Abstract
The increasing diversity of potential biosecurity threats makes their diagnosis a complicated and evolving area, requiring moving beyond traditional taxonomic species identification. New biosecurity diagnostic tools should provide a greater depth of information on threat biology to enable accurate risk assessment for the more efficient and effective deployment of biosecurity resources. Metabolomics is amongst the new approaches being explored for biosecurity diagnostics, where a broad spectrum of metabolites might signify relevant biological characteristics of an intercepted organism. Examples of these characteristics are physiological signatures of age, reproductive status, geographic origin, pathogen status of potential invertebrate vectors, and the distinction between diseases and abiotic plant stress symptoms. This broad-based approach is attractive, where several biological characteristics of an organism can be assessed with a single measurement. However, it can be impractical as several hundred biological replicates of the organism are needed to build a robust model of a species. New approaches such as mass spectral fingerprinting substantially reduce the time taken for metabolomics measurements, and more sophisticated modelling methods aid feasibility. Promising data are emerging for metabolomics and metabolite fingerprinting for potential biosecurity applications. We discuss the possibilities and potential uses for these new tools for post-detection decisions, diagnosis, and biosecurity.
1. Introduction
The Evolving Role of Diagnostics for Biosecurity
Biosecurity, broadly considered, aims to keep exotic organisms from invading or establishing in a geographical region to protect its environment, economy, health, social, and cultural values. This can include preventing the demise of indigenous species or the damage or destruction of valuable crops and livestock. Against a background of increasing numbers and diversity of insect pest and pathogen threats, there is growing pressure on biosecurity systems, ranging from implementing policies to risk assessment, eradication, and management [1]. Amongst the practical components, diagnostics is a cornerstone activity for post-detection decisions that support the most effective actions for eradication or management. Traditionally, diagnostics has been limited to taxonomically based species identification. Lately, this has become more dependent on molecular tools, from standard DNA barcoding [2] to high-throughput sequencing [3] and portable in-field solutions [4], supported by biochemical and serological tests and pathogenicity assays. These have been central to improving species-, population-, and subspecies-level identification. However, there are still occasions important to biosecurity where genetic markers are either unreliable [5,6] or are not useful for questions beyond taxonomic identification owing to inherited molecular sequences not changing in response to the environment. Over the last decade, alternative methods that extend the application of diagnostics have been emerging, but are not yet common (e.g., Holder, Van Hale et al. [7]). One tool that could complement or rationalise the existing methods is metabolomics [8] (Figure 1). The potential for metabolomic approaches in the diagnosis of human disease is well established [9], and it is also an emerging tool for animal [10] and plant disease diagnoses [11]. Additionally, the possibilities to apply metabolomics for diagnostics in biosecurity are now being considered in light of initial studies and work in areas allied to biosecurity, such as entomology and plant pathology, with examples of its potential application examined more closely below.
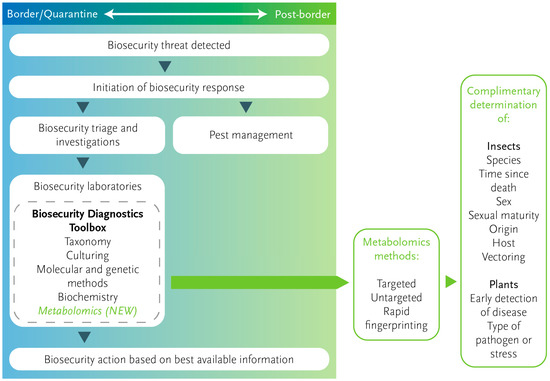
Figure 1.
Metabolomics in the context of a biosecurity response. Metabolomics could be included as part of the biosecurity diagnostics ‘toolbox’ to complement existing diagnostic methods. Figure adapted from the New Zealand Ministry for Primary Industries.
2. Metabolomics
2.1. An Overview of Metabolomics
Metabolomics uses analytical chemistry tools to measure a wide range of small molecules derived from metabolic processes (usually defined as less than 1500 Daltons) to understand biological systems [12]. This includes how biochemical pathways respond to treatments or differ between species and the discovery of novel metabolites that can serve as biomarkers of specific biological situations. Metabolomics is ideally suited for hypothesis-generating research where specific molecular targets concerning a scientific question may not be known. For example, two different plant pathogens may cause different symptoms in a plant, but the molecular basis for this difference is unknown. In that regard, it is a powerful opportunity to discover non-traditional markers of biological characteristics that are useful for biosecurity.
Metabolites are at the end of the genes–mRNA–proteins–posttranslational modification–metabolite cascade, reflecting the interaction between genetics, the environment, and the resulting phenotype [13]. Primary metabolites such as intermediates in energy metabolism, amino acids, and fatty acids fill a variety of roles in biology: structural and building blocks for other biomolecules, enzyme cofactors and substrates, gene expression regulators, energy metabolism, and waste products. In plants, there are also thousands of secondary metabolites not essential for growth and development but that fulfil a wide range of biological functions, including defence and intra- or inter-species communication. These include volatile molecules and polyphenols. Estimates are scant of the size of the metabolome (all measurable metabolites in an organism, potentially including exogenous metabolites) of biosecurity-relevant species. There are thousands of metabolites for some agriculture-relevant plants, such as kiwifruit (4062), potato (4470), and tomato (4194), included in the food metabolome database (www.foodb.ca, accessed on 23 May 2022). For comparison, the human metabolome database contains 217,920 entries [14], suggesting that the actual size of plant metabolomes is much higher (although the human metabolome database also includes compounds of plant and synthetic origin if they are detected in human samples). In practice, the number of metabolites detected and identified with any method is between 50 and 1000, depending on the sample and methods used to measure the metabolites.
Both genetic and external factors influence metabolite production and composition in a sample, and metabolome measurement of a given sample may include several different metabolomes. For example, an insect metabolome will consist of its own metabolome, that of its gut microbiome or parasites, its external microbiome, and also metabolites from the metabolome of its host plant. The metabolites from exogenous sources, including food compounds, pesticides, pollutants, and microbial compounds, are collectively referred to as the ‘exposome’ [15]. This high degree of complexity and heterogeneity presents several challenges. However, it also means that the relative ratios of metabolites can be specific for certain situations, including species and subspecies, host-plant association, and geographic origin (Figure 2).
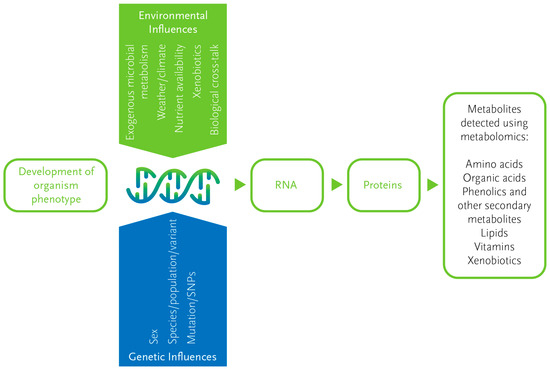
Figure 2.
Influences on the metabolome. Metabolites reflect the interaction between genetics and the environment, and together with proteins, reflect the resulting phenotype of an organism. This makes the metabolome highly responsive to both genetic and environmental changes. SNP = single nucleotide polymorphism.
Estimates of metabolomic heritability—the proportion of the metabolome that is stable from generation to generation—vary, with 5–47% based on genome-wide association studies of metabolite genes in tomatoes and rice, suggesting a significant impact of the environment on the expression of metabolites in plants [16]. There is also considerable heterogeneity between different plant species, with a single genetic locus explaining up to 90% of the observed metabolite variation in Arabidopsis thaliana and in rice, but at most 62% in maize [16]. Metabolomic patterns are complicated because metabolites are often not unique to one species or pathway. For example, amino acids are present in all biological samples, and their concentrations are regulated by several pathways, which limits the utility of single amino acids as biomarkers in many cases. This high degree of complexity is challenging, but can be harnessed with the chemometric (i.e., data-driven analysis of chemical information) and bioinformatic advances over the past decade for processing and understanding metabolomics data [17] to gain further knowledge of the organism or system of interest.
2.2. Metabolomics Methodologies
Nuclear magnetic resonance spectroscopy (NMR) and mass spectrometry (MS) are the main technologies used to measure the metabolome. NMR detection of metabolites is based on how the nuclei of certain atoms (e.g., hydrogen or carbon-13) resonate under an electromagnetic field. In metabolomics, the different resonances, or chemical shifts, are used to identify metabolites in complex mixtures. NMR usually requires minimal sample preparation and can be performed on solid samples (‘solid-state NMR’). It has an advantage in being quantitative (i.e., the strength of a signal is directly proportional to the number of molecules responsible for the signal). The main limitations to the wider use of NMR for metabolomics are its sensitivity relative to mass spectrometry, size, and expense, although smaller and cheaper instruments are available, with the trade-off of less sensitivity and resolution [18].
Alternatively, MS measures the mass of charged molecules. It uses a combination of the molecular weight (based on the mass of the charged molecule ratio, or m/z) and the fragmentation pattern of a molecule for identification. Usually, for metabolomics, mass spectrometers are coupled to chromatography to separate metabolites prior to their analysis to avoid overwhelming the ionisation process, where a charge is added to the metabolites before detection (a phenomenon known as ‘ion suppression’). The time taken for a metabolite to pass through a chromatography column (‘retention time’) is also used to aid compound identification. Gas chromatography (GC) and liquid chromatography (LC) are the main types of chromatography used. However, other separation methods, such as capillary electrophoresis or supercritical CO2 chromatography, are also used.
Mass spectrometry is more sensitive than NMR and can detect greater numbers of metabolites. While still expensive, it is cheaper and usually requires less laboratory infrastructure than NMR. The main limitations are that it is not quantitative without the use of standards (not all molecules ionise with the same intensity), and samples need to be extracted before analysis, which is time-consuming, especially for solid samples, and can add artefacts into the analyses [19]. As with all limitations, research in the field is on-going to mitigate these problems and improve the throughput and quality of metabolomics analyses.
The emerging use of ambient ionisation mass spectrometry (AI-MS) has relevance and potential for biosecurity diagnostics. AI-MS refers to a mass spectrometer with an interface that allows a sample to be measured without any or with minimal sample preparation; it produces a mass spectral ‘fingerprint’ within a few seconds of analysis. Many types of AI-MS work according to different principles of introducing samples to a mass spectrometer. Among these are rapid evaporative ionisation mass spectrometry (REIMS) [20,21] and direct analysis in real-time mass spectrometry (DART-MS) [22]. AI-MS fills a gap for rapid sample preparation and analysis that is not readily met by other instruments used for metabolomics (Figure 3). The main drawback of AI-MS is its limited ability to identify the molecules of interest. This is problematic in cases where understanding the biology of a sample is the primary focus, but for diagnostics, only a robust fingerprint of the metabolites present is required [23]. Tentative identification of metabolites can be made using AI-MS, depending on the mass spectrometer type, for example, high-resolution mass and mass fragmentation.
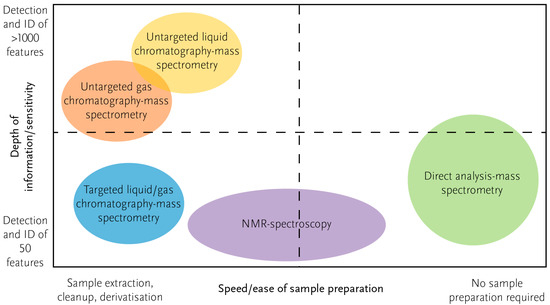
Figure 3.
Different methods for metabolomics have different strengths and weaknesses. With current methods there is a general trend that greater depth of information (e.g., more metabolites detected) requires more sample preparation. Other factors also need to be considered, such as ease of use, initial investment and maintenance costs, and depth of information required in relation to the application (e.g., scientific research versus diagnostic screening).
Along with instrumentation, data analysis and interpretation are critical for metabolomics. While outside the scope of this review, as in all areas of science, new data analysis tools are improving the use of the large amounts of data generated using metabolomics. As for all fields of measurement science, these improvements will make data analysis and interpretation more profound, simpler, and faster, further increasing the utility of metabolomics.
3. Exploring Metabolomics for Plant Health Biosecurity
Here we consider various characteristics of insect pests and plant pathogens that can help to describe their biosecurity risk, including and beyond species identification, to inform critical biosecurity responses. Based on the application of metabolomics to diagnostics elsewhere and on fundamental research and insights from the use of this technology in insect and plant pathogen studies, we indicate how this methodology could contribute to expanding the current role of diagnostics for plant health. This includes both comparative fingerprinting for diagnostics and for metabolite profiling to discover new associations between metabolites and phenotypes relevant to biosecurity. We have broadly divided this into two sections, one related to traits of interest to insect biosecurity threats, and the second to the detection of plant pathogens, including vectored pathogens.
3.1. Metabolomics Applied to Insect Traits Relevant to Biosecurity
3.1.1. Entometabolomics
The application of metabolomics to insects is termed entometabolomics [24], and a range of metabolite-based investigations, described below, illustrate how this can be translated for potential use in biosecurity. Amongst the key characteristics of interest in plant biosecurity are not only the invasive insect’s species, sex, age, and sexual maturity but also whether they are vectoring pathogenic microorganisms or not [24], if they are sterile (through post-harvest treatment or produced for the sterile insect technique (SIT) for eradication) or not, and, in the case of sessile species or life stages, whether they are dead or alive [25]. Each of these traits might provide insights into the invasive ability of a given insect, the status of the associated population if established, and the potential risk of damage to the invading area. This, in turn, could help direct responses regarding geographic surveillance needs and design, host plants at risk, and any need for eradication (Figure 1).
For any diagnostic test, irrespective of the questions being asked, one of the main challenges is the low amounts of insect, pathogen, or plant biomass available, particularly if multiple separate tests and technologies are required. Low biomass may be due to only one or very few small specimens being detected at the outset, so that pooling specimens is not an option, or to immature life stages or traces such as pupal casings or frass (i.e., insect faeces) being the only samples available. Destructive pre-test preparation prevents the use of other tests, especially if retaining the specimen as a morphological voucher is needed. Analysis of less than 1 mg of tissue presents a challenge for most existing analytical methods, and most studies have resorted to pooling insects or body parts together for analysis [24]. This could have negative consequences in the case, for instance, of the identification of cryptic species complexes. As instruments become more sensitive and better able to detect metabolites from low amounts of sample, one of the main challenges becomes practical: very small samples are difficult to sample reproducibly and can be easily blown about by laboratory airflows.
Cuticular hydrocarbons (CHCs) are a compound class present in the insect metabolome, originally evolved for the prevention of desiccation and as signalling molecules for insect chemical communication [26]. CHCs allow the identification of sex, species, and colony, among other traits, and this has consequently made the measurement of CHCs a popular method in insect biology and chemical ecology studies [27,28,29]. Gas chromatography coupled with mass spectrometry (GC-MS) is the standard analytical instrument for CHC analysis. GC-MS analysis of CHCs requires substantial sample preparation using organic solvents for extraction, often requiring a whole insect for analysis [22,30]. Some insects might retain their body shape and integrity after extraction for CHC analysis, but others do not, which can be impractical for biosecurity purposes. GC-MS analysis of CHCs also restricts the chain length detected to 40 carbon atoms [22,31].
A potentially non-destructive sample for metabolomics as an alternative to hydrocarbon analysis is the haemolymph. Haemolymph is the equivalent of vertebrate blood and circulates nutrients, signalling molecules, and waste products between different organs within an insect. Analytically, haemolymph can be seen as a pool of metabolites that reflects all organs and cells of the insect [32,33]. As haemolymph is present throughout the whole insect body, small aliquots can be taken without mutilating the entire insect. Samples can be drained from a leg or antenna or taken from an abdominal puncture [34], although it requires considerable skill. With haemolymph collection, the insect remains primarily intact for further dissection, preservation, or analyses. This provides a clear advantage over using the entire or part of an insect, especially for subsequent taxonomic analysis [33,35]. Studies have used haemolymph to detect differences between developmental stages and parasitized compared with healthy insects using GC-MS [32,33]. However, further work on insect haemolymph as a substrate for metabolomics is required, especially in handling the limited sample volumes available from haemolymph sampling.
Finally, the use of ether or ethanol to preserve insects or insects from desiccated collections as sources of samples to create reference ‘metabolomes’ also needs to be accounted for, as these types of specimens are often available for biosecurity purposes. Still, the preservative will invariably have an impact on the metabolites present in the insects. In other areas of biological science where metabolomics has been applied, there are strong recommendations for the standardisation of specimen collection and storage [36,37], and this is likely to be necessary if metabolomics is to be used for the routine analysis of biosecurity samples as well.
3.1.2. Species Identification
Species identification is arguably the most critical characteristic determining the type and timing of a biosecurity response. It underpins the assessed risk posed to health, agriculture, and the environment at the border [38]. Insect identification typically relies on either traditional morphological taxonomic characteristics, requiring increasingly rare advanced entomological knowledge and skills, or on molecular markers and laboratory plus bioinformatic skills. However, morphological or molecular markers are not always reliably distinctive. They can even be impractical, especially with taxonomically cryptic species (i.e., taxa that are morphologically indistinguishable but biologically different). Some species are endemic or exotic but not a pest, while others are a biosecurity threat. Entometabolomics as an alternative source of markers for species identification has been effective in species separation, showing great promise as an approach that could overcome some of the traditional hurdles to differentiating between cryptic species. For example, females within the Ceratitis (Diptera: Tephritadae) FAR complex are difficult to distinguish morphologically, and using 16 microsatellite loci is highly impractical and too expensive for routine biosecurity applications. Using two-dimensional gas chromatography with time-of-flight mass spectrometry, the CHC profiles of the genotypic clusters representing these species were significantly different, with the main diagnostic compounds being methyl-branched alkanes, alkenes, and alkatrienes [39]. This was consistent with branched and unsaturated alkanes being demonstrated elsewhere as useful chemotaxonomical markers in several dipteran species [40].
Screening of populations of the economically significant subterranean Reticulitermes (Blattodea: Rhinotermitidae) termites also revealed markedly different GC-MS CHC profiles. This led to the discovery of mitochondrial DNA clades correlated with important phenotypic differences in pest management, including geography, flight times, and aggressiveness, and even leading the authors to propose new undescribed species [41], showing that metabolites can inform genetics.
Developing species markers on a large scale has also been nicely illustrated in an animal biosecurity example. There are many barriers to accurately identifying sarcophagid flesh flies (Diptera: Sarcophagidae): females are often impossible to distinguish morphologically, genetic introgression has been reported, and DNA from key historical specimens cannot be retrieved. Using high-throughput GC-MS to measure the CHC profiles of 185 specimens, including some over 100 years old, 11 species and the sexes within species could be reliably differentiated [42]. Another study, undertaken using DART-MS, attempted to identify species at cryptic life stages (e.g., larvae) of various carrion (blow) flies (Diptera: Calliphoridae) [22]. Using an artificial neural network, it was possible to distinguish larvae from different species despite their being visually identical. Although this can also be determined using other molecular methods, the metabolic fingerprinting methodology represents a rapid and facile approach for identifying dipteran larvae in field-derived samples.
The key for diagnosticians is the knowledge that species taxonomy is reliable. In the case of the economically important Australian genus of sympatric weevils Gonipterus (Coleoptera: Curculionidae), the accurate identification of species, especially within the cryptic G. scutellatus complex where not all species are invasive in other countries, is extremely difficult: morphologically based species identification is possible only with destructive dissection of male genitalia or using DNA barcoding [31]. Using GC-MS, species’ CHC profiles were all shown to be significantly different, allowing successful species separation [31].
Mass spectral fingerprinting using REIMS, an AI-MS instrument, is another high-throughput analytical option that has also shown promise for insect species identification. A study using five Drosophila (Diptera: Drosophilidae) species demonstrated that an informative spectral fingerprint can be produced for each species and that the data generated convincingly resolved the Drosophila into clear species clusters [43]. REIMS was also able to separate morphological and molecular cryptic species in the Anopheles gambiae (Diptera: Culicidae) mosquito species complex [44]: Anopheles coluzzi, A. gambiae, and Anopheles arabiensis. Here, using approximately 200 specimens, the REIMS spectra were visually similar by eye, but clear differences were resolved using machine learning [45].
The results using CHC analysis and fingerprinting for otherwise difficult species identifications are very encouraging. However, destroying all or part of a sample to conduct the analysis is still a requirement, which creates a potential conflict, as outlined above, when a specimen needs to be vouchered or used for multiple assays. As an advance on those studies, we have recently performed analyses on individual weevil legs using a modified REIMS source for improved signal intensity [46], which resulted in a fingerprint that enabled differentiation between species (unpublished observations). This suggests that the destruction of a specimen is not necessary for many entometabolomic applications.
3.1.3. Sex Differentiation
Post-border detection of a high-risk species is the primary determinant of a biosecurity response plan, which may also be influenced by the sex of the species, as females of an exotic species may be able to establish founder populations in the invaded regions. Many species have sexually dimorphic traits, making this relatively straightforward to determine. However, the potential of a single measurement using, for example, fingerprinting mass spectrometry to identify multiple characteristics from a single measure, including sex, is advantageous when time and samples are limited, such as during an incursion [30,45]. Useful examples of metabolomics with minute species include the use of silver-assisted laser desorption/ionisation MS and DART-MS to distinguish the sexes of 48 h old parasitoid jewel wasps (Nasonia vitripennis) (Hymenoptera: Pteromalidae) based on their CHC profiles [30] and REIMS to differentiate sexes in marsh mosquitos (Anopheles spp.) [45] and across different Drosophila species (Diptera: Drosophilidae) [43,45].
3.1.4. Age and Sexual Maturity
Knowledge of the age of an insect can be used to trace back the likely timeline of incursion, including when insects are caught in traps, especially for traps that are not checked frequently. Age is also correlated with sexual maturity, which indicates the likelihood that a female has mated and possibly laid eggs, with apparent implications for biosecurity risk assessment.
Some CHCs function as contact pheromones and are involved in inter-individual communication [47]. Changes to the CHC phenotype often correlate with sexual maturation, including attractiveness [31]. Thus, metabolite profiles would be expected to reflect relevant physiological information (e.g., hormone or gene up-/down-regulation) related to an insect sample. Using laser-MS technologies for detecting longer-chain CHCs, age-specific profiles have been determined, for example, in D. melanogaster, to shift from short- to longer-chain-length CHCs responsible for a significant reduction in sexual attractiveness as the insects age [48]. Similarly, age-specific profiles, including novel, very-long-chain CHCs, were detected with the same MS instruments and confirmed with DART-MS on parasitoid wasps and mosquitos [30,45].
The discovery of other non-CHC age-related metabolites further builds the application of metabolomics to determine age and sexual maturity status in a given sample. For example, the different instars (moulting stages) of the silkworm (Bombyx mori) (Lepidoptera: Bombycidae) were determined from haemolymph samples using GC-MS- and NMR-based metabolomics, and changes across instars were related to differences in circulating carbohydrate type, as related to biochemical changes that occur with age [33]. Similarly, Kapranas et al. [49] showed with NMR spectra of low-biomass samples (1 mg) from individual parasitoid wasps that changes in glycerolipids, amino acids, and circulatory sugars between 0- and 7-day-old wasps were correlated with age. However, the authors emphasised that the metabolomics of aging will differ among insect species according to their life histories. The observed trends for parasitoids are linked to prolonged starvation, as they cannot generate lipid energy stores after adult eclosion owing to their parasitic lifestyle. However, for non-parasitoids, a post-eclosion diet can significantly alter the metabolomics of aging [49]. REIMS fingerprints could also clearly differentiate between mosquitos aged 0, 1–2, and 12–13 days, with the potential to differentiate age differences as small as 24 h. However, on some days, there was an overlap in fingerprints [45]. Of note with the latter, age and sex could be determined from the same fingerprint with an accuracy of 84 and 91%, respectively [45]. This supports the idea that a single measurement could be used to obtain information about several biological aspects of the same sample.
3.1.5. Time since Death and Storage
Estimating the time since the death of a specimen could help us understand how long a potentially undiscovered population may have been in the locale before the detection of an exotic species was made, for example, in a trap. Little entometabolomic research directly addresses determining the time of death in insects. However, from a recent study on the impact of different storage conditions over time, REIMS fingerprints of D. melanogaster and Anopheles species showed a deterioration of metabolic features at high masses (m/z 600–800, generally phospholipids, diglycerides, and triglycerides), but a stable profile at the lower mass-region [45]. ‘Storage’ experiments are helpful examples of whether the time of death could be determined, and even after storage for up to 10 weeks post mortem at temperatures from −20 °C to room temperature, it was still possible to discriminate between species [45]. However, some deterioration in the metabolite signal was observed with longer storage time [45], which suggests that through monitoring the rate of degradation of lipids or other compounds, it would be possible to build a model that allows accurate estimation of the time of death. Metabolomics has been successfully applied to determining the time of death in human forensics [50], and recent studies show a steady increase in the concentrations of several specific metabolites post mortem [51], supporting the findings in insects. Given the upregulation of genes and biochemical pathways involved in the degradation of tissues in any organism during the dynamic process of ‘death’ [52], more in-depth mass spectral fingerprinting studies with insects could be informative. This could include the method of death (e.g., caught in a trap, found dead on a chemically treated commodity), which would further enhance the depth of information that could be gained through entometabolomics for biosecurity diagnostics.
3.1.6. Geographical Origin
Determining the geographical origin of an insect is also valuable for biosecurity to help identify risk pathways. Molecular tools can be used in cases where the pest is genetically well-defined so that different populations can be linked through genetic loci [53]. The immediate within-generation origin of a detected high-risk exotic pest can inform if it is a new arrival from offshore or if it belongs to an already-established, yet undetected, locally breeding population. These are two scenarios that represent quite different biosecurity risks. In biosecurity and other forensic applications, non-metabolomics mass spectrometry tools have been used for this purpose, measuring light and heavy element bioavailable isotope ratios as environmental markers taken up into the tissues via the diet [7,54].
Owing to the efficacy and broad applicability of stable isotope measurements for determining the provenance of a sample, there has been little focus on other methods, such as metabolomics, for the same purpose. However, there is potential for mass spectral fingerprinting to be used to determine insect provenance. For instance, recently, REIMS could differentiate between the country of origin in lamb meat samples [23]. Biologically, it would be expected that the combined impact of differences in population genetics, host/feed, and environment would lead to measurable differences in the metabolite composition of insects of similar species but originating from different countries.
3.1.7. Plant Host
Metabolomic differences between insects fed on different plant diets have been demonstrated. For example, LC-MS metabolomics on frass could distinguish between Pieris rapae (Lepidoptera: Pieridae) larvae feeding on Brassicaceae or Cleomaceae [45]. There were different degrees of the breakdown of plant metabolites: metabolites from Brassicaceae were primarily broken down by larvae, while many metabolites from Cleome spinosa were detected directly in larvae feeding on the plant [55]. In another example, REIMS fingerprinting of black cricket (Gryllus bimaculatus) (Orthoptera: Gryllidae) frass fed on three different diets (leafy greens, oats, and potatoes) found that insect diet could be determined from the mass spectral fingerprint of frass [45]. Using LC-MS metabolomics, changes to the root microbiome of tomato plants were found to change the metabolome of Manduca sexta (Lepidoptera: Sphingidae) larvae feeding on the leaves of tomato plants [56]. These examples, while not useful for biosecurity as the larvae are always physically associated with their host plant, illustrate that diet could be detected in the metabolome. This could present complexity in interpreting mass spectral fingerprints for polyphagous species. However, with reference data for likely hosts, the dietary history of post-border detections of adult insects could be helpful. For example, for hemimetabolous (i.e., developing through incomplete metamorphosis) pests such as brown marmorated stink bug (Halyomorpha halys) (Hemiptera: Pentatomidae), where the diet is maintained through nymphs to adults, location of the population from a dispersing adult might be possible based on the plant host used during its nymphal stage. For holometabolous (i.e., developing through complete metamorphosis) insects, such as fruit flies, with distinct and discontinuous life stages where the trapped adults no longer feed, dietary history information may not be possible to determine, but is worthy of further research.
3.1.8. Insecticide Resistance
Biosecurity issues may also arise from introducing exotic phenotypes such as insecticide-resistant populations in many sectors [57,58], including horticulture [59,60]. Typically, detection is achieved with specific molecular tests, requiring knowledge of the gene target and considerable research and development [61]. Still, marker discovery might be more easily accomplished with metabolomics analysis. For instance, in mosquitoes resistant to deltamethrin, over 100 metabolites were detected using LC-MS metabolomics that differed between resistant and susceptible mosquitoes, with the biological finding that glycerophospholipid metabolism was upregulated in both larval and adult deltamethrin-resistant mosquitos [62]. As physiological responses such as chemical resistance continue to evolve, with no outward indication of the mechanism, this top-down, hypothesis-free diagnostic approach has the potential to be much more efficient than the equivalent molecular quantitative trait loci mapping or state-of-the-art genome sequencing.
3.1.9. Insects as Vectors of Plant Pathogens
Establishing whether an intercepted insect of a species that is known to vector high-risk plant pathogens has acquired a pathogen is another biosecurity concern. Diagnosis can be achieved using molecular laboratory or in-field tests [63] but requires knowledge of pathogen-specific gene targets and pre-analysis DNA extraction. While several metabolomic studies have investigated the impacts of pathogen infection on plants, few have considered the insects carrying the pathogen. Using GC-MS, substantial changes to energy-metabolism-related metabolites were observed in Asian citrus psyllids, Diaphorina citri (Hemiptera: Liviidae), reared on ‘Candidatus Liberibacter asiaticus’ (CLa) (Hyphomicrobiales: Rhizobiaceae)-infected oranges compared with those on healthy oranges [64]. An independent study using LC-MS in the same model also found a substantial alteration in amino acid and purine metabolism, along with changes to concentrations of several lipids in the psyllids [65]. There is insufficient evidence to infer whether these changes are pathogen-specific or pathogen-induced energetic stress responses [66]. These studies suggest detecting an altered metabolome could be a useful screening method to differentiate pathogen-carrying insects. However, further research is needed to determine if these responses are pathogen-specific, related to the overall vector stress response from carrying the pathogen, or due to an interaction between the infected plant host for the vector organism, the vector, and the pathogen.
3.2. Metabolomics Applied to the Biosecurity Detection of Plant Pathogens
The detection of plant pathogens in planta is achievable with many molecular genetic methods but remains highly challenging because of the cryptic nature of asymptomatic and latent infections. Tools that can quickly detect early-stage, asymptomatic, or latent infections in plants are desirable both at the border and in the event of an incursion to reduce costly time in quarantine and disease management, respectively [67]. Metabolomics-based methods have not been used in this context but have been useful for the scientific understanding of plant–pathogen interactions, which may have the potential for diagnostics. For example, plant responses to attacks by pathogens include cell wall strengthening, production of secondary metabolites (often phenolic compounds), and generation of reactive oxygen species [11]. Several metabolites found in plant–pathogen interactions have been ascribed ‘attack’ or ‘defence’ roles [11,68,69,70], suggesting that the metabolic profile of a plant, as a snapshot of its metabolic state, could be used to determine if a plant is under current pathogen attack and possibly from what pathogen. However, this can be complicated by a pathogen successfully switching off plant defences, as has been demonstrated with Pseudomonas syringae (Pseudomonadales: Pseudomonadaceae) preventing syringic, jasmonic, and abscisic acid signalling in the model brassica Arabidopsis (Brassicaceae) [71].
Specific metabolic profiles or fingerprints of plant–pathogen interactions face several confounding factors that may complicate the interpretation of the results. These include determining the origin of metabolites, as they may stem from the host plant, symbiotic microorganisms (e.g., endophytes, rhizobia), and/or pathogenic microorganisms [68,70]. Plant stress may also lead to the production of metabolites that could confound clear identification of the presence of a pathogen. Present studies suggest that heat and drought stress may also stimulate secondary metabolic pathways (e.g., the shikimate pathway), as well as primary pathways (e.g., energy, lipid, and amino acid metabolism) [72]. These influences on the plant metabolome also need to be assessed against natural plant variation, differences due to season and soil, and inter-plant interactions (allelopathy), among others [73].
Nevertheless, as symbiotic microorganisms both provide additional metabolites to a host plant and upregulate the production of specific metabolites by the host plant [68], the broad view of metabolism provided by metabolomics should make it possible to differentiate the relative impact of stressors, symbiotic, and pathogenic microorganisms. Supporting the concept of broad-based methods for detecting and characterising host–pathogen interactions, lipid-signalling molecules, including prostaglandins, were among the metabolites upregulated in winter wheat grains infected with the smut fungus Tilletia controvera Kühn (Tilletiales: Tilletiaceae) [74] suggesting that while most studies have focused on primary and secondary metabolites, lipids may also be helpful diagnostic molecules. As new model systems to study host–pathogen interactions are designed, these diverse interactions must be factored in if scientific results are also to be translated into practical diagnostic tools for biosecurity.
The detection of early-stage or latent infections in plants is challenging for metabolomics methods, as any metabolic response of a plant may take time to be initiated or may not be sufficient to detect above the baseline variation in metabolism (e.g., because of diurnal patterns, maturation). Alternatively, it may be possible to detect signals from the pathogen itself. Still, it may be complicated with early-stage infection, and it is unlikely if the pathogen load is low. Potentially, if the plant produces metabolite-based signals that keep the pathogen in a latent state, this may be detectable. Infection of strawberries by the fungus Colletotrichum theobromicola (Glomerellales: Glomerellaceae) was diagnosed using GC-MS metabolomics four days before symptoms were visible on leaves [75], although notably, the profile of predictive metabolites changed over time, reflecting the dynamic nature of plant infection by the pathogen and underpinning the importance of including ‘time since infection’ as a variable when designing metabolomics studies on plant–pathogen interactions. Another study on the grey mould Botrytis cinerea (Helotiales: Sclerotiniaceae) infection of strawberries found that metabolite differences in leaves could be detected as early as two days post-inoculation, with differences in fatty acids (hexadecenoic and octadecanoic acids) and carbohydrates (sucrose, β-lyxopyranose, and melibiose) before visible symptoms of infection [76]. As an example of temporal influences on the plant metabolome, rice plants infested with Cnaphalocrocis medinalis (Lepidoptera: Crambidae) caterpillars showed a differential metabolic response 1–12 h post-infestation, but a less marked difference at 24 h [77], and we speculate that similar temporal changes may be detectable with pathogen attacks on plants.
One of the more open questions for developing diagnostics for determining if plants are infected by pathogens or not is pathogen identification—especially if it is an exotic compared with an endemic pathogen. Intriguingly, using LC-MS metabolomics it was established that different plant species produced different metabolomic responses to infestation by either native or non-native pea aphids (Acyrthosiphon pisum (Hemiptera: Aphididae) [78]. Earlier work had established that insect–plant interactions are unique for different insects on the same plant [79]. A wide range of plant defence compounds (e.g., flavonoids, saponins, peptides) was produced in response to A. pisum infestation, and the type of response depended on whether the aphid usually fed on that species of plant or not [78,79]. Whether similar distinctions can be made for bacterial or fungal pathogens remains to be determined.
GC-MS metabolomics analysis of phloem sap and leaves after infection of mulberry plants (Morus multicaulis) (Rosales: Moraceae) by mulberry yellow dwarf disease found different metabolic responses to the disease in these two sampling areas [80], while ‘Candidatus Liberibacter’ sp. infection of citrus is noted as specifically located in the phloem rather than in other tissues, leading to corresponding differences in metabolomic profiles depending on the sampling site [11]. These findings are not surprising given that these two pathogens are phloem-limited and require an insect vector for transmission into the plant’s vascular system. Knowing that detection of and early response to a particular pathogen may depend on sampling from the correct site on a plant can help to streamline sampling strategies. This underlines the importance of understanding the interactions and the mechanistic basis of infection when developing metabolomics-based markers of plant infection.
Although based on isolated viruses, MS has been successfully used to identify plant viruses based on peptide fragments of viral coat proteins [81], supporting the ability of mass spectral methods to determine virus species based on non-genetic components. Similar work has been conducted on fungal plant pathogens, including Fusarium culmorum (Hypocreales: Nectriaceae), using LC-MS metabolomics to understand the potential mechanisms of host plant–pathogen interactions [82]. This approach could be compelling when combined with genomic and proteomic studies, which can highlight specific pathways and molecules that may be unique for a particular plant–pathogen system.
The use of metabolomics for high-throughput plant phenotyping has recently been advocated to understand the quality, variation, and added-value opportunities stemming from plant growth variation within greenhouses, fields, and orchards [83]. Automated systems, including sensors, imaging, and spectroscopic and mass spectrometric methods, are proposed to generate phenotypic data on individual plants quickly. Similar approaches could be envisaged for screening changes related to, for example, pathogen presence in quarantined plants. Using multiple methods beyond just metabolomics will enhance the sensitivity and specificity of such detection systems. There is a need for the greater use of ambient ionisation metabolomics to make this approach feasible, based on AI-MS instruments such as DART-MS and REIMS, which do not require extensive sample preparation before measurement.
3.3. Other Potential Biosecurity Applications
There are many other potential biosecurity applications where metabolomics could be helpful. Several countries restrict or ban the import of genetically modified organisms, and the area of targeted manipulation of agricultural genetic material has received an additional spotlight with the advent of CRISPR/Cas-based genetic modification. Several studies have found that untargeted metabolomics can detect differences between genetically modified crops and closely related breeds. However, the detected changes do not always match what would be expected from the genetic modification [84]. Most research literature has focused on metabolomics to discover unexpected effects of genetic modification on the expressed metabolome [85,86], rather than purely as a detection tool. As for identifying the traits in insects considered above, determining the genetic modification status of important plants or seeds would require a large reference population-based sample that captures natural variation or existing knowledge of possible genetic modifications and the metabolites that are likely to be affected.
Another related use of metabolomics for biosecurity is to detect possible herbicide resistance in new seeds or germplasm. Herbicide resistance has important implications for crop management. The spread of herbicide-resistance genes across related species will make weed management difficult [87], especially as farmers now have a greater impetus to reduce herbicide use. However, with many genetic mechanisms behind herbicide resistance, even for the same herbicide class, there is unlikely to be any single metabolite change or pathway that can be easily detected using metabolomics. As for other phenotypes of interest in insects and plants, linking differences in the measured metabolome to a particular phenotype of interest will require a combination of a relatively large number of samples to characterise a population, along with biological knowledge on how the phenotype might influence metabolism. New developments in chemometrics may help to discover or predict metabolome changes, and research in this area is necessary to obtain information beyond high-level sample categorisation, such as species and sex, and to delve into deeper information from metabolomics measurements [88] such as geographic origin and vector pathogen status.
Further applications could also include confirmation of quarantine treatment measures on plants, including devitalisation and fumigation or other chemical treatments to ensure that imported plants do not contain unwanted pests, as part of phytosanitation certification. While our literature search did not find any application of metabolomics for phytosanitation assurance, metabolomics methods could be adapted to include the targeted measurement of chemicals used in devitalisation and fumigation. A single way to detect the pathogen status of imported plants, if steps have been taken to ensure phytosanitation measures have been employed, is an interesting area of potential application for biosecurity metabolomics.
3.4. Translational Metabolomics for Biosecurity: Direct Analysis Mass Spectrometry
Translating the outcomes of metabolomics research into a practical application can present many hurdles [89]. A major limitation that might arise for biosecurity is the potential requirement for extensive sample preparation prior to some forms of analysis (e.g., chromatography-MS or NMR), which prevents metabolomics from being a truly high-throughput approach to sample analysis. However, in several examples presented above for insects, AI-MS, which does not need pre-analysis sample treatment, has been successfully used. This new, relatively inexpensive, and flexible approach can be of practical use in smaller ‘response’ laboratories and in the field (Figure 3), overcoming the hurdles associated with chromatography-mass spectrometry and NMR-based metabolomics, which are limited to use in well-equipped scientific laboratories.
AI-MS has several advantages over ‘traditional’ metabolomics, mainly in the speed of the analytical workflow—with little or no sample preparation—or the time required for chromatography. Although promising, the current applications of AI-MS in areas of biosecurity concern are limited. For example, species identification based on REIMS fingerprinting of faecal samples could differentiate species of rodent; strains of mice fed the same diet; and sex and age in mice [90]. Another type of AI-MS instrument, DART-MS, has been used to identify different species from morphologically similar seeds [91] and to detect the presence of herbicide residues in pasture seeds [92], along with detecting species of endangered trees from wood samples and insect species from pupal casings [22,93]. DART-MS has also been used to determine species of sarcophagous dipterans (i.e., maggots) and the time since infestation in tissues in decomposing meat [94]. These examples show that AI-MS has clear forensic applications and illustrate the depth of information that can be obtained from a mass spectral fingerprint. Notably, many studies reported here have used machine-learning algorithms to determine diagnostic fingerprints, underlining the potential to develop new data analysis tools to obtain diagnostic values from mass spectral fingerprints. In these studies, large sample sizes are generally required to adequately characterise a population, on the order of 50–400 per species, although with the advantage that a single MS fingerprint can provide biological information that is not only restricted to an individual’s species or sex.
Another challenge, shared across all analytical methods, is the small sample ordinarily available with a suspected biosecurity sample [24,49]. In many cases, this can be a single insect, which should be characterised using several methods, meaning there may only be a few mg of material available for analysis. To test this limitation, we have successfully used REIMS to analyse insect legs using a modified REIMS source [46], with a throughput of >400 legs/day (unpublished observations by the authors).
Other types of fingerprinting methods can also be applied, including matrix-assisted laser desorption ionisation-time of flight mass spectrometry (MALDI-TOF MS), which is widely used for determining microbial species [95]. MALDI-TOF MS requires a chemical matrix to be applied to a sample before analysis, which means it is not a true AI-MS instrument, although the result is a mass spectral fingerprint. MALDI-TOF MS has been successfully used to detect CLa in navel oranges [96], supporting that mass spectral fingerprints could be used for biosecurity diagnostics applications. Other methods that allow the direct measurement of metabolites or metabolite profiles in solid samples, such as solid-state NMR [97], provide additional possibilities for biosecurity diagnostics, but fall outside the scope of the present review.
3.5. The Interface between Diagnostics and Underpinning Metabolomics Science
Applied biosecurity diagnostics is focused on facilitating rapid decision-making on potential threats. The data acquired can also be of scientific importance to understand threat organisms and how they interact with hosts, biosecurity lures, and traps or behave in a new climate. Using broad-based methods such as metabolomics, where an overall ‘picture’ of a biosecurity threat organism is acquired, could allow the determination of traits relevant for biosecurity response decision-making (e.g., in the case of insects: species, strain, sexual maturity, time since death) as well as information that is of more research interest (e.g., metabolic pathway regulation, specific compounds that make up structural components, potential microbial metabolites related to host(s)). While it is more time-consuming to extract the scientific information from the metabolic profile, a broader understanding of the biosecurity threat from metabolic profiles can potentially lead to the development of better lures for traps, cheaper screening tools, and more effective mitigation measures. There are further benefits through using the same data to understand the mechanisms of how exotic and indigenous organisms differ, giving metabolomics utility beyond purely diagnostic applications.
For metabolomics methods to be effective for biosecurity diagnosis, practical aspects must be considered. Primary among these is the cost–benefit of what metabolomics measurements add to existing taxonomic and molecular tools. This includes the cost of the instrumentation and bringing in or training staff to operate and maintain instrumentation that would be unfamiliar in most current biosecurity diagnostics laboratories. After investment in instrumentation, service contracts and other maintenance costs, and staff, per-sample costs are low, covering solvents, standards, and, if needed, chromatography columns. ‘Traditional’ metabolomics instrumentation such as NMR, GC-MS, and LC-MS bring high precision (and sensitivity in the case of MS) but are lengthy in sample preparation and analysis time.
Conversely, although there is scant literature for biosecurity applications, AI-MS methods are an up-and-coming alternative for routine diagnostics owing to the higher throughput of up to several hundred samples per day and the lack of need to maintain a chromatography system in addition to the mass spectrometer. In our experience, a person with limited practical scientific experience, such as an undergraduate student, can be trained to run samples independently on an AI-MS instrument within a few days. Bioinformatics and data storage tools need to develop to enable real-time feedback on metabolomics measurements regarding past or open-source data. While this is a significant undertaking, such tools are already in place for genetics and genomics analyses for biosecurity. The uptake of metabolomics technologies across biosecurity laboratories worldwide will be a major advantage, as it will allow the sharing of experiences and the setting of common standards for biosecurity metabolomics.
4. Conclusions
With globalised tourism and trade networks and a changing climate, new plant biosecurity threats are emerging regularly, which require fast, accurate, and reliable diagnostic tools. Metabolomics is a rapidly maturing analytical science [89], including the advancement of ambient ionisation techniques, which now presents the opportunity to complement the taxonomic and molecular methods currently used to determine information about biosecurity incursions. The recent literature suggests that the depth of information obtained from a single metabolomics analysis includes an insect’s species, sex, time since death, host plant, parasitism, and origin. For plant pathogens, examples that provide specific evidence of metabolomics for biosecurity applications are lacking. However, based on what has been learnt from the range of published research on plant–pathogen interactions, extension to biosecurity applications with well-defined goals appears to be a feasible direction for new research. Although not addressed in this review, there is a similar potential for applying metabolomics to animal biosecurity. Metabolomics is a new tool for biosecurity. While its application has substantial potential, implementation will require significant research efforts to make it a viable tool for biosecurity diagnostics. Through such efforts, valuable tools for rapid and detailed diagnosis of biosecurity threats that complement existing tools could be developed, which will have the added advantage of furthering the scientific understanding of biosecurity threat species and their interactions with at-risk plants.
Author Contributions
Conceptualisation, A.B.R., K.F.A.; writing-original draft preparation, A.B.R., H.M., A.N.-R., K.F.A.; writing-review and editing, A.B.R., H.M., A.S., I.H., F.M., S.H., L.S., J.V., S.V., A.N.-R., K.F.A.; funding acquisition, A.B.R., K.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This review was supported by a grant from the Better Border Biosecurity (B3, b3nz.org.nz) research collaboration, project #D21.14, and through the New Zealand Strategic Science Investment Fund.
Data Availability Statement
No new data were created or analysed in this study. Data sharing is not applicable to this article.
Acknowledgments
We thank Alice Baillie (AgResearch) for her work on the figures used in this review; Gabby Drayton (Plant & Food Research) for technical support; and Robert Taylor, Lia Liefting, and Diane Anderson (Ministry for Primary Industries, New Zealand) for their comments.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations and Glossary
CHC: Cuticular hydrocarbon(s).
Chemometrics:
The application of mathematical and statistical tools to extract chemical information from a measurement. A fundamental process in metabolomics to turn acquired data into usable information.
AI-MS: Ambient ionisation-mass spectrometry:
A subgroup of mass spectrometers where the ionisation process needed to detect molecules occurs outside of the vacuum environment of the mass spectrometer. This enables analysis of samples with no or minimal sample processing. Includes DART-MS and REIMS.
DART-MS: Direct analysis in real time-mass spectrometry:
A type of mass spectrometry interface that uses heated helium to generate ions for rapid detection by a mass spectrometer.
Features:
In metabolomics, used to describe what is detected by a mass spectrometer prior to identification. A metabolomics analysis may detect hundreds to thousands of features, but these do not necessarily translate to the same number of identifiable metabolites. One metabolite may lead to several detected features, or a feature may be a metabolite that cannot be identified.
GC: Gas chromatography:
A chromatography method that can interface with mass spectrometry. Often used for measuring volatile metabolites, energy metabolism pathway metabolites, and free fatty acids.
LC: Liquid chromatography:
A chromatography method that can interface with mass spectrometry. Highly flexible owing to the range of solvents and chromatography columns. LC coupled to MS is the most common instrumentation used for metabolomics.
MS: Mass spectrometry.
NMR: Nuclear magnetic resonance spectroscopy.
PCA: Principal components analysis:
A type of unsupervised multivariate statistical analysis that models the variation between different samples based on measures associated with those samples. Commonly used as a basic overview method in metabolomics, and can be complemented with supervised multivariate statistical models, machine learning, and ‘traditional’ univariate statistical analyses.
REIMS: Rapid evaporative ionisation mass spectrometer:
A type of AI-MS interface that ionises a sample vapour or smoke for rapid detection with a mass spectrometer. It can be used in conjunction with a surgical knife or laser.
Targeted and untargeted metabolomics:
Metabolomics methods are often referred to as ‘targeted’ or ‘untargeted’. A targeted metabolomics method is set up to measure pre-defined metabolites in a sample. For example, a mass-spectrometer-based method may be set up to measure specific amino acids, organic acids, or lipids. Untargeted metabolomics methods are set up to detect everything in the sample that can be detected and require an identification step, usually after statistical analysis.
Generally, targeted methods have the advantage of greater confidence in identification, and some can be semi-quantitative, but are limited to measuring those metabolites that have been pre-defined. Quantitative analytical methods such as fatty acid or amino acid analysis are not true ‘targeted metabolomics methods’ as they will be applied to samples with a hypothesis that these metabolites will differ. Untargeted metabolomics detects a broader range of metabolites, including unknown ones, and compound identification can be a time-consuming step but is useful for discovery-type research questions.
References
- Waage, J.K.; Mumford, J.D. Agricultural biosecurity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, K.; Ball, S. DNA barcodes for biosecurity: Invasive species identification. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1813–1823. [Google Scholar] [CrossRef] [PubMed]
- Olmos, A.; Boonham, N.; Candresse, T.; Gentit, P.; Giovani, B.; Kutnjak, D.; Liefting, L.; Maree, H.J.; Minafra, A.; Moreira, A.; et al. High-throughput sequencing technologies for plant pest diagnosis: Challenges and opportunities. EPPO Bull. 2018, 48, 219–224. [Google Scholar] [CrossRef]
- Boykin, L.M.; Sseruwagi, P.; Alicai, T.; Ateka, E.; Mohammed, I.U.; Stanton, J.-A.L.; Kayuki, C.; Mark, D.; Fute, T.; Erasto, J.; et al. Tree Lab: Portable Genomics for Early Detection of Plant Viruses and Pests in Sub-Saharan Africa. Genes 2019, 10, 632. [Google Scholar] [CrossRef]
- Cleary, M.; Oskay, F.; Doğmuş, H.; Lehtijärvi, A.; Woodward, S.; Vettraino, A.M. Cryptic Risks to Forest Biosecurity Associated with the Global Movement of Commercial Seed. Forests 2019, 10, 459. [Google Scholar] [CrossRef]
- Congrains, C.; Zucchi, R.A.; de Brito, R.A. Phylogenomic approach reveals strong signatures of introgression in the rapid diversification of neotropical true fruit flies (Anastrepha: Tephritidae). Mol. Phylogenet. Evol. 2021, 162, 107200. [Google Scholar] [CrossRef]
- Holder, P.W.; Van Hale, R.; Frew, R.; George, S.; Armstrong, K.F. Natal origin of the invasive biosecurity pest, brown marmorated stink bug (Halyomorpha halys: Penatomidae), determined by dual-element stable isotope-ratio mass spectrometry. Pest Manag. Sci. 2020, 76, 1456–1463. [Google Scholar] [CrossRef]
- Reese, K.L.; Rasley, A.; Avila, J.R.; Jones, A.D.; Frank, M. Metabolic Profiling of Volatile Organic Compounds (VOCs) Emitted by the Pathogens Francisella tularensis and Bacillus anthracis in Liquid Culture. Sci. Rep. 2020, 10, 9333. [Google Scholar] [CrossRef]
- Trifonova, O.P.; Maslov, D.; Balashova, E.; Lokhov, P.G. Mass spectrometry-based metabolomics diagnostics-myth or reality? Expert Rev. Proteom. 2021, 18, 7–12. [Google Scholar] [CrossRef]
- Blakebrough-Hall, C.; Dona, A.; D’Occhio, M.J.; McMeniman, J.; A González, L. Diagnosis of Bovine Respiratory Disease in feedlot cattle using blood 1H NMR metabolomics. Sci. Rep. 2020, 10, 115. [Google Scholar] [CrossRef]
- Castro-Moretti, F.R.; Gentzel, I.N.; Mackey, D.; Alonso, A.P. Metabolomics as an Emerging Tool for the Study of Plant–Pathogen Interactions. Metabolites 2020, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Rasmussen, S.; Lane, G.A. Metabolomics or metabolite profiles? Trends Biotechnol. 2005, 23, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2021, 50, D622–D631. [Google Scholar] [CrossRef]
- Orešič, M.; McGlinchey, A.; Wheelock, C.E.; Hyötyläinen, T. Metabolic Signatures of the Exposome—Quantifying the Impact of Exposure to Environmental Chemicals on Human Health. Metabolites 2020, 10, 454. [Google Scholar] [CrossRef]
- Soltis, N.E.; Kliebenstein, D.J. Natural Variation of Plant Metabolism: Genetic Mechanisms, Interpretive Caveats, and Evolutionary and Mechanistic Insights. Plant Physiol. 2015, 169, 1456–1468. [Google Scholar] [CrossRef]
- Chong, J.; Yamamoto, M.; Xia, J. MetaboAnalystR 2.0: From raw spectra to biological insights. Metabolites 2019, 9, 57. [Google Scholar] [CrossRef]
- Draper, S.L.; McCarney, E.R. Benchtop nuclear magnetic resonance spectroscopy in forensic chemistry. Org. Magn. Reson. 2021, 61, 106–129. [Google Scholar] [CrossRef] [PubMed]
- Barding, G.A., Jr.; Béni, S.; Fukao, T.; Bailey-Serres, J.; Larive, C.K. Comparison of GC-MS and NMR for Metabolite Profiling of Rice Subjected to Submergence Stress. J. Proteome Res. 2013, 12, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Balog, J.; Szaniszlo, T.; Schaefer, K.-C.; Denes, J.; Lopata, A.; Godorhazy, L.; Szalay, D.; Balogh, L.; Sasi-Szabo, L.; Toth, M.; et al. Identification of Biological Tissues by Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2010, 82, 7343–7350. [Google Scholar] [CrossRef]
- Balog, J.; Sasi-Szabó, L.; Kinross, J.; Lewis, M.; Muirhead, L.; Veselkov, K.; Mirnezami, R.; Dezső, B.; Damjanovich, L.; Darzi, A.; et al. Intraoperative tissue identification using rapid evaporative ionization mass spectrometry. Sci. Transl. Med. 2013, 5, 194ra193. [Google Scholar] [CrossRef] [PubMed]
- Beyramysoltan, S.; Giffen, J.; Rosati, J.; Musah, R.A. Direct Analysis in Real Time-Mass Spectrometry and Ko-honen Artificial Neural Networks for Species Identification of Larva, Pupa and Adult Life Stages of Carrion Insects. Anal. Chem. 2018, 90, 9206–9217. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.; Brunius, C.; Chevallier, O.; Dervilly, G.; Elliott, C.; Guitton, Y.; Prenni, J.E.; Savolainen, O.; Hemeryck, L.; Vidkjær, N.H.; et al. Making complex measurements of meat composition fast: Application of rapid evaporative ionisation mass spectrometry to measuring meat quality and fraud. Meat Sci. 2020, 181, 108333. [Google Scholar] [CrossRef]
- Snart, C.J.; Hardy, I.C.; Barrett, D.A. Entometabolomics: Applications of modern analytical techniques to insect studies. Èntomol. Exp. Et Appl. 2015, 155, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.B.; Iline, I.I.; Richards, N.K.; Novoselov, M.; McNeill, M. Development and validation of a quick easily used biochemical assay for evaluating the viability of small immobile arthropods. J. Econ. Èntomol. 2013, 106, 2006–2019. [Google Scholar] [CrossRef]
- Holze, H.; Schrader, L.; Buellesbach, J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity 2020, 126, 219–234. [Google Scholar] [CrossRef]
- Kather, R.; Martin, S.J. Cuticular hydrocarbon profiles as a taxonomic tool: Advantages, limitations and technical aspects. Physiol. Èntomol. 2012, 37, 25–32. [Google Scholar] [CrossRef]
- Wang, Q.; Goodger, J.; Woodrow, I.; Elgar, M.A. Location-specific cuticular hydrocarbon signals in a social insect. Proc. Biol. Sci. 2016, 283, 20160310. [Google Scholar] [CrossRef]
- Würf, J.; Pokorny, T.; Wittbrodt, J.; Millar, J.G.; Ruther, J. Cuticular Hydrocarbons as Contact Sex Pheromone in the Parasitoid Wasp Urolepis rufipes. Front. Ecol. Evol. 2020, 8, 180. [Google Scholar] [CrossRef]
- Bien, T.; Gadau, J.; Schnapp, A.; Yew, J.Y.; Sievert, C.; Dreisewerd, K. Detection of very long-chain hydrocarbons by laser mass spectrometry reveals novel species-, sex-, and age-dependent differences in the cuticular profiles of three Nasonia species. Anal. Bioanal. Chem. 2019, 411, 2981–2993. [Google Scholar] [CrossRef]
- Souza, N.M.; Schröder, M.; Hayes, R.; Bello, J.; Nahrung, H.F. Cuticular hydrocarbons of Gonipterus weevils: Are there species differences? Chemoecology 2021, 31, 159–167. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Copley, T.; Jabaji, S. Gas chromatography–mass spectrometry metabolite profiling of worker honey bee (Apis mellifera L.) hemolymph for the study of Nosema ceranae infection. J. Insect Physiol. 2012, 58, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, H.; Hao, F.; Li, N.; Liu, X.; Wang, G.; Wang, Y.; Tang, H. Developmental Changes for the Hemolymph Metabolome of Silkworm (Bombyx mori L.). J. Proteome Res. 2015, 14, 2331–2347. [Google Scholar] [CrossRef] [PubMed]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Domaciuk, M.; Krutmuang, P.; Paleolog, J. A New Method for Quick and Easy Hemolymph Collection from Apidae Adults. PLoS ONE 2017, 12, e0170487. [Google Scholar] [CrossRef] [PubMed]
- Bozic, J.; Di Cesare, J.; Wells, H.; Abramson, C.I. Ethanol levels in honeybee hemolymph resulting from alcohol in-gestion. Alcohol 2007, 41, 281–284. [Google Scholar] [CrossRef]
- Stevens, V.L.; Hoover, E.; Wang, Y.; Zanetti, K.A. Pre-Analytical Factors that Affect Metabolite Stability in Human Urine, Plasma, and Serum: A Review. Metabolites 2019, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, R.; González-Domínguez, Á.; Sayago, A.; Fernández-Recamales, Á. Recommendations and Best Practices for Standardizing the Pre-Analytical Processing of Blood and Urine Samples in Metabolomics. Metabolites 2020, 10, 229. [Google Scholar] [CrossRef]
- Hulme, P.E. One Biosecurity: A unified concept to integrate human, animal, plant, and environmental health. Emerg. Top. Life Sci. 2020, 4, 539–549. [Google Scholar] [CrossRef]
- Vaníčková, L.; Virgilio, M.; Tomčala, A.; Břízová, R.; Ekesi, S.; Hoskovec, M.; Kalinová, B.; Nascimento, R.D.; De Meyer, M. Resolution of three cryptic agricultural pests (Ceratitis fasciventris, C. anonae, C. rosa, Diptera: Tephritidae) using cuticular hydrocarbon profiling. Bull. Entomol. Res. 2014, 104, 631–638. [Google Scholar] [CrossRef]
- De Meyer, M.; Delatte, H.; Ekesi, S.; Jordaens, K.; Kalinova, B.; Manrakhan, A.; Mwatawala, M.; Steck, G.; Van Cann, J.; Vancikova, L.; et al. An integrative approach to unravel the Ceratitis FAR (Diptera, Tephritidae) cryptic species complex: A review. Zookeys 2015, 540, 405–427. [Google Scholar] [CrossRef]
- Copren, K.A.; Nelson, L.J.; Vargo, E.L.; Haverty, M.I. Phylogenetic analyses of mtDNA sequences corroborate taxonomic designations based on cuticular hydrocarbons in subterranean termites. Mol. Phylogenet. Evol. 2005, 35, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Moore, H.E.; Hall, M.; Drijfhout, F.; Cody, R.; Whitmore, D. Cuticular hydrocarbons for identifying Sar-cophagidae (Diptera). Sci. Rep. 2021, 11, 7732. [Google Scholar] [CrossRef]
- Wagner, I.; Koch, N.; Sarsby, J.; White, N.; Price, T.; Jones, S.; Hurst, J.; Beynon, R.J. The application of rapid evaporative ionization mass spectrometry in the analysis of Drosophila species—A potential new tool in ento-mology. Open Biol. 2020, 10, 200196. [Google Scholar] [CrossRef]
- Chabi, J.; Van’t Hof, A.; N’Dri, L.K.; Datsomor, A.; Okyere, D.; Njoroge, H.; Pipini, D.; Hadi, M.P.; de Souza, D.K.; Suzuki, T.; et al. Rapid high throughput SYBR green assay for identifying the malaria vectors Anopheles ara-biensis, Anopheles coluzzii and Anopheles gambiae s.s. Giles. PLoS ONE 2019, 14, e0215669. [Google Scholar] [CrossRef] [PubMed]
- Wagner, I. Exploration of Rapid Evaporative Ionisation Mass Spectrometry as a Novel Tool for Insect Identification and Characterisation. Ph.D. Thesis, University of Liverpool, Liverpool, UK, 2021. [Google Scholar]
- Cameron, S.J.S.; Alexander, J.L.; Bolt, F.; Burke, A.; Ashrafian, H.; Teare, J.P.; Marchesi, J.R.; Kinross, J.M.; Li, J.V.; Takáts, Z. Evaluation of Direct from Sample Metabolomics of Human Feces Using Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2019, 91, 13448–13457. [Google Scholar] [CrossRef]
- Otte, T.; Hilker, M.; Geiselhardt, S. Phenotypic Plasticity of Cuticular Hydrocarbon Profiles in Insects. J. Chem. Ecol. 2018, 44, 235–247. [Google Scholar] [CrossRef]
- Kuo, T.-H.; Yew, J.; Fedina, T.; Dreisewerd, K.; Dierick, H.; Pletcher, S.D. Aging modulates cuticular hydro-carbons and sexual attractiveness in Drosophila melanogaster. J. Exp. Biol. 2012, 215 Pt 5, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Kapranas, A.; Snart, C.J.P.; Williams, H.; Hardy, I.C.W.; Barrett, D.A. Metabolomics of aging assessed in individual parasitoid wasps. Sci. Rep. 2016, 6, 34848. [Google Scholar] [CrossRef]
- Dawidowska, J.; Krzyżanowska, M.; Markuszewski, M.; Kaliszan, M. The Application of Metabolomics in Forensic Science with Focus on Forensic Toxicology and Time-of-Death Estimation. Metabolites 2021, 11, 801. [Google Scholar] [CrossRef]
- Pesko, B.K.; Weidt, S.; McLaughlin, M.; Wescott, D.J.; Torrance, H.; Burgess, K.; Burchmore, R. Postmortomics: The Potential of Untargeted Metabolomics to Highlight Markers for Time Since Death. OMICS A J. Integr. Biol. 2020, 24, 649–659. [Google Scholar] [CrossRef]
- Bonadio, R.S.; Nunes, L.B.; Moretti, P.N.S.; Mazzeu, J.F.; Cagnin, S.; Pic-Taylor, A.; de Oliveira, S.F. Insights into how environment shapes post-mortem RNA transcription in mouse brain. Sci. Rep. 2021, 11, 13008. [Google Scholar] [CrossRef]
- Barr, N.; Ruiz-Arce, R.; Armstrong, K. Using Molecules to Identify the Source of Fruit Fly Invasions. Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies: Lures, Area-Wide Programs, and Trade Implications; Shelly, T., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 321–378. [Google Scholar] [CrossRef]
- Holder, P.W.; Armstrong, K.; Van Hale, R.; Millet, M.-A.; Frew, R.; Clough, T.; Baker, J. Isotopes and Trace Elements as Natal Origin Markers of Helicoverpa armigera—An Experimental Model for Biosecurity Pests. PLoS ONE 2014, 9, e92384. [Google Scholar] [CrossRef]
- Riach, A.C.; Perera, M.V.L.; Florance, H.V.; Robinson, L.A.; Penfield, S.D.; Hill, J.K. Metabolic fingerprints reveal how an insect metabolome is affected by different larval host plant species. Arthropod-Plant Interact. 2019, 13, 571–579. [Google Scholar] [CrossRef]
- Papantoniou, D.; Vergara, F.; Weinhold, A.; Quijano, T.; Khakimov, B.; Pattison, D.; Bak, S.; van Dam, N.; Mar-tínez-Medina, A. Cascading Effects of Root Microbial Symbiosis on the Development and Metabolome of the Insect Her-bivore Manduca sexta L. Metabolites 2021, 11, 731. [Google Scholar] [CrossRef]
- Candas, M.; Loseva, O.; Oppert, B.; Kosaraju, P.; Bulla, L.A., Jr. Insect Resistance to Bacillus thuringiensis: Alterations in the Indianmeal Moth Larval Gut Proteome. Mol. Cell. Proteom. 2003, 2, 19–28. [Google Scholar] [CrossRef]
- Abougamos, H.R.; Sadler, R.; White, B. Managing evolving insecticide resistance in stored grain pests within Avon Region, Western Australia. J. Stored Prod. Postharvest Res. 2017, 8, 16–30. [Google Scholar]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Liu, S.; Liu, L.; Tay, W.T.; Walsh, T.K.; Yang, Y.; Wu, Y. CRISPR/Cas9 mediated genome editing of Helicoverpa armigera with mutations of an ABC transporter gene HaABCA2 confers resistance to Bacillus thuringiensis Cry2A toxins. Insect Biochem. Mol. Biol. 2017, 87, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Dermauw, W.; Mavridis, K.; Vontas, J. Significance and interpretation of molecular diagnostics for insecticide resistance management of agricultural pests. Curr. Opin. Insect Sci. 2020, 39, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Wang, G.; Li, J.; Zhang, M.; Wu, J.; Liang, C.; Zhou, H.; Tang, J.; Zhu, G. Differential metabolome responses to deltamethrin between resistant and susceptible Anopheles sinensis. Ecotoxicol. Environ. Saf. 2022, 237, 113553. [Google Scholar] [CrossRef]
- Pusz-Bochenska, K.; Perez-Lopez, E.; Dumonceaux, T.; Olivier, C.; Wist, T.J. A Rapid, Simple, Laboratory and Field-Adaptable DNA Extraction and Diagnostic Method Suitable for Insect-Transmitted Plant Pathogen and Insect Identifi-cation. Plant Health Prog. 2020, 21, 63–68. [Google Scholar] [CrossRef]
- Killiny, N.; Jones, S.E. Metabolic alterations in the nymphal instars of Diaphorina citri induced by Candidatus Liberibacter asiaticus, the putative pathogen of huanglongbing. PLoS ONE 2018, 13, e0191871. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; He, J.; Guan, Z.; Zhong, M.; Pang, R.; Han, Q. Transcriptomic and Metabolomic Analyses of Diaphorina citri Kuwayama Infected and Non-infected With Candidatus Liberibacter Asiaticus. Front. Physiol. 2021, 11, 630037. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Nehela, Y.; Hijaz, F.; Vincent, C.I. A plant pathogenic bacterium exploits the tricarboxylic acid cycle metabolic pathway of its insect vector. Virulence 2017, 9, 99–109. [Google Scholar] [CrossRef]
- Smith, G.R.; Fletcher, J.; Marroni, V.; Kean, J.; Stringer, L.; Vereijssen, J. Plant pathogen eradication: Determi-nants of successful programs. Australas. Plant Pathol. 2017, 46, 277–284. [Google Scholar] [CrossRef]
- Adeniji, A.A.; Babalola, O.O.; Loots, D.T. Metabolomic applications for understanding complex tripartite plant-microbes interactions: Strategies and perspectives. Biotechnol. Rep. 2020, 25, e00425. [Google Scholar] [CrossRef]
- Vo, K.T.X.; Rahman, M.; Rahman, M.; Trinh, K.T.T.; Kim, S.T.; Jeon, J.-S. Proteomics and Metabolomics Studies on the Biotic Stress Responses of Rice: An Update. Rice 2021, 14, 30. [Google Scholar] [CrossRef]
- Gupta, S.; Schillaci, M.; Roessner, U. Metabolomics as an emerging tool to study plant–microbe interactions. Emerg. Top. Life Sci. 2022, 6, 175–183. [Google Scholar] [CrossRef]
- De Torres-Zabala, M.; Truman, W.; Bennett, M.H.; Lafforgue, G.; Mansfield, J.W.; Egea, P.R.; Bögre, L.; Grant, M. Pseudomonas syringae pv. tomato hijacks the Arabidopsis abscisic acid signalling pathway to cause disease. EMBO J. 2007, 26, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Villate, A.; Nicolas, M.S.; Gallastegi, M.; Aulas, P.-A.; Olivares, M.; Usobiaga, A.; Etxebarria, N.; Aizpurua-Olaizola, O. Review: Metabolomics as a prediction tool for plants performance under environmental stress. Plant Sci. 2020, 303, 110789. [Google Scholar] [CrossRef] [PubMed]
- Tenenboim, H.; Brotman, Y. Omic Relief for the Biotically Stressed: Metabolomics of Plant Biotic Interactions. Trends Plant Sci. 2016, 21, 781–791. [Google Scholar] [CrossRef]
- Ren, Z.; Fang, M.; Muhae-Ud-Din, G.; Gao, H.; Yang, Y.; Liu, T.; Chen, W.; Gao, L. Metabolomics analysis of grains of wheat infected and noninfected with Tilletia controversa Kühn. Sci. Rep. 2021, 11, 18876. [Google Scholar] [CrossRef]
- Dai, T.; Chang, X.; Hu, Z.; Liang, L.; Sun, M.; Liu, P.; Liu, X. Untargeted Metabolomics Based on GC-MS and Chemometrics: A New Tool for the Early Diagnosis of Strawberry Anthracnose Caused by Colletotrichum theobromicola. Plant Dis. 2019, 103, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chang, X.; Dai, T.; Li, L.; Liu, P.; Wang, G.; Liu, P.; Huang, Z.; Liu, X. Metabolic Profiling to Identify the Latent Infection of Strawberry by Botrytis cinerea. Evol. Bioinform. 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Q.; Du, L.; Hallerman, E.; Li, Y. Transcriptomic and Metabolomic Responses of Rice Plants to Cnaphalocrocis medinalis Caterpillar Infestation. Insects 2020, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Arcos, C.; Kai, M.; Svatoš, A.; Gershenzon, J.; Kunert, G. Untargeted Metabolomics Approach Reveals Differences in Host Plant Chemistry before and after Infestation with Different Pea Aphid Host Races. Front. Plant Sci. 2019, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Drès, M.; Mallet, J. Host races in plant–feeding insects and their importance in sympatric speciation. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 471–492. [Google Scholar] [CrossRef]
- Gai, Y.P.; Han, X.; Li, Y.; Yuan, C.; Mo, Y.; Guo, F.; Liu, Q.; Ji, X.L. Metabolomic analysis reveals the po-tential metabolites and pathogenesis involved in mulberry yellow dwarf disease. Plant Cell Environ. 2014, 37, 1474–1490. [Google Scholar] [CrossRef]
- Blouin, A.G.; Greenwood, D.R.; Chavan, R.R.; Pearson, M.N.; Clover, G.R.; MacDiarmid, R.M.; Cohen, D. A generic method to identify plant viruses by high-resolution tandem mass spectrometry of their coat proteins. J. Virol. Methods 2010, 163, 49–56. [Google Scholar] [CrossRef]
- Schmidt, R.; Durling, M.; de Jager, V.; Menezes, R.; Nordkvist, E.; Svatoš, A.; Dubey, M.; Lauterbach, L.; Dickschat, J.; Karlsson, M.; et al. Deciphering the genome and secondary metabolome of the plant pathogen Fusarium culmorum. FEMS Microbiol. Ecol. 2018, 94, fiy078. [Google Scholar] [CrossRef]
- Hall, R.D.; D’Auria, J.; Ferreira, A.S.; Gibon, Y.; Kruszka, D.; Mishra, P.; van de Zedde, R. High-throughput plant phenotyping: A role for metabolomics? Trends Plant Sci. 2022, 27, 549–563. [Google Scholar] [CrossRef]
- Simó, C.; Ibáñez, C.; Valdés, A.; Cifuentes, A.; García-Cañas, V. Metabolomics of genetically modified crops. Inter-Natl. J. Mol. Sci. 2014, 15, 18941–18966. [Google Scholar] [CrossRef] [PubMed]
- Bedair, M.; Glenn, K.C. Evaluation of the use of untargeted metabolomics in the safety assessment of genetically modified crops. Metabolomics 2020, 16, 111. [Google Scholar] [CrossRef]
- Fraser, P.D.; Aharoni, A.; Hall, R.; Huang, S.; Giovannoni, J.; Sonnewald, U.; Fernie, A.R. Metabolomics should be deployed in the identification and characterization of gene-edited crops. Plant J. 2020, 102, 897–902. [Google Scholar] [CrossRef]
- Buddenhagen, C.E.; James, T.K.; Ngow, Z.; Hackell, D.L.; Rolston, M.P.; Chynoweth, R.J.; Gunnarsson, M.; Li, F.; Harrington, K.C.; Ghanizadeh, H. Resistance to post-emergent herbicides is becoming common for grass weeds on New Zealand wheat and barley farms. PLoS ONE 2021, 16, e0258685. [Google Scholar] [CrossRef]
- Pomyen, Y.; Wanichthanarak, K.; Poungsombat, P.; Fahrmann, J.; Grapov, D.; Khoomrung, S. Deep metabolome: Applications of deep learning in metabolomics. Comput. Struct. Biotechnol. J. 2020, 18, 2818–2825. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Goldansaz, S.A.; Jaine, J. Translational Metabolomics: Current Challenges and Future Opportunities. Metabolites 2019, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.B.; Koch, N.; Sarsby, J.; Jones, E.; Hurst, J.L.; Beynon, R.J. Rapid identification of species, sex and maturity by mass spectrometric analysis of animal faeces. BMC Biol. 2019, 17, 66. [Google Scholar] [CrossRef]
- Lesiak, A.D.; Cody, R.B.; Dane, A.J.; Musah, R.A. Plant Seed Species Identification from Chemical Fingerprints: A High-Throughput Application of Direct Analysis in Real Time Mass Spectrometry. Anal. Chem. 2015, 87, 8748–8757. [Google Scholar] [CrossRef] [PubMed]
- Subbaraj, A.K.; Barrett, B.; Wakelin, S.; Fraser, K. Using non-targeted direct analysis in real time-mass spec-trometry (DART-MS) to discriminate seeds based on endogenous or exogenous chemicals. Anal. Bioanal. Chem. 2015, 407, 8047–8058. [Google Scholar] [CrossRef]
- Musah, R.A.; Espinoza, E.; Cody, R.; Lesiak, A.; Christensen, E.; Moore, H.; Maleknia, S.; Drijfhout, F.P. A High Throughput Ambient Mass Spectrometric Approach to Species Identification and Classification from Chemical Finger-print Signatures. Sci. Rep. 2015, 5, 11520. [Google Scholar] [CrossRef] [PubMed]
- Beyramysoltan, S.; Ventura, M.; Rosati, J.; Giffen-Lemieux, J.; Musah, R.A. Identification of the Species Con-stituents of Maggot Populations Feeding on Decomposing Remains—Facilitation of the Determination of Post Mortem Interval and Time Since Tissue Infestation through Application of Machine Learning and Direct Analysis in Real Time-Mass Spec-trometry. Anal. Chem. 2020, 92, 5439–5446. [Google Scholar] [PubMed]
- Gekenidis, M.-T.; Studer, P.; Wüthrich, S.; Brunisholz, R.; Drissner, D. Beyond the matrix-assisted laser desorption ionization (MALDI) biotyping workflow: In search of microorganism-specific tryptic peptides enabling discrimination of sub-species. Appl. Environ. Microbiol. 2014, 80, 4234–4241. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, A.; Ding, L.; Hao, Y.; Liu, H.; Cui, M.; Liu, L.; Nie, Z.; Luo, L. Direct identification and metabolomic analysis of Huanglongbing associated with Candidatus Liberibacter spp. in navel orange by MALDI-TOF-MS. Anal. Bioanal. Chem. 2020, 412, 3091–3101. [Google Scholar] [CrossRef] [PubMed]
- Gullion, J.D.; Gullion, T. Solid-State NMR Study of the Cicada Wing. J. Phys. Chem. B 2017, 121, 7646–7651. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).