Abstract
Intensive use of chemicals in agriculture harms the soil, disrupts the ecological balance, and impacts microorganisms. Biofertilizers are gaining traction due to their eco-friendly and cost-effective benefits. This study evaluates the potential of the cyanobacterium MACC-612 (Nostoc piscinale) and plant growth-promoting bacteria (PGPB) (Azospirillum lipoferum, Pseudomonas fluorescens) in enhancing crop growth, yield, and soil health. A two-year field study was conducted using a factorial approach and a completely randomized block design, comprising four replications. The three levels of the cynobacterium (0, 0.3, or 1 g/L of N. MACC-612) and different bacteria strains were used in the experiments. The results demonstrated substantial enhancements in seed number per ear, kernel weight, and yield when using N. piscinale and PGPB, whether used individually or in combination. The soil pH, humus, (NO3− + NO2−)-nitrogen, and soil microbial biomass showed significant increases across both years. The combining application of the N. piscinale (0.3 g/L) with A. lipoferum increased grain yield by 33.20% in the first year and 31.53% in the second. The humus and (NO3− + NO2−)-nitrogen content significantly rose in treatments involving N. piscinale at 0.3 g/L combined with A. lipoferum at about 20.25% and 59.2%, respectively, in comparison to the untreated control. Hence, the most effective approach was the combined use of N. piscinale and A. lipoferum, which enhanced maize growth and soil fertility.
1. Introduction
Maize (Zea mays), recognized as the “queen of grains”, holds the greatest genetic production capacity among crops and stands as one of the foremost cereals globally, yielding the highest food output. It serves as a vital global crop, serving purposes in feeding livestock, human consumption, and various industrial applications [1]. It is assuming a multifaceted and ever-evolving role within global agricultural and food systems, contributing significantly to food and nutrition security [2,3]. A wide range of beneficial microorganisms engage in intricate partnerships with plants, acting as growth facilitators, and playing essential roles in promoting plant health and enriching soil fertility, with some microorganisms possessing known capabilities while others remain subjects of ongoing research. Preliminary research has demonstrated the advantageous potential of collaboration among diverse microorganisms, a concept of growing importance due to contemporary apprehensions surrounding the adverse consequences of agrochemicals, leading to heightened curiosity about advancing our comprehension of cooperative interactions within rhizosphere microbial communities and their potential applications in agriculture [4,5].
Soil microbiomes, led by plant growth-promoting bacteria (PGPB) like rhizospheric bacteria [6] and symbiotic rhizobia [7], are driving the emergence of a new era in sustainable agriculture, and these bacteria are recognized as plant health-promoting bacteria (PHPB) agents [8,9]. Additionally, cyanobacteria (blue-green algae) play a central role in sustainable agriculture by enhancing soil properties, providing nutrients, promoting plant growth, and acting as biocides against soil-borne pathogens, making them valuable biofertilizers and contributors to agricultural sustainability [10,11]. Microalgae and cyanobacteria, functioning as a primary producer, along with bacteria, collectively form the uppermost strata of soil known as the biological soil crust, and this intricate ecosystem plays a pivotal role in augmenting soil fertility and ultimately boosting crop productivity [12,13,14,15,16]. It enhances crop development and well-being through a processes such as nitrogen fixation, the release of trace elements into the soil, nutrient solubilization, production of exopolysaccharides, increasing organic matter, and improved nutrient retention within the plant-soil system, ultimately benefiting plant growth and providing an alternative to chemical fertilizers and pesticides [15,17,18,19,20,21,22,23].
The unreliability of single-strain inoculations in the rhizosphere can be addressed by using PGPB in multispecies consortia, presenting a promising approach for enhancing plant growth, and offering a novel method to discover complementary PGPB within root and soil communities for the development of advanced biofertilizers [24,25,26]. Recently, microalgae-bacteria interaction has been proposed as a potential strategy to improve crop productivity through the generation of phytohormones such as auxin and cytokinins, the synthesis of polysaccharides, which aid in nutrient absorption, and the regulation of numerous biochemical processes and improve soil health [27,28]. A symbiotic partnership between microalgae and bacteria operates through a reciprocal exchange of metabolites. Primarily, bacteria utilize organic carbon released during algal photosynthesis. In return, they facilitate their growth by consuming oxygen, producing carbon dioxide, providing essential nutrients, vitamins, and trace elements to support microalgal growth, and generating growth-promoting substances, chelators, and phytohormones [29,30]. Numerous research investigations indicate that heterotrophic bacteria play a widespread and crucial role in the growth and survival of algae through the provision of hormones and nitrogen sources [31,32,33,34].
Maize (Zea mays) exhibits promising responses when the soil was treated with cyanobacteria as a biofertilizer, demonstrating enhanced growth, nutrient use efficiency, and increased tolerance to abiotic stress [10,14,30,35,36]. Similarly, positive results were obtained by applying cyanobacterial extracts as foliar biostimulants and seed priming, which further contributed to the overall development and well-being of the plants [37,38,39]. Inoculating maize with Azospirillum spp. encourages root development, enhances the uptake of nitrogen from the soil, and promotes the release of carbon compounds from the roots, ultimately leading to improved nitrogen utilization efficiency [40,41]. Inoculating maize seeds with Pseudomonas spp. strains reduced leaf dehydration, boosted plant growth, raised relative water content, enhanced leaf water potential, improved root adhesion, increased soil/root tissue ratio, bolstered aggregate stability, grain yield, and fixed nitrogen [42,43]. To ensure the effectiveness of these consortia, it is crucial for their diverse members to maintain positive interactions with each other over an extended period. Hence, the current research was conducted under field conditions to evaluate the potential of two-member consortia consisting of cyanobacterial biomass and PGPB strains applied to maize (Z. mays) plants to measure their impact on plant growth and soil health.
2. Materials and Methods
2.1. Experimental Site and Design
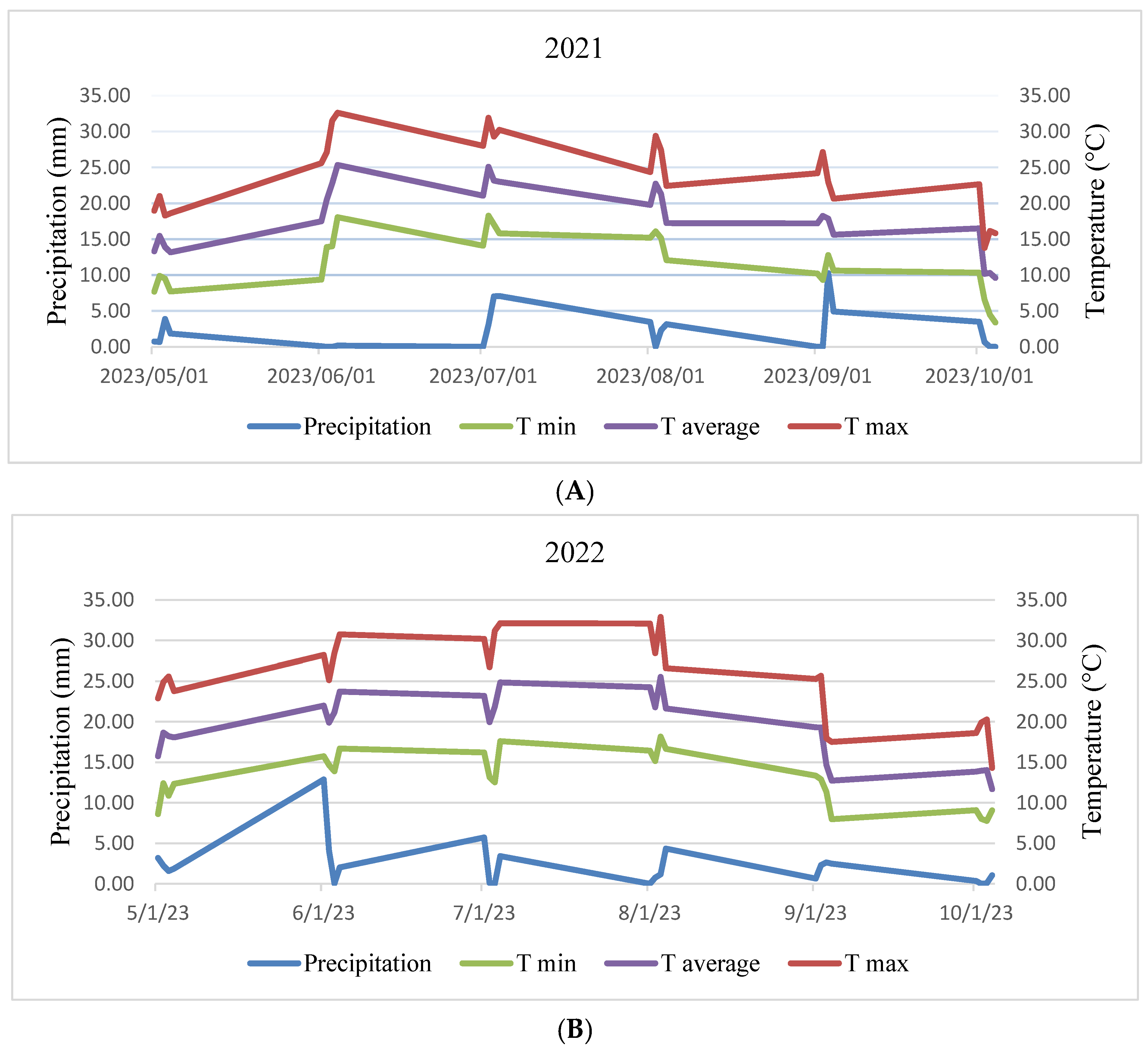
Experiments were conducted with same treatment in two different fields located at (47°53′32.3″ N 17°15′59.0″ E) and (47°54′26.7″ N 17°15′09.3″ E) in the university experimental farm of Széchenyi István University in Mosonmagyaróvár, Hungary, during two different seasons in 2021 and 2022. Figure 1 displays the recorded rainfall, maximum air temperature (T max), and minimum air temperature (T min) at both years throughout the duration of the research. Prior to initiating the experiment, soil samples were collected for chemical analysis, and the findings are displayed in Section 3 (Results section).
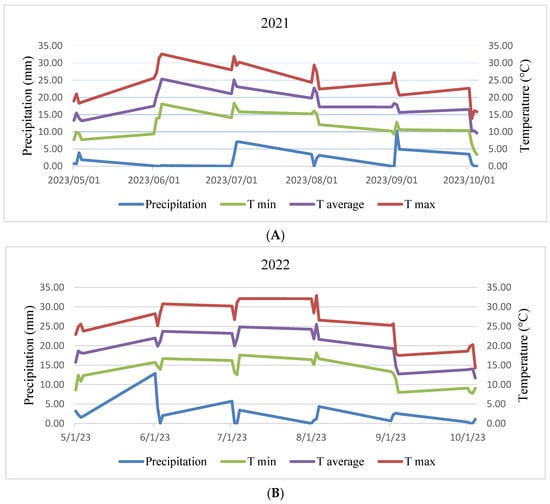
Figure 1.
Illustration of weekly mean precipitation and temperature data recorded in the experimental field during the interval across from sowing to harvest in the production years of 2021 (A) and 2022 (B). Where: T min = Temperature minimum, T average = Temperature average, T max = Temperature maximum.
The study followed a completely randomized block design (CRBD) with four replication and a total of 3 × 3 treatments. The experimental design included two main factors, which were: Cyanobacterial biomass (MACC-612, N. piscinale) and PGPB (such as A. lipoferum (strain NF5) and P. fluorescens (strain NCAIM B01666). The three levels of the cyanobacterial biomass (control, 0.3 g/L of MACC-612, and 1.0 g/L of MACC-612) and three levels of bacteria strains (control, A. lipoferum (NF5), and P. fluorescens (NCAIM B01666)) were used for the experiment (Table 1).

Table 1.
Treatments combination of the cyanobacteria and PGPB.
The Cyanobacterium strain (MACC-612, N. piscinale) was obtained from the Mosonmagyaróvár Algal Culture Collection (MACC), at the Albert Kázmér Faculty of Agricultural and Food Sciences in Mosonmagyaróvár, Széchenyi István University, Hungary. In order to generate the necessary biomass for the experiments, as previously detailed by [44], the strain was cultivated in a laboratory algae culture system, using a Tamiya medium for a duration of 6 days, following the outlined protocol [45]. The cyanobacterium biomass was produced in laboratory culture units. It was illuminated from below with a light intensity of 130 µmol m−2 s−1 and grown in a Tamiya nutrient solution [46], with a starting concentration of 10 mg/L algal dry weight (dwt). 20 L/ha of filtered compressed air enriched with 1.5% CO2 during the light period was used for aerating the culture strains [44]. The cultures grown in these conditions for 10 days were then centrifuged for 15 min at 3000 rpm (Sigma 6 K15, Sigma Laborzentrifugen GmbH, Osterode am Harz, Germany) and freeze-dried using Gamma 1–20 (Martin Christ Gefriertrocknungsanlagen GmbH, Osterode am Harz, Germany) and stored at −18 °C. Biomass samples were re-suspended in distilled water and sonicated (VirTis, VirSonic 600 Ultrasonic Cell Disruptor, SP VirTis, Gardiner, NY, USA) 3 min just before the soil application treatments. The freeze-dried biomass of MACC-612 (N. piscinale) was reconstituted in distilled water at varying concentrations stated above and then subjected to a 3 min sonication process using the VirTis VirSonic 600 Ultrasonic Cell Disruptor just prior to soil application. Each bacteria strain was randomly combined with each of the three levels of cyanobacterium biomass with four replications giving a total of 36 (3 × 3 × 4) plots. The plants underwent treatment using either tap water as a control or with the cyanobacterium biomass at concentrations of 0.3 or 1.0 g/L dry weight, with the selection of these concentrations being informed by previous research findings on maize [45].
A. lipoferum (NF5) and P. fluorescens (NCAIM B01666) were obtained in liquid-based formulation from BioFil Microbiological, Gene Technological and Biochemical LLC, Budapest, Hungary. The bacterial strains were cultured in a liquid medium that was enriched with yeast extract (3 g/L), glucose (5 g/L), sucrose (5 g/L) and then subjected to a 48 h incubation period in a gyrotary water bath shaker (New Brunswick Scientific Co., Inc. Edison, NJ, USA) set at 120 rotations per minute and maintained at a temperature of 37 °C. The cell concentration of A. lipoferum and P. fluorescens were 7.8 × 108 CFU/mL and 1.02 × 109 CFU/mL, respectively, measured by DEN-1, McFarland Densitometer (suspension turbidity detector).
The cyanobacterium biomass suspension and PGPB treatments were introduced to the soil using a 15 L knapsack sprayer during the sowing process, at an application rate of 300 L/ha. The planting was carried out using a 163-cc mini tractor. For the experimental cultivation, a type of Zea mays hybrid, obtained from (Saaten Union-Körner kernels Grains), was utilized. Sowing was conducted using a row spacing of 75 cm, a plant spacing of 20 cm, and a sowing depth of 6 cm. Each plot covered an area of 28.5 m2 (3 × 9.5 m), allowing for 256.5 m2 per replication and requiring a total of 1026 m2 for four replications. Plots were spaced 0.5 m apart, while blocks were kept 1 m apart.
2.2. Plant Yield Attributes
The study involved measuring several parameters from 4 plants taken from each central plot at the end of experiment, including plant height (cm), number of grains per ear, and the thousand kernel weight (kg). The overall yield in ton/ha was then determined based on the harvest from each individual plot. The harvest took place at the end of October in both consecutive years.
2.3. Soil Collection and Analysis
Soil samples were extracted from two consecutive years of drilling within the experimental field, spanning depths from 0 to 20 cm. The soil collected underwent a sieving process using a 2 mm mesh sieve, followed by thorough mixing to ensure uniformity. Soil pH, humus, (NO3− + NO2−)-nitrogen, P and K were subjected to analysis. The pH of the soil’s particles smaller than 2 mm was assessed using a solution of 1 M KCl in a 1:2.5 ratio, following established methods for determining soil pH as outlined by [47].
Flow Injection Analysis (FIA) was utilized to determine the concentrations of nitrate and nitrite nitrogen in the samples as outlined in the procedure specified by [48]. Samples were mixed with imidazole buffer and treated with copperized cadmium to convert nitrate to nitrite. The nitrite reacted with sulfanilamide, forming a red azo dye with N-(1-Naphthyl)-ethylenediamine (NED), which was detected spectrophotometrically at a specific wavelength (546 nm) using a flow cell or a reaction coil within the FIA system.
2.4. Soil Microbial Biomass Evaluation
The microorganisms were evaluated using the agar-plate method according to [49], which is the predominant cultural approach for assessing soil microbial populations, aiding in their identification and quantification. Bacterial populations were assessed by culturing them on nutrient agar [50]. Dextrose nitrate agar was formulated to quantify the population of actinomycetes according to [51]. In direct approaches, microorganisms in the soil are quantified by counting the number of colonies forming units (CFU) through a soil dilution series using the most probable number (MPN). The MPN method involves dispersing soil samples in a sequence of dilutions to estimate the density of the population by observing the presence or absence of microbial cells [52]. Therefore, if microbial growth is detected in the 10−4 dilution but not in the 10−5 dilution, the estimated number of cells falls within the range of 104 to 105. Utilizing a logarithmic scale, like log CFU, facilitates the representation of these counts in a more practical and informative fashion.
log CFU = log10(CFU)
In this equation, CFU denotes the precise tally of colonies, while log CFU signifies the base 10 logarithm of the colony count. Through this logarithmic transformation, values are condensed, facilitating the comparison and visualization of variations in microbial populations.
2.5. Statistical Analysis
All statistical computations were conducted using R software (version 4.3.1) [53], utilizing the software package known as “agricolae” (version 1.3-6) [54]. The outcomes from all the experiments, which exhibited a normal distribution pattern, underwent two-way analysis variance (ANOVA p ≤ 0.05) used to test yield attributes, soil chemical analysis and microbial abundance. The results displayed the average values of the treatments’ replicates ± standard deviation of the mean within each treatment. Following this, Tukey’s HSD post hoc analysis was applied at a significance level of p ≤ 0.05. To streamline data interpretation and the statistical analysis of the microbial biomass, we applied a logarithmic transformation, which resulted in condensed values. This approach simplifies the process of comparing and visualizing fluctuations within microbial populations.
3. Results
The soil type was Danubian alluvial soil at the experimental area. The soil of the experimental field was slightly basic. A decrease in humus content was observed in the soil from 2021 to 2022. The combined nitrate and nitrite nitrogen levels exhibited a slight increase from 2021 to 2022 (Table 2).
Table 2.
The soil chemical characteristics collected from the experimental field prior to planting.
Figure 1 displays meteorological data for the two years. In the 2022 field trial, there was a higher precipitation, particularly during the vegetative growth phase compared to the 2021 production year. However, the precipitation during the reproductive stage was relatively consistent between the first and second years.
3.1. Plant Yield Parameters
Table 3 provides data on how the plant growth parameters of Z. mays were affected by different concentrations of cyanobacterial biomass and PGPB strains. Statistical analysis showed a significant (p ≤ 0.05) increase in the number of seeds per ear, thousand kernel weight, and yield when N. piscinale and PGPB were applied alone or jointly in both production years. However, there was no statistically significant difference in plant height across the observed period (Table 3).
Table 3.
Impact of single and combined application cyanobacterial biomass and PGPB on Z. mays yield components.
The highest number of seeds per ear and thousand kernel weight was documented when using N. piscinale at a concentration of 0.3 g/L in combination with A. lipoferum in both years, whereas the lowest was observed in the untreated trials. However, in 2021, the number of seeds per ear was statistically comparable across treatments, except for the control group and N. piscinale at 1 g/L combined with P. fluorescens, where notable deviations were observed. Moreover, the thousand grain weight was statistically similar to both N. piscinale at 0.3 g/L combined with A. lipoferum and P. fluorescens, and N. piscinale at 1 g/L combined with A. lipoferum in 2021.
The utilization of N. piscinale at a concentration of 0.3 g/L in combination with A. lipoferum resulted in a significant upsurge in grain yield, demonstrating a remarkable 33.20% enhancement in the initial year and a substantial 31.53% increase in the subsequent year when compared with untreated plots. In the second year, there were no significant statistical differences in grain yield among the treatments that included a combination of A. lipoferum and P. fluorescens with a 0.3 g/L concentration of N. piscinale and A. lipoferum combined with a 1 g/L concentration of N. piscinale. In general, the grain yield demonstrated a high performance in the second year as opposed to the initial year.
3.2. Soil Chemical Properties
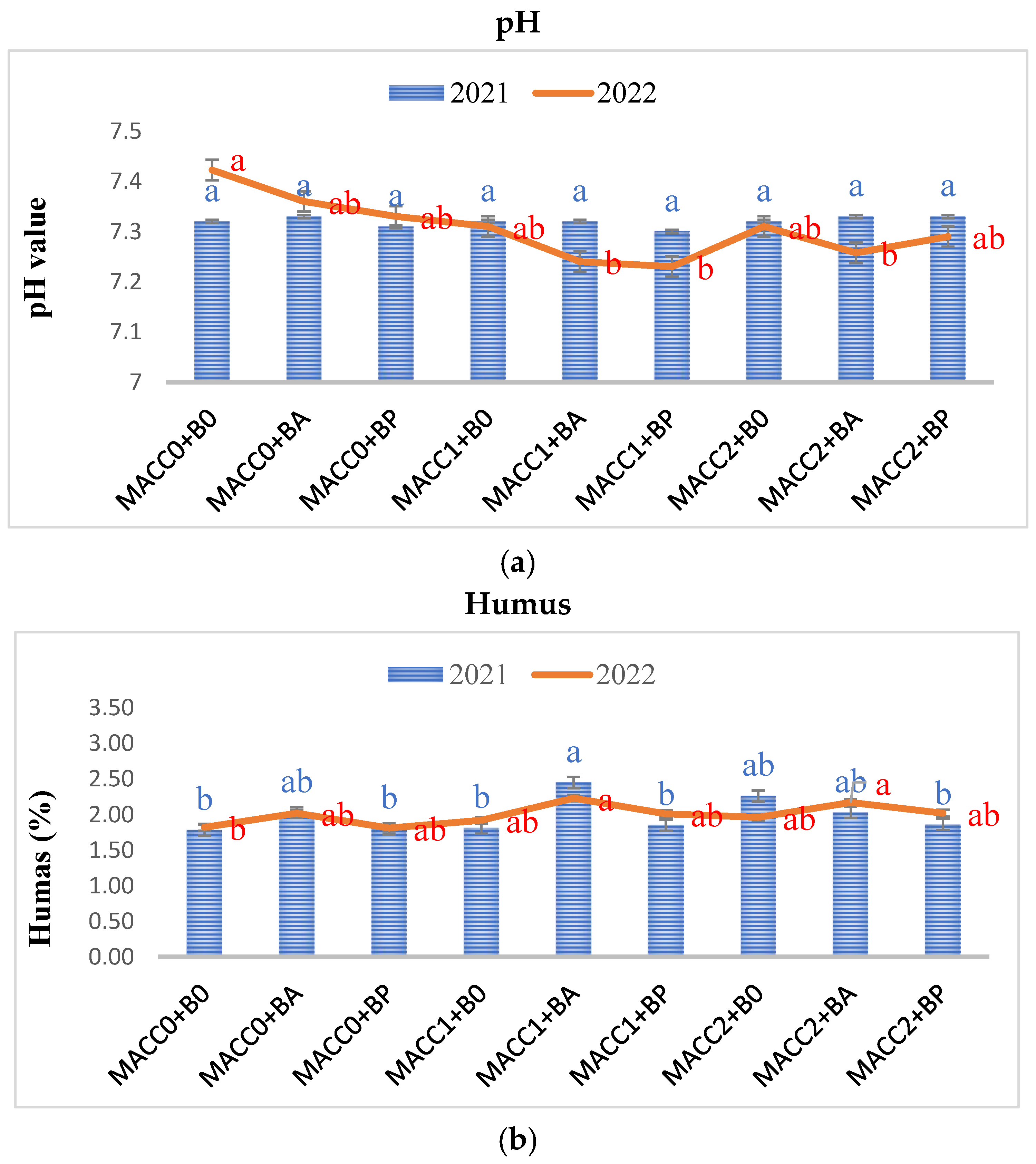
Following the Z. mays plant harvest from each treatment, the soil was gathered and subjected to analysis to assess the post-treatment impacts on soil properties, including pH, organic matter content, (NO3− + NO2−)-nitrogen, phosphorus (P), and potassium (K) levels. Overall, whether applied individually or in combination, different concentrations of N. piscinale and PGPB strains noticeably elevated soil pH, humus, and (NO3− + NO2−)-nitrogen in 2022, whereas P and K were statistically insignificant (p ≤ 0.05) in both years, as illustrated in Figure 2, Figure 3 and Figure 4. However, during the initial year of the investigation, no statistically significant differences were observed except in the humus. In contrast to the findings in the initial year, the results of the soil analysis during the second year showed a statistically significant effect between N. piscinale and PGPB applied in alone/combination with respect to pH, humus, and (NO3− + NO2−)-nitrogen indicating that the alone or combined application of N. piscinale and PGPB increases soil fertility.
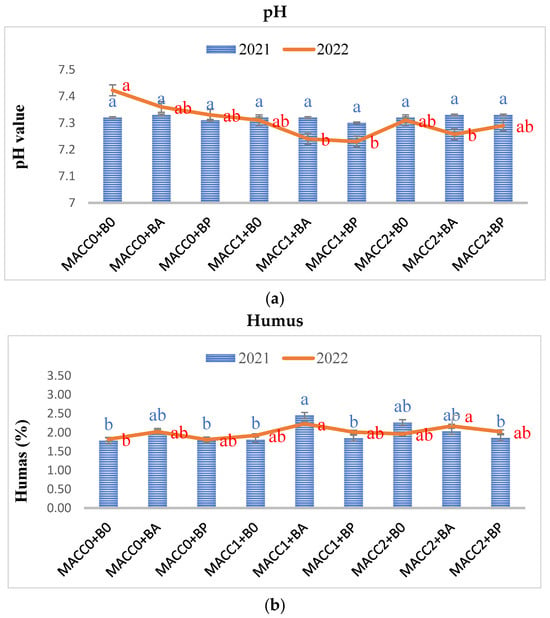
Figure 2.
Effect of different applications of N. piscinale biomass and PGPB on soil pH and humus content. Values marked a, b, letter shows significant difference (p ≤ 0.05) according to Tukey test. Where: MACC0 and B0 = control; MACC1 = 0.3 g/L of N. piscinale MACC-612; MACC2 = 1.0 g/L of N. piscinale MACC-612; BA = A. lipoferum; BP = P. fluorescens.
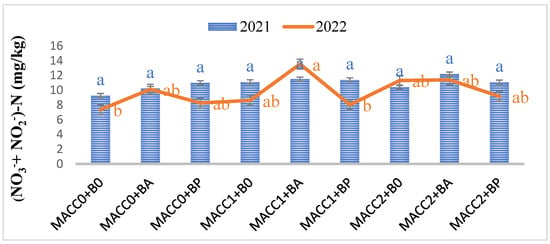
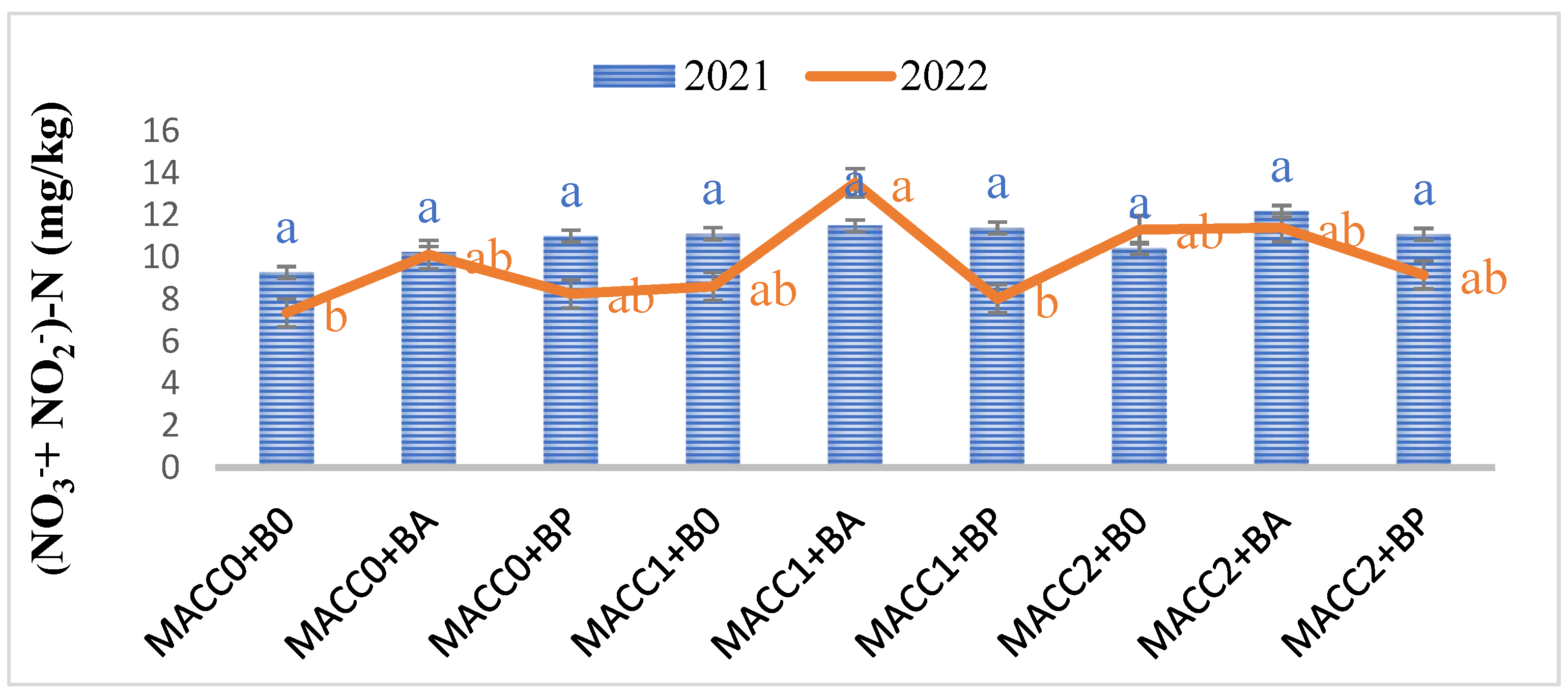
Figure 3.
Effect of different applications of N. piscinale biomass and PGPB on soil Nitrate-Nitrite Nitrogen content. Values marked a, b, letter shows significant difference (p ≤ 0.05) according to Tukey test. Where: MACC0 and B0 = control; MACC1 = 0.3 g/L of N. piscinale MACC-612; MACC2 = 1.0 g/L of N. piscinale MACC-612; BA = A. lipoferum; BP = P. fluorescens.
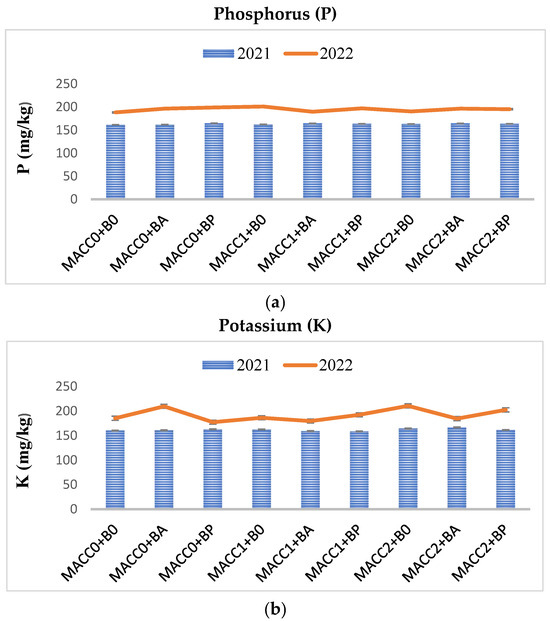
Figure 4.
Effect of different applications of N. piscinale biomass and PGPB on soil phosphorus and potassium content. (Where: MACC0 and B0 = control; MACC1 = 0.3 g/L of N. piscinale MACC-612; MACC2 = 1.0 g/L of N. piscinale MACC-612; BA = A. lipoferum; BP = P. fluorescens).
In 2022, the highest pH value (7.42) was observed in the control group, signifying slightly alkaline soil conditions. In contrast, lower pH values were recorded in the treatments, involving N. piscinale at a concentration of 0.3 g/L in combination with A. lipoferum (7.24) and P. fluorescens (7.23), as well as N. piscinale at a concentration of 1 g/L with A. lipoferum (7.26), indicating that the combined application of N. piscinale and PGPB slightly lowers the alkalinity level (Figure 2a). Moreover, during the second year, the combined use of N. piscinale at a concentration of 0.3 g/L in combination with A. lipoferum resulted in 20.25% increase in humus content compared with the control trails. Similarly, the application of N. piscinale at a concentration of 1 g/L along with treatments of A. lipoferum resulted in humus levels that were 17.54% higher compared with the untreated traits (Figure 2b). However, in both years, the untreated plots experienced a decrease in humus content compared to their initial levels. This decrease was likely attributed to various factors, such as microbial activity and decomposition processes within the soil.
During 2022, the analysis of bulk soil showed a noteworthy rise in (NO3− + NO2−)-nitrogen content within the treatments subjected to N. piscinale at a concentration of 0.3 g/L with A. lipoferum treatment (59.20%), N. piscinale at a concentration of 1 g/L along with treatments of A. lipoferum treatment (43.20%), and N. piscinale at a concentration of 1 g/L along with P. fluorescens treatment (21.82%), in comparison to the control group (Figure 3).
3.3. Microbial Activity of the Soil
Table 4 shows the activity of the bacteria and actinomycete population exhibited statistically significant differences in both years. Throughout the course of this study, the lowest levels of bacterial and actinomycete populations were consistently observed in the control group in both years.
Table 4.
Effect of the cyanobacteria-PGPB on activity of soil bacteria and actinomycete composition.
The results demonstrated that the highest bacterial biomass was recorded in N. piscinale at a concentration rate of 1 g/L when combined with A. lipoferum and P. fluorescens in the 2021 production season and in N. piscinale at a concentration rate of 0.3 g/L with A. lipoferum in the 2022 season.
Furthermore, the highest actinomycete count was observed in N. piscinale at a concentration rate of 1 g/L when combined with P. fluorescens in the 2021 season. Additionally, the highest actinomycete count was also recorded in N. piscinale at a concentration rate of 1 g/L when in association with A. lipoferum and P. fluorescens in the 2022 season.
4. Discussion
Z. mays holds a prominent position as one of the most frequently cultivated field crops on a global scale, being grown extensively across various regions of the world [55]. Z. mays requires a significant amount of nitrogen fertilizer, making it important to explore alternative fertilizer sources that offer benefits in agronomy, environmental sustainability, and economics [56,57]. Utilizing cyanobacteria and PGPB could serve as a viable alternative to enhance crop growth and yield in significant crops such as Z. mays, as these microorganisms have the capability to stimulate root system growth [58], enhance nutrient use efficiency (NUE) [59,60], and enhance the uptake of essential nutrients, including nitrogen (N) [38,61].
The current study evaluated the combined impact of cyanobacterial biomass and PGPB on Z. mays growth and soil fertility. The utilization of both cyanobacteria and PGPB in soil treatment resulted in a significant enhancement in the quantity of seeds number per ear, the thousand kernel weight, and overall crop yield while also enhancing soil properties including pH, humus, and (NO3− + NO2−)-N contents. However, the effectiveness of biofertilizer treatment on both plant growth and soil fertility is dependent on the concentration rate of N. piscinale and the PGPB strains used. Hence, the most effective approach was the joint use of N. piscinale and PGPB, followed by the single application of N. piscinale or PGPB, which also showed notable improvements compared with the control group.
The data clearly showed that using N. piscinale and PGPB alone/combination in the soil significantly boosted Z. mays growth, resulting in more seeds per ear, higher thousand kernel weight, and increased yield compared with the respective control. The improvement in various plant growth factors is likely a consequence of plants being better able to absorb essential nutrients from the soil. This enhanced nutrient uptake process makes vital nutrients more readily available to plants, supporting their overall growth and development. Furthermore, microbes have been reported to facilitate nutrient movement toward plant roots, and a substantial portion of soil microorganisms possess the capacity to improve plant nutrient uptake, offering eco-friendly strategies to address plant nutritional needs [62,63]. In earlier research, it has been demonstrated that applying cyanobacteria and PGPB, whether separately or together, leads to enhanced Z. mays growth and yield by either directly improving resource utilization and adjusting plant hormone levels or indirectly reducing the negative impact of various harmful agents [22,60,64].
The Azospirillum genus, consisting of free-living diazotrophs found in plant rhizospheres, is esteemed as a prime example of PGPB as a biofertilizer due to their beneficial influence on plant growth, crop yields, and nitrogen content [65]. In the present study, it was observed that the highest result in terms of the number of seeds per ear, seed weight, and yield was recorded at combined application N. piscinale at a concentration of 0.3 g/L with A. lipoferum (Table 3). However, the yield of Z. mays showed no statistical difference between the treatment using N. piscinale at 0.3 g/L combined with A. lipoferum and P. fluorescens, and the treatment with N. piscinale at 1 g/L combined with A. lipoferum.
The use of either N. piscinale or PGPB, as well as their combined application, resulted in a notable improvement in soil pH, humus content, and (NO3− + NO2−)-N levels. In both years, the untreated plots experienced a reduction in humus content compared to their initial levels. This decrease was likely attributed to various factors such as microbial activity and decomposition processes within the soil. Cyanobacterial growth improves soil properties, potentially aiding infertile soil reclamation when combined with PGPB, leading to enhanced plant growth, organic matter, improved soil fertility, nutrient utilization, and increased plant stress tolerance [10,11,27,66,67,68]. It may be due to the improvement of organic matter in the soil, which has had a positive impact on the soil’s physicochemical and biological properties. This enhancement of organic matter has been facilitated by beneficial microorganisms, including microalgae and bacteria, known for their ability to promote soil health and fertility [11,15,27,69,70]. In the current study, applying N. piscinale and PGPB either alone or in combination had no statistically significant impact on phosphorus and potassium levels. In contrast to this study, the research findings demonstrated that PGPB enhanced soil characteristics, elevating the availability of phosphorus and potassium content by a significant margin (100 and 70%), respectively, compared to the untreated soil [71].
In our current research, whether we applied N. piscinale or PGPB alone or in combination to the soil, we observed significant differences (p ≤ 0.05) in bacterial and actinomycete populations across both production years. Notably, the control group consistently exhibited the lowest levels of these populations throughout the study period. Cyanobacteria and PGPB play a pivotal role in regulating the abundance and functions of diverse soil microbial communities and enhancing plant growth [38,72]. The soil’s microbial communities in the rhizosphere play a crucial role in fostering eco-friendly agricultural practices, promoting sustainability, soil fertility, and ensuring agricultural productivity [73]. The incorporation of microbial biomass in the soil resulted in enhanced microbial diversity, changes in the abundance of organic matter-decomposing microorganisms, improved soil health, and the promotion of greater microbial variety [72,74].
5. Conclusions
This comprehensive study highlights the significant potential of utilizing cyanobacterial biomass and PGPB in enhancing Z. mays cultivation and improving soil fertility. Z. mays, a globally important crop, often relies heavily on nitrogen fertilizers, prompting the need for sustainable alternatives. The application of cyanobacterial biomass and PGPB, either individually or in combination, resulted in substantial improvements in Z. mays growth parameters, including seed quantity, seed weight, and overall yield, while also positively influencing key soil properties such as pH, humus content, and (NO3− + NO2−)-N levels. In this work, all strains displayed a high degree of compatibility for co-growth, however, the most optimal synergistic grouping were established by integrating both N. piscinale at a concentration of 0.3 g/L and A. lipoferum, resulting enhancing Z. mays yield and soil fertility. These findings support the role of microbial interventions in enhancing nutrient uptake by plants and promoting soil health. Moreover, the study highlights the potential of such eco-friendly strategies to address the nutritional needs of crops like Z. mays, offering promising avenues for sustainable agriculture and improved agricultural productivity.
Author Contributions
W.S. was responsible for the conceptualization, composition, and endorsement of the content, while L.M. and M.R. contributed to content creation and provided editorial support, and T.J. and Z.M. granted approval and assisted in editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project No. TKP2021-NKTA-06 funded by the Ministry of Innovation and Technology from the National Research Development and Innovation Fund, in the frame of the Thematic Excellence Program 2021.
Data Availability Statement
Data is contained within the article.
Acknowledgments
The authors extend their sincere appreciation to Giczi Zsolt and Takács Georgina for their invaluable assistance and support throughout the course of the experiment.
Conflicts of Interest
The authors do not have any pertinent financial or non-financial affiliations to declare.
References
- Singh, S.; Jagota, N.; Kaur, H.; Kaur, R.; Kaur, G.; Sandhu, S.; Sharma, A. Deciphering behavioral changes in maize plants in a quest to identify species specific plant growth promoting rhizobacteria. Total. Environ. Res. Themes 2023, 6, 100043. [Google Scholar] [CrossRef]
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009. [Google Scholar] [CrossRef]
- Poole, N.; Donovan, J.; Erenstein, O. Viewpoint: Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy 2021, 100, 101976. [Google Scholar] [CrossRef] [PubMed]
- Meena, R.S.; Kumar, S.; Datta, R.; Lal, R.; Vijayakumar, V.; Brtnicky, M.; Sharma, M.P.; Yadav, G.S.; Jhariya, M.K.; Jangir, C.K.; et al. Impact of agrochemicals on soil microbiota and management: A review. Land 2020, 9, 34. [Google Scholar] [CrossRef]
- Mahmud, K.; Missaoui, A.; Lee, K.; Ghimire, B.; Presley, H.W.; Makaju, S. Rhizosphere microbiome manipulation for sustainable crop production. Curr. Plant Biol. 2021, 27, 100210. [Google Scholar] [CrossRef]
- Sharma, S.; Kumawat, K.C. Role of rhizospheric microbiome in enhancing plant attributes and soil health for sustainable agriculture. In Core Microbiome; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2022; pp. 139–162. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Mohammed, M.; Ibny, F.Y.I.; Dakora, F.D. Rhizobia as a source of plant growth-promoting molecules: Potential applications and possible operational mechanisms. Front. Sustain. Food Syst. 2021, 4, 619676. [Google Scholar] [CrossRef]
- Chen, C.; Wang, M.; Zhu, J.; Tang, Y.; Zhang, H.; Zhao, Q.; Jing, M.; Chen, Y.; Xu, X.; Jiang, J.; et al. Long-term effect of epigenetic modification in plant–microbe interactions: Modification of DNA methylation induced by plant growth-promoting bacteria mediates promotion process. Microbiome 2022, 10, 36. [Google Scholar] [CrossRef]
- Khalil, A.T.; Shinwari, Z.K. Utilization of plant growth-promoting bacteria (pgpb) against phytopathogens. In Antifungal Metabolites of Rhizobacteria for Sustainable Agriculture; Sayyed, R.Z., Singh, A., Ilyas, N., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 53–63. [Google Scholar] [CrossRef]
- Eman, E.; Magdi, A.E.A.; Hassan, E.F. Perspective chapter: Cyanobacteria—A futuristic effective tool in sustainable agriculture. In Cyanobacteria; Archana, T., Ed.; IntechOpen: Rijeka, Croatia, 2023. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef]
- Abinandan, S.; Subashchandrabose, S.R.; Venkateswarlu, K.; Megharaj, M. Soil microalgae and cyanobacteria: The biotechnological potential in the maintenance of soil fertility and health. Crit. Rev. Biotechnol. 2019, 39, 981–998. [Google Scholar] [CrossRef]
- Vinoth, M.; Sivasankari, S.; Ahamed, A.K.K.; Al-Arjani, A.-B.F.; Abd_Allah, E.F.; Baskar, K. Biological soil crust (BSC) is an effective biofertilizer on Vigna mungo (L.). Saudi J. Biol. Sci. 2020, 27, 2325–2332. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Subramanian, J.; Gopalsamy, J.; Jayasingam, P.; Arumugam, A.; Kannadasan, S.; Sampathkumar, P. The impact of using microalgae as biofertilizer in maize (Zea mays L.). Waste Biomass- Valorization 2019, 10, 1101–1110. [Google Scholar] [CrossRef]
- Ramakrishnan, B.; Maddela, N.R.; Venkateswarlu, K.; Megharaj, M. Potential of microalgae and cyanobacteria to improve soil health and agricultural productivity: A critical view. Environ. Sci. Adv. 2023, 2, 586–611. [Google Scholar] [CrossRef]
- Glaser, K.; Albrecht, M.; Baumann, K.; Overmann, J.; Sikorski, J. Biological soil crust from mesic forests promote a specific bacteria community. Front. Microbiol. 2022, 13, 769767. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: A critical review of microalgae as renewable resources for agriculture. Algal Res. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Kang, Y.; Kim, M.; Shim, C.; Bae, S.; Jang, S. Potential of algae–bacteria synergistic effects on vegetable production. Front. Plant Sci. 2021, 12, 656662. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Bo, Y.; Feng, Y.; Tan, Y.; Zhou, C.; Yan, X.; Ruan, R.; Xu, Q.; Cheng, P. Potential applications for multifunctional microalgae in soil improvement. Front. Environ. Sci. 2022, 10, 1035332. [Google Scholar] [CrossRef]
- Lee, S.-M.; Ryu, C.-M. Algae as New Kids in the Beneficial Plant Microbiome. Front. Plant Sci. 2021, 12, 599742. [Google Scholar] [CrossRef]
- Berthon, J.-Y.; Michel, T.; Wauquier, A.; Joly, P.; Gerbore, J.; Filaire, E. Seaweed and microalgae as major actors of blue biotechnology to achieve plant stimulation and pest and pathogen biocontrol—A review of the latest advances and future prospects. J. Agric. Sci. 2021, 159, 523–534. [Google Scholar] [CrossRef]
- Reed, L.; Glick, B.R. The recent use of plant-growth-promoting bacteria to promote the growth of agricultural food crops. Agriculture 2023, 13, 1089. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Tavasolee, A.; Ghassemi-Golezani, K.; Torabian, S.; Monirifar, H.; Rahmani, H.A. Growth-promoting bacteria and natural regulators mitigate salt toxicity and improve rapeseed plant performance. Protoplasma 2020, 257, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Barua, N.; Clouse, K.M.; Diaz, D.A.R.; Wagner, M.R.; Platt, T.G.; Hansen, R.R. Screening the maize rhizobiome for consortia that improve Azospirillum brasilense root colonization and plant growth outcomes. Front. Sustain. Food Syst. 2023, 7, 1106528. [Google Scholar] [CrossRef]
- Khan, M.Y.; Nadeem, S.M.; Sohaib, M.; Waqas, M.R.; Alotaibi, F.; Ali, L.; Zahir, Z.A.; Al-Barakah, F.N.I. Potential of plant growth promoting bacterial consortium for improving the growth and yield of wheat under saline conditions. Front. Microbiol. 2022, 13, 958522. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mei, S.; Salles, J.F. Inoculated microbial consortia perform better than single strains in living soil: A meta-analysis. Appl. Soil Ecol. 2023, 190, 105011. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, L.M.; De-Bashan, L.E. The potential of microalgae–bacteria consortia to restore degraded soils. Biology 2023, 12, 693. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-Del-Valle, M.; Vílchez, C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef]
- González-González, L.M.; De-Bashan, L.E. Toward the enhancement of microalgal metabolite production through microalgae–bacteria consortia. Biology 2021, 10, 282. [Google Scholar] [CrossRef]
- Solomon, W.; Mutum, L.; Janda, T.; Molnár, Z. Potential benefit of microalgae and their interaction with bacteria to sustainable crop production. Plant Growth Regul. 2023, 101, 53–65. [Google Scholar] [CrossRef]
- Amin, S.A.; Hmelo, L.R.; Van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef]
- Kim, B.-H.; Ramanan, R.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Role of rhizobium, a plant growth promoting bacterium, in enhancing algal biomass through mutualistic interaction. Biomass- Bioenergy 2014, 69, 95–105. [Google Scholar] [CrossRef]
- Bunbury, F.; Deery, E.; Sayer, A.P.; Bhardwaj, V.; Harrison, E.L.; Warren, M.J.; Smith, A.G. Exploring the onset of B12 -based mutualisms using a recently evolved Chlamydomonas auxotroph and B12-producing bacteria. Environ. Microbiol. 2022, 24, 3134–3147. [Google Scholar] [CrossRef]
- Smith, M.J.; Francis, M.B. A Designed A. vinelandii–S. elongatus coculture for chemical photoproduction from air, water, phosphate, and trace metals. ACS Synth. Biol. 2016, 5, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Chittora, D.; Meena, M.; Barupal, T.; Swapnil, P.; Sharma, K. Cyanobacteria as a source of biofertilizers for sustainable agriculture. Biochem. Biophys. Rep. 2020, 22, 100737. [Google Scholar] [CrossRef]
- Prasanna, R.; Hossain, F.; Babu, S.; Bidyarani, N.; Adak, A.; Verma, S.; Shivay, Y.S.; Nain, L. Prospecting cyanobacterial formulations as plant-growth-promoting agents for maize hybrids. S. Afr. J. Plant Soil 2015, 32, 199–207. [Google Scholar] [CrossRef]
- Ördög, V.; Stirk, W.; Takács, G.; Pőthe, P.; Illés, A..; Bojtor, C.; Széles, A.; Tóth, B.; van Staden, J.; Nagy, J. Plant biostimulating effects of the cyanobacterium Nostoc piscinale on maize (Zea mays L.) in field experiments. S. Afr. J. Bot. 2021, 140, 153–160. [Google Scholar] [CrossRef]
- Sharma, V.; Prasanna, R.; Hossain, F.; Muthusamy, V.; Nain, L.; Das, S.; Shivay, Y.S.; Kumar, A. Priming maize seeds with cyanobacteria enhances seed vigour and plant growth in elite maize inbreds. 3 Biotech 2020, 10, 154. [Google Scholar] [CrossRef] [PubMed]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643. [Google Scholar] [CrossRef] [PubMed]
- Florio, A.; Pommier, T.; Gervaix, J.; Bérard, A.; Le Roux, X. Soil C and N statuses determine the effect of maize inoculation by plant growth-promoting rhizobacteria on nitrifying and denitrifying communities. Sci. Rep. 2017, 7, 8411. [Google Scholar] [CrossRef] [PubMed]
- Walker, V.; Couillerot, O.; Von Felten, A.; Bellvert, F.; Jansa, J.; Maurhofer, M.; Bally, R.; Moënne-Loccoz, Y.; Comte, G. Variation of secondary metabolite levels in maize seedling roots induced by inoculation with Azospirillum, Pseudomonas and Glomus consortium under field conditions. Plant Soil 2012, 356, 151–163. [Google Scholar] [CrossRef]
- Mohanty, P.; Singh, P.K.; Chakraborty, D.; Mishra, S.; Pattnaik, R. Insight into the role of pgpr in sustainable agriculture and environment. Front. Sustain. Food Syst. 2021, 5, 667150. [Google Scholar] [CrossRef]
- Jiang, S.; Li, J.; Wang, Q.; Yin, C.; Zhan, Y.; Yan, Y.; Lin, M.; Ke, X. Maize growth promotion by inoculation with an engineered ammonium-excreting strain of nitrogen-fixing pseudomonasstutzeri. Microorganisms 2022, 10, 1986. [Google Scholar] [CrossRef]
- Ördög, V. Apparatus for Laboratory Algal Bioassay. Int. Revue ges. Hydrobiol. 1982, 67, 127–136. [Google Scholar]
- Takács, G.; Stirk, W.; Gergely, I.; Molnár, Z.; van Staden, J.; Ördög, V. Biostimulating effects of the cyanobacterium Nostoc piscinale on winter wheat in field experiments. S. Afr. J. Bot. 2019, 126, 99–106. [Google Scholar] [CrossRef]
- Tamiya, H. Mass culture of algae. Annu. Rev. Plant Physiol. 1957, 8, 309–334. [Google Scholar] [CrossRef]
- Pansu, M.A.; Pansu, M.B. Handbook of soil analysis of the chapter. In Book Handbook of Soil Analysis; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- ISO 13395:1996; Water Quality—Determination of Nitrite Nitrogen and Nitrate Nitrogen and the Sum of Both by Flow Analysis (CFA and FIA) and Spectrometric Detection. ISO Central Secretariat: Geneva, Switzerland, 1996. Available online: https://infostore.saiglobal.com/en-gb/standards/iso-13395-1996-613022_saig_iso_iso_1406298/ (accessed on 20 August 2023).
- Clark, F.E. Agar-plate method for total microbial count. In Methods of Soil Analysis; Chemical and Microbiological Properties: Madson, NY, USA, 1965; pp. 1460–1466. [Google Scholar] [CrossRef]
- Olsen, R.A.; Bakken, L.R. Viability of soil bacteria: Optimization of plate-counting technique and comparison between total counts and plate counts within different size groups. Microb. Ecol. 1987, 13, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.T.; Goodfellow, M.; Alderson, G.; Wellington, E.M.H.; Sneath, P.H.A.; Sackin, M.J. Numerical classification of streptomyces and related genera. Microbiology 1983, 129, 1743–1813. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M. Most-probable-number method for microbial populations. Methods Soil Anal. Part 2 Chem. Microbiol. Prop. 1965, 9, 1467–1472. [Google Scholar] [CrossRef]
- R Core Team, R. R: A language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Mendiburu, F.E.D. Agricolae: Statistical Procedures for Agricultural Research. 2023, (Version 1.3-7). Available online: https://cran.r-project.org/web/packages/agricolae/agricolae.pdf (accessed on 24 October 2023).
- FAO. World Food and Agriculture—Statistical Yearbook 2022; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2022. [Google Scholar]
- Alves, D.K.M.; Teixeira, M.B.; Cunha, F.N.; Filho, F.R.C.; Cunha, G.N.; Andrade, C.L.L.D. Grain yield of maize crops under nitrogen fertigation using wastewater from swine and fish farming. Agronomy 2023, 13, 1834. [Google Scholar] [CrossRef]
- Hungria, M.; Barbosa, J.Z.; Rondina, A.B.L.; Nogueira, M.A. Improving maize sustainability with partial replacement of N fertilizers by inoculation with Azospirillum brasilense. Agron. J. 2022, 114, 2969–2980. [Google Scholar] [CrossRef]
- Li, H.; Zhao, Y.; Jiang, X. Seed soaking with bacillus sp. Strain hx-2 alleviates negative effects of drought stress on maize seedlings. Chil. J. Agric. Res. 2019, 79, 396–404. [Google Scholar] [CrossRef]
- Pandey, C.; Dheeman, S.; Prabha, D.; Negi, Y.K.; Maheshwari, D.K. Plant growth-promoting bacteria: Effective tools for increasing nutrient use efficiency and yield of crops. In Endophytes: Mineral Nutrient Management; Maheshwari, D.K., Dheeman, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; Volume 3, pp. 293–313. [Google Scholar] [CrossRef]
- Di Benedetto, N.A.; Corbo, M.R.; Campaniello, D.; Cataldi, M.P.; Bevilacqua, A.; Sinigaglia, M.; Flagella, Z. The role of plant growth promoting bacteria in improving nitrogen use efficiency for sustainable crop production: A focus on wheat. AIMS Microbiol. 2017, 3, 413–434. [Google Scholar] [CrossRef]
- Múnera-Porras, L.M.; García-Londoño, S.; Ríos-Osorio, L.A. Ríos-Osorio. Action mechanisms of plant growth promoting cyanobacteria in crops in situ: A systematic review of literature. Int. J. Agron. 2020, 2020, 2690410. [Google Scholar] [CrossRef]
- Singh, S.K.; Wu, X.; Shao, C.; Zhang, H. Microbial enhancement of plant nutrient acquisition. Stress Biol. 2022, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant Sci. 2015, 6, 815. [Google Scholar] [CrossRef] [PubMed]
- Gavilanes, F.Z.; Andrade, D.S.; Zucareli, C.; Horácio, E.H.; Yunes, J.S.; Barbosa, A.P.; Alves, L.A.R.; Cruzatty, L.G.; Maddela, N.R.; de Fátima Guimarães, M. Co-inoculation of Anabaena cylindrica with Azospirillum brasilense increases grain yield of maize hybrids. Rhizosphere 2020, 15, 100224. [Google Scholar] [CrossRef]
- Vuolo, F.; Novello, G.; Bona, E.; Gorrasi, S.; Gamalero, E. Impact of plant-beneficial bacterial inocula on the resident bacteriome: Current knowledge and future perspectives. Microorganisms 2022, 10, 2462. [Google Scholar] [CrossRef]
- Singh, J.S. Cyanobacteria: A vital bio-agent in eco-restoration of degraded lands and sustainable agriculture. Clim. Change Environ. Sustain. 2014, 2, 133–137. [Google Scholar]
- Mutale-Joan, C.; Sbabou, L.; Hicham, E.A. Microalgae and cyanobacteria: How exploiting these microbial resources can address the underlying challenges related to food sources and sustainable agriculture: A review. J. Plant Growth Regul. 2023, 42, 1–20. [Google Scholar] [CrossRef]
- Prasanna, R.; Hossain, F.; Saxena, G.; Singh, B.; Kanchan, A.; Simranjit, K.; Ramakrishnan, B.; Ranjan, K.; Muthusamy, V.; Shivay, Y.S. Analyses of genetic variability and genotype x cyanobacteria interactions in biofortified maize (Zea mays L.) for their responses to plant growth and physiological attributes. Eur. J. Agron. 2021, 130, 126343. [Google Scholar] [CrossRef]
- Mutum, L.; Janda, T.; Ördög, V.; Molnár, Z. Biologia Futura: Potential of different forms of microalgae for soil improvement. Biol. Futur. 2022, 73, 1–8. [Google Scholar] [CrossRef]
- Kumar, S.; Diksha; Sindhu, S.S.; Kumar, R. Biofertilizers: An ecofriendly technology for nutrient recycling and environmental sustainability. Curr. Res. Microb. Sci. 2022, 3, 100094. [Google Scholar] [CrossRef]
- Schoebitz, M.; Mengual, C.; Roldán, A. Combined effects of clay immobilized Azospirillum brasilense and Pantoea dispersa and organic olive residue on plant performance and soil properties in the revegetation of a semiarid area. Sci. Total. Environ. 2014, 466–467, 67–73. [Google Scholar] [CrossRef]
- Ranjan, K.; Priya, H.; Ramakrishnan, B.; Prasanna, R.; Venkatachalam, S.; Thapa, S.; Tiwari, R.; Nain, L.; Singh, R.; Shivay, Y.S. Cyanobacterial inoculation modifies the rhizosphere microbiome of rice planted to a tropical alluvial soil. Appl. Soil Ecol. 2016, 108, 195–203. [Google Scholar] [CrossRef]
- Uzoh, I.M.; Babalola, O.O. Rhizosphere biodiversity as a premise for application in bio-economy. Agric. Ecosyst. Environ. 2018, 265, 524–534. [Google Scholar] [CrossRef]
- Alobwede, E.; Cotton, A.; Leake, J.R.; Pandhal, J. The fate and distribution of microalgal nitrogen when applied as an agricultural soil fertiliser and its effect on soil microbial communities. Phycology 2022, 2, 297–318. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).