

Alternative Cover Crops and Soil Management Practices Modified the Macronutrients, Enzymes Activities, and Soil Microbial Diversity of Rainfed Olive Orchards (cv. Chetoui) under Mediterranean Conditions in Tunisia
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
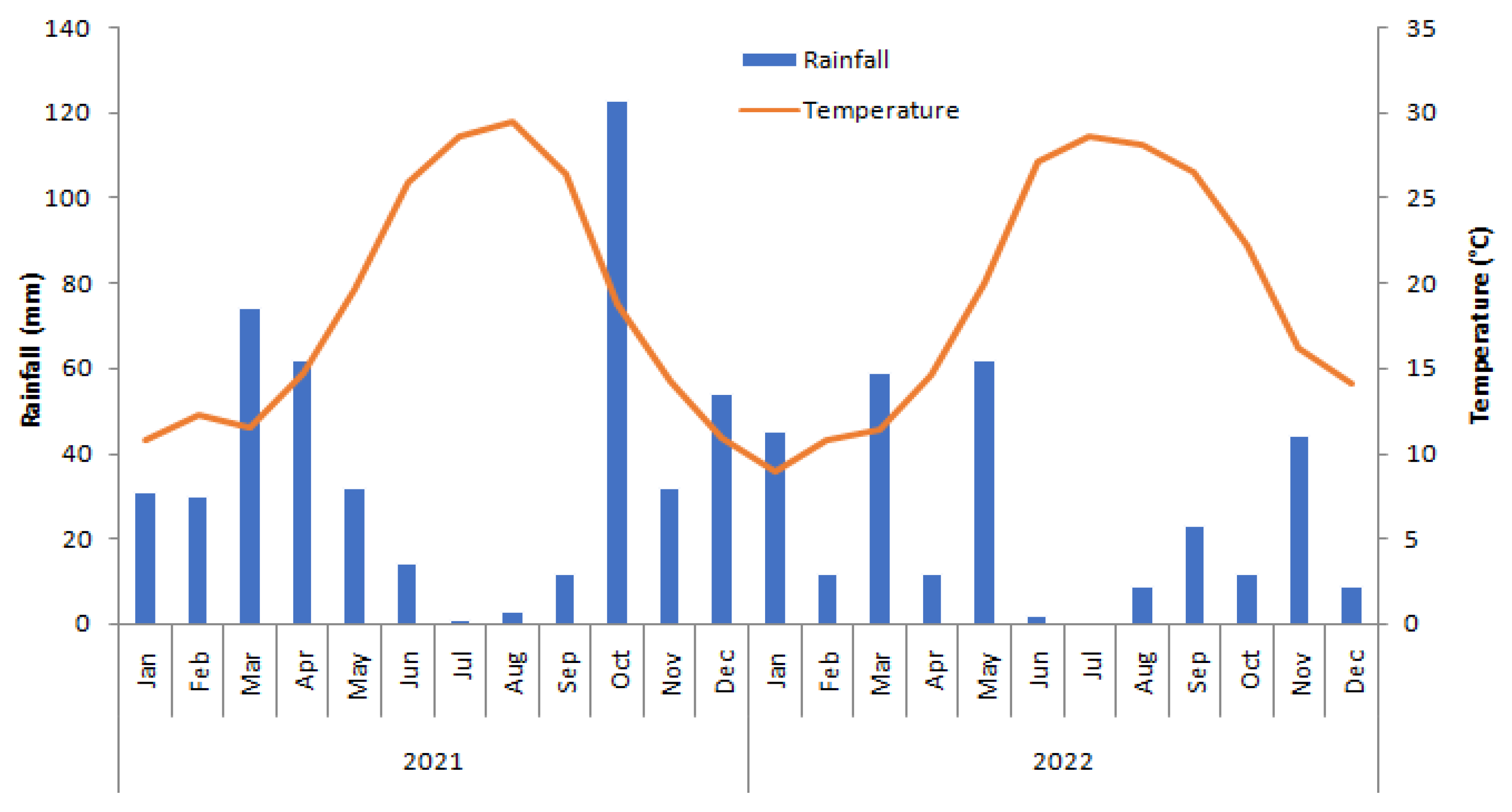
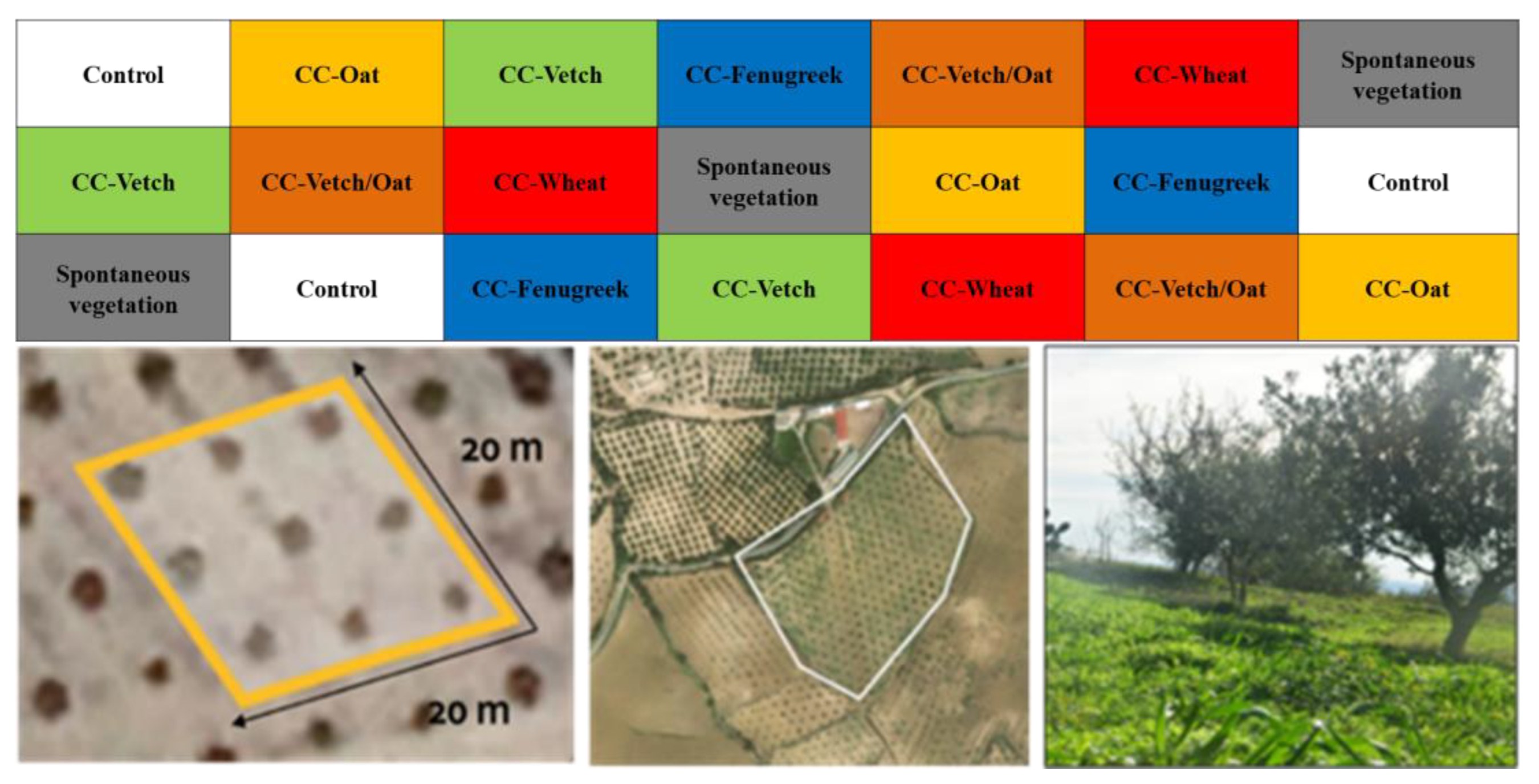
2.1. Description of the Study Area and Experimental Design
2.2. Experimental Design
2.3. Cover Crop Sampling and Analysis
2.4. Soil Sampling
2.5. Soil Properties
2.6. Soil Functional Quality
2.6.1. Enzymatic Activity
2.6.2. BiologTM Ecoplate and Soil Microbial Community Catabolic Profiling
2.7. Statistical Analysis
3. Results
3.1. Dry Biomass of Cover Crops and Residues Incorporated into the Soil
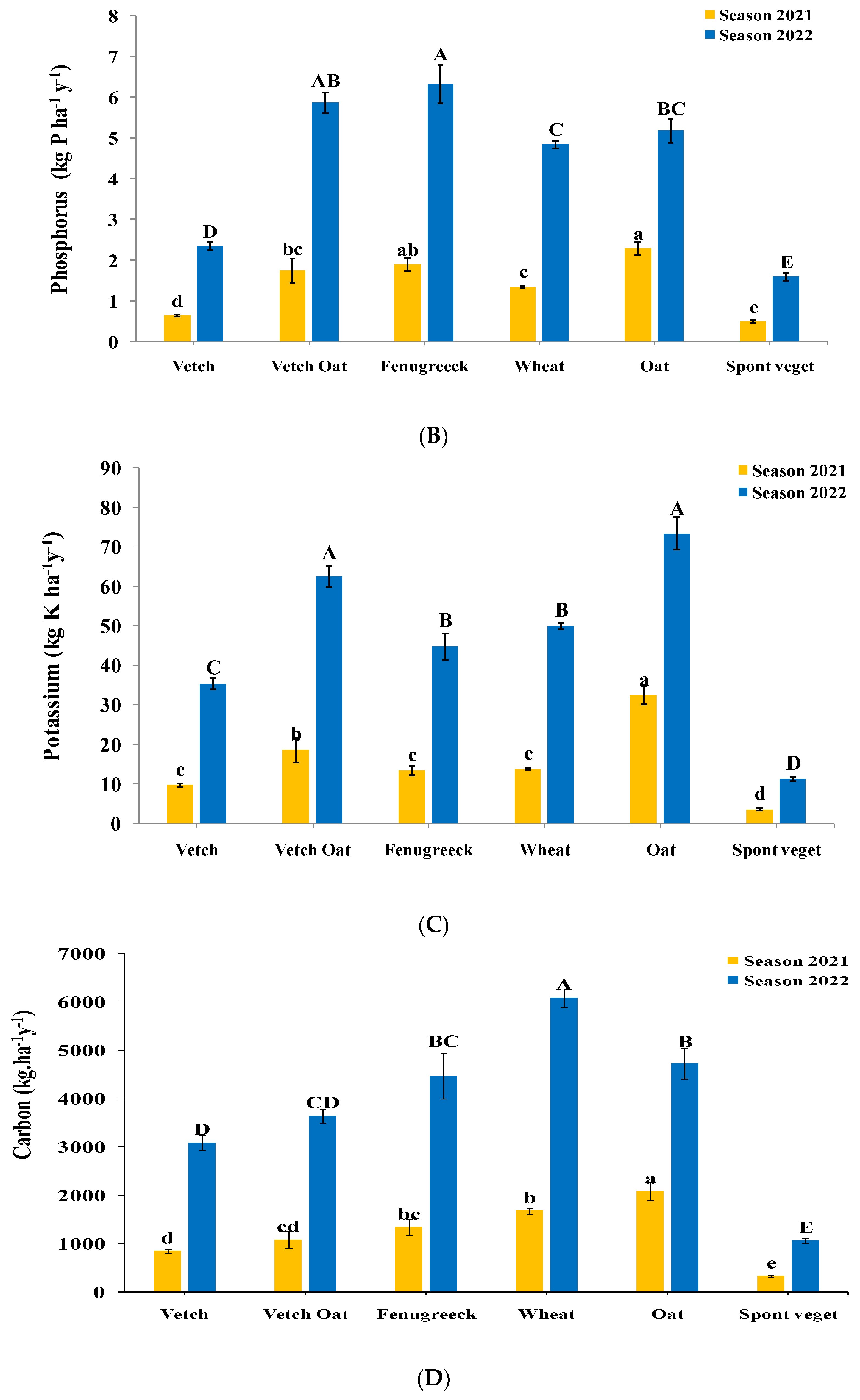
3.2. Nutrient and Carbon Retention by Cover Crops and Residues Incorporated in the Soil
3.3. Soil Organic Matter and Macronutrient Availabilities after the Incorporation of Residues and Cover Crops
3.4. Soil Functional Quality
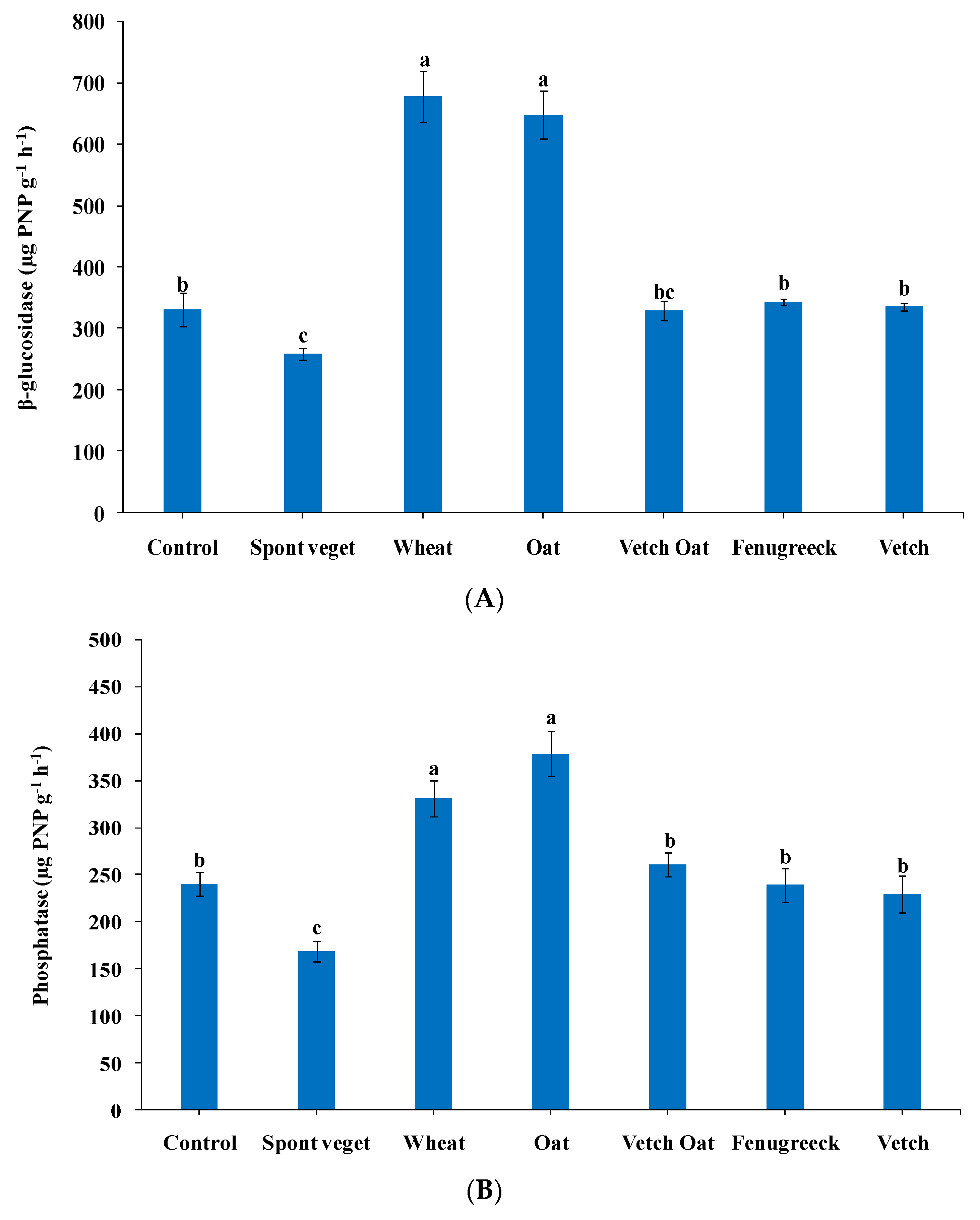
3.4.1. Soil Enzyme Activities
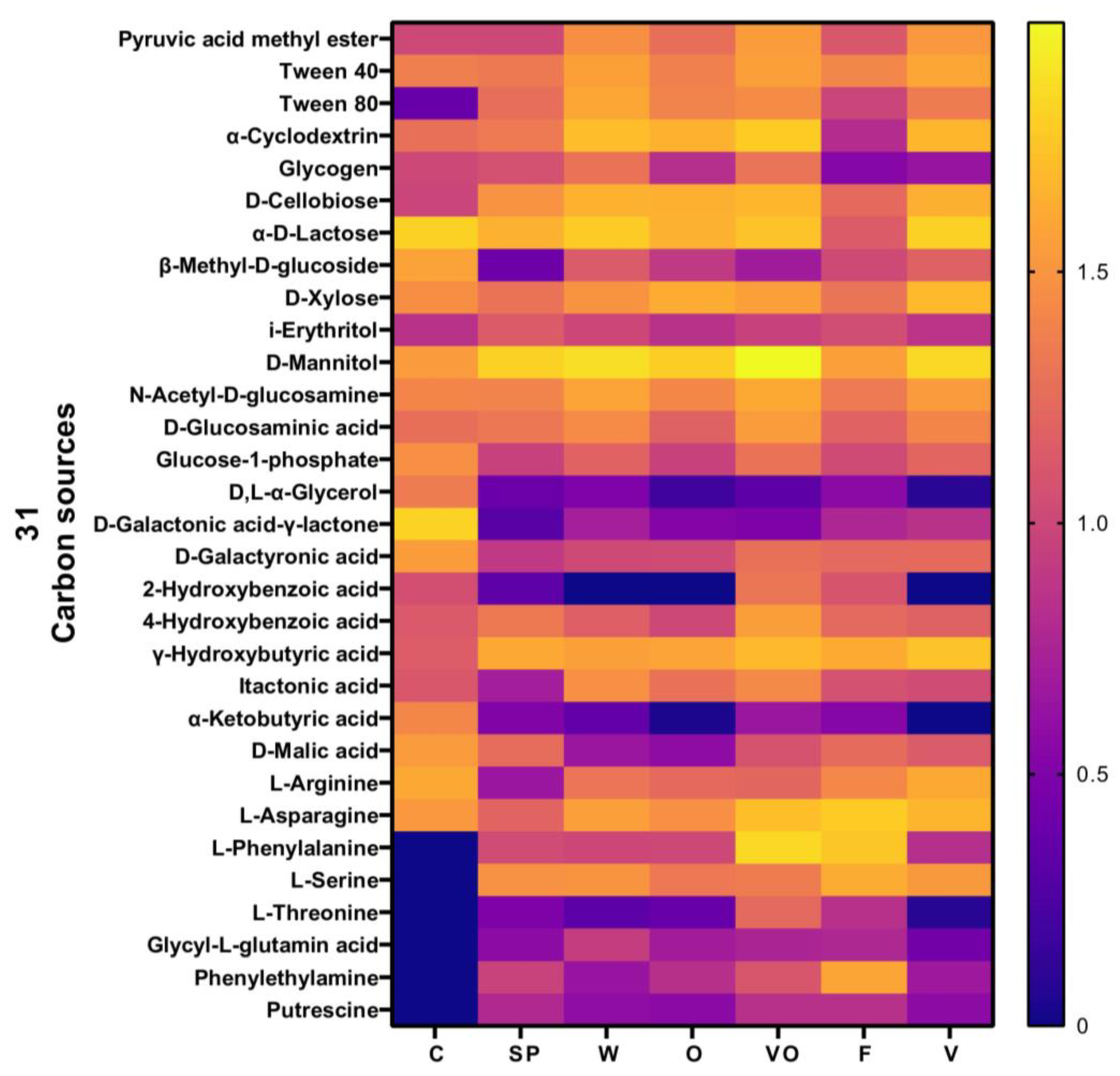
3.4.2. Carbon Utilization Sources
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OLIVAE. Official Journal of the International Olive Council. 2017. Available online: www.internationaloliveoil.org (accessed on 13 April 2024).
- Ben Abdallah, S.; Elfkih, S.; Suárez-Rey, E.M.; Parra-López, C.; Romero-Gámez, M. Evaluation of the environmental sustainability in the olive growing systems in Tunisia. J. Clean. Prod. 2021, 282, 124526. [Google Scholar] [CrossRef]
- Elfkih, S.; Hadiji, O.; Ben Abdallah, S.; Boussadia, O. Water Accounting for Food Security: Virtual Water and Water Productivity in the Case of Tunisian Olive Oil Value Chain. Agriculture 2023, 13, 1205. [Google Scholar] [CrossRef]
- DGPA. Document de la Direction Générale de la Production Agricole en Tunisie; DGPA: Tunis, Tunisia, 2021. [Google Scholar]
- ONH. Rapport de l’Office National de l’Huile en Tunisie; ONH: Tunis, Tunisia, 2023. [Google Scholar]
- Gargouri, K.; Mhiri, A. Relalionship between soil fertility, phosphorus and potassium nutrition on the olive in Tunisia. Opt. Mediterr. 2003, 50, 199–204. [Google Scholar]
- Chehab, H.; Tekaya, M.; Ouhibi, M.; Gouiaa, M.; Zakhama, H.; Mahjoub, Z.; Laamari, S.; Sfina, H.; Chihaoui, B.; Boujnah, D.; et al. Effects of compost, olive mill wastewater and legume cover cropson soil characteristics, tree performance and oil quality of olive trees cv. Chemlali grown under organic farming system. Sci. Hortic. 2019, 253, 163–171. [Google Scholar] [CrossRef]
- Issaoui, M.; Flamini, G.; Brahmi, F.; Dabbou, S.; Ben Hassine, K.; Taamali, A.; Chehab, H.; Ellouz, M.; Zarrouk, M.; Hammami, M. Effect of the growing area conditions on differentiation between Chemlali and Chétoui olive oils. Food Chem. 2010, 119, 220–225. [Google Scholar] [CrossRef]
- Ben Youssef, N.; Zarrouk, W.; Carrasco-Pancorbo, A.; Zarrouk, M. Effect of olive ripeness on chemical properties and phenolic composition of Chétoui virgin olive oil. J. Sci. Food Agric. 2010, 90, 199–204. [Google Scholar] [CrossRef]
- Li, T.; Wang, Y.; Kamran, M.; Chen, X.; Tan, H.; Long, M. Effects of Grass Inter-Planting on Soil Nutrients, Enzyme Activity, and Bacterial Community Diversity in an Apple Orchard. Front. Plant Sci. 2022, 13, 901143. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.; Patterson, J. How can science policy help to deliver the global goals. Guard. Sci. Policy Blog. 2015, 10, 17–25. [Google Scholar]
- Koudahe, K.; Allen, S.C.; Djaman, K. Critical review of the impact of cover crops on soil properties. Int. Soil Water Conserv. Res. 2022, 10, 343–354. [Google Scholar] [CrossRef]
- Piccolo, A. The Nature of Soil Organic Matter and Innovative Soil Managements to Fight Global Changes and Maintain Agricultural Productivity. In Carbon Sequestration in Agricultural Soils A Multidisciplinary Approach to Innovative Methods; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–19. [Google Scholar]
- Blanco-Canqui, H.; Mikha, M.M.; Presley, D.R.; Claassen, M.M. Addition of Cover Crops Enhances No-Till Potential for Improving Soil Physical Properties. Soil Sci. Soc. Am. J. 2011, 75, 1471–1482. [Google Scholar] [CrossRef]
- Beniaich, A.; Guimarães, V.D.; Avanzi, J.C.; Silva, B.M.; Salvador, F.A.G.; Santos, W.P.; Silva, M.L.N. Spontaneous vegetation as an alternative to cover crops in olive orchards reduces water erosion and improves soil physical properties under tropical conditions. Agric. Water Manag. 2023, 279, 108186. [Google Scholar] [CrossRef]
- Ordóñez-Fernández, R.; de Torres, M.A.R.R.; Márquez-García, J.; Moreno-García, M.; Carbonell-Bojollo, R.M. Legumes used as cover crops to reduce fertilization problems improving soil nitrate in an organic orchard. Eur. J. Agron. 2018, 95, 1–13. [Google Scholar] [CrossRef]
- Huertas, A.J.; Cuartero, J.; Ros, M.; Pascual, J.A.; Parras-Alcántara, L.; González-Rosado, M.; Özbolat, O.; Zornoza, R.; Egea-Cortines, M.; Hurtado-Navarro, M.; et al. How binomial (traditional rainfed olive grove-Crocus sativus) crops impact the soil bacterial community and enhance microbial capacities. J. Environ. Manag. 2023, 3023, 345. [Google Scholar]
- Adetunji, A.T.; Ncube, B.; Mulidzi, R.; Lewu, F.B. Management impact and benefit of cover crops on soil quality: A review. Soil Tillage Res. 2020, 204, 104717. [Google Scholar] [CrossRef]
- Chehab, H.; Tekaya, M.; Gouiaa, M.; Mahjoub, Z.; Laamari, S.; Sfina, H.; Chihaoui, B.; Boujnah, D.; Mechri, B. The use of legume and grass cover crops induced changes in ion accumulation, growth and physiological performance of young olive trees irrigated with high-salinity water. Sci. Hortic. 2018, 232, 170–174. [Google Scholar] [CrossRef]
- Ovalle, C.; Del Pozo, A.; Peoples, M.B.; Lavín, A. Estimating the contribution of nitrogen from legume cover crops to the nitrogen nutrition of grapevines using a 15 N dilution technique. Plant Soil 2010, 334, 247–259. [Google Scholar] [CrossRef]
- Rodrigues, M.Â.; Correia, C.M.; Claro, A.M.; Ferreira, I.Q.; Barbosa, J.C.; Moutinho-Pereira, J.M.; Bacelar, E.A.; Fernandes-Silva, A.A.; Arrobas, M. Soil nitrogen availability in olive orchards after mulching legume cover crop residues. Sci. Hortic. 2013, 158, 45–51. [Google Scholar] [CrossRef]
- Murray, J.D.; Liu, C.W.; Chen, Y.; Miller, A.J. Nitrogen sensing in legumes. J. Exp. Bot. 2017, 68, 1919–1926. [Google Scholar] [CrossRef]
- Sofo, A.; Palese, A.; Maria, C.; Teresa, C.; Giuseppe, R.P.; Curci, M.; Crecchio, C.; Xiloyannis, C. Genetic, Functional, and Metabolic Responses of Soil Microbiota in a Sustainable Olive Orchard. Soil Sci. 2010, 175, 81–88. [Google Scholar] [CrossRef]
- Sofo, A.; Ciarfaglia, A.; Scopa, A.; Camele, I.; Curci, M.; Crecchio, C.; Xiloyannis, C.; Palese, A.M. Soil microbial diversity and activity in a Mediterranean olive orchard using sustainable agricultural practices. Soil Use Manag. 2014, 30, 160–167. [Google Scholar] [CrossRef]
- Montes-Borrego, M.; Metsis, M.; Blanca, B.; Landa, B.B. Arbuscular Mycorhizal Fungi Associated with the Olive Crop across the Andalusian Landscape: Factors Driving Community Differentiation. PLoS ONE 2014, 9, e96397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xue, W.; Xue, J.; Zhang, J.; Qiu, L.; Chen, X.; Hu, F.; Kardol, P.; Liu, M. Leveraging functional traits of cover crops to coordinate crop productivity and soil health. J. Appl. Ecol. 2022, 59, 2627–2641. [Google Scholar] [CrossRef]
- Bernnan, B.E.; Veronica, A.M. Soil microbial biomass and enzyme data after six years of cover crop and compost treatments in organic vegetable production. Soil Sci. Soc. Am. J. 2019, 83, 624–637. [Google Scholar] [CrossRef]
- Herencia, J.F. Enzymatic activities under different cover crop management in a Mediterranean olive orchard. Biol. Agric. Hortic. 2015, 31, 45–52. [Google Scholar] [CrossRef]
- Henriquez, C.; Uribe, L.; Valenciano, A.; Nogales, R. Soil enzyme activity-dehidrogenase, ß-glucosidase, Phosphatase and urease-under different crops. Agron. Costarric. 2014, 38, 43–54. [Google Scholar]
- Weber, K.; Legge, R. One-dimensional metric for tracking bacterial community divergence using sole carbon source utilization patterns. J. Microbiol. Methods 2009, 79, 55–61. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis: Microbiological and Biochemical Properties: Part 2; SSSA Book Ser; Soil Science Society of America, Inc.: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Grzadiel, J.; Furtak, K.; Galazka, A. Community-Level physiological profiles of microorganism from different types of soil that are characteristic to Poland-a long-term microplot experiment. Sustainability 2019, 11, 56. [Google Scholar] [CrossRef]
- Ben Zineb, A.; Barkaoui, K.; Karray, F.; Mhiri, N.; Sayadi, S.; Mliki, A.; Gargouri, M. Olive agroforestry shapes rhizosphere microbiome networks associated with annual crops and impacts the biomass production under low-rainfed conditions. Front. Microbiol. 2022, 13, 977797. [Google Scholar] [CrossRef]
- Amassaghrou, A.; Barkaoui, K.; Bouaziz, A.; Alaoui, S.B.; Fatemi, Z.E.A.; Daoui, K. Yield and related traits of three legume crops grown in olive-based agroforestry under an intense drought in the South Mediterranean. Saudi J. Biol. Sci. 2023, 30, 103597. [Google Scholar] [CrossRef]
- Temani, F.; Bouaziz, A.; Daoui, K.; Wery, J.; Barkaoui, K. Olive agroforestry can improve land productivity even under low water availability in the South Mediterranean. Agric. Ecosyst. Environ. 2021, 307, 107234. [Google Scholar] [CrossRef]
- Torrús-Castillo, M.; Domouso, P.; Herrera-Rodríguez, J.M.; Calero, J.; García-Ruiz, R. Aboveground Carbon Fixation and Nutrient Retention in Temporary Spontaneous Cover Crops in Olive Groves of Andalusia. Front. Environ. Sci. 2022, 10, 868410. [Google Scholar] [CrossRef]
- Arruda, B. Manipulation of the soil microbiome regulates the colonization of plants by arbuscular mycorrhizal fungi. Mycorrhiza 2021, 31, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.Â.; Dimande, P.; Pereira, E.L.; Ferreira, I.Q.; Freitas, S.; Correia, C.M.; Moutinho-Pereira, J.; Arrobas, M. Early-maturing annual legumes: An option for cover cropping in rainfed olive orchards. Nutr. Cycl. Agroecosyst. 2015, 103, 153–166. [Google Scholar] [CrossRef]
- Soriano, M.; Cabezas, J.M.; Gomez, A.J. Field evaluation of selected autochthonous herbaceous species for cover crops in Mediterranean woody crops. Eur. J. Agron. 2023, 143, 126723. [Google Scholar] [CrossRef]
- Tul, S.; Manolikaki, I.; Digalaki, N.; Psarras, G.; Koufakis, I.; Kalaitzaki, A.; Sergentani, C.; Koubouris, G. Contribution of a Seeded Cover Crop Mixture on Biomass Production and Nutrition Status Compared to Natural Vegetation in a Mediterranean Olive Grove. Int. J. Plant Biol. 2022, 13, 235–244. [Google Scholar] [CrossRef]
- Gomez, A.J. Sustainability using cover crops in Mediterranean trees crops, olives and vines-challenges and current knowledge. Hung. Geogr. Bull. 2017, 66, 13–28. [Google Scholar]
- Ferreira, I.Q.; Rodrigues, M.Â.; Claro, A.M.; Arrobas, M. Management of Nitrogen-Rich Legume Cover Crops as Mulch in Traditional Olive Orchards. Commun. Soil Sci. Plant Anal. 2015, 46, 1881–1894. [Google Scholar] [CrossRef]
- Stein, S.; Hartung, J.; Perkons, U.; Möller, K.; Zikeli, S. Plant and soil N of different winter cover crops as green manure for subsequent organic white cabbage. Nutr. Cycl. Agroecosyst. 2023, 127, 10306–10309. [Google Scholar] [CrossRef]
- Lee, A.; Neuberger, P.; Omokanye, A.; Hernandez-Ramirez, G.; Kim, K.; Gorzelak, A. Arbuscular mycorrhizal fungi in oat-pea intercropping. Sci. Rep. 2023, 13, 390. [Google Scholar] [CrossRef]
- Afzal, A.; Bano, A. Rhizobium and phosphate solubilizing bacteria improve the yield and phosphorus uptake in wheat (Ttiticum aestivum). Int. J. Agric. Biol. 2010, 1814, 9596. [Google Scholar]
- Rodrigues, M.; Withers, P.J.A.; Soltangheisi, A.; Vargas, V.; Holzschuh, M.; Pavinato, P.S. Tillage systems and cover crops affecting soil phosphorus bioavailability in Brazilian Cerrado Oxisols. Soil Tillage Res. 2021, 205, 104770. [Google Scholar] [CrossRef]
- De Torres, M.A.R.R.; Carbonell, B.R.; Alcantara, B.C.; Rodriguez, L.A.; Ordonèz, F.R. Carbon sequestration potential of residues of different types of cover crops in olive groves under Mediterranean climate. Span. J. Argic. Res. 2012, 10, 649–661. [Google Scholar] [CrossRef]
- Fernandes, G.; Marques, A.; César, R.; Ribeiro, B.R.; Cardoso Paula, S. Potassium Uptake Kinetics in Native Forage Grass Species From Pampa Biome. Soil Sci. Cienc. Rural 2022, 52, 8478cr2020. [Google Scholar] [CrossRef]
- Guesmi, H.; Aichi, H.; Bendhafer, G.; Fouad, Y.; Menasseri, S.; Chaar, H. Assessment of radial variation of soil properties in an olive tree-barley/common vetch agroforestry system under low-input conditions in a Tunisian semi-arid climate after three cropping seasons. J. Res. Environ. Earth Sci. 2022, 11, 330–339. [Google Scholar]
- Porwollik, V.; Rolinski, S.; Heinke, J.; Von Bloh, W.; Schaphoff, S.; Müller, C. The role of cover crops for cropland soil carbon, nitrogen leaching, and agricultural yields -A global simulation study with LPJmL (V. 5.0-tillage-cc). Biogeosciences 2022, 19, 957–977. [Google Scholar] [CrossRef]
- Rattan, L. Regenerative agriculture for food and climate. J. Soil Water Conserv. 2020, 75, 123A. [Google Scholar]
- Wang, Y.; Huang, Q.; Gao, H.; Zhang, R.; Yang, L.; Guo, Y.; Li, H.; Awasthi, M.K.; Li, G. Long-term cover crops improved soil phosphorus availability in a rain-fed apple orchard. Chemosphere 2021, 275, 130093. [Google Scholar] [CrossRef] [PubMed]
- Hansen, V.; Müller-Stöver, D.; Gómez-Muñoz, B.; Oberson, A.; Magid, J. Differences in cover crop contributions to phosphorus uptake by ryegrass in two soils with low and moderate P status. Geoderma 2022, 426, 116075. [Google Scholar] [CrossRef]
- Ferreira, M.A.; Jose’, M.P.; Carlos, C.; Angelo, R.M. Olive response to potassium applications under different water regimes and cultivars Isabel Q. Nutr. Cycl. Agroecosyst. 2018, 112, 387–401. [Google Scholar] [CrossRef]
- Chavarría, D.N.; Verdenelli, R.A.; Serri, D.L.; Restovich, S.B.; Andriulo, A.E.; Meriles, J.M.; Vargas-Gil, S. Effect of cover crops on microbial community structure and related enzyme activities and macronutrient availability. Eur. J. Soil Biol. 2016, 76, 74–82. [Google Scholar] [CrossRef]
- Martín, M.P.R.; Fernández-Ondoño, E.; Ortiz-Bernad, I.; Abreu, M.M. Influence of Intensive and Super-Intensive Olive Grove Management on Soil Quality—Nutrients Content and Enzyme Activities. Plants 2023, 12, 2779. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Sekaran, U.; Wang, T.; Kumar, S. On-farm assessment of cover cropping effects on soil C and N pools, enzyme activities, and microbial community structure. J. Agric. Sci. 2021, 159, 216–226. [Google Scholar] [CrossRef]
- Stegarescu, G.; Reintam, E.; Tõnutare, T. Cover crop residues effect on soil structural stability and phosphatase activity. Acta Agric. Scand. Sect. B Soil Plant Sci. 2021, 71, 992–1005. [Google Scholar] [CrossRef]
- Peregrina, F.; Pérez-Álvarez, E.P.; García-Escudero, E. The short-term influence of aboveground biomass cover crops on C sequestration and β-glucosidase in a vineyard ground under semiarid conditions. Span. J. Agric. Res. 2014, 12, 1000–1007. [Google Scholar] [CrossRef]
- Navas, M.; Benito, M.; Rodríguez, I.; Masaguer, A. Effect of five forage legume covers on soil quality at the Eastern plains of Venezuela. Appl. Soil Ecol. 2011, 49, 242–249. [Google Scholar] [CrossRef]
- Roper, M.M.; Ophel-Keller, K.M. Soil microflora as bio-indicators of soil health. In Biological Indicators of Soil Health; Pankhurst, C.E., Doube, B.M., Gupta, V.V.S.R., Eds.; CAB International: New York, NY, USA, 1997. [Google Scholar]
- Marais, A.; Labuschagne, J.; Booyse, M. Influence of oats cover crop preceding dryland lucerne establishment on some aspects of soil microbial ecology. S. Afr. J. Plant Soil 2020, 37, 87–89. [Google Scholar] [CrossRef]
- Gomez, M.B.; Pittroff, S.M.; De Neergaard, A.; Jensen, L.S.; Nicolaisen, M.H.; Magid, J. Penicillium bilaii effects on maize growth and P uptake from soil and localized sewage sludge in a rhizobox experiment. Biol. Fertil. Soils 2017, 53, 23–35. [Google Scholar] [CrossRef]
- Lucas, M.M. The Symbiosome: Legume and Rhizobia Co-evolution toward a Nitrogen-Fixing Organelle. Front. Plant Sci. 2018, 22, 2229. [Google Scholar]
- Chinthalapudi, D.P.M.; Pokhrel, S.; Kingery, W.L.; Shankle, M.W.; Ganapathi Shanmugam, S. Exploring the Synergistic Impacts of Cover Crops and Fertilization on Soil Microbial Metabolic Diversity in Dryland Soybean Production Systems Using Biolog EcoPlates. Appl. Biosci. 2023, 2, 328–346. [Google Scholar] [CrossRef]
- Siczek, A.; Lipiec, J. Impact of faba bean-seed rhizobial inoculation on microbial activity in the rhizosphere soil during growing season. Int. J. Mol. Sci. 2016, 17, 784. [Google Scholar] [CrossRef]
- Aguilera-Huertas, J.; Parras-Alcántara, L.; González-Rosado, M.; Lozano-García, B. Intercropping in rainfed Mediterranean olive groves contributes to improving soil quality and soil organic carbon storage. Agric. Ecosyst. Environ. 2024, 361, 108826. [Google Scholar] [CrossRef]
- Pantigoso, H.A.; Manter, D.K.; Fonte, S.J.; Vivanco, J.M. Root exudate-derived compounds stimulate the phosphorus solubilizing ability of bacteria. Sci. Rep. 2023, 13, 4050. [Google Scholar] [CrossRef]
- Samuel, I.; Haruna, N.N. Influence of Cover Crop, Tillage, and Crop Rotation Management on Soil Nutrients. Soil Biol. Biochem. 2020, 25, 142–149. [Google Scholar]
- Michael, F.D. Key roles of microsymbiont amino acid metabolism in rhizobia-legume interactions. Crit. Rev. Microbiol. 2015, 44, 411–451. [Google Scholar]
- Doud, D.D.; Naghahashi, G.; Pfeffer, P.F.; Kayser, W.M.; Reider, C. On farm production and utilization of arbuscular mycorrhizal fungi inoculums. Can. J. Plant Sci. 2005, 85, 15–21. [Google Scholar]
- Lehman, R.M.; Taheri, W.I.; Osborne, S.L.; Buer, J.S.; Doud, D.D. Fall cover cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Appl. Soil Ecol. 2012, 61, 300–304. [Google Scholar] [CrossRef]
- Tekaya, M.; Dabbaghi, O.; Guesmi, A.; Attia, F.; Chehab, H.; Khezami, L.; Algathami, F.; Ben Hamadi, N.; Hammami, M.; Prinsen, E.; et al. Arbuscular mycorrhizas modulate carbohydrate, phenolic compounds and hormonal metabolism to enhance water deficit tolerance of olive trees (Olea europaea). Agric. Water Manag. 2022, 274, 107947. [Google Scholar]
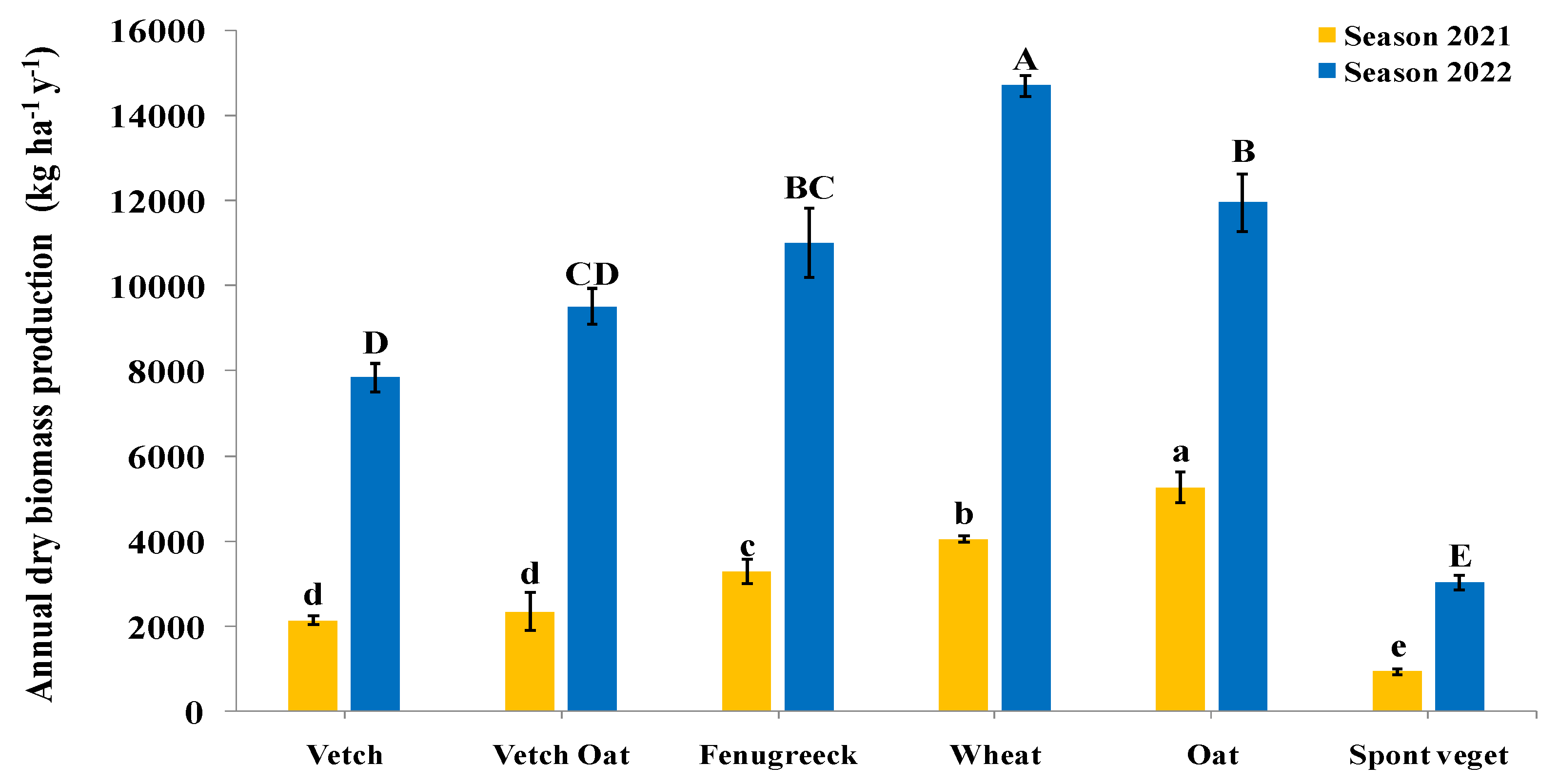




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | SOM (%) | Total N (%) | Available P (µg g−1) | Available K (mg g−1) | ||||
|---|---|---|---|---|---|---|---|---|
| Season | 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | 2021 | 2022 |
| Control | 1.62 ± 0.03 b | 2.59 ± 0.04 e | 0.088 ± 0.00 b | 0.1 ± 0.02 b | 2.68 ± 0.05 c | 2.62 ± 0.04 d | 2.85 ± 0.03 c | 2.48 ± 0.07 d |
| Spont. | 3.03 ± 0.03 a | 3.08 bc | 0.157 ± 0.039 a | 0.154 ± 0.02 a | 3.82 ± 0.02 ab | 4.08 ± 0.09 c | 3.41 ± 0.03 bc | 3.01 ± 0.03 c |
| Wheat | 2.84 ± 0.05 a | 3.72 ± 0.043 b | 0.143 ± 0.02 a | 0.163 ± 0.06 a | 4.12 ± 0.12 a | 5.88 ± 0.07 a | 3.95 ± 0.09 abc | 2.87 ± 0.07 c |
| Oat | 2.5 ± 0.02 a | 4.23 ± 0.02 a | 0.1 ± 0.02 b | 0.168 ± 0.02 a | 3.34 ± 0.06 b | 4.55 ± 0.04 bc | 4.50 ± 0.09 ab | 2.80 ± 0.04 c |
| Vetch–oat | 3.24 ± 0.05 a | 3.51 ± 0.04 bc | 0.157 ± 0.03 a | 0.168 ± 0.03 a | 3.38 ± 0.09 b | 4.70 ± 0.06 b | 4.82 ± 0.07 a | 3.39 ±0.03 ab |
| Fenugreek | 3.04 ± 0.05 a | 3.42 ± 0.03 c | 0.143 ± 0.04 a | 0.133 ± 0.02 a | 3.48 ± 0.11 b | 4.13 ± 0.09 c | 3.06 ± 0.087 c | 3.54 ± 0.05 a |
| Vetch | 3.07 ± 0.01 a | 3.17 ± 0.03 d | 0.150 ± 0.017 a | 0.143 ± 0.04 a | 3.81 ± 0.05 ab | 4.80 ± 0.07 b | 2.91 ± 0.073 c | 3.07 ±0.05 bc |
| ACWD | H’ | S | E | |
|---|---|---|---|---|
| Control | 1.21 ± 0.12 a | 4.70 ± 0.09 a | 30.0 ± 1.00 a | 1.38 ± 0.03 a |
| Non-tilled | 1.03 ± 0.37 a | 4.70 ± 0.25 a | 28.0 ± 4.36 a | 1.41 ± 0.01 a |
| Wheat | 1.16 ± 0.12 a | 4.77 ± 0.02 a | 29.3 ± 0.58 a | 1.41 ± 0.01 a |
| Oat | 1.04 ± 0.11 a | 4.71 ± 0.07 a | 28.3 ± 2.08 a | 1.41 ± 0.01 a |
| Vetch–oat | 1.31 ± 0.11 a | 4.84 ± 0.09 a | 30.3 ± 1.15 a | 1.42 ± 0.01 a |
| Fenugreek | 1.23 ± 0.65 a | 4.80 ± 0.18 a | 29.0 ± 9.85 a | 1.42 ± 0.01 a |
| Vetch | 1.11 ± 0.02 a | 4.68 ± 0.05 a | 27.6 ± 1.15 a | 1.41 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elhaddad, F.; González, J.A.C.; Abdelhamid, S.; Garcia-Ruiz, R.; Chehab, H. Alternative Cover Crops and Soil Management Practices Modified the Macronutrients, Enzymes Activities, and Soil Microbial Diversity of Rainfed Olive Orchards (cv. Chetoui) under Mediterranean Conditions in Tunisia. Sustainability 2024, 16, 5329. https://doi.org/10.3390/su16135329
Elhaddad F, González JAC, Abdelhamid S, Garcia-Ruiz R, Chehab H. Alternative Cover Crops and Soil Management Practices Modified the Macronutrients, Enzymes Activities, and Soil Microbial Diversity of Rainfed Olive Orchards (cv. Chetoui) under Mediterranean Conditions in Tunisia. Sustainability. 2024; 16(13):5329. https://doi.org/10.3390/su16135329
Chicago/Turabian StyleElhaddad, Fadoua, Julio Antonio Calero González, Sofiane Abdelhamid, Roberto Garcia-Ruiz, and Hechmi Chehab. 2024. "Alternative Cover Crops and Soil Management Practices Modified the Macronutrients, Enzymes Activities, and Soil Microbial Diversity of Rainfed Olive Orchards (cv. Chetoui) under Mediterranean Conditions in Tunisia" Sustainability 16, no. 13: 5329. https://doi.org/10.3390/su16135329