


The Effect of Clearing Diseased Wood on the Soil’s Physicochemical Properties in Black Pine Forests


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Manipulation and Design
2.2.1. Sample Design
2.2.2. Meteorological Observation
2.2.3. The Soil’s Physicochemical Properties
2.3. Data Analysis
3. Results
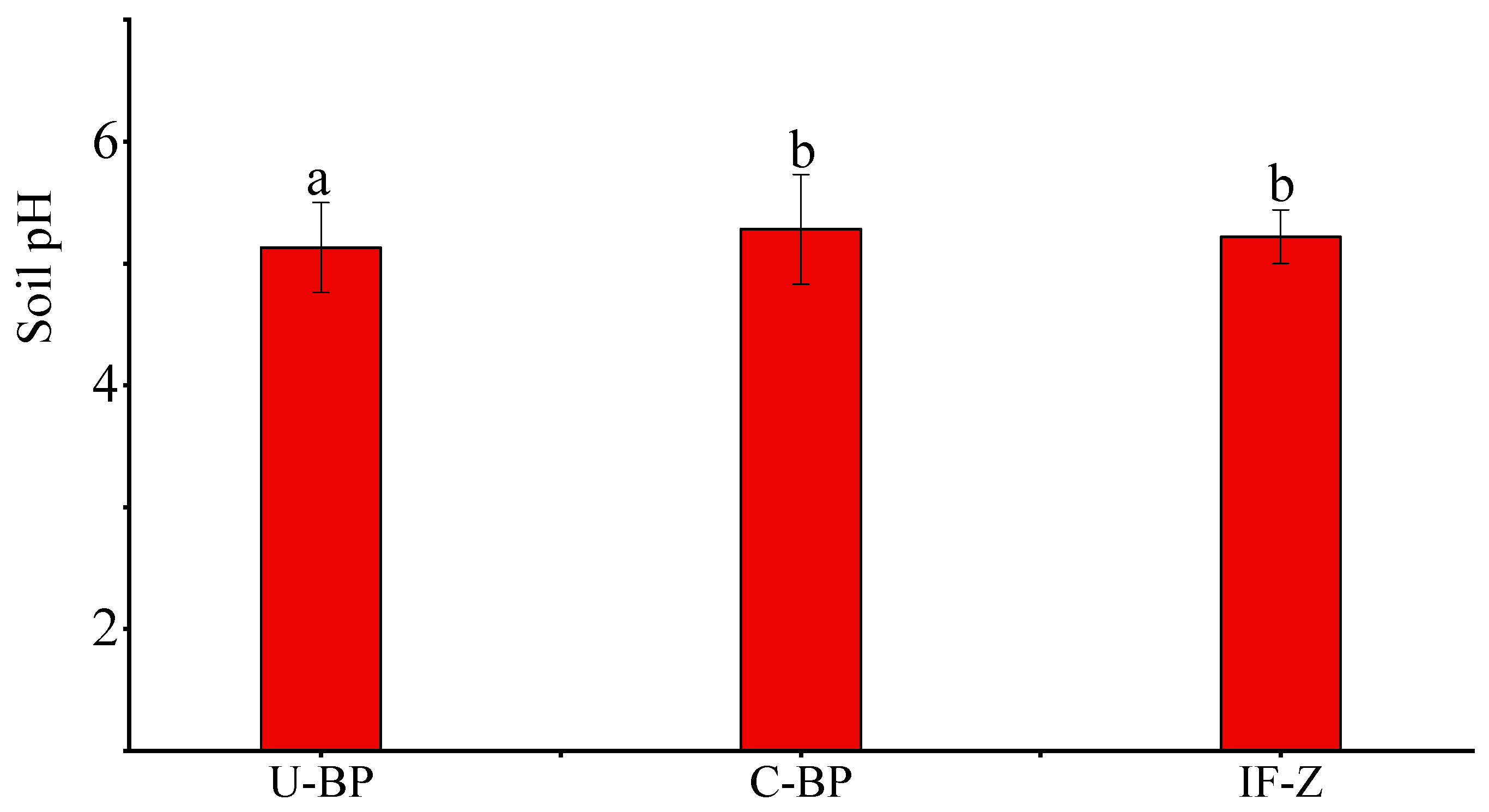
3.1. Soil pH
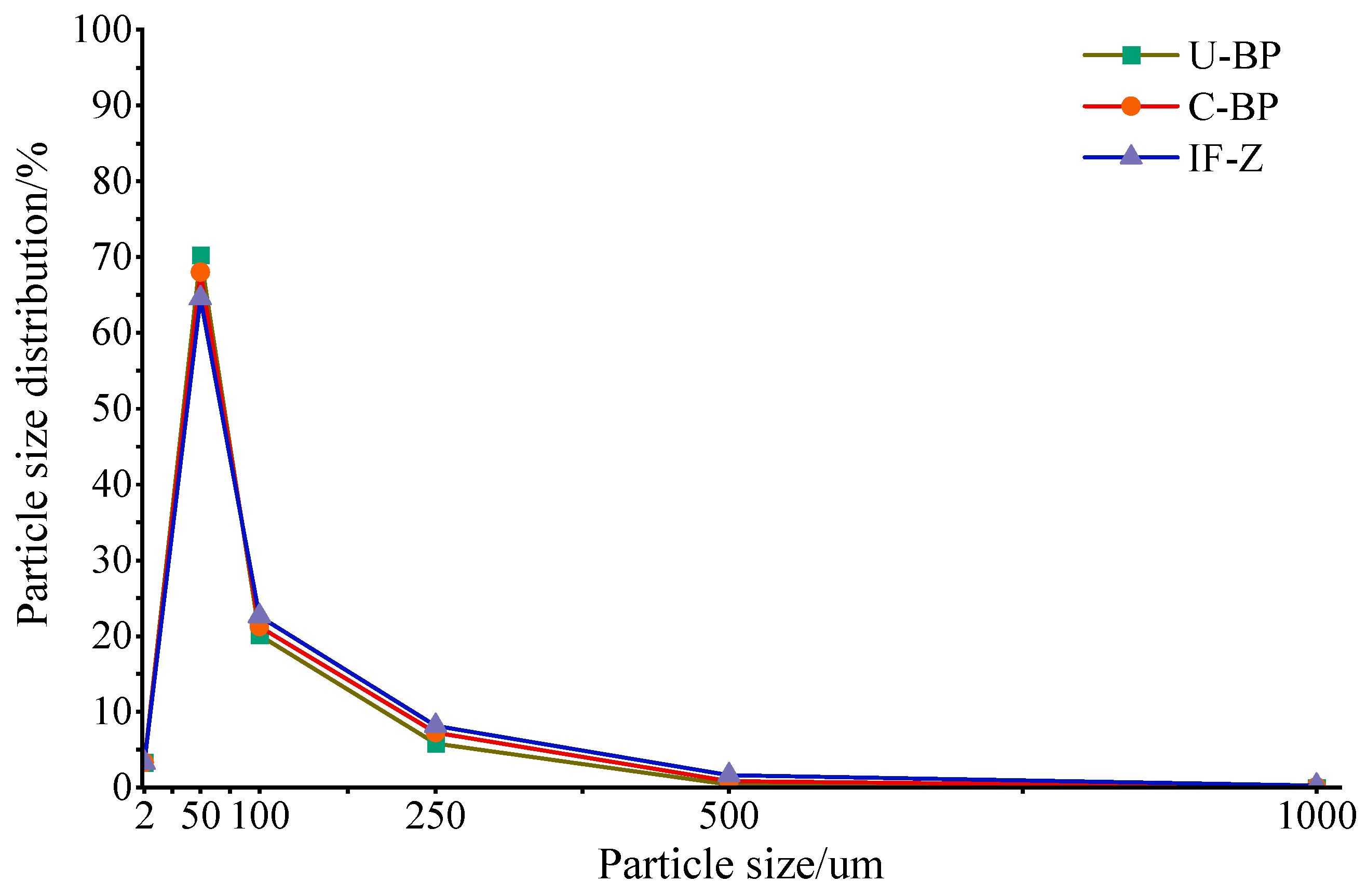
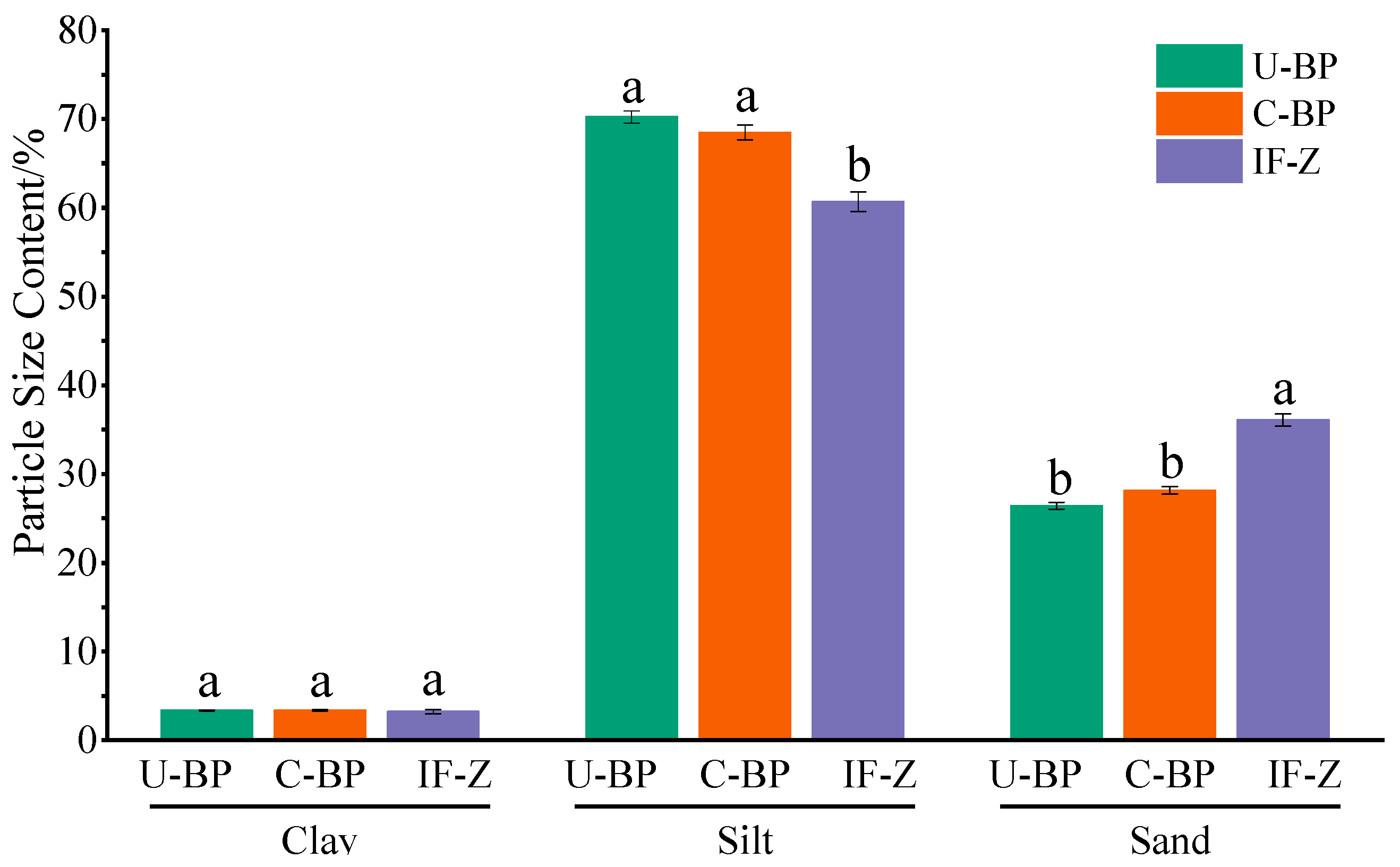
3.2. Soil Particle Size Distribution and Fractal Dimension
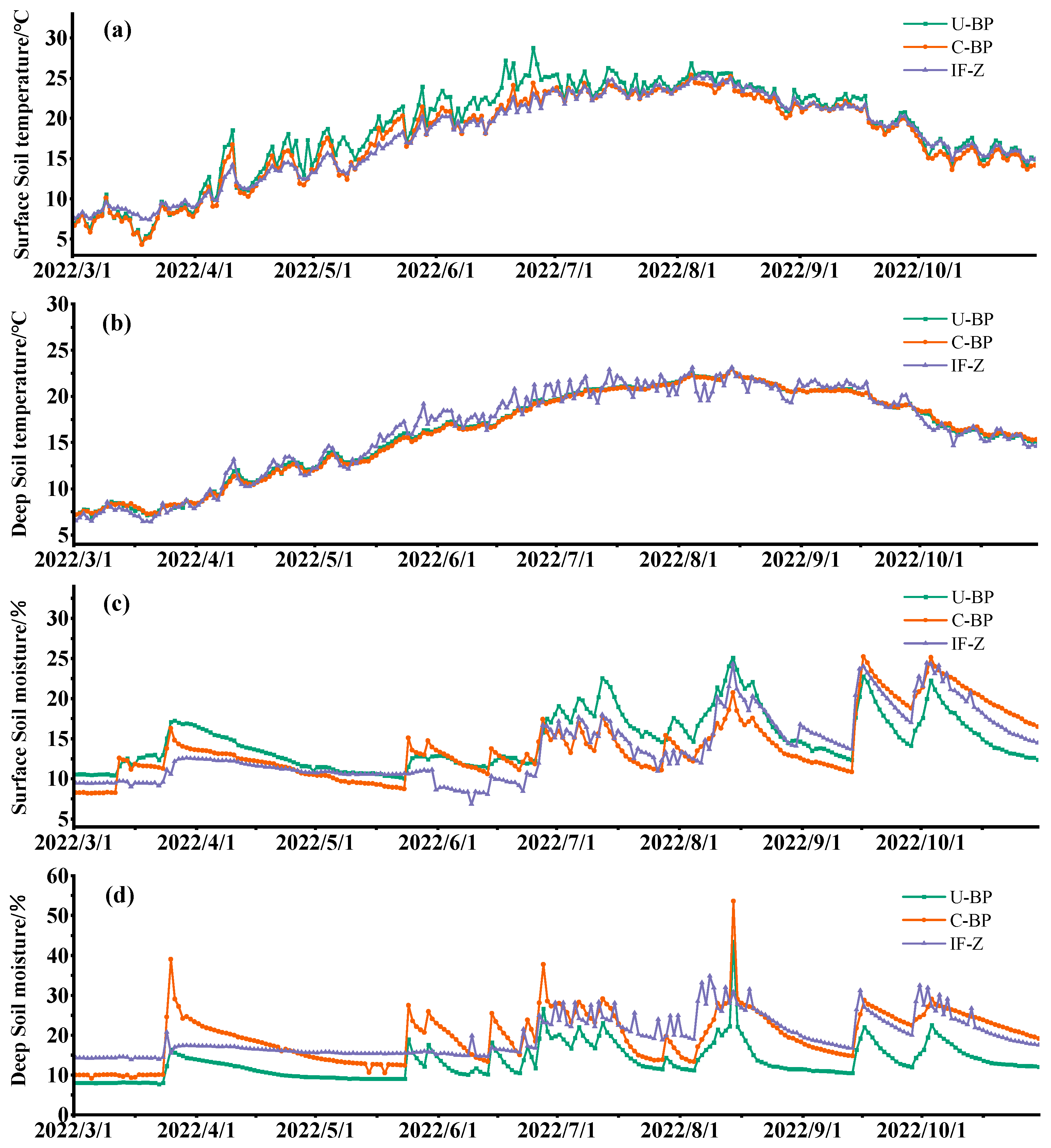
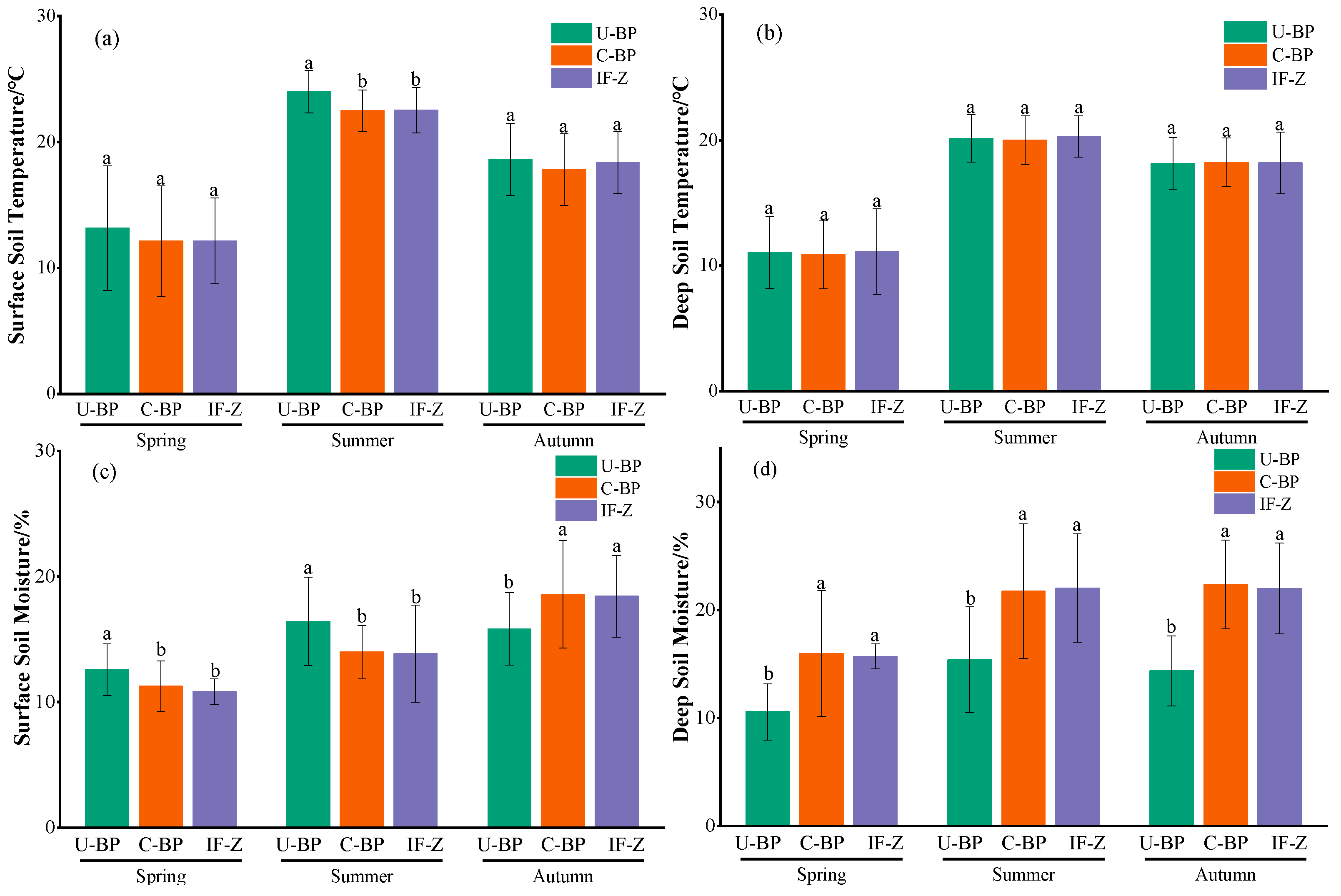
3.3. Dynamics of Soil Temperature
3.4. Dynamics of Soil Moisture
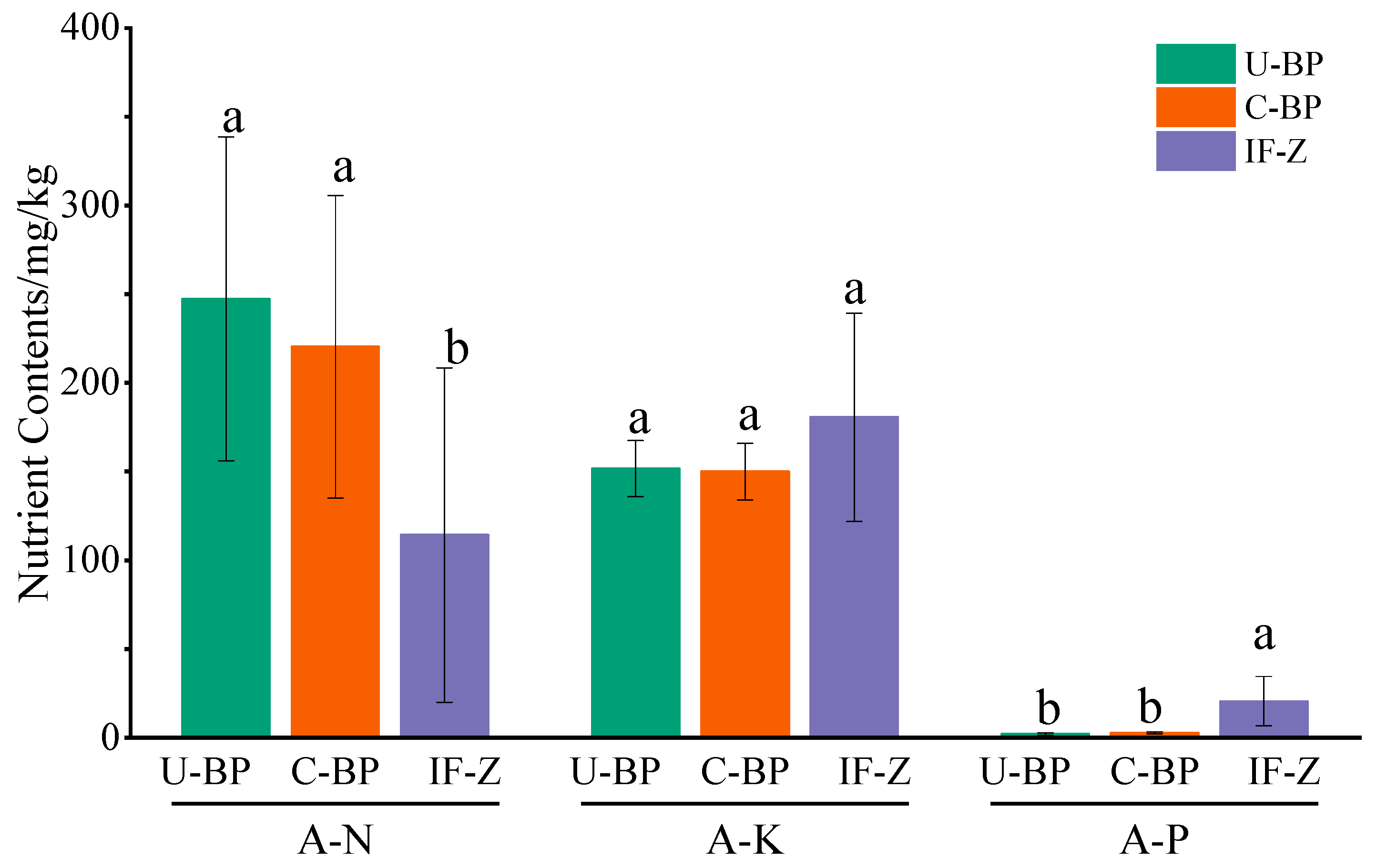
3.5. Soil Nutrients
3.6. Soil Salinity
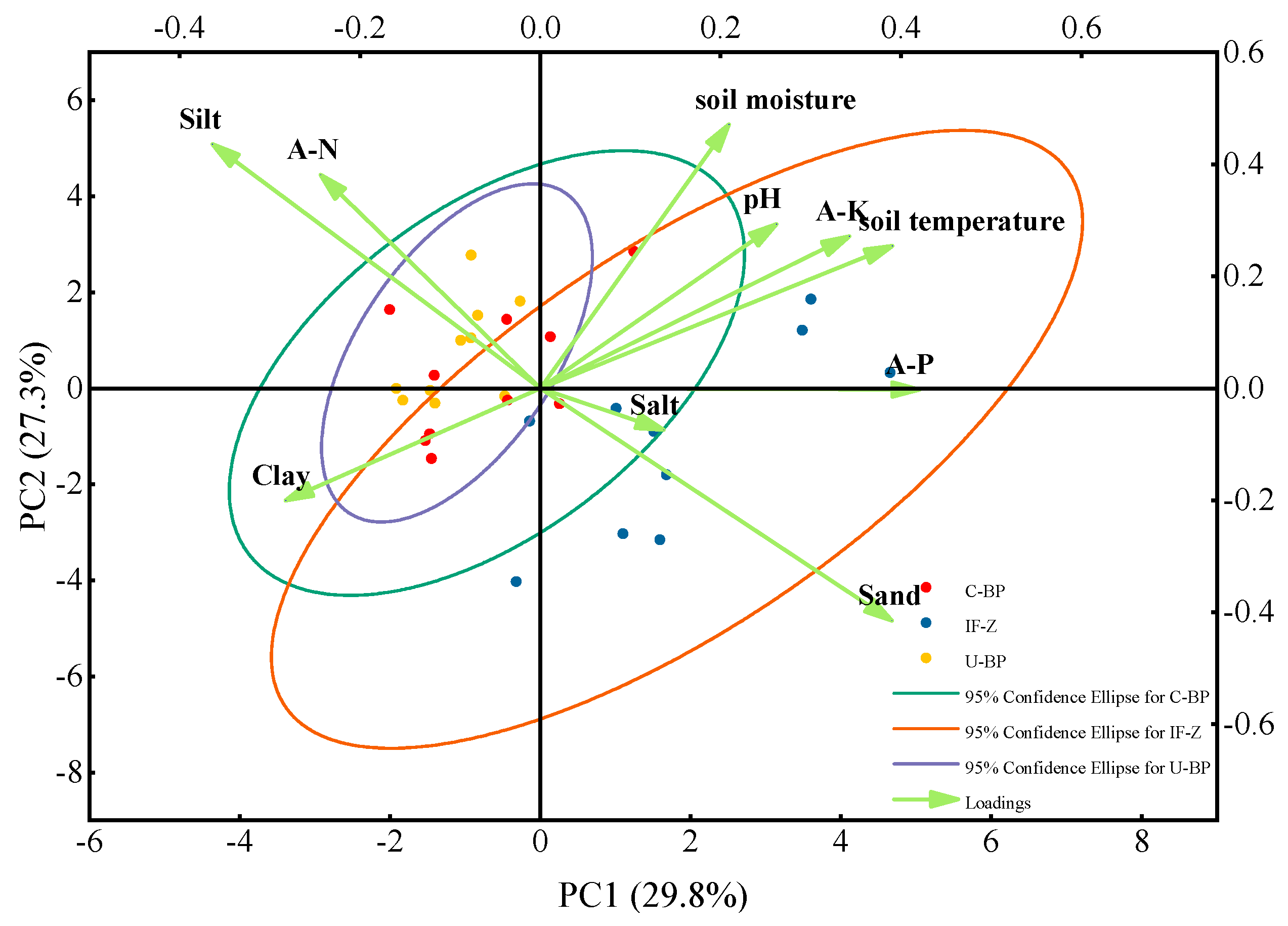
3.7. Similarity Analysis of the Sample Sites Based on Soil Properties
4. Discussion
4.1. Response of Soil pH to the Removal of Diseased P. thunbergii Wood
4.2. Changes in Soil Texture after the Removal of Diseased Wood
4.3. Effects of Wood Clearing on Soil Temperature and Moisture
4.4. Effects of Clearing Diseased Wood on Soil Nutrients and Salinity
4.5. The Direction of Succession of the Forests after Clearing Diseased Wood
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, Q.; Zhang, X.; Li, J.; Ren, J.; Ren, L.; Luo, Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests 2023, 14, 174. [Google Scholar] [CrossRef]
- Li, Z.; Tao, J.; Zong, S. Cold Tolerance in Pinewood Nematode Bursaphelenchus xylophilus Promoted Multiple Invasion Events in Mid-Temperate Zone of China. Forests 2022, 13, 1100. [Google Scholar] [CrossRef]
- Dou, G.; Yan, D.-H. Research Progress on Biocontrol of Pine Wilt Disease by Microorganisms. Forests 2022, 13, 1047. [Google Scholar] [CrossRef]
- Martini, X.; Hughes, M.A.; Conover, D.; Smith, J. Use of Semiochemicals for the Management of the Redbay Ambrosia Beetle. Insects 2020, 11, 796. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-L.; Wu, X.-Q.; Ye, J.-R.; Huang, L. Molecular characterization and functional analysis of three pathogenesis-related cytochrome P450 genes from Bursaphelenchus xylophilus (Tylenchida: Aphelenchoidoidea). Int. J. Mol. Sci. 2015, 16, 5216–5234. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Inácio, M.L. Insights into the Role of Fungi in Pine Wilt Disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef]
- Kim, B.-N.; Kim, J.H.; Ahn, J.-Y.; Kim, S.; Cho, B.-K.; Kim, Y.-H.; Min, J. A short review of the pinewood nematode, Bursaphelenchus xylophilus. Toxicol. Environ. Health Sci. 2020, 12, 297–304. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Inácio, M.L.; Mota, M. The Potential of Esteya spp. for the Biocontrol of the Pinewood Nematode, Bursaphelenchus Xylophilus. Microorganisms 2022, 10, 168. [Google Scholar] [CrossRef]
- Holuša, J.; Hlásny, T.; Modlinger, R.; Lukášová, K.; Kula, E. Felled trap trees as the traditional method for bark beetle control: Can the trapping performance be increased? For. Ecol. Manag. 2017, 404, 165–173. [Google Scholar] [CrossRef]
- Lim, C.-H.; Choi, Y.; Kim, M.; Jeon, S.W.; Lee, W.-K. Impact of Deforestation on Agro-Environmental Variables in Cropland. North Korea. Sustainability 2017, 9, 1354. [Google Scholar] [CrossRef]
- Rannestad, M.M.; Gessesse, T.A. Deforestation and Subsequent Cultivation of Nutrient Poor Soils of Miombo Woodlands of Tanzania: Long Term Effect on Maize Yield and Soil Nutrients. Sustainability 2020, 12, 4113. [Google Scholar] [CrossRef]
- Wang, B.; Wang, G.; Myo, S.T.Z.; Li, Y.; Xu, C.; Lin, Z.; Qian, Z.; Tang, L. Deforestation for Agriculture Temporarily Improved Soil Quality and Soil Organic Carbon Stocks. Forests 2022, 13, 228. [Google Scholar] [CrossRef]
- Junior, C.R.P.; Pereira, M.G.; Filho, J.D.S.O.; Beutler, S.J. Can topography affect the restoration of soil properties after deforestation in a semiarid ecosystem? J. Arid. Environ. 2019, 162, 45–52. [Google Scholar] [CrossRef]
- Durrer, A.; Margenot, A.J.; Silva, L.C.R.; Bohannan, B.J.M.; Nusslein, K.; van Haren, J.; Andreote, F.D.; Parikh, S.J.; Rodrigues, J.L.M. Beyond total carbon: Conversion of amazon forest to pasture alters indicators of soil C cycling. Biogeochemistry 2021, 152, 179–194. [Google Scholar] [CrossRef]
- Lemos, L.N.; Pedrinho, A.; de Vasconcelos, A.T.R.; Tsai, S.M.; Mendes, L.W. Amazon deforestation enriches antibiotic resistance genes. Soil Biol. Biochem. 2021, 153, 108110. [Google Scholar] [CrossRef]
- Alexander, S.P. What Drives Deforestation in the Brazilian Amazon? J. Environ. Econ. Manag. 1999, 37, 26–43. [Google Scholar] [CrossRef]
- Mueller, R.C.; Paula, F.S.; Mirza, B.S.; Rodrigues, J.L.; Nüsslein, K.; Bohannan, B.J. Links between plant and fungal communities across a deforestation chronosequence in the Amazon rainforest. ISME J. 2014, 8, 1548–1550. [Google Scholar] [CrossRef]
- Nourbakhsh, F. Decoupling of soil biological properties by deforestation. Agric. Ecosyst. Environ. 2007, 121, 435–438. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Z.; Yao, Y.; Kong, W.; Zhao, Z.; Shao, M.; Wei, X. Effects of slope morphology and position on soil nutrients after deforestation in the hilly loess region of China. Agric. Ecosyst. Environ. 2021, 321, 107615. [Google Scholar] [CrossRef]
- Fujisaki, K.; Perrin, A.-S.; Garric, B.; Balesdent, J.; Brossard, M. Soil organic carbon changes after deforestation and agrosystem establishment in Amazonia: An assessment by diachronic approach. Agric. Ecosyst. Environ. 2017, 245, 63–73. [Google Scholar] [CrossRef]
- Chernov, T.I.; Zhelezova, A.D.; Tkhakakhova, A.K.; Ksenofontova, N.A.; Zverev, A.O.; Tiunov, A.V. Soil microbiome, organic matter content and microbial abundance in forest and forest-derived land cover in Cat Tien National Park (Vietnam). Appl. Soil Ecol. 2021, 165, 103957. [Google Scholar] [CrossRef]
- Klanderud, K.; Mbolatiana, H.Z.H.; Vololomboahangy, M.N.; Radimbison, M.A.; Roger, E.; Totland, Ø.; Rajeriarison, C. Recovery of plant species richness and composition after slash-and-burn agriculture in a tropical rainforest in Madagascar. Biodivers. Conserv. 2010, 19, 187–204. [Google Scholar] [CrossRef]
- Getaneh, S.; Honnay, O.; Desie, E.; Helsen, K.; Couck, L.; Shibru, S.; Muys, B. Impact of tree litter identity, litter diversity and habitat quality on litter decomposition rates in tropical moist evergreen forest. For. Ecosyst. 2022, 9, 100023. [Google Scholar] [CrossRef]
- Miao, R.; Ma, J.; Liu, Y.; Liu, Y.; Yang, Z.; Guo, M. Variability of Aboveground Litter Inputs Alters Soil Carbon and Nitrogen in a Coniferous–Broadleaf Mixed Forest of Central China. Forests 2019, 10, 188. [Google Scholar] [CrossRef]
- Alfaro, F.D.; Manzano, M.; Marquet, P.A.; Gaxiola, A. Microbial communities in soil chronosequences with distinct parent material: The effect of soil pH and litter quality. J. Ecol. 2017, 105, 1709–1722. [Google Scholar] [CrossRef]
- Peng, Y.; Li, Y.-J.; Song, S.-Y.; Chen, Y.-Q.; Chen, G.-T.; Tu, L.-H. Nitrogen addition slows litter decomposition accompanied by accelerated manganese release: A five-year experiment in a subtropical evergreen broadleaf forest. Soil Biol. Biochem. 2022, 165, 108511. [Google Scholar] [CrossRef]
- Luan, J.; Li, S.; Wang, Y.; Ding, L.; Cai, C.; Liu, S. Decomposition of diverse litter mixtures affected by drought depends on nitrogen and soil fauna in a bamboo forest. Soil Biol. Biochem. 2022, 173, 108783. [Google Scholar] [CrossRef]
- Tie, L.; Hu, J.; Peñuelas, J.; Sardans, J.; Wei, S.; Liu, X.; Zhou, S.; Huang, C. The amounts and ratio of nitrogen and phosphorus addition drive the rate of litter decomposition in a subtropical forest. Sci. Total Environ. 2022, 833, 155163. [Google Scholar] [CrossRef]
- Rodtassana, C.; Tanner, E.V.J. Litter removal in a tropical rain forest reduces fine root biomass and production but litter addition has few effects. Ecology 2018, 99, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, Y.; Yu, D.; Wang, Y.; Zhao, X.; Zhang, R.; Zhang, W.; Wang, Q.; Xu, M.; Chen, L.; et al. The decomposition of green leaf litter is less temperature sensitive than that of senescent leaf litter: An incubation study. Geoderma 2021, 381, 114691. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, H.; Liu, S.; Li, G.; Hu, X. Asymmetric effects between tree and understorey litters on mixed litter decomposition in temperate Quercus variabilis forest. Sci. Total Environ. 2022, 806, 150939. [Google Scholar] [CrossRef] [PubMed]
- Rawlik, K.; Kasprowicz, M.; Nowiński, M.; Jagodziński, A.M. The afterlife of herbaceous plant species: A litter decomposition experiment in a temperate oak-hornbeam forest. For. Ecol. Manag. 2022, 507, 120008. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, S.; Liu, Q.; Jiang, J. Influence of Soil Moisture on Litter Respiration in the Semiarid Loess Plateau. PLoS ONE 2014, 9, e114558. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Du, C.; Guo, X.; Shi, W. Heterogeneity of water-retention capacity of forest and its influencing factors based on meta-analysis in the Beijing-Tianjin-Hebei region. J. Geogr. Sci. 2021, 31, 69–90. [Google Scholar] [CrossRef]
- Kim, T.; Kim, J.; Lee, J.; Kim, H.S.; Park, J.; Im, S. Water Retention Capacity of Leaf Litter According to Field Lysimetry. Forests 2023, 14, 478. [Google Scholar] [CrossRef]
- Li, Q.; Lee, Y.E.; Im, S.J. Characterizing the Interception Capacity of Floor Litter with Rainfall Simulation Experiments. Water 2020, 12, 3145. [Google Scholar] [CrossRef]
- Li, N.; Huo, L.; Zhang, X. Classification of pine wilt disease at different infection stages by diagnostic hyperspectral bands. Ecol. Indic. 2022, 142, 109198. [Google Scholar] [CrossRef]
- Lu, X.; Huang, J.; Li, X.; Fang, G.; Liu, D. The interaction of environmental factors increases the risk of spatiotemporal transmission of pine wilt disease. Ecol. Indic. 2021, 133, 108394. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, L.; Wu, C.; Yu, G.; Wang, Z.; Fan, J.; Tu, C. The Effect of Regulating Soil pH on the Control of Pine Wilt Disease in a Black Pine Forest. Forest 2023, 14, 1583. [Google Scholar] [CrossRef]
- Lee, J.-W.; Nam, I.; Park, J.-H.; Huh, M.-J.; Park, I.-K. Nematicidal activity of (Z)-ligustilide isolated from Angelica tenuissima Nakai root extract against the pine wood nematode Bursaphelenchus xylophilus. J. Asia Pac. Entomol. 2022, 25, 101957. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Deep sequencing analyses of pine wood nematode Bursaphelenchus xylophilus microRNAs reveal distinct miRNA expression patterns during the pathological process of pine wilt disease. Gene 2015, 555, 346–356. [Google Scholar] [CrossRef]
- Qin, J.; Wang, B.; Wu, Y.; Lu, Q.; Zhu, H. Identifying Pine Wood Nematode Disease Using UAV Images and Deep Learning Algorithms. Remote Sens. 2021, 13, 162. [Google Scholar] [CrossRef]
- Dale, A.G.; Frank, S.D. Warming and drought combine to increase pest insect fitness on urban trees. PLoS ONE 2017, 12, e0173844. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, W.; Wang, L.; Liu, Y.; Liu, Y.; Feng, Q. Relationship between soil water content and soil particle size on typical slopes of the Loess Plateau during a drought year. Sci. Total Environ. 2019, 648, 943–954. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lv, D. Fractal expression of soil particle-size distribution at the basin scale. Open Geosci. 2022, 14, 70–78. [Google Scholar] [CrossRef]
- Gui, D.; Lei, J.; Zeng, F.; Mu, G.; Zhu, J.; Wang, H.; Zhang, Q. Characterizing variations in soil particle size distribution in oasis farmlands—A case study of the Cele Oasis. Math. Comput. Model. 2010, 51, 1306–1311. [Google Scholar] [CrossRef]
- Fu, X.; Ding, H.; Sheng, Q.; Zhang, Z.; Yin, D.; Chen, F. Fractal Analysis of Particle Distribution and Scale Effect in a Soil–Rock Mixture. Fractal Fract. 2022, 6, 120. [Google Scholar] [CrossRef]
- Zhong, W.; He, N.; Cosgrove, T.; Zhu, Y.J.; Fu, L. Analysis of the correlation between fractal dimension of gravelly soil and debris-flow initiation through in-situ experiments. Appl. Ecol. Environ. Res. 2019, 17, 7573–7589. [Google Scholar] [CrossRef]
- Li, Q.; Lee, Y.E.; Im, S. Effect of Pennisetum giganteum z.x.lin mixed nitrogen-fixing bacterial fertilizer on the growth. quality, soil fertility and bacterial community of Pakchoi (Brassica chinensis L.). PLoS ONE 2020, 15, e0228709. [Google Scholar]
- Cheng, X.; Zhou, T.; Liu, S.; Sun, X.; Zhou, Y.; Xu, L.; Xie, B.; Ying, J.; Shi, Y. Effects of Climate on Variation of Soil Organic Carbon and Alkali-Hydrolyzed Nitrogen in Subtropical Forests: A Case Study of Zhejiang Province, China. Forest 2023, 14, 914. [Google Scholar] [CrossRef]
- Štípek, K.; Vaněk, V.; Száková, J.; Černý, J.; Šilha, J. Temporal variability of available phosphorus, potassium and magnesium in arable soil. Plant Soil Environ. 2004, 50, 547–551. [Google Scholar] [CrossRef]
- Soleimanifard, S.; Hamdami, N. Modelling of the Sorption Isotherms and Determination of the Isosteric Heat of Split Pistachios, Pistachio Kernels and Shells. Czech J. Food Sci. 2018, 36, 268–275. [Google Scholar] [CrossRef]
- Afonso, J.C.; Renata, M.A. Chemical analysis of copper salts manufactured between 1877 and 1995. Quim. Nova 2004, 27, 678–683. [Google Scholar] [CrossRef]
- Gewers, F.L.; Ferreira, G.R.; De Arruda, H.F.; Silva, F.N.; Comin, C.H.; Amancio, D.R.; Costa, L.D.F. Principal Component Analysis: A Natural Approach to Data Exploration. ACM Comput. Surv. 2021, 54, 70. [Google Scholar] [CrossRef]
- Jin, Z.; Luo, D.; Yu, Y.; Yang, S.; Zhang, J.; Cao, G. Soil pH changes in a small catchment on the Chinese Loess Plateau after long-term vegetation rehabilitation. Ecol. Eng. 2022, 175, 106503. [Google Scholar] [CrossRef]
- Tao, J.; Zuo, J.; He, Z.; Wang, Y.; Liu, J.; Liu, W.; Cornelissen, J.H.C. Traits including leaf dry matter content and leaf pH dominate over forest soil pH as drivers of litter decomposition among 60 species. Funct. Ecol. 2019, 33, 1798–1810. [Google Scholar] [CrossRef]
- Rigueiro-Rodríguez, A.; Mosquera-Losada, M.R.; Fernández-Núñez, E. Afforestation of agricultural land with Pinus radiata D. don and Betula alba L. in NW Spain: Effects on soil PH, understorey production and floristic diversity eleven years after establishment. Land Degrad. Dev. 2012, 23, 227–241. [Google Scholar] [CrossRef]
- Mabuhay, J.A.; Nakagoshi, N. Response of soil microbial communities to changes in a forest ecosystem brought about by pine wilt disease. Landsc. Ecol. Eng. 2012, 8, 189–196. [Google Scholar] [CrossRef]
- Liu, X.; Chen, S.; Li, X.; Yang, Z.; Xiong, D.; Xu, C.; Wanek, W.; Yang, Y. Soil warming delays leaf litter decomposition but exerts no effect on litter nutrient release in a subtropical natural forest over 450 days. Geoderma 2022, 427, 116139. [Google Scholar] [CrossRef]
- Li, A.; Fan, Y.; Chen, S.; Song, H.; Lin, C.; Yang, Y. Soil warming did not enhance leaf litter decomposition in two subtropical forests. Soil Biol. Biochem. 2022, 170, 108716. [Google Scholar] [CrossRef]
- Santos, A.F.A.; Carneiro, A.C.P.; Martinez, D.T.; Caldeira, S.F. Eucalyptus Litter Capacity of Stock and Water Retention. Floresta Ambiente 2017, 24, e20150303. [Google Scholar] [CrossRef]
- Xie, J.; Su, D. Water-Holding Characteristics of Litter in Meadow Steppes with Different Years of Fencing in Inner Mongolia, China. Water 2020, 12, 2374. [Google Scholar] [CrossRef]
- Zagyvai-Kiss, K.A.; Kalicz, P.; Szilágyi, J.; Gribovszki, Z. On the specific water holding capacity of litter for three forest ecosystems in the eastern foothills of the Alps. Agric. For. Meteorol. 2019, 278, 107656. [Google Scholar] [CrossRef]
- Wang, Q.-W.; Robson, T.M.; Pieristè, M.; Kenta, T.; Zhou, W.; Kurokawa, H. Canopy structure and phenology modulate the impacts of solar radiation on C and N dynamics during litter decomposition in a temperate forest. Sci. Total Environ. 2022, 820, 153185. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Huang, Y.; Zhi, Y.; Yao, J.; Wang, F.; Yang, W.; Han, L.; Lin, D.; He, Q.; Wei, B.; et al. Assessing the impacts of urbanization on stream ecosystem functioning through investigating litter decomposition and nutrient uptake in a forest and a hyper-eutrophic urban stream. Ecol. Indic. 2022, 138, 108859. [Google Scholar] [CrossRef]
- Bazihizina, N.; Veneklaas, E.J.; Barrett-Lennard, E.G.; Colmer, T.D. Hydraulic redistribution: Limitations for plants in saline soils. Plant Cell Environ. 2017, 40, 2437–2446. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Wan, H.; Qi, H.; Shang, S. Estimating soil water and salt contents from field measurements with time domain reflectometry using machine learning algorithms. Agric. Water Manag. 2023, 285, 108364. [Google Scholar] [CrossRef]
- Lu, D.; Moran, E.; Mausel, P. Linking amazonian secondary succcession forest growth to soil properties. Land Degrad. Develop. 2002, 13, 331–343. [Google Scholar] [CrossRef]
- Rasiah, V.; Florentine, S.; Williams, B.; Westbrooke, M. The impact of deforestation and pasture abandonment on soil properties in the wet tropics of Australi. Geoderma 2004, 120, 35–45. [Google Scholar] [CrossRef]
- Brown, D.D.; Curtis, T.; Adams, E.C. Effects of clear-felling versus gradual removal of conifer trees on the survival of understorey plants during the restoration of ancient woodlands. For. Ecol. Manag. 2015, 248, 15–22. [Google Scholar] [CrossRef][Green Version]
- Veldman, J.W.; Buisson, E.; Durigan, G.; Fernandes, G.W.; Le Stradic, S.; Mahy, G.; Negreiros, D.; Overbeck, G.E.; Veldman, R.G.; Zaloumis, N.P.; et al. Toward an old-growth concept for grasslands, savannas, and woodlands. Front. Ecol. Environ. 2015, 13, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Mottl, O.; Plowman, N.S.; Novotny, V.; Gewa, B.; Rimandai, M.; Klimes, P. Secondary succession has surprisingly low impact on arboreal ant communities in tropical montane rainforest. Ecosphere 2019, 10, e02848. [Google Scholar] [CrossRef]
- Rezende, J.S.; Freire, F.; Filho, J.A.; Freire, M.D.S.; de Almeida, B.G.; Santos, L.C. Impact of deforestation on the soil physical and chemical attributes, and humic fraction of organic matter in dry environments in Brazil. iForest 2021, 15, 465–475. [Google Scholar] [CrossRef]
- Hassler, S.K.; Zimmermann, B.; van Breugel, M.; Hall, J.S.; Elsenbeer, H. Recovery of saturated hydraulic conductivity under secondary succession on former pasture in the humid tropics. For. Ecol. Manag. 2011, 261, 1634–1642. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
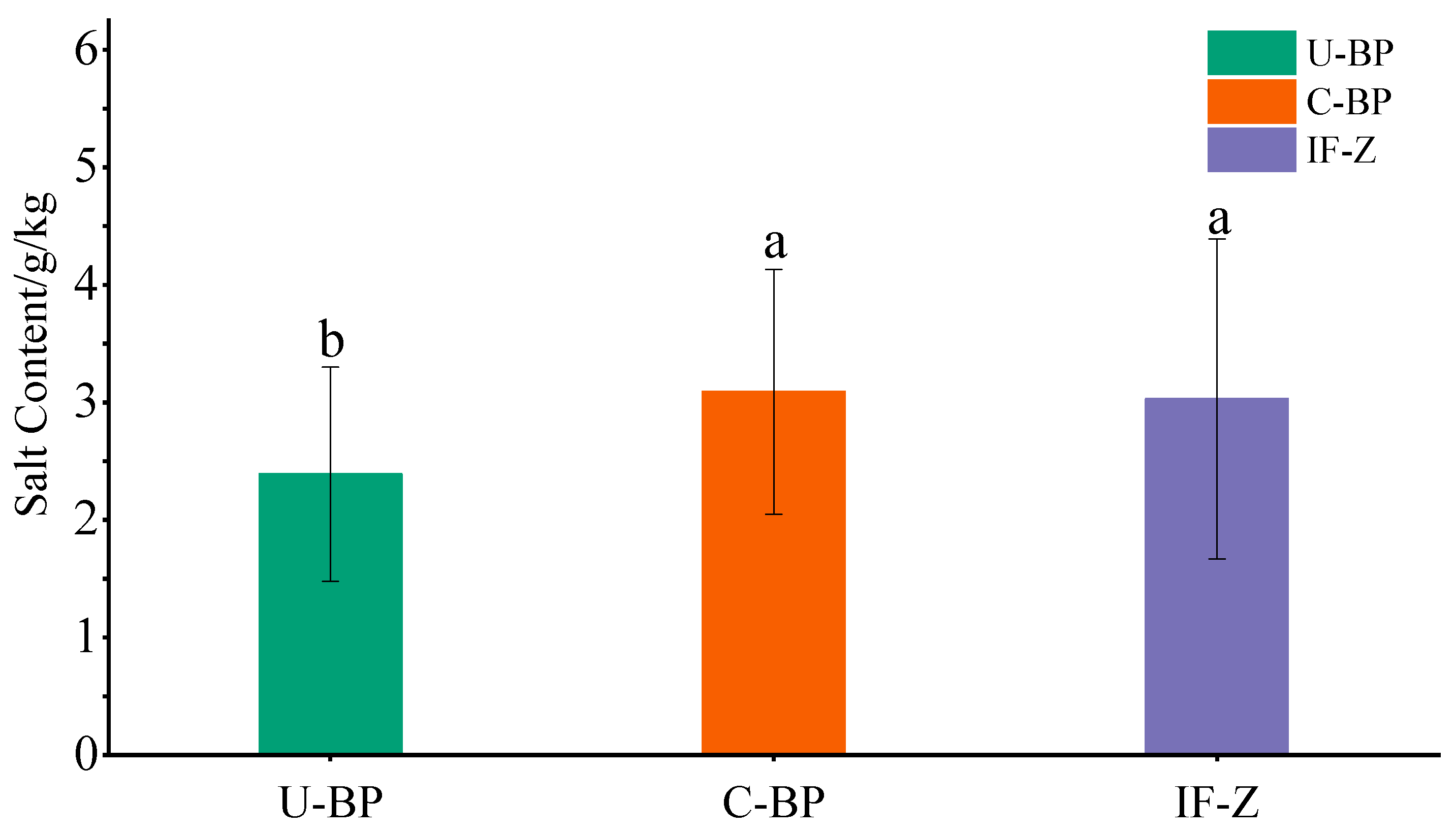{kind=link}
{kind=link}
| Soil Sample Type | Soil Texture Type | Particle Size Content (%) | D | |||||
|---|---|---|---|---|---|---|---|---|
| Clay | Silt | Superfine Sand | Fine Sand | Medium Sand | Coarse Sand | |||
| 0–2 μm | 2–50 μm | 50–100 μm | 100–250 μm | 250–500 μm | 500–1000 μm | |||
| U-BP | loamy sand | 3.35 ± 0.08 | 70.25 ± 0.69 | 20.12 ± 0. 68 | 5.84 ± 0.34 | 0.43 ± 0.12 | 0.01 ± 0.00 | 2.42 ± 0.01 |
| C-BP | sandy loam | 3.37 ± 0.09 | 68.48 ± 0.84 | 21.58 ± 0. 82 | 6.12 ± 0.13 | 0.43 ± 0.19 | 0.02 ± 0.01 | 2.47 ± 0.01 |
| IF-Z | sandy loam | 3.21 ± 0.25 | 60.70 ± 1.10 | 23.61 ± 1. 31 | 9.04 ± 0.58 | 2.85 ± 0.65 | 0.59 ± 0.20 | 2.50 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Tu, C.; Wang, Z.; Fan, J.; Lv, Z.; Fan, Y. The Effect of Clearing Diseased Wood on the Soil’s Physicochemical Properties in Black Pine Forests. Sustainability 2023, 15, 15980. https://doi.org/10.3390/su152215980
Wu C, Tu C, Wang Z, Fan J, Lv Z, Fan Y. The Effect of Clearing Diseased Wood on the Soil’s Physicochemical Properties in Black Pine Forests. Sustainability. 2023; 15(22):15980. https://doi.org/10.3390/su152215980
Chicago/Turabian StyleWu, Chuchen, Chengyi Tu, Zhengyu Wang, Jingjing Fan, Zhenglin Lv, and Ying Fan. 2023. "The Effect of Clearing Diseased Wood on the Soil’s Physicochemical Properties in Black Pine Forests" Sustainability 15, no. 22: 15980. https://doi.org/10.3390/su152215980
APA StyleWu, C., Tu, C., Wang, Z., Fan, J., Lv, Z., & Fan, Y. (2023). The Effect of Clearing Diseased Wood on the Soil’s Physicochemical Properties in Black Pine Forests. Sustainability, 15(22), 15980. https://doi.org/10.3390/su152215980