CTC, explaining the growth inhibition in this group. Flavobacterium is not only a kind of pathogenic bacteria, but also an important aerobic denitrifying bacteria, and has been proved to be related to NO2−–N and NO3−–N [68,69]. In this study, high-dose CTC reduced the Flavobacterium indicated that CTC could significantly inhibit the growth of this bacteria.
RDA analysis showed soil pH, NO3−–N, and NO2−–N had strong correlation with the soil microbial community. Soil pH and nutrients are closely related to the functional diversity of microbial community, since a large number of previous studies have proved that soil pH, organic carbon, and other nutrients have different degrees of influence on soil microorganisms [70,71]. Conversely, the change in soil microorganisms leads to the change in soil physical and chemical properties. Due to the complexity of the microbial community structure, it is unavoidable and reasonable for microbes to form a mutually facilitative or inhibitory relationship which is typically involves competition for resources and energy to help to determine the presence and relative abundance of species to some extent [72]. In this study, Rhodoplanes demonstrated a positive correlation with Methylosinus and Adhaeribacter. Actinocorallia and Sedimentibacter exhibited a positive correlation. Flavobacterium exhibited a positive correlation with Flavisolibacter and Segetibacter. These microbes play important roles in soil nutrient cycling and plant growth, and their positive correlation may be beneficial to improve the adaptability of plants under exogenous stress [62,66,67,68,69]. A heat map of the Pearson correlation demonstrated that soil pollutants, soil microorganisms, soil properties, and plant interact with and affect each other. However, due to the limited number of samples in our study, any correlation between these environmental factors and bacterial biodiversity requires further examination.
5. Conclusions
In this study, CTC treatment changed the soil properties and inhibited the growth of eggplant. The primary explanation may be the disruption of chlorophyll fluorescence parameters by CTC, particularly the light damage it causes. Additionally, varying doses of CTC disrupted the bacterial communities in the soil, particularly the rhizobacteria that promote plant growth and the bacterial communities that aid in denitrification. The mechanisms of CTC toxicity to eggplant should focus more on the metabolism of soil microbes and plant secretions. RDA showed CTC significantly influenced soil microbial community, and soil properties are closely related to the soil microbial community. Pearson correlation analysis demonstrated that CTC, soil microbes, soil nutrients, and the plant can interact with and affect each other. These results highlight the need for a thorough assessment of the dangers associated with tetracycline antibiotics and help us understand how they affect terrestrial ecosystems, to promote sustainable agricultural development. However, this study did not focus on the effects of CTC on the expression of plant-related resistance genes, and low-dose CTC exposure risk should be investigated in the future.
Author Contributions
Conceptualization, Y.C. and L.L.; methodology, Y.C., L.L. and Y.X.; software, H.W.; validation, Y.C. and L.L.; formal analysis, L.L.; investigation, Y.C.; resources, Y.X.; data curation, Y.C. and L.L.; writing—original draft preparation, L.L.; writing—review and editing, Y.C. and L.L.; visualization, L.L. and H.W.; supervision, Y.C.; project administration, Y.C. and L.L.; funding acquisition, Y.C. and L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Anhui University Outstanding Top-of-the-line Talent Cultivation Project, grant number [NO. gxbjZD2021073], the Natural Science Foundation of Anhui Province, China, grant number [NO. 2008085QC103] and Hefei Normal University High-level Talents Research Start-up Fund, grant number [NO. 2020rcjj49].
Institutional Review Board Statement
The study did not require ethical approval.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks to the suggestion of “State Key Laboratory of Pollution Control and Resource Reuse, School of the Environment, Nanjing University” during the whole experiment and manuscript design. Thanks to the sequencing analysis of JiangSu ZhongYiJinDa Analytical & Testing Co., Ltd.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Daghrir, R.; Drogui, P. Tetracycline antibiotics in the environment: A review. Environ. Chem. Lett. 2013, 11, 209–227. [Google Scholar] [CrossRef]
- Guo, A.; Pan, C.; Su, X.; Zhou, X.; Bao, Y. Combined effects of oxytetracycline and microplastic on wheat seedling growth and associated rhizosphere bacterial communities and soil metabolite profiles. Environ. Pollut. 2022, 302, 119046. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Yang, L.; Zhang, L.; Ye, B.; Wang, L. Antibiotics in soil and water in China-a systematic review and source analysis. Environ. Pollut. 2020, 266, 115147. [Google Scholar] [CrossRef] [PubMed]
- Patyra, E.; Kowalczyk, E.; Grelik, A.; Przeniosło-Siwczyńska, M.; Kwiatek, K. Screening method for the determination of tetracyclines and fluoroquinolones in animal drinking water by liquid chromatography with diode array detector. Pol. J. Vet. Sci. 2015, 18, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Chen, L.; Yang, L.; Li, S.; Sun, L.; Yu, X. Distribution, dynamics and determinants of antibiotics in soils in a peri-urban area of Yangtze River Delta, Eastern China. Chemosphere 2018, 211, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; He, T.; Zhang, S.; Zhu, L.; Shang, B.; Li, Z.; Wang, R. Occurrence of seventeen veterinary antibiotics and resistant bacterias in manure-fertilized vegetable farm soil in four provinces of China. Chemosphere 2019, 215, 234–240. [Google Scholar] [CrossRef]
- Pan, Z.; Yang, S.; Zhao, L.; Li, X.; Weng, L.; Sun, Y.; Li, Y. Temporal and spatial variability of antibiotics in agricultural soils from Huang-Huai-Hai Plain, northern China. Chemosphere 2021, 272, 129803. [Google Scholar] [CrossRef]
- Hamscher, G.; Sczesny, S.; Höper, H.; Nau, H. Determination of persistent tetracycline residues in soil fertilized with liquid manure by high-performance liquid chromatography with electrospray ionization tandem mass spectrometry. Anal. Chem. 2002, 74, 1509–1518. [Google Scholar] [CrossRef]
- Lee, C.; An, J.; Lee, Y.S.; Choi, K.; Kim, J.Y. Uncertainty-based concentration estimation of chlortetracycline antibiotics in swine farms and risk probability assessment for agricultural application of manure. J. Hazard. Mater. 2021, 402, 123763. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Ma, J.; Zhu, D.; Sheng, G.D.; O’Connor, P.; Zhu, Y.G. Soil oxytetracycline exposure alters the microbial community and enhances the abundance of antibiotic resistance genes in the gut of Enchytraeus crypticus. Sci. Total Environ. 2019, 673, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.T.; Pan, X.; Chen, L.K.; Liu, W.X.; Christie, P.; Luo, Y.M.; Wu, L.H. Effects of different concentrations and application frequencies of oxytetracycline on soil enzyme activities and microbial community diversity. Eur. J. Soil Biol. 2016, 76, 53–60. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Grinyer, J.; Reich, P.B.; Singh, B.K. Relative importance of soil properties and microbial community for soil functionality: Insights from a microbial swap experiment. Funct. Ecol. 2016, 30, 1862–1873. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]
- Chessa, L.; Pusino, A.; Garau, G.; Mangia, N.P.; Pinna, M.V. Soil microbial response to tetracycline in two different soils amended with cow manure. Environ. Sci. Pollut. Res. Int. 2016, 23, 5807–5817. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, W.; Xu, W. Effects of tetracycline antibiotics in chicken manure on soil microbes and antibiotic resistance genes (ARGs). Environ. Geochem. Health 2022, 44, 273–284. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, J.; Gao, L.; Kong, F.; Shen, G.; Wang, R.; Gao, J.; Zhang, J. The Effects of Tetracycline Residues on the Microbial Community Structure of Tobacco Soil in Pot Experiment. Sci. Rep. 2020, 10, 8804. [Google Scholar] [CrossRef]
- Han, L.; Zhang, H.; Long, Z.; Ge, Q.; Mei, J.; Yu, Y.; Fang, H. Exploring microbial community structure and biological function in manured soil during ten repeated treatments with chlortetracycline and ciprofloxacin. Chemosphere 2019, 228, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lv, Y.; Xu, K.; Xiao, X.; Xi, B.; Lu, S. Response of ginger growth to a tetracycline-contaminated environment and residues of antibiotic and antibiotic resistance genes. Chemosphere 2018, 201, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Liu, X.; Wang, J.; Li, S. Accumulation and risk assessment of antibiotics in edible plants grown in contaminated farmlands: A review. Sci. Total Environ. 2022, 853, 158616. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Chen, L.; Wu, L.; Christie, P.; Luo, Y. Toxicity of OTC to Ipomoea aquatica Forsk. and to microorganisms in a long-term sewage-irrigated farmland soil. Environ. Sci. Pollut. Res. Int. 2016, 23, 15101–15110. [Google Scholar] [CrossRef]
- Ding, D.; Huang, X.Y.; Gu, J.Y.; Chen, C.Y.; Long, X.X.; Zeng, Q.Y. Distribution Characteristics and Risk Assessment of Tetracycline Antibiotics (TCs) in Soil-Vegetable System with Soil Fertilized with Animal Manure. Environ. Sci. 2023, 44, 4440–4447. (In Chinese) [Google Scholar]
- Xiang, L.; Wu, X.L.; Jiang, Y.N.; Yan, Q.Y.; Li, Y.W.; Huang, X.P.; Cai, Q.Y.; Mo, C.H. Occurrence and risk assessment of tetracycline antibiotics in soil from organic vegetable farms in a subtropical city, south China. Environ. Sci. Pollut. Res. Int. 2016, 23, 13984–13995. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Chu, L.M. Phytotoxicity of veterinary antibiotics to seed germination and root elongation of crops. Ecotoxicol. Environ. Saf. 2016, 126, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, J.; Lu, C.; Liao, Q.; Gudda, F.O.; Ling, W. Antibiotics in animal manure and manure-based fertilizers: Occurrence and ecological risk assessment. Chemosphere 2020, 255, 127006. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Guo, Y.; Shi, M.; Qiu, T.; Gao, M.; Tian, S.; Wang, X. Effect of antibiotic type and vegetable species on antibiotic accumulation in soil-vegetable system, soil microbiota, and resistance genes. Chemosphere 2021, 263, 128099. [Google Scholar] [CrossRef]
- Wang, W.Z.; Chi, S.L.; Xu, W.H. Biological Effect of Tetracycline Antibiotics on a Soil-Lettuce System and Its Migration Degradation Characteristics. Environ. Sci. 2021, 42, 1545–1558. (In Chinese) [Google Scholar]
- Cheong, M.S.; Choe, H.; Jeong, M.S.; Yoon, Y.E.; Jung, H.S.; Lee, Y.B. Different Inhibitory Effects of Erythromycin and Chlortetracycline on Early Growth of Brassica campestris Seedlings. Antibiotics 2021, 10, 1273. [Google Scholar] [CrossRef]
- Grayston, S.J.; Wang, S.Q.; Campbell, C.D.; Anthony, C.E. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biol. Biochem. 1998, 30, 369–378. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Gürbüz, N.; Uluişik, S.; Frary, A.; Frary, A.; Doğanlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.; Barber, S.A. Effect of ammonium and nitrate fertilization on phosphorus uptake as related to root-Induced pH changes at the root-soil interface. Soil Sci. Soc. Amer. Proc. 1971, 35, 301–306. [Google Scholar] [CrossRef]
- Liu, F.; Mo, X.; Kong, W.; Song, Y. Soil bacterial diversity, structure, and function of Suaeda salsa in rhizosphere and non-rhizosphere soils in various habitats in the Yellow River Delta, China. Sci. Total Environ. 2020, 740, 140144. [Google Scholar] [CrossRef]
- NY/T 1377-2007; Determination of pH in Soil. Ministry of Agriculture of the People’s Republic of China. Standards Press of China: Beijing, China, 2007.
- NY/T 1121.6-2006; Soil Testing Part 6: Method for Determination of Soil Organic Matter. Ministry of Agriculture of the People’s Republic of China. Standards Press of China: Beijing, China, 2006.
- HJ 634-2012; Soil-Determination of Ammonium, Nitrite and Nitrate by Extraction with Potassium Chloride Solution-Spectrophotometric Methods. Ministry of Environmental Protection of the People’s Republic of China. China Environmental Press: Beijing, China, 2012.
- Wu, J.S.; Lin, Q.M.; Huang, Q.Y.; Xiao, H.A. Method for Calculating Soil Microbial Biomass and Its Application; China Meteorological Press: Beijing, China, 2006; pp. 54–71. [Google Scholar]
- Xue, Y.G.; Liu, F.; Zhou, L.L.; Jin, S.; Jiang, Y.; Wang, Y.C.; Jiang, X.D.; Wang, Q.; Shi, X.L.; Xue, K. Comparison study of bacterial community structure between groundwater and soil in industrial park based on high throughput sequencing. Asian J. Ecotoxicol. 2017, 12, 107–115. (In Chinese) [Google Scholar]
- Mao, B.; Zeng, Y.; Lai, C.T.; Yang, Y.; Zhou, C.H.; Li, Z.T. Effects of nitrogen reduction fertilization on sugarcane biomass, soil nitrate nitrogen and ammonium nitrogen. Chin. J. Ecol. 2023. Available online: https://kns.cnki.net/kcms/detail/21.1148.q.20230303.1019.006.html (accessed on 6 March 2023). (In Chinese).
- Pashaei, R.; Zahedipour-Sheshglani, P.; Dzingelevičienė, R.; Abbasi, S.; Rees, R.M. Effects of pharmaceuticals on the nitrogen cycle in water and soil: A review. Environ. Monit. Assess. 2022, 194, 105. [Google Scholar] [CrossRef] [PubMed]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed]
- Kotzerke, A.; Sharma, S.; Schauss, K.; Heuer, H.; Thiele-Bruhn, S.; Smalla, K.; Wilke, B.M.; Schloter, M. Alterations in soil microbial activity and N-transformation processes due to sulfadiazine loads in pig-manure. Environ. Pollut. 2008, 153, 315–322. [Google Scholar] [CrossRef]
- Toth, J.D.; Feng, Y.C.; Dou, Z.X. Veterinary antibiotics at environmentally relevant concentrations inhibit soil iron reduction and nitrification. Soil Biol. Biochem. 2011, 43, 2470–2472. [Google Scholar] [CrossRef]
- Yadava, U.L. A rapid and nondestructive method to determine chlorophyll in intact leaves. Hortscience 1986, 21, 1449–1500. [Google Scholar] [CrossRef]
- Park, S.Y.; Yu, J.W.; Park, J.S.; Li, J.; Yoo, S.C.; Lee, N.Y.; Lee, S.K.; Jeong, S.W.; Seo, H.S.; Koh, H.J.; et al. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell 2007, 19, 1649–1664. [Google Scholar] [CrossRef]
- Shimoda, Y.; Ito, H.; Tanaka, A. Arabidopsis STAY-GREEN, Mendel’s Green Cotyledon Gene, Encodes Magnesium-Dechelatase. Plant Cell 2016, 28, 2147–2160. [Google Scholar] [CrossRef] [PubMed]
- Rydzyński, D.; Piotrowicz-Cieślak, A.I.; Grajek, H.; Wasilewski, J. Investigation of chlorophyll degradation by tetracycline. Chemosphere 2019, 229, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Liu, Y.; Wang, P.; Wu, T.; Zhang, S.; Shan, X.; Lu, J. Toxic effects of chlortetracycline on maize growth, reactive oxygen species generation and the antioxidant response. J. Environ. Sci. 2012, 24, 1099–1105. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- Cheong, M.S.; Yoon, Y.E.; Kim, J.W.; Hong, Y.K.; Kim, S.C.; Lee, Y.B. Chlortetracycline inhibits seed germination and seedling growth in Brassica campestris by disrupting H2O2 signaling. Appl. Biol. Chem. 2020, 63, 1. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Jägerbrand, A.K.; Kudo, G. Short-Term Responses in Maximum Quantum Yield of PSII (Fv/Fm) to ex situ Temperature Treatment of Populations of Bryophytes Originating from Different Sites in Hokkaido, Northern Japan. Plants 2016, 5, 22. [Google Scholar] [CrossRef]
- Zhang, S.R. A discussion on Chlorophyll Fluorescence kinetics parameters and their significance. Chin. Bull. Bot. 1999, 17, 444–448. [Google Scholar]
- Demmig-Adams, B.; Adams, W.W.I. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Du, P.Z.; Liao, S.B.; Sun, B.; Chen, Y.; Luo, S.X.; Chen, L.; Huang, Y.F. Photosynthetic pigments and chlorophyll fluorescent characteristics of 17 provenances of Banksia seedlings. J. Central South Univ. Forest. Technol. 2014, 34, 49–54. (In Chinese) [Google Scholar]
- Costa, E.S.; Bressan-Smith, R.; Oliveira, J.G.; Campostrini, E. Chlorophyll a fluorescence analysis in response to excitation irradiance in bean plants (Phaseolus vulgaris L. and unguiculata L. Walp) submitted to high temperature stress. Photosynthetica 2003, 41, 77–82. [Google Scholar] [CrossRef]
- Shi, S.B.; Zhou, D.W.; Li, T.C.; De, K.J.; Gao, X.Z.; Ma, J.L.; Sun, T.; Wang, F.L. Responses of photosynthetic function of Kobresia pygmaea to simulated nocturnal low temperature on the Qingzang Plateau. Chin. J. Plant Ecol. 2023, 47, 361–373. (In Chinese) [Google Scholar] [CrossRef]
- Liu, M.; Cao, J.; Wang, C. Bioremediation by earthworms on soil microbial diversity and partial nitrification processes in oxytetracycline-contaminated soil. Ecotoxicol. Environ. Saf. 2020, 189, 109996. [Google Scholar] [CrossRef]
- Zhao, L.; Pan, Z.; Sun, B.; Sun, Y.; Weng, L.; Li, X.; Ye, H.; Ye, J.; Pan, X.; Zhou, B.; et al. Responses of soil microbial communities to concentration gradients of antibiotic residues in typical greenhouse vegetable soils. Sci. Total Environ. 2023, 855, 158587. [Google Scholar] [CrossRef]
- Adams, D.G.; Duggan, P.S. Cyanobacteria-bryophyte symbioses. J. Exp. Bot. 2008, 59, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Liu, F.; Xu, S.; Wu, S.; Zhuang, G.; Deng, Y.; Wu, J.; Zhuang, X. Myriophyllum aquaticum Constructed Wetland Effectively Removes Nitrogen in Swine Wastewater. Front. Microbiol. 2017, 8, 1932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chang, L.; Liu, X.; Wang, J.; Ge, X.; Cheng, J.; Xie, J.; Lin, Y.; Fu, Y.; Jiang, D.; et al. Rapeseed Domestication Affects the Diversity of Rhizosphere Microbiota. Microorganisms 2023, 11, 724. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, T.A.; Daghio, M.; González, M.; Papacchini, M.; Franzetti, A.; Seeger, M. Toluene degradation by Cupriavidus metallidurans CH34 in nitrate-reducing conditions and in Bioelectrochemical Systems. FEMS Microbiol. Lett. 2018, 365, 4996784. [Google Scholar]
- Bravo, G.; Vega-Celedón, P.; Gentina, J.C.; Seeger, M. Bioremediation by Cupriavidus metallidurans Strain MSR33 of Mercury-Polluted Agricultural Soil in a Rotary Drum Bioreactor and Its Effects on Nitrogen Cycle Microorganisms. Microorganisms 2020, 8, 1952. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.Y.; Klotz, M.G. Nitrifying and denitrifying pathways of methanotrophic bacteria. Biochem. Soc. Trans. 2011, 39, 1826–1831. [Google Scholar] [CrossRef]
- Shinjo, R.; Oe, F.; Nakagawa, K.; Murase, J.; Asakawa, S.; Watanabe, T. Type-specific quantification of particulate methane monooxygenase gene of methane-oxidizing bacteria at the oxic-anoxic interface of a surface paddy soil by digital PCR. Environ. Microbiol. Rep. 2023, 15, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Qiu, T.; Zhu, F.Y.; Xiao, J.L.; Wei, L.; Liang, Z.H. Calcium oxide regulation of fusarium wilt and rhizosphere bacterial community. J. Hunan Agric. Univ. (Nat. Sci.) 2018, 44, 620–624. (In Chinese) [Google Scholar]
- Zhang, T.; Shao, M.F.; Ye, L. 454 pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J. 2012, 6, 1137–1147. [Google Scholar] [CrossRef]
- Zhu, J.S.; Qin, H.L.; Sun, Q.Y.; Wang, B.Z.; Gao, R.X.; Guo, R.L.; Li, W.B. Microbial Diversity and Influencing Factors in a Small Watershed in winter. Environ. Sci. 2020, 41, 5016–5026. (In Chinese) [Google Scholar]
- Li, H.Y.; Yao, T.; Zhang, J.G.; Gao, Y.M.; Ma, Y.C.; Lu, Y.W.; Zhang, H.R.; Yang, X.L. Relationship between soil bacterial community and environmental factors in the degraded alpine grassland of eastern Qilian Mountains, China. Chin. J. Appl. Ecol. 2018, 29, 3793–3801. (In Chinese) [Google Scholar]
- Guo, H.; Tang, W.P. Enzyme activity and microbial community diversity in rhizosphere and non-rhizosphere of Larix principis-rupprechtii. Ecol. Environ. Sci. 2020, 29, 2163–2170. (In Chinese) [Google Scholar]
- Gilbert, J.A.; Steele, J.A.; Caporaso, J.G.; Steinbrück, L.; Reeder, J.; Temperton, B.; Huse, S.; McHardy, A.C.; Knight, R.; Joint, I.; et al. Defining seasonal marine microbial community dynamics. ISME J. 2012, 6, 298–308. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).