Abstract
The Antarctic region is facing a higher risk of hydrocarbon pollution due to increased human activities. Compounds such as polycyclic aromatic hydrocarbons (PAHs) and heterocyclic compounds available in fuel are highly stable and can reside in the environment for prolonged periods if left untreated. The isolation of native strains is needed to develop bioremediation applications suitable for Antarctica. Strain BS19 was isolated as heterocyclic compound carbazole-degrading bacterium from Antarctic soil through culture enrichment. The 16S rRNA gene sequences identified strain BS19 as a member of the Sphingonium genus. Strain BS19 could remove 75% of carbazole after 15 days of culture at 15 °C. Whole genome sequencing resulted in incomplete genomes of 4.77 Mb in 96 contigs with the lowest GC content among Sphingobium sp. strains. The analyses revealed car gene cluster and ant genes and cat gene cluster required for the complete metabolism of carbazole as a source of carbon and energy. The comparison of the car gene cluster showed a similarity to the car gene cluster of Novosphingobium KA1. The expression of the car gene cluster was confirmed with an RT-PCR analysis indicating the involvement of the predicted genes in carbazole degradation. The findings from this study could provide more insight into developing bioremediation applications and approaches for Antarctica and other cold environments.
1. Introduction
Increased anthropogenic activities in the Antarctic region have elevated the risk of oil contamination, primarily from oil spills polluting soil and/or seawater, endangering the pristine ecosystem of the last explored continent in the world. Apart from high molecular weight aliphatic hydrocarbons (AHs), oil also contains polycyclic aromatic hydrocarbons (PAHs), which tend to reside in the environment longer than equivalent aliphatic compounds. Both AHs and PAHs have been detected and reported in Antarctic soil sediments, seawater, and aquatic bivalve species [1,2,3].
Closely related to PAHs are a group of compounds known as heterocyclic compounds. These are cyclic compounds containing at least one non-carbon atom (heteroatom) incorporated in a ring structure. The heteroatom may be oxygen, nitrogen, or sulfur. Other than petrogenic sources, heterocyclic compounds are known to originate from industries manufacturing pharmaceuticals, pesticides, and dyes [4]. Heterocyclic compounds possess a stable structure with an increased solubility than their hydrocarbon homologs, which allows them to contaminate the environment more readily [5]. The ecotoxicity and mutagenicity of heterocyclic compounds are also points for concern. A study reported that heterocyclic compounds such as dibenzothiophene, dibenzofuran, and carbazole showed a high ecotoxicity potential on algae and daphnids with an EC50 lower than 10 mg/L [6].
Bioremediation provides potential solutions to completely remove AHs, PAHs, and heterocyclic compounds from the environment [7,8,9,10]. Various approaches and techniques have been employed to ensure that bioremediation efforts lead to positive outcomes despite the challenging environmental conditions in Antarctica [11,12]. However, the application of bioremediation in Antarctica requires the use of bacteria natively present in Antarctica as dictated by the Antarctic Treaty System (ATS). Hence, it is necessary to isolate native bacterial strains and develop bioremediation applications specifically for Antarctica [13].
For bioremediation in extreme cold regions including Antarctica, several potentially useful bacterial strains have been isolated and studied. Oleispira antarctica RB-8 was isolated from crude-oil-enriched Antarctic seawater. Genomic analyses of strain RB-8 revealed three genes encoding alkane monooxygenases responsible for its alkane-degrading ability. As a psychrophilic strain, growth and oil degradation were more efficient at 4 °C than 15 °C [14]. Similarly, cold-adapted bacterial species were also reported to be responsible in hydrocarbon degradation in Antarctica. Sphingomonas sp. or Pseudomonas sp., which were isolated from hydrocarbon-contaminated soil, showed a positive ability to grow on a range of substrates within the linear alkane, monocyclic hydrocarbon, and polycyclic hydrocarbon groups [15]. Several strains belonging to Sphingomonas sp. and Pseudomonas sp. were identified as psychrothophic or psychrotolerant strains, having a wide range of growth temperatures from 4 to 30 °C.
In this study, focus is placed on the heterocyclic compound, carbazole (CAR). CAR is a tricyclic compound consisting of two benzene rings joined by a single nitrogen containing five membered rings in the middle (Figure 1A). This structure provides a rigid planar structure similar to compounds such as dibenzofuran and dibenzo-p-dioxin (Figure 1B,C). Here, we report on the isolation of carbazole-degrading bacterium from Antarctic soil. The degradation ability, growth characteristics, and genomic sequences were also studied to gain more insight into heterocyclic compounds degrading bacteria in cold environments.
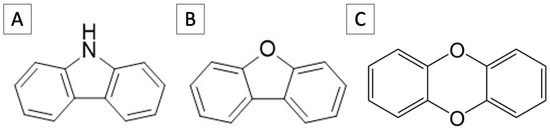
Figure 1.
Chemical structures of (A) carbazole, (B) dibenzofuran, and (C) dibenzo-p-dioxin.
2. Materials and Methods
2.1. Collection of Sample and Isolation of Strain
Soil sample was collected nearby Base General Bernardo O’Higgins Riquelme (63°19,020.600 S 57°53,053.600 W). Soil sample 3–5 cm from the surface was collected using a sterilized spatula and transferred into a sterilized 50 mL plastic tube. Collected sample was transported and stored at 4 °C until use. Approximately 500 mg was transferred into minimal salt medium (MSM) supplemented with 0.1% (w/v) carbazole as sole carbon source to start enrichment culture process. The culture was incubated at 15 °C on a 120 rpm orbital shaker for 14 days. Enrichment culture process was repeated 3 times before confirmation of single colonies were performed using minimal salt medium agar supplemented with 0.1% (w/v) CAR. Isolated strain was stored in 15% glycerol stock −80 °C freezer until use.
2.2. Bacterial Cell Enumeration
To determine the growth of isolate during the degradation of heterocyclic compounds, enumeration using the colony forming units (CFU) method was employed. Aliquots of 50 µL were sampled from culture flasks and diluted by 102 to 106 times using distilled water. Diluted samples were plated onto half-strength Luria–Bertani medium agar (5 g tryptone, 2.5 g yeast extract, 5 g NaCl, 15 g agar powder per liter) and incubated at 25 °C until colonies were visible.
2.3. Carbazole Degradation Analysis
Carbazole degradation ability of isolate was determined using gas chromatography with flame ionization detector (GC-FID, Shimadzu GC 2014, Kyoto, Japan). Aliquots were sampled every 72 h from flasks growing isolate strain in MSM supplemented with 0.1% (w/v) carbazole. Residual CAR were extracted using 1.0 mL ethyl acetate for GC-FID analysis using SH-Rtx-5 fused silica capillary column (ID: 0.53 mm, 15 m) at 250 °C at the injector, and 300 °C at the detector, with column temperature maintained at 250 °C and split-less injection configuration.
2.4. PAHs and Heterocyclic Compound Utilization Test
To assess the isolate’s ability to utilize PAHs and heterocyclic compounds other than CAR, isolate was cultured in flasks containing MSM supplemented with dibenzofuran, dibenzothiophene, fluorene, phenanthrene, and biphenyl. Flasks were incubated at 15 °C on a 120 rpm orbital shaker for 7 days. Growth from each flask was determined using CFU counts. Utilization of a specific compound was confirmed only when growth beyond 103 cfu/mL was observed in comparison to the initial day.
2.5. DNA Sequencing Analysis
2.5.1. Total DNA Extraction
Isolate was cultured in 50 mL of MSM supplemented with CAR in a shaking incubator at 15 °C for approximately 7 days. Total volume of cultured broth was transferred to centrifuge tubes, and cells were harvested at 15,000× g for 2 min. Total DNA was extracted using protocols as described [16]. Extracted DNA sample was visualized using 1.0% (w/v) agarose gel electrophoresis stained with 0.5 µg/mL ethidium bromide (Vivantis, Oceanside, CA, USA) at 100 V for 30 min. Concentration and quality of extracted total DNA sample was measured using Nanodrop spectrophotometer (Bio-Rad, Hercules, CA, USA) and Qubit dsDNA assay (Invitrogen, Carlsbad, CA, USA) prior to specific sample preparation as required by DNA sequencing service provider.
2.5.2. 16S ribosomal RNA Gene Sequence Analysis
To accurately determine the taxonomic classification of the isolated strain, the 16S ribosomal RNA gene was sequenced. PCR was performed using primer 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and primer 1492R (5′-TACGGYTACCTTGTTACGACTT-3′) [17]. The size of the PCR product of approximately 1500 bp was confirmed using 1.5% (w/v) agarose gel electrophoresis before submitting to DNA sequencing service provider Macrogen Japan Corp., Tokyo, Japan. Result from the DNA sequencing was analyzed using online standard nucleotide BLAST tool (https://blast.ncbi.nlm.nih.gov/ (accessed on 3 July 2022) and submitted to NCBI GenBank (accession no.: LC749818).
2.6. Whole Genome Sequencing, Assembly and Analyses
The whole genome of the isolated strain was sequenced using 2 × 150 base paired-end reads of Illumina HiSeq platform (Illumina, San Diego, CA, USA) performed by Azenta Life Sciences, Suzhou, China. High-quality reads trimming were performed prior to assembly and scaffolding using Velvet assembler (version 1.2.10), SSPACE (version 3.0), and GapFiller (version 1-10). To detect encoding genes for bacteria, Prodigal (version 2.6.3) was used while tRNA and rRNA were identified using tRNAscan-SE and RNAmmer. Initial gene annotation was performed using RAST (https://rast.nmpdr.org/ (accessed on 10 December 2022)). For further annotation, BLAST was utilized with NCBI non-redundant (NR) database together with confirmation using KEGG, cluster of orthologous groups of proteins (COG), Pfam, and Swiss_Prot databases [18,19,20]. Whole genome sequencing data was submitted to NCBI GenBank (accession no.: BSEF01000000).
2.7. Gene Expression Detection for Genes Involved in Carbazole Degradation
Total RNA samples were extracted from isolate grown in MSM supplemented with 0.1% (w/v) carbazole. For RT-PCR, primers were designed using Primer3 [21] to amplify approximately 650 bp fragment of gene carAa and gene catA. Designed primers were validated by confirming single amplification products of expected size for each primer set. RT–PCR was conducted as follows: 50 °C for 30 min; 94 °C for 2 min; 30 cycles of 94 °C for 2 min, 55 °C for 15 s; 72 °C for 30 s; and 72 °C for 5 min. Total RNA extractions and RT-PCR preparations were conducted as reported previously [14]. The primer sets used were CarAa-Fw (5′-agtgctattccaagaacaccatatc-3′) and CARAa-Rv (5′-aagtagaggtggctggtctcat-3′) specific for carAa gene, CatA-Fw (5′-gaagaccgtatcaagaatattgtcc-3′) and CatA-Rv (5′-gatgttgatctgtgtcgtcaaatag-3′) specific for antA gene, and 27F and 1492R specific for 16S rRNA.
3. Results
3.1. Isolation and Identification of Carbazole-Degrading Bacterium from Antarctic Soil
During the enrichment culture process, the degradation of carbazole was determined primarily by visually observing the reduction in insoluble carbazole particles and the increase in turbidity due to cell growth. Carbazole is hydrophobic and remains as insoluble white powder in MSM until degraded by bacterial isolate. After three cycles of enrichment culture, uniform colonies were observed on MSM agar supplemented with carbazole as the sole carbon source. The colonies were convex, smooth, approximately 0.5 mm in diameter, and bright yellow in color. When grown on half-strength Luria–Bertani medium agar, the colonies formed appeared to be light cream colored. This isolate was designated as strain BS19.
The growth temperature for strain BS19 was characterized and confirmed from temperatures ranging from 10 °C to 30 °C. At 5 °C, strain BS19 grew at a significantly lower rate. The 16S ribosomal RNA gene sequencing result determined approximately 1400 bp of sequence which was used to compare with taxonomically meaningful sequences from curated type strains using EZBioCloud [22]. Closely related strains are listed in Table 1. Strain BS19 was determined to belong to the Sphingobium genus.

Table 1.
Strains with highest similarities to strain BS19 when analyzed with EZBioCloud.
3.2. Degradation of Carbazole and Utilization of Similar Compounds
Carbazole degradation by strain BS19 was determined by quantitatively measuring the residual carbazole concentration in the MSM medium using GC-FID, and cell growth was measured using CFU counts. The residual carbazole was reduced to 25% after 15 days of culture while bacterial cell concentration increased from 105 to the highest of 108 on day 9 (Figure 2). The growth of strain BS19 utilizing dibenzofuran, dibenzothiophene, fluorene, phenanthrene, and biphenyl was also confirmed.
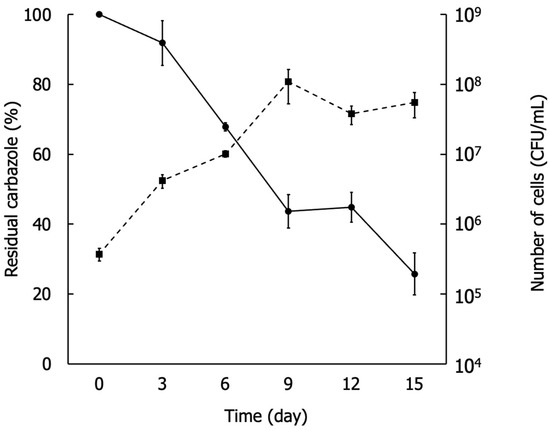
Figure 2.
Degradation of carbazole analysis by measuring residual carbazole (•) and bacterial cell growth by CFU count (▪).
3.3. Whole Genome Sequencing of Strain BS19
The results from the whole genome sequencing of strain BS19 is summarized in Table 2. The genome sequencing yielded an incomplete genome of 4,773,423 base pairs in 96 contigs. The average GC content was 60.51%. A total of 4690 genes were predicted with 4 rRNA genes and 47 tRNA genes identified. When compared to all completely assembled genomes from the Sphingobium sp. obtained from the NCBI database, the genome size and number of protein coding sequence of strain BS19 were typical, as the average genome size for Sphingobium sp. was 4.92457 Mb with the average number of coding sequence being 4597 (Table 3). However, the total genomic GC content for strain BS19 was the lowest.
Table 2.
Genomic features of Sphingobium sp. BS19.
Table 3.
Comparison of genomic features of strain BS19 with all completely assembled genomes from strains within Sphingobium genus obtained from NCBI database.
3.4. Gene Function Annotation
The gene function annotation was performed by comparing the predicted protein sequences to the protein contained in the database. The best results of the alignment (E-value < 0.05 and contain more than 60% of reference sequences when using BLAST) were chosen. The genus/species classification was confirmed by comparing the analysis results with the species information provided by the NR annotation database (Figure 3). The annotated genes showed the highest identity to the genes from the Sphingobium genus with 57.86% being from unclassified Sphingobium species.
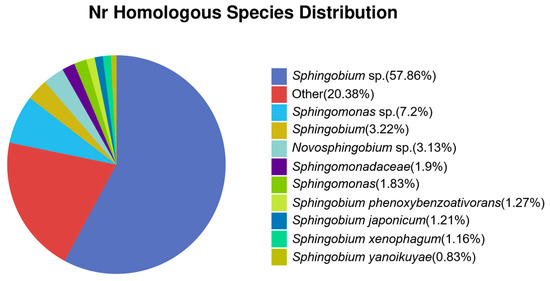
Figure 3.
Homologous species distribution from strain BS19 gene annotation using NR database.
For functional prediction, the genes are annotated to COGs using the COG database. Each gene was assigned to a single COG based on the best E-value and was classified according to protein function (Figure 4). A total of 3424 genes were assigned to COGs with 1668 COG clusters identified.
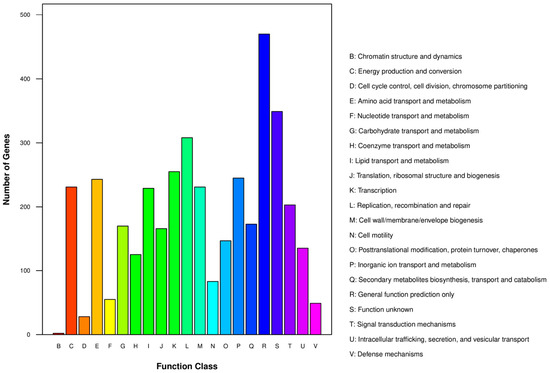
Figure 4.
COG functional classification of genes from strain BS19.
3.5. Identification of Gene Clusters Involved in Carbazole Degradation Pathway
The car gene cluster and cat gene cluster were identified (accession no.: LC749817) and predicted to be responsible in completely metabolizing carbazole (Table 4). Both clusters were separated by 6563 bp. The car gene cluster contained eight genes encoding for transcriptional regulator (carR), carbazole 1,9a-dioxygenase (carA), 2-subunit meta-cleavage enzyme (carBa, carBb), meta-cleavage compound hydrolase (carC), and ferredoxin (carAc). The cat gene cluster, containing genes encoding for catechol 1,2-dioxygenase (catA), cis,cis-muconate cycloisomerase (catB), muconolactone isomerase (catC), and transcriptional regulator (catR), was predicted to be involved in catechol metabolism. The genes predicted to be encoding large subunit (antA) and small subunit (antB) of anthranilate 1,2-dioxygenase were also identified.
Table 4.
Identified genes and gene clusters predicted to be involved in the complete metabolism of carbazole in strain BS19.
The comparison of the structure for the carbazole gene cluster from strain BS19 to well-studied car gene clusters is shown in Figure 5. The structure from strain BS19 was found to be similar with the car cluster I from Novosphingobium sp. KA1 [35]. The search for ferredoxin reductase (carAd) within strain BS19 genome revealed two genes identified as pyridine nucleotide-disulfide oxidoreductase (Sbs19_13590 and Sbs19_42570) to be highly similar to carAd from strain KA1.
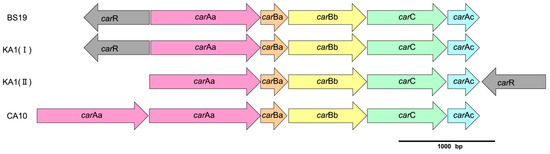
Figure 5.
Structures of car gene clusters from strain BS19, Novosphingobium sp. KA1 and Pseudomonas resinovorans CA10.
3.6. Detection of Car Gene and Cat Gene Cluster Expression
The expression of the car gene cluster and cat gene cluster were confirmed using reverse-transcription PCR using the total RNA extracted from strain BS19 grown in MSM, supplemented with carbazole as the sole carbon source. A PCR run was conducted using genomic DNA first to confirm the validity of the selected primers. The expression of genes from the car cluster was confirmed when the carAa gene was detected. The band corresponding to the catA gene from the cat gene cluster was not observed (Figure 6).
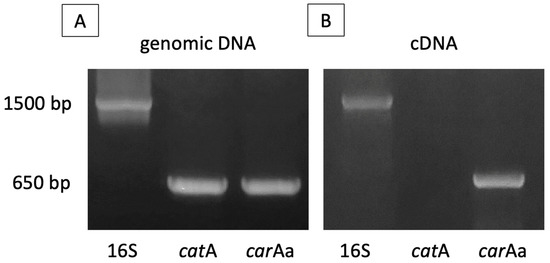
Figure 6.
(A): Agarose gel electrophoresis of PCR product of 16S rRNA, catA and carAa gene from genomic DNA of strain BS19 grown in MSM supplemented with carbazole. (B): Agarose gel electrophoresis of reverse-transcription-based PCR product (cDNA) of the same genes from total RNA extracted.
4. Discussion
Strain BS19 was isolated as carbazole-degrading bacteria from the Antarctic soil sample using the culture enrichment method. The 16S ribosomal RNA gene sequence confirmed strain BS19 taxonomic association as Sphingobium sp. matching closest to Sphingobium algorifonticola TLA-22T at 97.945%. This was further verified when CDS annotation using the genome sequencing data revealed at least 57.86% of genes match the genes from Sphingobium sp. The isolated strains from the Sphingobium genus have been isolated from various environmental samples such as soil, wastewater, and activated sludge, with many of them studied for their ability to degrade hydrocarbons and xenobiotics. This highlights the potential and versatility for Sphingobium sp. to be utilized in bioremediation applications.
Several Sphingobium species were also reported to be psychrotolerant and cold-active. Sphingobium sp. VY55, which was isolated from the highly oligothophic water of the permanently ice covered Lake Vanda in Antarctica [36], could adapt to a wide range of growth temperatures from 0 to 33 °C with the optimum growth occurring at 20 °C. Additionally, as summarized in Table 3, strain KCTC 72723 and strain PAMC28499 were isolated from the Antarctic soil and the glaciers of Uganda, respectively. Sphingobium sp. strains are not recognized as psychrophilic bacteria according to the definition by Gounot [37]. On the contrary, general members of the Sphingobium genus are identified as mesophiles. Strain BS19 was isolated and studied at 15 °C but can be grown at temperatures up to 30 °C, placing it in the psychrotrophic group of microorganisms. Even though investigation on the genetical differences between the psychrotrophic (psychrotolerant) strains and mesophilic strains of Sphingobium sp. is beyond the scope of this study, it may provide insight into understanding bacterial adaptation in cold environments.
The degradation ability of carbazole is an important parameter if strain BS19 were to be used for bioremediation application. Previously, Nocardioides sp. strain IC177, which was isolated from soil, degraded 18% of carbazole in four days [38]. However, the particularly rapid degrader Pseudomonas sp. strain CA06 and strain CA10 showed 100% degradation of carbazole within 20 h [39]. Similarly, seven Pseudomonas sp. isolates (strain BC039—strain BC046) completely utilized all carbazole within 24 h in addition to having denitrification ability [40]. In comparison to these isolates, strain BS19 showed inferior degradation ability with 75% carbazole removal within 15 days. However, carbazole degradation experiments for strain BS19 were conducted at 15 °C, which is significantly lower compared to previously reported mesophilic isolates. Despite the variability of experimental conditions between studies leading to difficulty in direct comparisons, it is believed that strain BS19 is well adapted to low temperatures with the ability to degrade carbazole at 5 °C.
The car gene cluster of P. resinovorans strain CA10 was the first reported and most extensively studied set of genes related to carbazole metabolisms in bacteria [41]. This gene cluster contains a total of 10 genes (carAaAaBaBbCAcAdDFE) which are responsible for encoding enzymes involved in the conversion of carbazole and 2-hydroxypenta-2,4-dienoate. Carbazole is converted to anthranilate and 2-hydroxypenta-2,4-dienoate through the activities of the terminal dioxygenase or carbazole 1,9a-dioxygenase (encoded by carAa), 2-subunit meta-cleavage enzyme (encoded by carBa and carBb), and meta-cleavage compound hydrolase (encoded by carC). Through anthranilate 1,2-dioxygenase (encoded by antA and antB), anthranilate is converted to catechol, which is further metabolized following the catechol degradation pathway involving enzymes encoded by genes catA, catB and catC. On the other hand, the complete metabolism of 2-hydroxypenta-2,4-dienoate is achieved through the 2-hydroxypenta-2,4-dienoate degradation pathway involving enzymes encoded by carD, carE and carF. The 2-hydroxypenta-2,4-dienoate degradation pathway is also commonly utilized during the complete metabolism of other xenobiotic aromatic hydrocarbon compounds such as toluene, ethylbenzene, dioxin, and biphenyl. Catechol and 2-hydroxypenta-2,4-dienoate are two metabolites shared among many aromatic compound degradation pathways, and are not exclusive to the carbazole degradation pathway.
The complete genomic sequencing of strain CA10 revealed a 199,035 bp plasmid pCAR1 harboring the car gene cluster and ant gene cluster [42]. However, the cat gene cluster is not available in plasmid pCAR1. Another carbazole degrader Novosphingobium sp. strain KA1 was also reported to possess a plasmid carrying car genes designated as plasmid pCAR3 [43]. In contrast to the plasmid originated from strain CA10, strain KA1 harbors two clusters of car degradation genes (car-I and car-II), and gene cluster (for anthranilate degradation) and cat gene cluster in its pCAR3 plasmid. In a separate study, the pCAR1 plasmid was shown to be transferred conjugatively Pseudomonas and non-Pseudomonas strains when carbazole was added [44]. This result indicated that car gene clusters are distributed widely across different genera of bacteria through the horizontal gene transfer mechanism, similar to the extensively reported toluene and xylene degradation genes carried by TOL plasmids [45].
In this study, whole genome sequencing of strain BS19 yielded incomplete genomic sequencing, therefore limiting the ability to identify key genes and operons involved in the degradation of xenobiotics. Despite this limitation, sufficient reads were obtained to identify the car gene cluster and compare gene cluster structures with extensively reported car gene clusters from the Pseudomonas resinovorans strain CA10 [42] and Novosphingobium sp. strain KA1 [43]. The car gene cluster of strain BS19 showed the highest similarity with car-I gene cluster from strain KA1 due to the position of the carR gene. This result also assisted in identifying two copies of the carAd gene encoding ferredoxin reductase, which is required for the activity of carbazole 1,9a-dioxygenase (carAa). However, further studies are required to confirm the functionality and compatibility of these predicted carAd genes.
The localization of the car gene cluster in strain BS19 cannot be confirmed with the current incomplete sequencing result. Alternatively, it is possible to distinguish whether bacterial strains possess plasmids involved in specific functions (namely, degradation of aromatic compounds, virulence, and antibiotic resistance) using the plasmid curing experiment. Assuming the existence of pCAR1 or pCAR3-like plasmid, an attempt for plasmid curing of strain BS19 was conducted. However, the result was inconclusive. Regardless of this result, there are compelling reasons to believe that strain BS19 could be harboring the car genes on a plasmid, as apart from strain PAMC28499 and strain LF-16, all other completely sequenced Sphingobium sp. strains possess at least one plasmid.
Sphingobium sp. Ant-17, which was isolated from oil-contaminated Antarctic soil, is believed to be involved in xenobiotics degradation processes at low temperatures [46]. While strain Ant-17 was reported to be able to grow on phenanthrene and naphthalene, a simple search through the genome revealed multiple genes encoding uncharacterized ring hydroxylating dioxygenase, which may indicate its ability to utilize more PAHs. On the other hand, strain BS19 was confirmed to also grow on dibenzofuran, dibenzothiophene, fluorene, phenanthrene, and biphenyl. However, genes responsible for the metabolism of these compounds were not found in the sequenced genome. A complete genome sequence is required to uncover more genes involved in xenobiotics degradation and to fully understand the potential for strain BS19.
The almost-complete set of carbazole, anthranilate, and catechol metabolism genes of strain BS19 offer convincing molecular confirmation of the ability to completely metabolize carbazole as source of carbon and energy. It was also predicted that strain BS19 would metabolize carbazole through an identical pathway as described for CAR-SF [47]. Regardless, efforts to confirm gene expression through RT-PCR was inconclusive, as only the carAa gene was detected but not the catA gene. The existence of multiple catechol metabolism operon may be a possible explanation for this result. Hence, obtaining a completed genome sequence is a requirement to study the involvement of these metabolic genes.
5. Conclusions
Strain BS19 was isolated as a carbazole degrader from Antarctic soil through culture enrichment. The carbazole-degrading ability of strain BS19 at low temperatures together with its ability to utilize dibenzofuran, dibenzothiophene, fluorene, phenanthrene, and biphenyl showed promising potential for future bioremediation applications. The genomic sequence of strain BS19 revealed the molecular basis for metabolizing carbazole completely. The car genes, ant genes and cat genes were organized in clusters typically seen in previously reported aromatic compound degrading bacteria. This indicated the widespread distribution of xenobiotic aromatic compound metabolism genes even to the Antarctic region. The expression of carbazole 1,9a-dioxygenase (encoded carA) was confirmed when grown with carbazole, which confirmed the functionality of car genes in strain BS19. However, due to the incomplete genomic data, the functional analyses of genes in this study were limited. Future studies are necessary to improve our understanding of heterocyclic compound degrading bacteria in cold environments.
Author Contributions
Conceptualization, A.Z.; methodology, K.S., S.T. and A.Z.; software, K.S. and S.T.; validation, S.A.A., C.G.-F. and A.Z.; formal analysis, K.S., S.T., C.G.-F. and A.Z.; investigation, K.S.; resources, S.A.A. and A.Z.; writing—original draft preparation, K.S. and A.Z.; writing—review and editing, S.A.A., C.G.-F., K.S. and A.Z.; supervision, S.A.A. and A.Z.; project administration, A.Z.; funding acquisition, S.A.A. and A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
S.A. Ahmad is supported by YPASM Smart Partnership Initiative (6300247) by Sultan Mizan Antarctic Research Foundation (YPASM). C.G. Fuentes is supported by Centro de Investigacion y Monitoreo Ambiental Antàrctico (CIMAA) Project.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This project was funded by the Sultan Mizan Antarctic Research Foundation (YPASM), Universiti Putra Malaysia (UPM), Center for Research and Antarctic Environmental Monitoring (CIMAA) and Shibaura Institute of Technology (SIT).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bícego, M.C.; Zanardi-Lamardo, E.; Taniguchi, S.; Martins, C.C.; da Silva, D.A.M.; Sasaki, S.T.; Albergaria-Barbosa, A.C.R.; Paolo, F.S.; Weber, R.R.; Montone, R.C. Results from a 15-Year Study on Hydrocarbon Concentrations in Water and Sediment from Admiralty Bay, King George Island, Antarctica. Antarct. Sci. 2009, 21, 209–220. [Google Scholar] [CrossRef]
- Martins, C.C.; Bícego, M.C.; Taniguchi, S.; Montone, R.C. Aliphatic and Polycyclic Aromatic Hydrocarbons in Surface Sediments in Admiralty Bay, King George Island, Antarctica. Antarct. Sci. 2004, 16, 117–122. [Google Scholar] [CrossRef]
- Cripps, G.C.; Priddle, J. Hydrocarbon Content of an Antarctic Infaunal Bivalve—Historical Record or Life Cycle Changes? Antarct. Sci. 1995, 7, 127–136. [Google Scholar] [CrossRef]
- Sims, G.K.; O’Loughlin, E.J.; Crawford, R.L. Degradation of Pyridines in the Environment. Crit. Rev. Environ. Control 1989, 19, 309–340. [Google Scholar] [CrossRef]
- Padoley, K.V.; Mudliar, S.N.; Pandey, R.A. Heterocyclic Nitrogenous Pollutants in the Environment and Their Treatment Options—An Overview. Bioresour. Technol. 2008, 99, 4029–4043. [Google Scholar] [CrossRef]
- Eisentraeger, A.; Brinkmann, C.; Hollert, H.; Sagner, A.; Tiehm, A.; Neuwoehner, J. Heterocyclic compounds: Toxic effects using algae, daphnids, and the salmonella/microsome test taking methodical quantitative aspects into account. Environ. Toxicol. Chem. 2008, 27, 1590. [Google Scholar] [CrossRef]
- Abdulrasheed, M.; Zakaria, N.N.; Roslee, A.F.A.; Shukor, M.Y.; Zulkharnain, A.; Napis, S.; Convey, P.; Alias, S.A.; Gonzalez-Rocha, G.; Ahmad, S.A. Biodegradation of Diesel Oil by Cold-Adapted Bacterial Strains of Arthrobacter spp. from Antarctica. Antarct. Sci. 2020, 32, 5. [Google Scholar] [CrossRef]
- Maliki, I.M.; Abdul-Manas, N.H.; Ahmad, S.A.; Fuse, H.; Ramírez-Moreno, N.; Zulkharnain, A. Removal of Heterocyclic Compound Carbazole Using Cell Immobilization of Thalassospira Profundimaris Strain M02. Rev. Mex. Ing. Quim. 2020, 20, 413–422. [Google Scholar] [CrossRef]
- Zahri, K.N.M.; Zulkharnain, A.; Gomez-Fuentes, C.; Sabri, S.; Ahmad, S.A. Study of Growth Kinetics of Antarctic Bacterial Community for Biodegradation of Waste Canola Oil. Desalination Water Treat 2021, 213, 128–138. [Google Scholar] [CrossRef]
- Lee, G.L.Y.; Ahmad, S.A.; Yasid, N.A.; Zulkharnain, A.; Convey, P.; Wan Johari, W.L.; Alias, S.A.; Gonzalez-Rocha, G.; Shukor, M.Y. Biodegradation of Phenol by Cold-Adapted Bacteria from Antarctic Soils. Polar. Biol. 2018, 41, 553–562. [Google Scholar] [CrossRef]
- Roslee, A.F.A.; Ahmad, S.A.; Gomez-Fuentes, C.; Shaharuddin, N.A.; Khalil, K.A.; Zulkharnain, A. Scientometric Analysis of Diesel Pollutions in Antarctic Territories: A Review of Causes and Potential Bioremediation Approaches. Sustainability 2021, 13, 7064. [Google Scholar] [CrossRef]
- Lim, Z.S.; Wong, R.R.; Wong, C.-Y.; Zulkharnain, A.; Shaharuddin, N.A.; Ahmad, S.A. Bibliometric Analysis of Research on Diesel Pollution in Antarctica and a Review on Remediation Techniques. Appl. Sci. 2021, 11, 1123. [Google Scholar] [CrossRef]
- Verasoundarapandian, G.; Wong, C.-Y.; Shaharuddin, N.A.; Gomez-Fuentes, C.; Zulkharnain, A.; Ahmad, S.A. A Review and Bibliometric Analysis on Applications of Microbial Degradation of Hydrocarbon Contaminants in Arctic Marine Environment at Metagenomic and Enzymatic Levels. Int. J. Environ. Res. Public. Health 2021, 18, 1671. [Google Scholar] [CrossRef] [PubMed]
- Gentile, G.; Bonsignore, M.; Santisi, S.; Catalfamo, M.; Giuliano, L.; Genovese, L.; Yakimov, M.M.; Denaro, R.; Genovese, M.; Cappello, S. Biodegradation Potentiality of Psychrophilic Bacterial Strain Oleispira Antarctica RB-8 T. Mar. Pollut. Bull. 2016, 105, 125–130. [Google Scholar] [CrossRef]
- Aislabie, J.; Foght, J.; Saul, D. Aromatic Hydrocarbon-Degrading Bacteria from Soil near Scott Base, Antarctica. Polar. Biol. 2000, 23, 183–188. [Google Scholar] [CrossRef]
- Lee, G.L.Y.; Zakaria, N.N.; Futamata, H.; Suzuki, K.; Zulkharnain, A.; Shaharuddin, N.A.; Convey, P.; Zahri, K.N.M.; Ahmad, S.A. Metabolic Pathway of Phenol Degradation of a Cold-Adapted Antarctic Bacteria, Arthrobacter sp. Catalysts 2022, 12, 1422. [Google Scholar] [CrossRef]
- Sung, J.Y.; Hwang, Y.; Shin, M.H.; Park, M.S.; Lee, S.H.; Yong, D.; Lee, K. Utility of Conventional Culture and MALDI-TOF MS for Identification of Microbial Communities in Bronchoalveolar Lavage Fluid in Comparison with the GS Junior Next Generation Sequencing System. Ann. Lab. Med. 2018, 38, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.J. A Genomic Perspective on Protein Families. Science (1979) 1997, 278, 631–637. [Google Scholar] [CrossRef]
- Makarova, K.; Wolf, Y.; Koonin, E. Archaeal Clusters of Orthologous Genes (ArCOGs): An Update and Application for Analysis of Shared Features between Thermococcales, Methanococcales, and Methanobacteriales. Life 2015, 5, 818–840. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic. Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Kasai, D.; Masai, E.; Katayama, Y.; Fukuda, M. Degradation of 3-O-Methylgallate in Sphingomonas Paucimobilis SYK-6 by Pathways Involving Protocatechuate 4,5-Dioxygenase. FEMS Microbiol. Lett. 2007, 274, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-R.; Jang, S.-M.; Chi, Y.M.; Kim, B.; Jung, S.-H.; Lee, Y.M.; Uetake, J.; Lee, J.H.; Park, H.; Oh, T.-J. Complete Genome Sequence of Sphingobium sp. Strain PAMC 28499 Reveals a Potential for Degrading Pectin with Comparative Genomics Approach. Genes Genom. 2020, 42, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hong, Q.; Li, Q.; Li, C.; Cao, L.; Sun, J.-Q.; Yan, X.; Li, S.-P. Characterization of Isoproturon Biodegradation Pathway in Sphingobium sp. YBL2. Int. Biodeterior. Biodegrad. 2012, 70, 8–13. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lee, P.C. Complete Genome Sequence of Yellow Pigment-Producing Sphingobium sp. Strain HAL-16. Microbiol Resour. Announc. 2020, 9, 41. [Google Scholar] [CrossRef]
- Tabata, M.; Ohhata, S.; Kawasumi, T.; Nikawadori, Y.; Kishida, K.; Sato, T.; Ohtsubo, Y.; Tsuda, M.; Nagata, Y. Complete Genome Sequence of a γ-Hexachlorocyclohexane Degrader, Sphingobium sp. Strain TKS, Isolated from a γ-Hexachlorocyclohexane-Degrading Microbial Community. Genome Announc. 2016, 4, 2. [Google Scholar] [CrossRef]
- Jiang, W.; Zhang, M.; Gao, S.; Zhu, Q.; Qiu, J.; Yan, X.; Xin, F.; Jiang, M.; Hong, Q. Comparative Genomic Analysis of Carbofuran-Degrading Sphingomonads Reveals the Carbofuran Catabolism Mechanism in Sphingobium sp. Strain CFD-1. Appl. Environ. Microbiol. 2022, 88, 22. [Google Scholar] [CrossRef]
- Jiang, W.; Gao, Q.; Zhang, L.; Wang, H.; Zhang, M.; Liu, X.; Zhou, Y.; Ke, Z.; Wu, C.; Qiu, J.; et al. Identification of the Key Amino Acid Sites of the Carbofuran Hydrolase CehA from a Newly Isolated Carbofuran-Degrading Strain Sphingbium sp. CFD-1. Ecotoxicol. Environ. Saf. 2020, 189, 109938. [Google Scholar] [CrossRef]
- Rangu, S.S.; Muralidharan, B.; Tripathi, S.C.; Apte, S.K. Tributyl Phosphate Biodegradation to Butanol and Phosphate and Utilization by a Novel Bacterial Isolate, Sphingobium sp. Strain RSMS. Appl. Microbiol. Biotechnol. 2014, 98, 2289–2296. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Brower, A.L.; Xu, Z.-J.; Xu, Y.; Spain, J.C.; Zhou, N.-Y. Molecular Basis and Evolutionary Origin of 1-Nitronaphthalene Catabolism in Sphingobium sp. Strain JS3065. Appl. Environ. Microbiol. 2023, 89, 1. [Google Scholar] [CrossRef] [PubMed]
- Tabata, M.; Ohhata, S.; Nikawadori, Y.; Sato, T.; Kishida, K.; Ohtsubo, Y.; Tsuda, M.; Nagata, Y. Complete Genome Sequence of a γ-Hexachlorocyclohexane-Degrading Bacterium, Sphingobium sp. Strain MI1205. Genome Announc. 2016, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Fixen, K.R.; Starkenburg, S.R.; Hovde, B.T.; Johnson, S.L.; Deodato, C.R.; Daligault, H.E.; Davenport, K.W.; Harwood, C.S.; Cattolico, R.A. Genome Sequences of Eight Bacterial Species Found in Coculture with the Haptophyte Chrysochromulina Tobin. Genome Announc. 2016, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liang, B.; Qi, M.; Yun, H.; Shi, K.; Li, Z.; Guo, Y.; Yan, P.; Liu, S.-J.; Wang, A. Novel Pathway for Chloramphenicol Catabolism in the Activated Sludge Bacterial Isolate Sphingobium sp. CAP-1. Environ. Sci. Technol. 2020, 54, 7591–7600. [Google Scholar] [CrossRef] [PubMed]
- Nojiri, H.; Nam, J.-W.; Kosaka, M.; Morii, K.-I.; Takemura, T.; Furihata, K.; Yamane, H.; Omori, T. Diverse Oxygenations Catalyzed by Carbazole 1,9a-Dioxygenase from Pseudomonas sp. Strain CA10. J. Bacteriol. 1999, 181, 3105–3113. [Google Scholar] [CrossRef]
- vander Schaaf, N.; Cunningham, A.; Cluff, B.; Kraemer, C.; Reeves, C.; Riester, C.; Slater, L.; Madigan, M.; Sattley, W. Cold-Active, Heterotrophic Bacteria from the Highly Oligotrophic Waters of Lake Vanda, Antarctica. Microorganisms 2015, 3, 391–406. [Google Scholar] [CrossRef]
- Gounot, A.-M. Psychrophilic and Psychrotrophic Microorganisms. Experientia 1986, 42, 1192–1197. [Google Scholar] [CrossRef]
- Inoue, K.; Habe, H.; Yamane, H.; Omori, T.; Nojiri, H. Diversity of Carbazole-Degrading Bacteria Having the Car Gene Cluster: Isolation of a Novel Gram-Positive Carbazole-Degrading Bacterium. FEMS Microbiol. Lett. 2005, 245, 145–153. [Google Scholar] [CrossRef]
- Ouchiyama, N.; Zhang, Y.; Omori, T.; Kodama, T. Biodegradation of Carbazole by Pseudomonas Spp. CA06 and CA10. Biosci. Biotechnol. Biochem. 1993, 57, 455–460. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, Y.; Li, X.; Wen, D.; Tang, X. Biodegradation of Carbazole by the Seven Pseudomonas sp. Strains and Their Denitrification Potential. J. Hazard Mater. 2011, 190, 253–259. [Google Scholar] [CrossRef]
- Nojiri, H.; Sekiguchi, H.; Maeda, K.; Urata, M.; Nakai, S.I.; Yoshida, T.; Habe, H.; Omori, T. Genetic Characterization and Evolutionary Implications of a Car Gene Cluster in the Carbazole Degrader Pseudomonas sp. Strain CA10. J. Bacteriol. 2001, 183, 12. [Google Scholar] [CrossRef]
- Maeda, K.; Nojiri, H.; Shintani, M.; Yoshida, T.; Habe, H.; Omori, T. Complete Nucleotide Sequence of Carbazole/Dioxin-Degrading Plasmid PCAR1 in Pseudomonas Resinovorans Strain CA10 Indicates Its Mosaicity and the Presence of Large Catabolic Transposon Tn4676. J. Mol. Biol. 2003, 326, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Shintani, M.; Urata, M.; Inoue, K.; Eto, K.; Habe, H.; Omori, T.; Yamane, H.; Nojiri, H. The Sphingomonas Plasmid PCAR3 Is Involved in Complete Mineralization of Carbazole. J. Bacteriol. 2007, 189, 2007–2020. [Google Scholar] [CrossRef] [PubMed]
- Shintani, M.; Fukushima, N.; Tezuka, M.; Yamane, H.; Nojiri, H. Conjugative Transfer of the IncP-7 Carbazole Degradative Plasmid, PCAR1, in River Water Samples. Biotechnol. Lett. 2008, 30, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Assinder, S.J.; Williams, P.A. The TOL Plasmids: Determinants of the Catabolism of Toluene and the Xylenes. Adv. Microb. Physiol. 1990, 31, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, E.M.; Guerrero, L.D.; Makhalanyane, T.P.; Aislabie, J.M.; Cowan, D.A. Draft Genome Sequence of the Aromatic Hydrocarbon-Degrading Bacterium Sphingobium sp. Strain Ant17, Isolated from Antarctic Soil. Genome Announc. 2014, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, H.; Zulkharnain, A.B.; Maeda, R.; Fuse, H.; Iwata, K.; Omori, T. Cloning and Nucleotide Sequences of Carbazole Degradation Genes from Marine Bacterium Neptuniibacter sp. Strain CAR-SF. Curr. Microbiol. 2010, 61, 50–56. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).