Abstract
As valuable sources of plant-based protein, leguminous vegetables (grain legumes) are essential for global food security and contribute to body growth and development in humans as well as animals. Climate change is a major challenge for agriculture development that creates major problems for the growth and development of plants. However, legume productivity is threatened by climate change factors, including rising temperatures, shifting precipitation patterns, increased atmospheric carbon dioxide levels, intensified extreme events, and altered pest/pathogen activity. This review synthesizes approximately 136 studies to assess the climate effects on major legume crops. Under all the global emissions trajectories, the mean temperatures are projected to rise beyond the optimal legume growing thresholds by 2050, carrying yield reductions between 10 and 49% for beans, soybeans, cowpeas, and lentils without adaptation measures. The elevated carbon dioxide may transiently enhance the yields up to 18%, but the benefits dramatically decline above 550 ppm and cannot offset the other climate impacts. Altered rainfall along with recurrent drought and heat waves are also expected to decrease the legume crop yields, seed quality, and soil nitrogen levels worldwide. Furthermore, the proliferation of legume pests and fungal diseases poses significant risks, amplified by climate shifts in 84% of the reviewed studies. These multifaceted impacts threaten the productivity gains in leguminous vegetables essential to sustainably meeting the global protein demand. Realizing resilience will require the accelerated development of heat/drought-tolerant legume varieties, enhanced climate-informed agronomic practices, strong policy interventions, and social safety nets explicitly supporting legume producers, in addition to the policies/steps that governments are taking to address the challenges of the climate crisis. This review highlights the essential adaptations and mechanisms required for legume crops to thrive and fulfill their significant roles in global nutrition. It explores how these crops can be improved to better withstand the environmental stresses, enhance their nutritional profiles, and increase their yields. Additionally, the review discusses the importance of legumes in sustainable agriculture and food security, emphasizing their potential to address the future challenges in feeding the growing global population. By focusing on these critical aspects, the review aims to underscore the importance of legumes in ensuring a healthy and sustainable food supply.
1. Introduction
The world population is predicted to double by 2050, and the increasing population presents concerns regarding food security [1]. The importance of legume crops is widely acknowledged, and increasing the nitrogen fixation, physical conditions, and biodiversity is more necessary than ever, as suggested in [2,3]. In addition, legume crops are important food and feed crops, as well as staple crops, being key components of the diets in different regions and helping to fulfill the nutritional requirements of human beings [4]. Legumes are the second-largest family on Earth and rank second in importance regarding the current and past agricultural activities for humans. Legumes represent a natural precious gift for humankind and are also called the poor man’s meat as they are rich in protein; apart from protein, grain legumes are rich in nutritional components including carbohydrates, vitamins etc. [5]. Moreover, legumes play a role in conserving/fixing the nitrogen in the soil. Grain legumes serve the ecosystem by fixing nitrogen, and they also have deep root systems that help to reduce soil erosion. Legumes’ popularity is rising due to the increasing demand for human health [6,7].
Economically the most important vegetables, the legumes consumed as vegetables are green pods, green pea pods, cowpeas, common peas, and faba beans [8]. In many countries, legumes are consumed after cooking, but they are also consumed as sprouts, such as with mung beans. In addition, legumes have different organoleptic properties that are considered to be important sources of carbohydrates, essential minerals, vitamins, and some other antioxidants and health-promoting compounds [9,10]. On the other hand, legumes are considered to be a low-fat food stuff. Among all legumes, the vegetables are consumed the most.
Furthermore, legumes, when consumed as vegetables, contain more water compared with pulses. Thereby, the soluble materials like carbohydrates are richer, and the starch content is much lower in vegetable legumes [11,12]. Additionally, vegetable legumes are more rich in healthy compounds such as vitamins, minerals, chlorophyll, phenolic acid, and are also a rich source of vitamin C [13]. On the other hand, vegetable legumes are preserved for future use when required; therefore, they are being continuously grown due to the increasing awareness of their well-balanced material [14]. There are several minor vegetable legume crops that are known for their value as they contribute to human health [15]. In addition to their nutritional superiority, legumes have also been ascribed economic, cultural, physiological, and medicinal roles.
Thus, although legumes have more beneficial nutrients, despite the many advantages of legumes, the popularity and cultivation of legumes remain lower compared to cereals [16]. The study reveals that, accompanied by a traditional system with industrialization, a largely cereal-based system is heavily reliant upon fossil fuel. The fossil fuel system significantly increases CO2, which has a harmful impact on Earth and causes direct negative effects on land [17].
Legume crops are more prone to biotic and abiotic factors, which have been known as the main constraints that restrain their crop yield [18]. Legume crops are grown across India and the world, but the yield and yield-related traits are affected by the climatic conditions/factors, such as drought and heat, indicating the threat of water scarcity for legumes in the future [19]. Water scarcity causes a severe loss of yield during the vegetative growth period and especially in the reproductive phase. Severe droughts limit the grain yield and related components of legumes. Back-to-back heatwaves or episodes of extreme weather deny plants any recovery windows to compensate for the reproductive losses. Sequential heat stresses slash the grain yields far more than single events in legumes [20], lentils [21], and other crops. Moreover, shifts toward recurrent and concurrent heat and drought will multiplicatively pressure legume production. For example, the chickpea seed yield under five days of 40/30 °C stress coupled with a water deficit declined 81–91% versus 55–64% under heat or drought alone [22]. The yield reduction depends on the duration of the climatic factor: as the duration increases, the yield reduction yield increases and vice versa.
The sustainable development and integration of several approaches for developing climate-resilient varieties are the primary elements that contribute to enhanced legume productivity in such difficult environments [23]. Considering the above fact, developing climate-resilient varieties may lead to enhanced crop yields. Scientists and the government have implemented several approaches and policies to mitigate the effects of climate change [24]. Below figure shows the Benefits of Legume for human and soil (Figure 1).
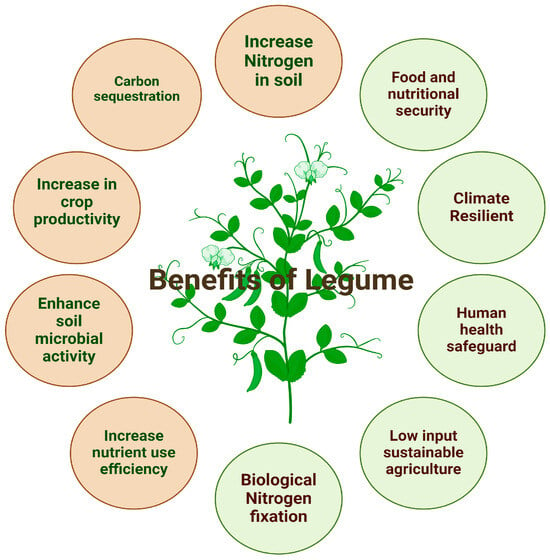
Figure 1.
Schematic representation of the benefits of legumes for humans and soil.
2. As Global Temperatures Rise, Grain Legumes Face Serious Threats
Climate change is affecting the food system globally as well as the human food security system and the health of the human population [25]. Globally, the average temperatures have already risen 1 °C above the pre-industrial averages and are firmly on track to increase by 1.5 °C between 2030 and 2052 at the current trajectories. Under optimistic projections limiting the warming to 1.5 °C, the global legume crop yields are expected to decrease by about 10%. However, under business-as-usual scenarios, the temperatures may rise 2.5–4.8 °C by 2100 [26], carrying median yield losses between 20 and 49% for major legumes like beans, chickpeas, cowpeas, lentils, and soybeans [27].
Nearly all legume species demonstrate marked declines in the key physiological processes beyond 30–35 °C [28], with extreme heat exposure over 40 °C capable of inducing complete reproductive failure [29]. Intensive agricultural and industrial development change the agricultural production and productivity of legume crops. Due to severe climate change, the productivity of legumes is decreasing day by day as many harms to legumes have occurred as a result of climate change. Thus, climate change presents a serious and severe threat to legumes. In this regard, we will also discuss the impact of biomass [30].
The adverse effects of climate change on legumes call for everyone to address this issue as legume crops can provide better yields [31,32]. Nowadays, the greenhouse gas (GHG) effect is increasing on crops, especially among legume crops, which makes it difficult to achieve the yield targets. According to the literature, it is evident that the temperature and CO2 level exert a direct and indirect effect on the physiological and biochemical characteristics of grain legumes [33]. The below table shows some different legumes and their threshold temperatures (Table 1).

Table 1.
The heat stress threshold temperature ranges of some leguminous crops.
3. Global Legume Security Goals under Threat
With the global population projected to increase to 9.7 billion by 2050, the demand for plant-based proteins is projected to rise by 51% between 2000 and 2050 [49]. Food systems are essential to achieving the goals of sustainable development, particularly zero hunger and zero poverty by 2030 [50].
Climate change is one of the major challenges that affects legume crops directly and indirectly [51]. Many tragic yet logical factors are initiated by cultivation to facilitate better crop production and tackle climate change [52,53]. Leguminous vegetables including lentils, dry beans, chickpeas, cowpeas, and soybeans already provide 33% of the total dietary protein nitrogen for humans worldwide [54,55], underscoring their indispensability to global food security. However, the legume yields in the top producer regions like sub-Saharan Africa, South Asia, and Australia are highly likely to fall below the baseline levels necessary to meet the future nutritional demands under all but the most ambitious climate change scenarios [56].
Tens of thousands of species are threatened with extinction as a result of human activities [57]. Here, we explore how the extinction risks of terrestrial mammals and birds might change in the next 50 years. The future population growth and economic development are forecasted to impose unprecedented levels of extinction risk on many more species worldwide, especially the large mammals of tropical Africa, Asia, and South America [58]. Yet, these threats are not inevitable. Proactive international efforts to increase crop yields, minimize land clearing and habitat fragmentation, and protect natural lands could increase the food security in developing nations and preserve much of Earth’s remaining biodiversity [59].
4. Realizing Climate Resilience in Leguminous Vegetables Is an Urgent Necessity
Food security has long been linked with nourishment, but now this picture has changed as the concept of nutritional security with nutritional demand has become prevalent [60]. In addition to producing legumes for livestock, vegetable legumes help to ensure the long-term viability of crop production [61]. Vegetable production provides food security as well as boosting the rural and urban economies.
The fact that vegetables need little land and can yield a high profit in a short time makes vegetable farming appealing. This is especially true for vegetable legumes like peas, soybeans (also known as edamame), fava beans, broad beans, and cowpeas [62]. Leguminous vegetables have long been sources of micronutrients for humans [63,64]. Their consumption prevents diseases such as cardiovascular and metabolic diseases [8]. Vegetable legumes possess various beneficial properties as they include minerals and vitamins, and they play a role as co-factors of metabolic reactions and coenzymes [65]. In addition, legume vegetables have a short duration for completing development and growth; they can be grown more than once a year and involve low costs during their harvest. Therefore, more attention needs to be paid to the production of vegetable legumes [66]. However, climate change affects vegetable production [67]. Without urgent efforts to bolster the legume climate resilience, the progress on malnutrition reduction, protein transition, smallholder farmer livelihoods, and sustainable agriculture worldwide hangs in the balance [68]. The reviewed literature highlights the multifaceted risks posed to legume producers globally while emphasizing the priority adaptations and mechanisms for managing the climate uncertainty to enable legumes to sustainably fulfill the current and future food security roles.
5. Rising Carbon Dioxide: Limited Benefits Unable to Offset Other Factors
The atmospheric CO2 concentration has risen since the beginning of the Industrial Revolution [69]. CO2 affects plants and decreases their yield due to the increase in global temperatures, thus directly harming crop plants [70,71]. Understanding how plants respond and might be adapted to a future increase in [CO2] will also help us to understand how they are currently responding and how they may have adapted to the increase that has already occurred [72]. The effects of increases in [CO2] on the physiology and development of plants have been the subject of much research over the past 20 years and have been the subject of many detailed reviews, as discussed below [73,74].
5.1. Elevated Atmospheric CO2 Accelerates Legume Photosynthesis and Also Increases Yields
As the atmospheric CO2 concentrations have risen above 400 ppm—levels unprecedented in over 800,000 years—the so-called “CO2 fertilization effect” directly stimulates photosynthesis and water retention in C3 plants like legumes [75]. On average, across diverse environments and genetic material, elevated CO2 enhances the legume photosynthetic rates by 55% and water use efficiency by 48% [76]. However, multiple long-term free-air CO2 enrichment (FACE) studies show that the actual resulting yields increase by just 18% under elevated (550 ppm) versus ambient CO2 levels [75]. The accelerated photosynthesis from CO2 fertilization often fails to translate to higher yields due to other limiting factors affecting the carbon allocation and reproductive development [77].
Here, we show that, despite the many potential metabolism aspects that are affected by [CO2], there is clear evidence only for effects on the Rubisco and stomatal movement in the range of [CO2] that are relevant to global change (i.e., 270–1000 µmol mol−1) [78]. Plants can only perceive a change in the atmospheric concentration through tissues that are exposed to the open air [79]. With the exception of some reproductive organs, only the photosynthetic organs of the plant have direct contact with the atmosphere [80]. The protective cuticle of higher-plant leaves and other photosynthetic organs mean that only the inner surfaces of the guard cells of the stomata and the mesophyll can directly sense a change in atmospheric [CO2] [81]. Although many steps in metabolism utilize or respond to CO2, the only sites where there is convincing evidence for a response in the concentration range of relevance (240–1000 µmol mol−1) are Rubisco and a yet-undefined metabolic step affecting the stomatal aperture [82]. The direct increase in photosynthesis due to the elevation of [CO2] results from two properties of Rubisco terrestrial C3 plants.
(a) The Km of the enzyme for CO2 is close to the current atmospheric concentration, so elevated [CO2] increases the velocity of carboxylation. The table shows the effect of long-term elevated CO2 on legumes.
(b) CO2 competitively inhibits the oxygenation reaction, which produces glycolate, leading to photorespiration [80]. This latter effect is particularly important because it increases the efficiency of the net carbon CO2 uptake by decreasing the photorespiratory CO2 loss and diverting ATP and NADPH (generated by light reactions) away from photorespiratory metabolism to photosynthetic assimilation [83]. Thus, because the efficiency of the net photosynthesis increases, the rate increases regardless of whether other factors limit the gross photosynthetic rate. Although the short-term response of C3 photosynthesis to increased [CO2] may be closely predicted from Rubisco’s properties, a mystery still surrounds the mechanism by which the stomatal aperture responds to variations in [CO2] [84]. Below table shows the effect of long term e[CO2] on legumes (Table 2).
Table 2.
Examples of effects of long-term e[CO2] on biomass production and seed yield in a selection of grain and groundnut legumes [76].
5.2. The CO2 Fertilization Effect Plateaus around 550 ppm and Will Be Increasingly Constrained by Other Climate Factors
The stimulatory effect of rising CO2 on legume crop yields plateaus around 550 ppm across the bean, soybean, and peanut varieties studied to date [85]. Yet, under all but the most ambitious emission reduction scenarios, the atmospheric CO2 is projected to rise beyond 700 ppm by 2050 and exceed 900 ppm by 2100 (IPCC, 2022). The temperatures over the same period are highly likely to surpass the optimal legume growing thresholds, constraining any transient CO2 benefits [56]. The threshold of 550 ppm in CO2 concentration refers to the level at which significant responses in plant yield are observed. This threshold has been identified through various scientific studies and experiments that explore the effects of elevated CO2 levels on plant growth, photosynthesis, and overall productivity. The threshold can be identified by a controlled environment study and FACE (Free-Air CO2 enrichment) experiment. The increased heat-induced flower abortion and restricted reproductive duration will overshadow any enhancements in the operational photosynthetic capacity [23]. Moreover, while the CO2 fertilization effect may slightly buoy the legume yields over the coming decade, it cannot protect the global legume production from the many other existential threats posed by rising emissions, like extreme temperature, water availability, and soil and nutrient availability.
In tropical regions like West Africa’s Sahel belt, the climate models project intensified intra-seasonal precipitation variability under climate change [86,87]. Increased rainfall uncertainty poses serious challenges for legume farmers in terms of planting timing, fertilizer application, and harvest scheduling. Excess rainfall also elevates the risk of fungal disease outbreaks, as demonstrated for foliar Bean Common Mosaic Virus, where the infection rates doubled under simulated future high precipitation regimes [88]. Below table shows the effects of different eCO2 concentration on legume yield (Table 3).
Table 3.
Effects of different eCO2 concentration on legume yield.
5.3. Drought Frequency in Key Legume Regions Threatens Yield Stability
The substantial development and integration of advanced approaches for dry environments are the primary elements that contribute to enhanced legume productivity in harsh environments [97,98]. Approaches such as the development of various traits for drought tolerance, innovative breeding, and water efficient practices, for instance, the use of drip irrigation and mulching, are promising ways to mitigate the devastating effects of drought [99]. The adverse effects of drought on several other crops have been previously reviewed, but no updated and comprehensive study is available on the impacts of DS in legume crops [98]. Studies regarding the effects, mechanisms, and management strategies may lead to managing the devastating effects of DS (drought stress) and to developing drought-tolerant genotypes in dry environments [100]. By the middle of this century, the climate projections suggest that the total annual precipitation will decline across India, the Mediterranean basin, southwestern Australia, the central United States, and other major legume-producing areas—even as the extreme rainfall events intensify [101]. Prolonged drought slashes legume yields, with the chickpea seed yields decreasing by 65% during dry years in Australia [102]. Just a 1–2-week moisture deficit during the critical legume reproductive stages can decimate the yields of short-season cowpeas and beans [103].
Legumes are very sensitive to drought; the final yield is significantly decreased due to severe drought. Due to drought, the germination percentage and decline in photosynthetic activity result in a decrease in yield [104,105]. During flowering time, decreased assimilation translocation and carbon fixation repress the flowering time and reproductive stage, thus causing poor seed set and a decline in the sink activity, which results in a decreased yield. Drought affects many plant aspects, including poor germination, shoot and root development, photosynthesis activity, reproductive activity, and the grain-filling stages [106,107]. Due to global climate change, drought is one of the uncontrolled and unpredictable phenomena that directly affects legume crops; it has been observed by studies that drought disturbs the plant morphology, physiology, and growing period. There are some legume crop stages listed in Table 4 that are affected by drought [108,109]. Below table shows that some legume crops effect by drought (Table 4).
Table 4.
Some legume crop stages that are affected by drought.
5.4. Extreme Heat Exposure Critically Threatens Key Legumes in Development Stages and Flowering Phases
The rising temperatures coupled with declining moisture availability create a high risk for compounding legume crop losses. The peanut plant mortality triples following simultaneous exposure to drought and 40 °C heat relative to 30 °C without a water deficit [127]. The flower abortion in heat-stressed soybean plants also soars from 27% to 74% alongside soil drying [128]. With the climate shifts decreasing the rainfall in areas like the Mediterranean and US high plains, aridity will exacerbate the lethal heat effects during the susceptible legume growth phases [129,130]. Breeding legume cultivars with enhanced drought or heat tolerance shows strong potential to counteract the shifting precipitation regimes [131,132]. However, the adaptation pace must accelerate rapidly as those areas presently growing up to 70% of the global legumes face precipitation declines [101]. While drought-protective plant traits are inherently complex [133], emerging genomic tools can enable better assessment of the available genetic diversity for climate resilience within legume germplasm collections [134]. Investing in irrigation infrastructure is also critical as supplemental water during episodic moisture deficits may reduce the legume flower abortion by 29–39% even under moderated heat stress (United States Department of Agriculture-USDA, 2012). Empowering farmers to access improved irrigation will prove essential to stabilizing the legume yields and farm incomes amidst the intensifying precipitation variability [135,136].
In addition, legumes possess various morphological attributes, such as deep root systems, higher water use efficiency, nitrogen fixing ability, the ability to host beneficial microbes in the rhizosphere to solubilize phosphate, etc., as well as low water and nutrient requirements [137,138]. In addition to having these properties, legumes often experience heat resistance during the reproductive stage. The majority of pulses are sensitive to high temperatures above 35 °C, leading to the inhibition of photosynthesis and reduction in pollen fertility [139,140]. Improving the water relation characteristics could be one of the best strategies to address heat tolerance. The deep roots, osmotic adjustment, lower 13C/12C carbon isotope discrimination ratio, and lower stomatal aperture contribute significantly toward improving the water use efficiency, although cellular-level tolerance aspects, including the expression of antioxidative enzymes, heat shock proteins, acquired thermotolerance, membrane stability, and altered carbohydrate metabolizing enzymes, play important roles regarding tolerance [141,142].
The male reproductive development in higher plants is very sensitive to heat stress at all the growth stages. In particular, high-temperature stress results in a lower seed set due to male sterility in most legume crops, including chickpea, common bean, and field pea [34]. In most legumes, the male gametophyte is more sensitive to high temperatures than the female gametophyte [88].
Regarding the female gametophyte, temperatures > 30 °C reduce the stigmatic receptivity and stigmatic pollen germination, stigma and style growth, and ovule penetration [43]. Heat stress abruptly affects almost all the aspects of female gametophyte development, e.g., reduced stigma receptivity in chickpea at 40/30 and 45/35 °C, and reduced ovule number and viability in common bean at 30 °C [88]. Figure 2 shows the effect of heat stress during reproductive stage.
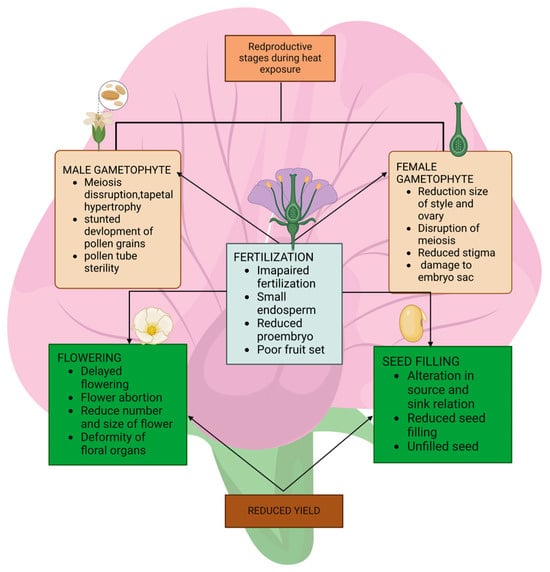
Figure 2.
Effects of heat stress during the reproductive phase.
Heat stress during the flowering stage is a serious threat to the quality and productivity of the legumes. The temperature for legumes during flowering varies from crop to crop, basically according to the season; the winter temperature should be 12–18 °C, and the summer temperature should be 21–28 °C [143,144]. The magnitude of the damage caused by high temperatures depends on the different developmental stages. Grain legumes exhibit maximum heat stress sensitivity during the flowering, pod set, and seed filling growth phases [29]. Brief periods of extreme temperatures of 35 °C or above increase the flower or pod abortion by 22–45% across legume species [103], with chickpeas demonstrating near 100% loss of their reproductive capacity beyond 36 °C even with ample moisture [22]. Just one day of extreme heat exposure during the susceptible flowering stages can slash the legume pod and seed numbers for the entire season. Heat stress affects the grain development because it disintegrates the tapetum layer, which reduces the nutrients’ supply to the microspore, leading to premature anther dehiscence, which controls the synthesis and distribution of carbohydrates to grain, all of which ultimately contribute to a reduction in yield [6,145].
However, seed-filling is also highly vulnerable to the transient heat waves that reduce the grain-fill duration, which represents a primary pathway for impaired legume yields under transient heat waves [146]. Episodes above 34 °C during the seed development stage decreased the harvested dry bean seed weights up to 61% relative to the ambient conditions [147]. Soybeans also exhibited a 5% decline in their seed oil and protein contents over the entire growing season, featuring just 4–5 days of heatwave conditions (40/30 °C day/night temperatures) centered around the critical seed-filling phase [128]. Table 5 shows the effects of heat stress on food legume at different growth stages.
Table 5.
Effects of heat stress on food legumes at different stages of growth.
5.5. Increasing Pest and Pathogen Pressure Creates New Uncertainty
Insect pests are cold-blooded, having body temperatures that are the same as that of the environment; therefore, temperature is a key climatic factor that influences the growth and development of insect pests [154]. Due to climate change, plant insect pests may change their physiology and behavior because of the modification of the host plant resistance variety and interaction with other insects and natural enemies [155]. Increasing temperatures accelerate the reproductive cycles of poikilothermic crop pests, allowing additional generations per season and amplifying their population sizes [156]. Vegetable legumes are a rich source of protein and are damaged by insects, pests, and pathogens under both the field and storage conditions [157]. There are many insect pests, but, among all pests, pod bore (Maruca vitrata) is one of the most dangerous insects [158]. But, nowadays, a vast number of plant insect pests and pathogens harm legume crops. Their effects range from mild to severe symptoms that cause yield losses directly in legumes.
The warmer conditions also expand the viable spatial ranges for legume pests, enabling influxes into historically unaffected areas. For example, the future warming is projected to increase the global regions suited for bean bruchid beetle (Acanthoscelides obtectus) reproduction by 5–20% by 2100, posing novel threats to bean cultivation zones like Eastern Africa [159].
Multiple aggressive legume pathogens including Sclerotinia stem rot, charcoal rot, and Cercospora leaf spot are exhibiting accelerated growth and heightened virulence under elevated temperatures and precipitation variability [160,161]. The field observations underscore the climatic influence on the emerging disease pressure, with the warming-linked range expansion of peanut rust (Puccinia arachidis.) already documented across Africa and Australia over the past decade [162].
On the other hand, grain legumes like faba beans, cowpeas, and soybean-producing hybrid seeds rely on insect cross-pollination for maximal reproduction rates. However, the advances in legume flowering times under climate change scenarios exceed the predicted shifts in the bee pollinator emergence patterns within the same region [163]. The resulting phenological mismatch could impair the legume yield trajectories. For example, experiments indicate that the climate warming-induced temporal divergence between faba plants and Bombus terrestrosin pollinators decreased the reproductive success by over 60% due to insufficient cross-pollination during the target flowering period [163].
The proliferation of the existing legume pests and pathogens alongside the possible emergence of novel threats under changing climate represent a formidable challenge for food security. Integrating the climatic risk factors into pest management frameworks will be essential going forward [159]. Improved landscape diversification via cropping system rotations and targeted agroforestry interventions may additionally impart some buffering of the pest pressures amplified under the climate shifts [156]. However, breeding durable multi-trait insect and pathogen resistance should be a top priority. Ultimately, however, the complexity of the climate change impacts on legume agroecosystems leave the conventional approaches insufficient to offset the risks. Instead, more adaptive, flexible solutions supported by global coordination will be necessary to detect and respond to unanticipated ecological surprises [164].
5.6. Response of Insect Pests to Changeable Precipitation Patterns
Precipitation variations in terms of quantity, severity, and frequency are crucial markers of climate change. The majority of incidents have shown that, while the precipitation intensity has increased, the precipitation frequency has decreased. The frequency of droughts and floods has been favored by this kind of rainfall pattern. Rainfall patterns that overlap have an immediate impact on insect species that hibernate in the soil. To put it briefly, prolonged water stagnation and floods can result from severe rains. Insect survival is threatened by this occurrence, which also impacts their diapause. Furthermore, intense rain and flooding have the potential to wash away insect eggs and larvae [165]. When it rains a great deal, small-bodied pests like jassids, aphids, mites, and whiteflies, among others, might be washed away [166]. Variable rainfall can have a major impact on insect populations. For example, in grassland plots, the authors of [167] investigated the effects of drought and increased summer rainfall on the soil-dwelling wireworm (Agriotes lineatus L.). Particularly when cultivated in grassland plots, wireworms are a very destructive pest of crops like potatoes, corn, sugar beet, etc., and the forecasts indicate that their impact will only increase due to climate change [168]. In contrast to ambient and drought circumstances, Staley et al. [167] observed that the wireworm populations grew quickly in the top soil as a result of higher summer rainfall episodes [169]. Drought affects herbivorous insects in a few ways: (I) dry regions may offer favorable climatic conditions for the development and expansion of herbivorous insects; (II) some insect species may be drawn to plants that are stressed by drought: for example, when plants decrease in humidity through transpiration, water columns in the xylem damage or cavitate, generating an ultrasonic acoustic emission that is picked up by dangerous bark beetles (Scolytidae); and (III) plants under stress from drought produce fewer secondary metabolites, which serve as defense mechanisms, making them more vulnerable to insect attacks [170] (Figure 3).
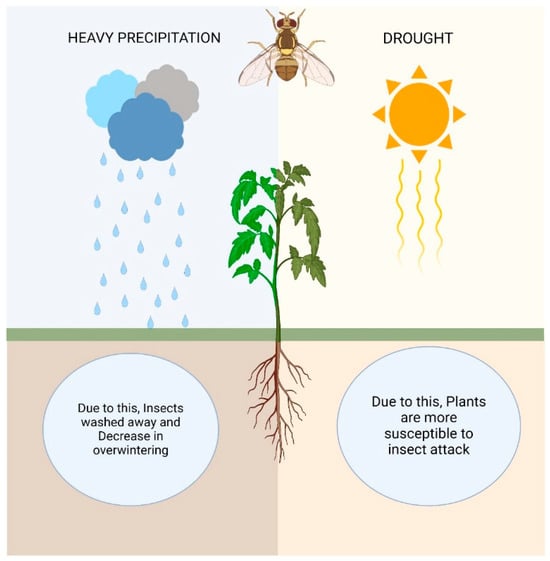
Figure 3.
Impacts of heavy precipitation and drought on agricultural insect pests.
5.7. Nitrogen Fixation Disruption and Soil Fertility Decline
The vital symbiotic relationship enabling atmospheric nitrogen fixation by Rhizobia bacteria into plant-usable forms exhibits steep declines beyond 30–35 °C [23]. Drought stress impairs nitrogen fixation through nodule damage and carbohydrate partitioning changes well before the manifestation of visible yield losses [171]. The nitrogenase enzyme kinetics slows below the optimal temperatures while amino acid deformities accumulate in the nodule tissue under heat stress, markedly suppressing the overall nitrogen fixation capacity [172].
As legume producers face increasing precipitation uncertainty, soil drying is likely to decrease the symbiotic nitrogen inputs below the critical thresholds necessary to support the sustainable intensification goals without external fertilizer amendments [173]. The loss of functional nodules and fixed nitrogen during the early crop establishment under recurrent heat waves reverberates through the entire cropping cycles through compounded soil nutrient deficiency [174]. The insufficient soil nitrogen triggered by climate extremes may erase any transient boosts in the operational photosynthetic productivity from CO2 fertilization [175].
On the other hand, the intersection of rising temperatures, shifting rainfall, and recurrent extremes stand to unravel the legume crop productivity through exacerbated soil nutrient deficits [176]. The combined drought and heat events (40/30 °C day/night temperatures) imposed during the seed filling phase decreased the nitrogen fixation rates in faba bean, chickpea, and lentil plots by 29–76% relative to the ambient conditions [104]. Prolonged elevated temperatures additionally hasten native soil organic matter decomposition and release substantial CO2 stores back into the atmosphere, thus further impeding farm system resilience [177].
6. Broader Socioeconomic Challenges Compound Climate Risks
6.1. Legume Producers Face Widespread Poverty and Constrained Adaptive Capacity
Grain legumes act as crucial poverty alleviation crops across Africa, Asia, and Latin America, providing income generation and nutritious food access for smallholder farmers with limited resources [55]. However, poverty and social inequality fundamentally constrain the adaptive capacity within rural legume production landscapes [178]. Poor farmers often lack the financial means to access improved seeds, irrigation infrastructure, or crop insurance to buffer the climate impacts [179]. Following extreme weather disasters, under-resourced farmer communities frequently struggle to recover without strong social safety nets or private savings [180]. Financing shortfalls thereby compound the climate risks faced by the globally indispensable legume basket regions already facing agricultural marginalization.
6.2. Access to Actionable Climate Information and Early Warning Systems Uneven
While the seasonal forecasting models provide valuable tools for climate risk projection, straightforward actionable information rarely reaches remote smallholder farmers [181]. Language barriers, low rates of smartphone penetration, and a lack of rural agricultural extension services hamper the dissemination efforts. Lacking reliable advance information on the impending extreme events or shifting agro-advisories, vulnerable legume producers cannot proactively implement protective measures [180]. Reinforcing participatory climate communication channels through the existing social structures is critical so that farming communities worldwide can brace for the acute shocks and stressors to come.
6.3. Policy Incentives Remain Aligned to Cereal Commodity Crops despite Nutritional Needs
Although the rising protein demands underscore the urgent need to bolster legume production, the agricultural policy frameworks concentrate primarily on stabilizing the yields, oversupply, and subsidization for cereal staples like wheat, rice, and maize [46]. Lacking the political capital of major commodity crops, grain legumes attract little targeted breeding investment relative to their global importance for food security [182]. This imbalance constrains the legume genetic yield potential improvements necessary to offset the climate change impacts, raising equity concerns given the high nutrition and livelihood benefits conferred by pulses for poor consumers and small-scale producers worldwide [46].
6.4. Avoiding the Protein Transition Crisis Requires Explicit Prioritization of Legume Crops
Given the perfect storm of the climate threats, booming protein demand, and policy distortion toward cereals instead of legumes, the world appears to be headed toward a protein transition crisis without deliberate interventions [183]. Achieving food system resilience hinges on spurring large-scale transitions toward diverse nutrition production typified by grain legumes [184]. However, landscape-level shifts cannot materialize under status quo policy inertia. Avoiding the looming protein shortfall this century necessitates the explicit prioritization of legumes within agricultural development, food security, and climate adaptation agendas [185]. Overcoming the disparities in public and private research funding, strengthening the extension services promoting sustainable intensification based on legume integration, and subsidizing the climate-risk reduction mechanisms for small-scale producers represent potential actions linking human livelihoods, nutrition, and ecological health through next-generation resilient legume agroecosystems [183].
7. Opportunities for Multi-Functional Legume-Supported Adaptation Pathways
7.1. Integrating Legumes Enhances Climate Resilience across Cropping Systems
Strategically incorporating appropriate grain legume crops into the prevailing regional cropping cycles shows major promise for bolstering climate resilience. As natural nitrogen-fixers, legumes reduce the reliance on synthetic fertilizers and minimize the nitrous oxide climate emissions relative to the production gains [186]. Legume root residues additionally contribute soil carbon relative to other crops, with the potential to offset the fossil fuel emissions. Mixed cereal–legume intercropping techniques leveraging underutilized vertical canopy space likewise enhance the biomass yields, water use efficiency, and economic stability under variable climate conditions in Africa and India [186]. Crop diversification through legume integration thereby supports multifaceted adaptation, mitigation, and productivity co-benefits that are relevant across scales.
7.2. Transitioning Local Diets through Legume Consumption: Part of the Puzzle
In many climate-vulnerable regions that are heavily reliant on cereals, traditional legume dishes remain underutilized nutritious alternatives [187]. Mainstreaming legume foods like cowpea, pea, chickpea, or pigeon pea through cultural promotion and value chain development programs can expand the domestic market access to locally adapted crops, incentivizing farmers to maintain production even during periods of climatic hardship. Given the vast nutritional benefits of pulses and the ability to substitute substantial proportions of meat demand via plant-based protein [56], supporting legume-based dietary transitions through community nutrition education and food procurement policies also constitutes a healthful, low-emission tactic for enhancing the future food system resilience [188].
7.3. Biodiverse Agroforestry Schemes Link Legume Services across Broader Landscapes
Enhancing the agricultural landscape complexity through carefully designed agroforestry systems interspersing multi-functional leguminous cover crops, hedgerows, and fallow vegetation offers another climate adaptation strategy centered on legumes [67]. Alongside stabilizing the soils and fostering pest regulation ecosystem services, the deep-rooted tendencies of perennial grain legumes like carob and mesquite offer niche drought resilience [189]. While still experimental, innovating toward integrated leguminous agroforestry production models may hold unique promise for sustainably adapting marginal lands to intensifying climate uncertainty [190].
7.4. Ultimately Realizing That the Legume Future Hinges on Collective Global Commitment
Fulfilling the fundamental ecosystem services and nutritional roles of leguminous vegetables under accelerating environmental change ultimately requires rapid, ambitious actions across production zones, market systems, and consumer populations worldwide. No single technological innovation or policy measure alone can enable the grain legumes to deliver on the current and projected demands that are essential for global food security and ecological sustainability [191]. Instead, the coming decades necessitate unprecedented global coordination across agricultural research institutions, policymakers, development programs, and farmer support mechanisms explicitly oriented toward realizing robust, equitable, and nutritious legume-based future food systems for a climate-disrupted planet [192]. There are no comparable alternatives to grain legumes in terms of blending the climate-resilient productivity, nutritional quality, and environmental stewardship at scale [55]. Whether the world rises to sufficiently harness the unique multifunctional potential of legumes through unprecedented collaboration and investment remains the most pivotal open question determining the fate of sustainable human development worldwide [193].
8. Future Outlook: Emerging Innovations to Accelerate Legume Climate Resilience
8.1. Selecting Superior Rhizobia Strains and Integrating Fertilizers Vital for Adaptation
Maintaining adequate nitrogen availability amid climate change will require multi-pronged approaches. Developing legume cultivars that are capable of sustaining effective nodulation under intermittent moisture stress and heat waves is essential and may be enabled through the rapid genomic screening of diverse germplasms [134]. However, the host plant genetics cannot be relied upon alone. The inoculation with elite Rhizobium strains exhibiting superior heat tolerance represents an accessible tactic for producers in the immediate future. Additionally, integrating judicious inorganic fertilizer applications to stabilize the yield goals may also be necessary where climate disruptions overwhelm the biological nitrogen contributions [185], albeit with care taken to avoid inhibitory effects on the symbiotic fixation.
8.2. Advanced Genomic Approaches Enable Faster Development of Climate-Resilient Legumes
Leveraging whole-genome sequencing, precision phenotyping platforms, and machine learning algorithms offers tremendous yet under-tapped potential to accelerate the delivery of climate-adapted legume varieties to producers’ fields [134]. High-throughput genomic selection now enables the simultaneous screening of complex traits like heat tolerance across expansive germplasm collections, which is not feasible through the traditional field-based approaches [190]. These novel techniques, coupled with the expanding public accessibility to international crop genebanks via digital interfaces, democratize the access to invaluable breeding material [56]. However, realizing resilience gains requires additional supportive financing to integrate the identified hardy lineages into pre-breeding pipelines and commercial variety advancement systems targeting climate-vulnerable geographies in the developing world [143].
8.3. Gene Editing Holds Unique Promise for Climate-Protective Legume Innovations
Gene editing techniques like CRISPR-Cas expand the possibility to develop transgenic, climate-resilient legume varieties at accelerated timelines compared to conventional breeding [194,195]. However, the novel gene editing applications in legumes currently remain confined to proof-of-concept research nearly exclusively targeting model organisms like soybean and Medicago truncatula. Directing further tool development and regulatory enabling environments to support the editing applications in orphan, nutrition-critical legumes like cowpeas, faba beans, and lentils is essential to democratize the climate-resilience benefits for vulnerable producers and consumers worldwide [192]. Particularly for complex, quantitative climate tolerance traits not feasibly manipulated through traditional crossing, strategic gene editing may offer uniquely rapid, precise, and revolutionary adaptation solutions if ethically deployed toward the needs of subsistence farmers [196,197,198].
8.4. Microbiome Biofertilizers and Biopesticides Expand Climate Risk Protection Options
Harnessing beneficial plant–microbe interactions is garnering rising interest in enhancing agricultural resilience [199]. Custom microbiome biostimulant products and biofertilizers containing elite plant growth-promoting rhizobacteria demonstrate the capacity to enhance the legume germination, nutrient uptake, and pathogen resistance under variable climate stresses [99]. These bio-solutions offer accessible adaptation tactics deployable on-farm by legume producers themselves. Microbial seed coatings encapsulating nitrogen-fixing rhizobia also help to retain the inoculant viability under environmental flux [200]. Foliar biopesticides harnessing antifungal bacteria or essential oils provide chemical-free options to manage the yield losses from virulent foliar diseases triggered after extreme weather. Scaling the commercialization and farmer education around the emerging microbiome and biopesticide innovations tailored for grain legume crops is paramount to expanding the climate risk management toolkits available across the legume production landscape [193].
8.5. Accelerating Heat-Tolerant Legume Breeding Is Imperative but Challenging
Developing legume cultivars that are tolerant to the emergent extremes in heat and aridity represents a top climate change adaptation priority. However, the complex polygenic architecture underlying the heat stress response [144] poses difficulties for rapid variety improvement. The further integration of quantitative trait loci mapping, genome-wide association studies, and genomic selection approaches can help to expedite the delivery of hardy legume germplasms to producers’ fields [179]. However, plant breeding alone is unlikely to elicit adaptation fast enough without parallel efforts to curb the emission trajectories. Under the projected mid-century temperatures, even heat-tolerant lines yet to be developed will struggle to deliver the productivity gains desperately needed [201,202,203,204].
9. Processing and Macroeconomic Analysis of Legumes
9.1. Deep Processing of Legumes
The deep processing of legumes involves various techniques to transform raw legumes into valuable products. These processes improve the nutritional value, digestibility, and palatability of legumes, making them more suitable for consumption and use in different food products [205]. Here are some common deep-processing methods for legumes: cleaning and sorting, soaking, dehulling, milling, extrusion, fermentation, sprouting/germination, cooking, canning, drying, enzymatic treatment, and fortification. These processing methods not only improve the quality and usability of legumes but also help in developing a variety of innovative food products that cater to different dietary needs and preferences [206].
9.2. Macroeconomic Analysis of Legumes
The legume industry plays a crucial role in the global economy due to its significant contributions to food security, soil health, and sustainable agricultural practices [204]. Legumes, such as beans, lentils, and peas, are not only rich in protein and essential nutrients but also have the unique ability to fix atmospheric nitrogen, thereby enhancing the soil fertility and reducing the need for synthetic fertilizers. This natural nitrogen fixation is particularly beneficial for organic farming, which relies on eco-friendly agricultural practices. Additionally, legumes are instrumental in greening pastures and combating desertification as their deep root systems improve the soil structure, prevent erosion, and enhance water retention [207]. The rising demand for organic food has further spurred interest in legume cultivation, promoting a shift toward more sustainable farming practices. Consequently, the macroeconomic analysis of the legume industry indicates a promising outlook, driven by its environmental benefits, increasing consumer awareness of healthy eating, and the global push toward sustainable agriculture [208].
10. Conclusions
Climate change poses serious threats to achieving sustainable legume-supported food systems worldwide. However, the coming decades of transformation also present the possibility to unlock legumes’ unique environmental services toward productive, nutrition-secure, and ecologically multifunctional agriculture on a climate-disrupted planet. The emerging innovations highlighted in this review—spanning omics-empowered crop improvement platforms, gene editing techniques, and plant microbiome harnessing combined with global coordination around the explicit policy prioritization and financing of legume crops—can help to accelerate the realization of a resilient future in which our health and climate needs converge. There exist no better-positioned alternatives than grain legumes to anchor the necessary protein transitions emerging across societies worldwide. The eternal pragmatism of legume producers themselves now depends on rapid external investments to appropriately position leguminous vegetables at the forefront of any genuine attempts toward future global food security and environmental sustainability. What emerges in terms of the research, development, and policy commitments supporting vulnerable smallholder legume producers over the coming years will ultimately determine the balance between crisis and opportunity for nourishing the world under climate change.
Realizing climate resilience is an urgent multidimensional challenge. This review illuminates the serious multidimensional risks climate change poses for legume productivity and global food security. Accompanying any transient CO2 fertilization benefits, rising temperatures, shifting precipitation extremes, and increased climate variability stand primed to destabilize agroecosystems directly and indirectly across the primary legume basket regions. Realizing climate-smart legume production to nourish the rising population will require acceleration across a suite of adaptive interventions:
- Heat/drought-tolerant legume cultivar development;
- Enhanced climate-informed agronomic management;
- Refined biological nitrogen fixation solutions;
- Strengthened climate pest/disease monitoring systems;
- Supportive agricultural development policies explicitly targeting smallholder legume producer resilience.
Tackling these challenges additionally necessitates the recommitment to ambitious emission reduction targets, recognizing that global temperature trajectories beyond 1.5 °C markedly ratchet the risk exposure for vulnerable farming communities already struggling with seasonal variability. There are no silver-bullet solutions. However, given the rising calls for protein diversification and ecological intensification, the climate resilience of leguminous vegetables now plays an indispensable role in determining the sustainability of our entire global food system. This urgently demands dedicated research, policymaking, and public engagement efforts if legumes are to sustainably fulfill the current and future food security needs.
Author Contributions
Conceptualization, K.D., P.K., A.K. and N.D.; resources, P.S.D., A.Y. and M.J.; data curation, P.K., K.D. and M.J.; writing—original draft preparation, K.D., N.D., M.J. and A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rahut, D.B.; Aryal, J.P.; Manchanda, N.; Sonobe, T. Chapter 6—Expectations for household food security in the coming decades: A global scenario. In Future Foods; Bhat, R., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 107–131. [Google Scholar] [CrossRef]
- Dhaliwal, S.K.; Talukdar, A.; Gautam, A.; Sharma, P.; Sharma, V.; Kaushik, P. Developments and prospects in imperative underexploited vegetable legumes breeding: A review. Int. J. Mol. Sci. 2020, 21, 9615. [Google Scholar] [CrossRef]
- Palmero, F.; Fernandez, J.A.; Garcia, F.O.; Haro, R.J.; Prasad, P.V.V.; Salvagiotti, F.; Ciampitti, I.A. A quantitative review into the contributions of biological nitrogen fixation to agricultural systems by grain legumes. Eur. J. Agron. 2022, 136, 126514. [Google Scholar] [CrossRef]
- Semba, R.D.; Ramsing, R.; Rahman, N.; Kraemer, K.; Bloem, M.W. Legumes as a sustainable source of protein in human diets. Glob. Food Secur. 2021, 28, 100520. [Google Scholar] [CrossRef]
- Yvonne, M.; Victoria, A.J. The Role of Legumes in Human Nutrition. In Functional Food; Maria Chavarri, H., Ed.; IntechOpen: Rijeka, Croatia, 2017; Chapter 6. [Google Scholar] [CrossRef]
- Kumar, A.; Saini, K.S.; Rolaniya, L.K.; Singh, L.K.; Kaushik, P. root system architecture and symbiotic parameters of summer mung bean (Vigna radiata) under different conservation agriculture practices. Sustainability 2022, 14, 3901. [Google Scholar] [CrossRef]
- Bennetau-Pelissero, C. Plant proteins from legumes. Bioact. Mol. Food 2019, 1, 223–265. [Google Scholar]
- Ntatsi, G.; Gutiérrez-Cortines, M.E.; Karapanos, I.; Barros, A.; Weiss, J.; Balliu, A.; dos Santos Rosa, E.A.; Savvas, D. The quality of leguminous vegetables as influenced by preharvest factors. Sci. Hortic. 2018, 232, 191–205. [Google Scholar] [CrossRef]
- Carbas, B.; Machado, N.; Pathania, S.; Brites, C.; Rosa, E.A.; Barros, A.I. Potential of legumes: Nutritional value, bioactive properties, innovative food products, and application of eco-friendly tools for their assessment. Food Rev. Int. 2023, 39, 160–188. [Google Scholar] [CrossRef]
- Aleem, M.; Aleem, S.; Sharif, I.; Aleem, M.; Shahzad, R.; Khan, M.I.; Batool, A.; Sarwar, G.; Farooq, J.; Iqbal, A. Whole-genome identification of APX and CAT gene families in cultivated and wild soybeans and their regulatory function in plant development and stress response. Antioxidants 2022, 11, 1626. [Google Scholar] [CrossRef]
- Guillon, F.; Champ, M.-J. Carbohydrate fractions of legumes: Uses in human nutrition and potential for health. Br. J. Nutr. 2002, 88, 293–306. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Ucak, İ.; Jain, S.; Elsheikh, W.; Ali Redha, A.; Kurt, A.; Toker, O.S. Impact of drying on techno-functional and nutritional properties of food proteins and carbohydrates-A comprehensive review. Dry. Technol. 2024, 42, 592–611. [Google Scholar] [CrossRef]
- Alemu, T. Nutritional contribution of fruit and vegetable for human health: A review. Int. J. Health Policy Plann. 2024, 3, 1–9. [Google Scholar]
- Carlini, B.; Lucini, C.; Velázquez, J. The role of legumes in the sustainable mediterranean diet: Analysis of the consumption of legumes in the mediterranean population over the last ten years a prisma statement methodology. Sustainability 2024, 16, 3081. [Google Scholar] [CrossRef]
- Aghora, T.; Thangam, M.; Patil, N. Legume Vegetables for Human Nutrition and Entrepreneurship. In Vegetables for Nutrition and Entrepreneurship; Springer: Berlin/Heidelberg, Germany, 2023; pp. 439–462. [Google Scholar]
- Odeku, O.A.; Ogunniyi, Q.A.; Ogbole, O.O.; Fettke, J. Forgotten gems: Exploring the untapped benefits of underutilized legumes in agriculture, nutrition, and environmental sustainability. Plants 2024, 13, 1208. [Google Scholar] [CrossRef]
- Nunes, L.J. The rising threat of atmospheric CO2: A review on the causes, impacts, and mitigation strategies. Environments 2023, 10, 66. [Google Scholar] [CrossRef]
- Laskar, R.A.; Dowarah, B.; Sheikh, N. Germplasm diversity and breeding approaches for genetic improvement of mungbean. In Advanced Crop Improvement, Volume 2: Case Studies of Economically Important Crops; Springer: Berlin/Heidelberg, Germany, 2023; pp. 173–196. [Google Scholar]
- Gayacharan; Parida, S.K.; Mondal, N.; Yadav, R.; Vishwakarma, H.; Rana, J.C. Mining legume germplasm for genetic gains: An Indian perspective. Front. Genet. 2023, 14, 996828. [Google Scholar]
- Siebers, M.H.; Yendrek, C.R.; Drag, D.; Locke, A.M.; Rios Acosta, L.; Leakey, A.D.; Ainsworth, E.A.; Bernacchi, C.J.; Ort, D.R. Heat waves imposed during early pod development in soybean (Glycine max) cause significant yield loss despite a rapid recovery from oxidative stress. Glob. Chang. Biol. 2015, 21, 3114–3125. [Google Scholar] [CrossRef]
- Siddique, K.; Loss, S.; Regan, K.; Jettner, R. Adaptation and seed yield of cool season grain legumes in Mediterranean environments of south-western Australia. Aust. J. Agric. Res. 1999, 50, 375–388. [Google Scholar] [CrossRef]
- Thomey, M.L.; Slattery, R.A.; Köhler, I.H.; Bernacchi, C.J.; Ort, D.R. Yield response of field-grown soybean exposed to heat waves under current and elevated [CO2]. Glob. Chang. Biol. 2019, 25, 4352–4368. [Google Scholar] [CrossRef] [PubMed]
- Ngalamu, T.; Galla, J.O.; Ofori, K.; Meseka, S.K. Genetic improvement for development of a climate resilient food legume Crops: Relevance of cowpea breeding approach in improvement of food legume crops for future. In Climate Change and Legumes; CRC Press: Boca Raton, FL, USA, 2023; pp. 97–120. [Google Scholar]
- Burgess, M.G.; Van Boven, L.; Wagner, G.; Wong-Parodi, G.; Baker, K.; Boykoff, M.; Converse, B.A.; Dilling, L.; Gilligan, J.M.; Inbar, Y. Supply, demand and polarization challenges facing US climate policies. Nat. Clim. Chang. 2024, 14, 134–142. [Google Scholar] [CrossRef]
- Vermeulen, S.J.; Campbell, B.M.; Ingram, J.S. Climate change and food systems. Annu. Rev. Environ. Resour. 2012, 37, 195–222. [Google Scholar] [CrossRef]
- Asseng, S.; Martre, P.; Maiorano, A.; Rötter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2019, 25, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Aleem, M.; Aleem, S.; Sharif, I.; Wu, Z.; Aleem, M.; Tahir, A.; Atif, R.M.; Cheema, H.M.N.; Shakeel, A.; Lei, S. Characterization of SOD and GPX gene families in the soybeans in response to drought and salinity stresses. Antioxidants 2022, 11, 460. [Google Scholar] [CrossRef]
- Izaurralde, R.C.; Thomson, A.M.; Morgan, J.; Fay, P.; Polley, H.; Hatfield, J.L. Climate impacts on agriculture: Implications for forage and rangeland production. Agron. J. 2011, 103, 371–381. [Google Scholar] [CrossRef]
- Devasirvatham, V.; Tan, D.; Gaur, P.; Raju, T.; Trethowan, R. High temperature tolerance in chickpea and its implications for plant improvement. Crop Pasture Sci. 2012, 63, 419–428. [Google Scholar] [CrossRef]
- Basu, P.; Singh, U.; Kumar, A.; Praharaj, C.; Shivran, R. Climate change and its mitigation strategies in pulses production. Indian J. Agron. 2016, 61, S71–S82. [Google Scholar]
- Vadez, V.; Berger, J.D.; Warkentin, T.; Asseng, S.; Ratnakumar, P.; Rao, K.P.C.; Gaur, P.M.; Munier-Jolain, N.; Larmure, A.; Voisin, A.-S. Adaptation of grain legumes to climate change: A review. Agron. Sustain. Dev. 2012, 32, 31–44. [Google Scholar] [CrossRef]
- Sena, L.; Mica, E.; Valè, G.; Vaccino, P.; Pecchioni, N. Exploring the potential of endophyte-plant interactions for improving crop sustainable yields in a changing climate. Front. Plant Sci. 2024, 15, 1349401. [Google Scholar] [CrossRef]
- Tariq, A.; Graciano, C.; Sardans, J.; Zeng, F.; Hughes, A.C.; Ahmed, Z.; Ullah, A.; Ali, S.; Gao, Y.; Peñuelas, J. Plant root mechanisms and their effects on carbon and nutrient accumulation in desert ecosystems under changes in land use and climate. New Phytol. 2024, 242, 916–934. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, N.; Gupta, K.; Bhandhari, K.; Kumar, S.; Thakur, P.; Nayyar, H. Proline induces heat tolerance in chickpea (Cicer arietinum L.) plants by protecting vital enzymes of carbon and antioxidative metabolism. Physiol. Mol. Biol. Plants 2011, 17, 203–213. [Google Scholar] [CrossRef]
- Kigel, J.; Konsens, I.; Ofir, M. Branching, flowering and pod-set patterns in snap-bean (Phaseolus vulgaris L.) as affected by temperature. Can. J. Plant Sci. 1991, 71, 1233–1242. [Google Scholar] [CrossRef]
- Prasad, P.V.; Boote, K.J.; Allen, L.H., Jr.; Thomas, J.M. Effects of elevated temperature and carbon dioxide on seed-set and yield of kidney bean (Phaseolus vulgaris L.). Glob. Chang. Biol. 2002, 8, 710–721. [Google Scholar] [CrossRef]
- Laing, D.; Jones, P.; Davis, J.; Goldsworthy, P.; Fisher, N. The Physiology of Tropical Field Crops; Wiley: Hoboken, NJ, USA, 1984. [Google Scholar]
- Craufurd, P.; Summeifield, R.; Ellis, R.; Roberts, E.; Singh, B.; Mohan Raj, D.; Dashiell, K.; Jackai, L. Advances in Cowpea Research; IITA: Ibadan, Nigeria, 1997. [Google Scholar]
- Bishop, J.; Potts, S.G.; Jones, H.E. Susceptibility of faba bean (Vicia faba L.) to heat stress during floral development and anthesis. J. Agron. Crop Sci. 2016, 202, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.V.V.; Craufurd, P.Q.; Kakani, V.G.; Wheeler, T.R.; Boote, K.J. Influence of high temperature during pre-and post-anthesis stages of floral development on fruit-set and pollen germination in peanut. Funct. Plant Biol. 2001, 28, 233–240. [Google Scholar] [CrossRef]
- Barghi, S.S.; Mostafaii, H.; Peighami, F.; Zakaria, R.A. Path analysis of yield and its components in lentil under end season heat condition. J. Agriculturae 2012, 2, 969–974. [Google Scholar]
- Mylona, P.; Pawlowski, K.; Bisseling, T. Symbiotic nitrogen fixation. Plant Cell 1995, 7, 869. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kaur, R.; Kaur, N.; Bhandhari, K.; Kaushal, N.; Gupta, K.; Bains, T.; Nayyar, H. Heat-stress induced inhibition in growth and chlorosis in mungbean (Phaseolus aureus Roxb.) is partly mitigated by ascorbic acid application and is related to reduction in oxidative stress. Acta Physiol. Plant. 2011, 33, 2091–2101. [Google Scholar] [CrossRef]
- Gladish, D.K.; Rost, T.L. The effects of temperature on primary root growth dynamics and lateral root distribution in garden pea (Pisum sativum L., cv. “Alaska”). Environ. Exp. Bot. 1993, 33, 243–258. [Google Scholar] [CrossRef]
- Duke, J. Handbook of Legumes of World Economic Importance; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- FAOSAT. Food and Agriculture Organization of the United Nations. 2018, p. 403. Available online: http://faostat.fao.org (accessed on 27 June 2024).
- Peart, R.M.; Shoup, W.D. Agricultural Systems Modeling and Simulation; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Shirsath and Bhosale Agro India Ltd. Modern Agrotechniques for Cultivation of Black Gram/Urdbean (Vigna mungo L.); Shirsath and Bhosale Agro India Ltd.: Ahmednagar, India, 2017. [Google Scholar]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Fanzo, J.; Haddad, L.; Schneider, K.R.; Béné, C.; Covic, N.M.; Guarin, A.; Herforth, A.W.; Herrero, M.; Sumaila, U.R.; Aburto, N.J. Rigorous monitoring is necessary to guide food system transformation in the countdown to the 2030 global goals. Food Policy 2021, 104, 102163. [Google Scholar] [CrossRef]
- Begam, A.; Paramanik, B.; Dutta, S.; Dutta, G.; Bhattacharjee, S. Combating the Effects of Climate Change through Regenerative Organic Agriculture. In Regenerative Agriculture; CRC Press: Boca Raton, FL, USA; pp. 180–192.
- Stern, N. Why Are We Waiting?: The Logic, Urgency, and Promise of Tackling Climate Change; MIT Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Martínez-Valderrama, J.; Olcina, J.; Delacámara, G.; Guirado, E.; Maestre, F.T. Complex policy mixes are needed to cope with agricultural water demands under climate change. Water Resour. Manag. 2023, 37, 2805–2834. [Google Scholar] [CrossRef]
- Dhaliwal, S.K.; Salaria, P.; Kaushik, P. Pea seed proteins: A nutritional and nutraceutical update. In Grain and Seed Proteins Functionality; IntechOpen: London, UK, 2021; pp. 105–114. [Google Scholar]
- Foyer, C.H.; Lam, H.-M.; Nguyen, H.T.; Siddique, K.H.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 16112. [Google Scholar] [CrossRef] [PubMed]
- FAO. Crops and Climate Change Impact Briefs: Climate-Smart Agriculture for More Sustainable, Resilient, and Equitable Food Systems; Food & Agriculture Organization: Rome, Italy, 2022. [Google Scholar]
- Best, S. Failed Species: The Rise and Fall of the Human Empire and the Age of the Anthropocene. In Lost Kingdom: Animal Death in the Anthropocene; Vernon Press: Wilmington, DE, USA, 2024; p. 87. [Google Scholar]
- Fletcher, C.; Ripple, W.J.; Newsome, T.; Barnard, P.; Beamer, K.; Behl, A.; Bowen, J.; Cooney, M.; Crist, E.; Field, C. Earth at risk: An urgent call to end the age of destruction and forge a just and sustainable future. PNAS Nexus 2024, 3, pgae106. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Clark, M.; Williams, D.R.; Kimmel, K.; Polasky, S.; Packer, C. Future threats to biodiversity and pathways to their prevention. Nature 2017, 546, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Burlingame, B.; Lawrence, M.; Macdiarmid, J.; Dernini, S.; Oenema, S. IUNS task force on sustainable diets-linking nutrition and food systEMS. Trends Food Sci. Technol. 2022, 130, 42–50. [Google Scholar] [CrossRef]
- Tulbek, M.C.; Wang, Y.L.; Hounjet, M. Pea—A Sustainable Vegetable Protein Crop. In Sustainable Protein Sources; Elsevier: Amsterdam, The Netherlands, 2024; pp. 143–162. [Google Scholar]
- Mannucci, P.M.; Jolliet, O.; Meijaard, E.; Slavin, J.; Rasetti, M.; Aleta, A.; Moreno, Y.; Agostoni, C. Sustainable nutrition and the case of vegetable oils to match present and future dietary needs. Front. Public Health 2023, 11, 1106083. [Google Scholar] [CrossRef] [PubMed]
- Praveen Kumar, P.K.; Bishnoi, S.; Prashant Kaushik, P.K. Genetic variability, heritability and genetic advance for seed yield and other agro-morphological traits in fababean (Vicia faba L.) genotypes of different origin. Trends Biosci. 2017, 10, 1246–1248. [Google Scholar]
- Kumar, P.; Kaushik, P. Evaluation of genetic diversity in cultivated and exotic germplasm sources of faba bean using important morphological traits. bioRxiv 2020, 1, 918284. [Google Scholar] [CrossRef]
- Yahia, E.M.; García-Solís, P.; Celis, M.E.M. Contribution of fruits and vegetables to human nutrition and health. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Elsevier: Amsterdam, The Netherlands, 2019; pp. 19–45. [Google Scholar]
- Watson, C.A.; Reckling, M.; Preissel, S.; Bachinger, J.; Bergkvist, G.; Kuhlman, T.; Lindström, K.; Nemecek, T.; Topp, C.F.; Vanhatalo, A. Grain legume production and use in European agricultural systems. In Advances in Agronomy; Academic Press: Cambridge, CA, USA, 2017; pp. 235–303. [Google Scholar]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of climate change on agriculture and its mitigation strategies: A review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Varzakas, T.; Smaoui, S. Global food security and sustainability issues: The road to 2030 from nutrition and sustainable healthy diets to food systems change. Foods 2024, 13, 306. [Google Scholar] [CrossRef]
- Wang, T.; Yao, P.-H.; Shen, C.-C.; Chawchai, S.; Torfstein, A.; Sinha, A.; Xu, H.; Yu, T.-L.; Lin, F.; Wang, X. Anthropogenically-induced atmospheric Pb cycle in low-latitude Asia since the industrial revolution recorded by high-resolution stalagmites. Glob. Planet. Chang. 2024, 232, 104337. [Google Scholar] [CrossRef]
- Sharma, M.; Kaushal, R.; Kaushik, P.; Ramakrishna, S. Carbon farming: Prospects and challenges. Sustainability 2021, 13, 11122. [Google Scholar] [CrossRef]
- Vijayalakshmi, D.; Priya, J.R.; Vinitha, A.; Ramya, G. Interactive effects of elevated CO2 with combined heat and drought stresses on the physiology and yield of C3 and C4 plants. J. Crop Sci. Biotechnol. 2024, 27, 1–16. [Google Scholar] [CrossRef]
- Heckman, R.W.; Pereira, C.G.; Aspinwall, M.J.; Juenger, T.E. Physiological responses of C4 perennial bioenergy grasses to climate change: Causes, consequences, and constraints. Annu. Rev. Plant Biol. 2024, 75. [Google Scholar] [CrossRef] [PubMed]
- Streck, N.A. Climate change and agroecosystems: The effect of elevated atmospheric CO2 and temperature on crop growth, development, and yield. Ciência Rural. 2005, 35, 730–740. [Google Scholar] [CrossRef]
- Ebi, K.L.; Anderson, C.L.; Hess, J.J.; Kim, S.-H.; Loladze, I.; Neumann, R.B.; Singh, D.; Ziska, L.; Wood, R. Nutritional quality of crops in a high CO2 world: An agenda for research and technology development. Environ. Res. Lett. 2021, 16, 064045. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob. Chang. Biol. 2021, 27, 27–49. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.D.; Chatterton, S.; Soolanayakanahally, R.Y.; Subedi, U.; Chen, G.; Acharya, S.N. Potential effects of a high CO2 future on leguminous species. Plant-Environ. Interact. 2020, 1, 67–94. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.A.; Leakey, A.D.; Ainsworth, E.A. How seasonal temperature or water inputs affect the relative response of C3 crops to elevated [CO2]: A global analysis of open top chamber and free air CO2 enrichment studies. Food Energy Secur. 2014, 3, 33–45. [Google Scholar] [CrossRef]
- Coast, O.; Scafaro, A.P.; Bramley, H.; Taylor, N.L.; Atkin, O.K. Photosynthesis in newly developed leaves of heat-tolerant wheat acclimates to long-term nocturnal warming. J. Exp. Bot. 2024, 75, 962–978. [Google Scholar] [CrossRef]
- Franks, P.J.; Adams, M.A.; Amthor, J.S.; Barbour, M.M.; Berry, J.A.; Ellsworth, D.S.; Farquhar, G.D.; Ghannoum, O.; Lloyd, J.; McDowell, N. Sensitivity of plants to changing atmospheric CO2 concentration: From the geological past to the next century. New Phytol. 2013, 197, 1077–1094. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Gulías, J.; Figueroa, C.M.; Iñiguez, C.; Clemente-Moreno, M.J.; Nunes-Nesi, A.; Fernie, A.R.; Cavieres, L.A.; Bravo, L.A.; García-Plazaola, J.I. How do vascular plants perform photosynthesis in extreme environments? An integrative ecophysiological and biochemical story. Plant J. 2020, 101, 979–1000. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising atmospheric carbon dioxide: Plants FACE the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Ainsworth, E.; Bernacchi, C.; Davey, P.; Morgan, P.; Hymus, G.; Leakey, A.; Osborne, C. Long-term responses of photosynthesis and stomata to elevated [CO2] in managed systems. Manag. Ecosyst. CO2 Case Stud. Process. Perspect. 2006, 187, 253–270. [Google Scholar]
- Kolaksazov, M.I. Part 2: Aspects of the relation between photosynthesis and crop productivity. Bot. Lett. 2024, 1–14. [Google Scholar] [CrossRef]
- Shah, M.H.; Pramanik, K.; Ajaharuddin, S.; Kundu, S.; Alam, M.A.; Islam, S.; Pal, A.; Atta, K.; Pande, C.B. Effects of climate change on plant growth and development. In Climate-Resilient Agriculture; Apple Academic Press: Cambridge, MA, USA, 2024; pp. 9–79. [Google Scholar]
- Soares, J.; Deuchande, T.; Valente, L.M.P.; Pintado, M.; Vasconcelos, M.W. Growth and Nutritional Responses of Bean and Soybean Genotypes to Elevated CO2 in a Controlled Environment. Plants 2019, 8, 465. [Google Scholar] [CrossRef] [PubMed]
- Brar, N.S.; Kumar, T.; Kaushik, P. Integration of technologies under climate change for profitability in vegetable cultivation: An outlook. Agric. Sci. Agron. 2020, preprint. [Google Scholar]
- Kebe, I.; Diallo, I.; Sylla, M.B.; De Sales, F.; Diedhiou, A. Late 21st century projected changes in the relationship between precipitation, African easterly jet, and African easterly waves. Atmosphere 2020, 11, 353. [Google Scholar] [CrossRef]
- Kumari, S.; Najar, A.; Nader, A.; Moukahel, A. Epidemiology and Management of Legume and Cereal Viruses in Arab and Mediterranean Regions. 2022. Available online: https://hdl.handle.net/10568/128128 (accessed on 27 June 2024).
- Cabrerizo, P.M.; González, E.M.; Aparicio-Tejo, P.M.; Arrese-Igor, C. Continuous CO2 enrichment leads to increased nodule biomass, carbon availability to nodules and activity of carbon-metabolising enzymes but does not enhance specific nitrogen fixation in pea. Physiol. Plant. 2001, 113, 33–40. [Google Scholar] [CrossRef]
- Saha, S.; Chakraborty, D.; Pal, M.; Nagarajan, S. Impact of elevated CO2 on utilization of soil moisture and associated soil biophysical parameters in pigeon pea (Cajanus cajan L.). Agric. Ecosyst. Environ. 2011, 142, 213–221. [Google Scholar] [CrossRef]
- Sathish, P.; Vijay Kumar, G.; Jyothi Lakshmi, N.; Vanaja, M.; Yadav, S.; Vagheera, P. Impact of CO2 enhancement on photosynthesis and protein profile-response studies with a CO2 responsive blackgram genotype. Int. J. Appl. Biol. Pharm. Technol. 2014, 5, 93–98. [Google Scholar]
- Nasser, R.R.; Fuller, M.; Jellings, A. Effect of elevated CO2 and nitrogen levels on lentil growth and nodulation. Agron. Sustain. Dev. 2008, 28, 175–180. [Google Scholar] [CrossRef]
- Mishra, A.K.; Agrawal, S. Biochemical and physiological characteristics of tropical mung bean (Vigna radiata L.) cultivars against chronic ozone stress: An insight to cultivar-specific response. Protoplasma 2015, 252, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Srinivasarao, C.; Kundu, S.; Shanker, A.K.; Naik, R.P.; Vanaja, M.; Venkanna, K.; Sankar, G.M.; Rao, V. Continuous cropping under elevated CO2: Differential effects on C4 and C3 crops, soil properties and carbon dynamics in semi-arid alfisols. Agric. Ecosyst. Environ. 2016, 218, 73–86. [Google Scholar] [CrossRef]
- Yamakawa, T.; Ikeda, T.; Ishizuka, J. Effects of CO2 concentration in rhizosphere on nodulation and N2 fixation of soybean and cowpea. Soil Sci. Plant Nutr. 2004, 50, 713–720. [Google Scholar] [CrossRef]
- Dey, S.K.; Chakrabarti, B.; Prasanna, R.; Pratap, D.; Singh, S.D.; Purakayastha, T.J.; Pathak, H. Elevated carbon dioxide level along with phosphorus application and cyanobacterial inoculation enhances nitrogen fixation and uptake in cowpea crop. Arch. Agron. Soil Sci. 2017, 63, 1927–1937. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Siddique, K.H.; Farooq, M.; Thornton, P.K.; Ortiz, R. Using biotechnology-led approaches to uplift cereal and food legume yields in dryland environments. Front. Plant Sci. 2018, 9, 393361. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef]
- Gupta, S.; Ranjan, R.; Bijlwan, A.; Jha, A.; Jena, R.K. Enhancing Water Use Efficiency Through Trait-Based Approaches. In Plant Functional Traits for Improving Productivity; Springer: Berlin/Heidelberg, Germany, 2024; pp. 133–147. [Google Scholar]
- Ghadirnezhad Shiade, S.R.; Fathi, A.; Taghavi Ghasemkheili, F.; Amiri, E.; Pessarakli, M. Plants’ responses under drought stress conditions: Effects of strategic management approaches—A review. J. Plant Nutr. 2023, 46, 2198–2230. [Google Scholar] [CrossRef]
- Toreti, A.; Belward, A.; Perez-Dominguez, I.; Naumann, G.; Luterbacher, J.; Cronie, O.; Seguini, L.; Manfron, G.; Lopez-Lozano, R.; Baruth, B. The exceptional 2018 European water seesaw calls for action on adaptation. Earth’s Future 2019, 7, 652–663. [Google Scholar] [CrossRef]
- Das, A.; Thoithoi Devi, M.; Babu, S.; Ansari, M.; Layek, J.; Bhowmick, S.; Yadav, G.S.; Singh, R. Cereal-legume cropping system in indian himalayan region for food and environmental sustainability. Legum. Soil Health Sustain. Manag. 2018, 33–76. [Google Scholar] [CrossRef]
- Yang, L.; Song, W.; Xu, C.; Sapey, E.; Jiang, D.; Wu, C. Effects of high night temperature on soybean yield and compositions. Front. Plant Sci. 2023, 14, 1065604. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Effects of drought, heat and their interaction on the growth, yield and photosynthetic function of lentil (Lens culinaris Medikus) genotypes varying in heat and drought sensitivity. Front. Plant Sci. 2017, 8, 298024. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S.; Kataria, S.; Rastogi, A. The interactive effect of high temperature and water deficit stress on nitrogen fixation, photosynthesis, chlorophyll fluorescence, seed yield and quality in soybean (Glycine max). Plant Physiol. Rep. 2024, 29, 125–140. [Google Scholar] [CrossRef]
- Bennett, E.J.; Roberts, J.A.; Wagstaff, C. The role of the pod in seed development: Strategies for manipulating yield. New Phytol. 2011, 190, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, E.A.; Nikolic, M.; White, P.J.; Xu, G. Mineral nutrition, yield, and source–sink relationships. In Marschner’s Mineral Nutrition of Plants; Elsevier: Amsterdam, The Netherlands, 2023; pp. 131–200. [Google Scholar]
- Rawal, H.; Nautiyal, Y.; Sharma, B.; Tiwari, S. Microbial Inoculants and Their Role in Abiotic Stress Management. In Microbial Inoculants: Applications for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2024; pp. 163–201. [Google Scholar]
- Singh, S.; Singh, D.; Kumar, G.; Archana, T.; Rana, M.; Srivastava, S. Symbiotic associations between microbes and host plants. In Microbiome Drivers of Ecosystem Function; Elsevier: Amsterdam, The Netherlands, 2024; pp. 145–179. [Google Scholar]
- Samarah, N.; Mullen, R.; Cianzio, S.; Scott, P. Dehydrin-like proteins in soybean seeds in response to drought stress during seed filling. Crop Sci. 2006, 46, 2141–2150. [Google Scholar] [CrossRef]
- Maleki, A.; Naderi, A.; Naseri, R.; Fathi, A.; Bahamin, S.; Maleki, R. Physiological performance of soybean cultivars under drought stress. Bull. Environ. Pharmacol. Life Sci. 2013, 2, 38–44. [Google Scholar]
- Kobraee, S.; Shamsi, K.; Rasekhi, B. Soybean production under water deficit conditions. Ann. Biol. Res. 2011, 2, 423–434. [Google Scholar]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Nayyar, H.; Kaur, S.; Singh, S.; Upadhyaya, H.D. Differential sensitivity of Desi (small-seeded) and Kabuli (large-seeded) chickpea genotypes to water stress during seed filling: Effects on accumulation of seed reserves and yield. J. Sci. Food Agric. 2006, 86, 2076–2082. [Google Scholar] [CrossRef]
- Varshney, R.K.; Thudi, M.; Nayak, S.N.; Gaur, P.M.; Kashiwagi, J.; Krishnamurthy, L.; Jaganathan, D.; Koppolu, J.; Bohra, A.; Tripathi, S. Genetic dissection of drought tolerance in chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2014, 127, 445–462. [Google Scholar] [CrossRef]
- Ogbonnaya, C.; Sarr, B.; Brou, C.; Diouf, O.; Diop, N.; Roy-Macauley, H. Selection of cowpea genotypes in hydroponics, pots, and field for drought tolerance. Crop Sci. 2003, 43, 1114–1120. [Google Scholar] [CrossRef]
- Kyei-Boahen, S.; Savala, C.E.; Chikoye, D.; Abaidoo, R. Growth and yield responses of cowpea to inoculation and phosphorus fertilization in different environments. Front. Plant Sci. 2017, 8, 238588. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Serna, R.; Kohashi-Shibata, J.; Acosta-Gallegos, J.A.; Trejo-López, C.; Ortiz-Cereceres, J.; Kelly, J.D. Biomass distribution, maturity acceleration and yield in drought-stressed common bean cultivars. Field Crops Res. 2004, 85, 203–211. [Google Scholar] [CrossRef]
- Martínez, J.-P.; Silva, H.; Ledent, J.-F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 26, 30–38. [Google Scholar] [CrossRef]
- Ghanbarı, A.A.; Mousavı, S.H.; Gorjı, A.M.; Rao, I. Effects of water stress on leaves and seeds of bean (Phaseolus vulgaris L.). Turk. J. Field Crops 2013, 18, 73–77. [Google Scholar]
- Nam, N.; Chauhan, Y.; Johansen, C. Effect of timing of drought stress on growth and grain yield of extra-short-duration pigeonpea lines. J. Agric. Sci. 2001, 136, 179–189. [Google Scholar] [CrossRef]
- Ahmad, A.; Selim, M.M.; Alderfasi, A.; Afzal, M. Effect of drought stress on mung bean (Vigna radiata L.) under arid climatic conditions of Saudi Arabia. Ecosyst. Sustain. Dev. 2015, 192, 185–193. [Google Scholar]
- Ranawake, A.; Dahanayaka, N.; Amarasingha, U.; Rodrigo, W.; Rodrigo, U. Effect of water stress on growth and yield of mung bean (Vigna radiata L.). Trop. Agric. Res. Ext. 2012, 14, 76–79. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Hosseinzadeh-Mahootchy, A. Changes in seed vigour of faba bean (Vicia faba L.) cultivars during development and maturity. Seed Sci. Technol. 2009, 37, 713–720. [Google Scholar] [CrossRef]
- Allahmoradi, P.; Mansourifar, C.; Saiedi, M.; Jalali Honarmand, S. Effect of different water deficiency levels on some antioxidants at different growth stages of lentil (Lens culinaris L.). Adv. Environ. Biol 2013, 7, 535–543. [Google Scholar]
- Shrestha, R.; Turner, N.; Siddique, K.; Turner, D.; Speijers, J. A water deficit during pod development in lentils reduces flower and pod numbers but not seed size. Aust. J. Agric. Res. 2006, 57, 427–438. [Google Scholar] [CrossRef]
- Chilakala, A.R.; Mali, K.V.; Irulappan, V.; Patil, B.S.; Pandey, P.; Rangappa, K.; Ramegowda, V.; Kumar, M.N.; Puli, C.O.R.; Mohan-Raju, B. Combined drought and heat stress influences the root water relation and determine the dry root rot disease development under field conditions: A study using contrasting chickpea genotypes. Front. Plant Sci. 2022, 13, 890551. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Prasad, P.V.; Boyle, D.; Schapaugh, W. High-temperature stress and soybean leaves: Leaf anatomy and photosynthesis. Crop Sci. 2011, 51, 2125–2131. [Google Scholar] [CrossRef]
- Norton, M.R.; Malinowski, D.P.; Volaire, F. Plant drought survival under climate change and strategies to improve perennial grasses. A review. Agron. Sustain. Dev. 2016, 36, 29. [Google Scholar] [CrossRef]
- Barendrecht, M.H.; Matanó, A.; Mendoza, H.; Weesie, R.; Rohse, M.; Koehler, J.; de Ruiter, M.; Garcia, M.; Mazzoleni, M.; Aerts, J.C. Exploring drought-to-flood interactions and dynamics: A global case review. Wiley Interdiscip. Rev. Water 2024, 11, e1726. [Google Scholar] [CrossRef]
- Subbarao, G.; Johansen, C.; Slinkard, A.; Nageswara Rao, R.; Saxena, N.; Chauhan, Y.; Lawn, R. Strategies for improving drought resistance in grain legumes. Crit. Rev. Plant Sci. 1995, 14, 469–523. [Google Scholar] [CrossRef]
- Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.M.; Anwar, M.P.; Fahad, S.; Datta, R.; Islam, A.A. Drought stress in grain legumes: Effects, tolerance mechanisms and management. Agronomy 2021, 11, 2374. [Google Scholar] [CrossRef]
- Fang, X.; Turner, N.C.; Yan, G.; Li, F.; Siddique, K.H. Flower numbers, pod production, pollen viability, and pistil function are reduced and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef]
- Varshney, R.K.; Roorkiwal, M.; Sorrells, M.E. Genomic Selection for Crop Improvement: New Molecular Breeding Strategies for Crop Improvement; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Newsham, A.; Kohnstamm, S.; Naess, L.O.; Atela, J. Agricultural Commercialisation Pathways: Climate Change and Agriculture; IDS/Future Agricultures Consortium: Brighton, UK, 2018. [Google Scholar]
- Garwi, J.; Masengu, R.; Chiwaridzo, O.T. Emerging Technologies and Marketing Strategies for Sustainable Agriculture; IGI Global: Hershey, PA, USA, 2024. [Google Scholar]
- Mehboob, I.; Naveed, M.; Zahir, Z.A.; Sessitsch, A. Potential of rhizosphere bacteria for improving Rhizobium-legume symbiosis. In Plant Microbe Symbiosis: Fundamentals and Advances; Springer: Berlin/Heidelberg, Germany, 2013; pp. 305–349. [Google Scholar]
- Naqqash, T.; Malik, K.A.; Imran, A.; Hameed, S.; Shahid, M.; Hanif, M.K.; Majeed, A.; Arshad, M.; van Elsas, J.D. Isolation and characterization of Rhizobium from non-leguminous potato plants: New frontiers in Rhizobium research. Biol. Fertil. Soils 2024, 60, 307–325. [Google Scholar] [CrossRef]
- Baruah, U.; Das, S.; Kalita, P.; Saikia, M.; Bhougal, S.; Pal, S.; Das, R. High-night temperature-induced changes in chlorophyll fluorescence, gas exchange, and leaf anatomy determine grain yield in rice varieties. J. Plant Growth Regul. 2023, 42, 5538–5557. [Google Scholar] [CrossRef]
- Sharma, A.; Yadav, R.; Sheoran, R.; Kaushik, D.; Mohanta, T.K.; Sharma, K.; Yadav, A.; Dhanda, P.S.; Kaushik, P. Estimation of heterosis and the combining ability effect for yield and its attributes in field pea (Pisum sativum L.) using PCA and GGE biplots. Horticulturae 2023, 9, 256. [Google Scholar] [CrossRef]
- Aisha Renju, N. Evaluation of Selectively Fertilized Coconut Hybrids (Cocos nucifera L.) for Water Use Efficiency through Stable Isotope Discrimination; Department of Plant Physiology, College of Agriculture, Vellayani: Kerala, India, 2015. [Google Scholar]
- Basu, P.; Singh, U.; Meena, S.K.; Gurumurthy, S.; Kumar, V.; Tewari, K.; Das, K.; Sharma, K.; Chaturvedi, S. Implication of Climate Change on the Productivity of Legumes. In Climate Change and Legumes; CRC Press: Boca Raton, FL, USA, 2023; pp. 207–250. [Google Scholar]
- Nelson, M.N.; Berger, J.D.; Erskine, W. Flowering time control in annual legumes: Prospects in a changing global climate. Plant Sci. Rev. 2010, 5, 49–62. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; HanumanthaRao, B.; Nair, R.M.; Vara Prasad, P.; Kumar, S.; Gaur, P.M.; Farooq, M.; Siddique, K.H.; Varshney, R.K. Food legumes and rising temperatures: Effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front. Plant Sci. 2017, 8, 298433. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.S.; Farooq, M.; Asch, F.; Krishna, J.S.; Prasad, P.V.; Siddique, K.H. Thermal stress impacts reproductive development and grain yield in rice. Plant Physiol. Biochem. 2017, 115, 57–72. [Google Scholar] [CrossRef]
- Gan, Y.; Miller, P.; McConkey, B.; Zentner, R.; Stevenson, F.; McDonald, C. Influence of diverse cropping sequences on durum wheat yield and protein in the semiarid northern great plains. Agron. J. 2003, 95, 245–252. [Google Scholar] [CrossRef]
- Praba, M.L.; Cairns, J.; Babu, R.; Lafitte, H. Identification of physiological traits underlying cultivar differences in drought tolerance in rice and wheat. J. Agron. Crop Sci. 2009, 195, 30–46. [Google Scholar] [CrossRef]
- Redden, R.J.; Hatfield, J.L.; Vara Prasad, P.; Ebert, A.W.; Yadav, S.S.; O’Leary, G.J. Temperature, climate change, and global food security. Temp. Plant Dev. 2014, 181–202. [Google Scholar] [CrossRef]
- Farooq, M.; Nadeem, F.; Gogoi, N.; Ullah, A.; Alghamdi, S.S.; Nayyar, H.; Siddique, K.H. Heat stress in grain legumes during reproductive and grain-filling phases. Crop Pasture Sci. 2017, 68, 985–1005. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Bhandari, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Impact of heat stress during seed filling on seed quality and seed yield in lentil (Lens culinaris Medikus) genotypes. J. Sci. Food Agric. 2018, 98, 5134–5141. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.V.; Boyle, D.; Schapaugh, W. Soybean pollen anatomy, viability and pod set under high temperature stress. J. Agron. Crop Sci. 2013, 199, 171–177. [Google Scholar] [CrossRef]
- Abeysingha, G.L. The Effect of Auxins on Seed Yield Parameters in Wheat, Pea and Canola Grown under Controlled Environment and Western Canadian Field Conditions. Master’s Thesis, University of Alberta Libraries, Edmonton, AB, Canada, 2015. [Google Scholar]
- Bindumadhava, H.; Nair, R.; Nayyar, H.; Riley, J.; Easdown, W. Mungbean production under a changing climate-insights from growth physiology. Mysore J. Agric. Sci. 2017, 51, 21–26. [Google Scholar]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Karthik, S.; Reddy, M.; Yashaswini, G. Climate change and its potential impacts on insect-plant interactions. In The Nature, Causes, Effects and Mitigation of Climate Change on the Environment; IntechOpen: London, UK, 2021; Volume 10. [Google Scholar]
- Macfadyen, S.; McDonald, G.; Hill, M.P. From species distributions to climate change adaptation: Knowledge gaps in managing invertebrate pests in broad-acre grain crops. Agric. Ecosyst. Environ. 2018, 253, 208–219. [Google Scholar] [CrossRef]
- Mohapatra, D.; Kar, A.; Giri, S.K. Insect pest management in stored pulses: An overview. Food Bioprocess Technol. 2015, 8, 239–265. [Google Scholar] [CrossRef]
- Dutta, A.; Trivedi, A.; Nath, C.P.; Gupta, D.S.; Hazra, K.K. A comprehensive review on grain legumes as climate-smart crops: Challenges and prospects. Environ. Chall. 2022, 7, 100479. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Mwaipopo, B.; Rajamäki, M.-L.; Ngowi, N.; Nchimbi-Msolla, S.; Njau, P.J.; Valkonen, J.P.; Mbanzibwa, D.R. Next-generation sequencing-based detection of common bean viruses in wild plants from Tanzania and their mechanical transmission to common bean plants. Plant Dis. 2021, 105, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Xavier, A.; Hall, B.; Hearst, A.A.; Cherkauer, K.A.; Rainey, K.M. Genetic architecture of phenomic-enabled canopy coverage in Glycine max. Genetics 2017, 206, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, L.; Ogle, S.; Cheng, K.; Xu, X.; Li, Y.; Pan, G. Future climate change may pose pressures on greenhouse gas emission reduction in China’s rice production. Geoderma 2023, 440, 116732. [Google Scholar] [CrossRef]
- Scaven, V.L.; Rafferty, N.E. Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions. Current Zoology 2013, 59, 418–426. [Google Scholar] [CrossRef]
- Kyriakopoulos, G.L.; Sebos, I.; Triantafyllou, E.; Stamopoulos, D.; Dimas, P. Benefits and synergies in addressing climate change via the implementation of the common agricultural policy in Greece. Appl. Sci. 2023, 13, 2216. [Google Scholar] [CrossRef]
- Shrestha, S. Effects of climate change in agricultural insect pest. Acta Sci. Agric. 2019, 3, 74–80. [Google Scholar] [CrossRef]
- Pathak, H.; Aggarwal, P.K.; Singh, S.D. Climate Change Impact, Adaptation and Mitigation in Agriculture: Methodology for Assessment and Applications; Indian Agricultural Research Institute: New Delhi, India, 2012; Volume 302, p. 19. [Google Scholar]
- Staley, J.T.; Hodgson, C.J.; Mortimer, S.R.; Morecroft, M.D.; Masters, G.J.; Brown, V.K.; Taylor, M.E. Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates. Eur. J. Soil Biol. 2007, 43, 189–198. [Google Scholar] [CrossRef]
- Johnson, S.N.; Anderson, E.A.; Dawson, G.; Griffiths, D.W. Varietal susceptibility of potatoes to wireworm herbivory. Agric. For. Entomol. 2008, 10, 167–174. [Google Scholar] [CrossRef]
- Gregory, P.J.; Johnson, S.N.; Newton, A.C.; Ingram, J.S. Integrating pests and pathogens into the climate change/food security debate. J. Exp. Bot. 2009, 60, 2827–2838. [Google Scholar] [CrossRef] [PubMed]
- Yihdego, Y.; Salem, H.S.; Muhammed, H.H. Agricultural pest management policies during drought: Case studies in Australia and the state of Palestine. Nat. Hazards Rev. 2019, 20, 05018010. [Google Scholar] [CrossRef]
- Nasr Esfahani, M.; Sulieman, S.; Schulze, J.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. Mechanisms of physiological adjustment of N2 fixation in Cicer arietinum L. (chickpea) during early stages of water deficit: Single or multi-factor controls. Plant J. 2014, 79, 964–980. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, L.K.; Robertson, A.; Schipanski, M.; Calderon, F.J.; Wallenstein, M.D.; Cotrufo, M.F. Addressing the soil carbon dilemma: Legumes in intensified rotations regenerate soil carbon while maintaining yields in semi-arid dryland wheat farms. Agric. Ecosyst. Environ. 2022, 330, 107906. [Google Scholar] [CrossRef]
- Naira, K.P. Microbial fertilization as a preferable choice than synthetic chemical fertilization in modern farming. Adv. Agron. 2024, 183, 251. [Google Scholar]
- Kim, K.; Daly, E.J.; Gorzelak, M.; Hernandez-Ramirez, G. Soil organic matter pools response to perennial grain cropping and nitrogen fertilizer. Soil Tillage Res. 2022, 220, 105376. [Google Scholar] [CrossRef]
- Oduor, K.; Mburu, W.; Wanjiru, T. The nexus between climate change and nutrition security and the impact of this relationship on diet-related non-communicable diseases. East Afr. J. Health Sci. 2024, 7, 60–79. [Google Scholar] [CrossRef]
- Lehmann, J.; Bossio, D.A.; Kögel-Knabner, I.; Rillig, M.C. The concept and future prospects of soil health. Nat. Rev. Earth Environ. 2020, 1, 544–553. [Google Scholar] [CrossRef]
- Morton, J.F. The impact of climate change on smallholder and subsistence agriculture. Proc. Natl. Acad. Sci. USA 2007, 104, 19680–19685. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.A.; Saborio-Rodríguez, M.; Martinez-Rodríguez, M.R.; Viguera, B.; Chain-Guadarrama, A.; Vignola, R.; Alpizar, F. Climate change impacts and adaptation among smallholder farmers in Central America. Agric. Food Secur. 2018, 7, 57. [Google Scholar] [CrossRef]
- Krishna, V.V.; Feleke, S.; Marenya, P.; Abdoulaye, T.; Erenstein, O. A Strategic Framework for Adoption and Impact Studies in the CGIAR Research Program on Maize (CRP MAIZE); CIMMYT–IITA: Texcoco, Mexico, 2019; p. 24. Available online: https://repository.cimmyt.org/handle/10883/20220 (accessed on 27 June 2024).
- Varshney, R.K.; Thudi, M.; Pandey, M.K.; Tardieu, F.; Ojiewo, C.; Vadez, V.; Whitbread, A.M.; Siddique, K.H.; Nguyen, H.T.; Carberry, P.S. Accelerating genetic gains in legumes for the development of prosperous smallholder agriculture: Integrating genomics, phenotyping, systems modelling and agronomy. J. Exp. Bot. 2018, 69, 3293–3312. [Google Scholar] [CrossRef] [PubMed]
- Carr, E.R.; Onzere, S.N. Really effective (for 15% of the men): Lessons in understanding and addressing user needs in climate services from Mali. Climate Risk Management 2018, 22, 82–95. [Google Scholar] [CrossRef]
- Gordon, L.J.; Bignet, V.; Crona, B.; Henriksson, P.J.; Van Holt, T.; Jonell, M.; Lindahl, T.; Troell, M.; Barthel, S.; Deutsch, L. Rewiring food systems to enhance human health and biosphere stewardship. Environ. Res. Lett. 2017, 12, 100201. [Google Scholar] [CrossRef]
- Tigchelaar, M.; Leape, J.; Micheli, F.; Allison, E.H.; Basurto, X.; Bennett, A.; Bush, S.R.; Cao, L.; Cheung, W.W.L.; Crona, B.; et al. The vital roles of blue foods in the global food system. Glob. Food Secur. 2022, 33, 100637. [Google Scholar] [CrossRef]
- Heckelman, A.; Chappell, M.J.; Wittman, H. A polycentric food sovereignty approach to climate resilience in the Philippines. Elementa 2022, 10, 00033. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.B.; Boddey, R.M.; Gresshoff, P.M.; Hauggaard-Nielsen, H.; JR Alves, B.; Morrison, M.J. Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron. Sustain. Dev. 2012, 32, 329–364. [Google Scholar] [CrossRef]
- Kang, Y.; Khan, S.; Ma, X. Climate change impacts on crop yield, crop water productivity and food security–A review. Prog. Nat. Sci. 2009, 19, 1665–1674. [Google Scholar] [CrossRef]
- Yadav, I.; Sharma, V.; Kumar, M.; Yadav, L.P.; Mishra, A.; Singh, V.; Singh Dhanda, P.; Yadav, A.; Yadav, M.; Singh, S.K. Assessment of gene action and identification of heterotic hybrids for enhancing yield in field pea. Horticulturae 2023, 9, 997. [Google Scholar] [CrossRef]
- Singh, H.; Sethi, S.; Kaushik, P.; Fulford, A. Grafting vegetables for mitigating environmental stresses under climate change: A review. J. Water Clim. Chang. 2020, 11, 1784–1797. [Google Scholar] [CrossRef]
- Haddad, F.; Ariza, C.; Malmer, A. Building Climate-Resilient Dryland Forests and Agrosilvopastoral Production Systems: An Approach for Context-Dependent Economic, Social and Environmentally Sustainable Transformations; Food & Agriculture Organization: Rome, Italy, 2021. [Google Scholar]
- Yu, J.; Holland, J.B.; McMullen, M.D.; Buckler, E.S. Genetic design and statistical power of nested association mapping in maize. Genetics 2008, 178, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Omomowo, O.I.; Babalola, O.O. Constraints and prospects of improving cowpea productivity to ensure food, nutritional security and environmental sustainability. Front. Plant Sci. 2021, 12, 751731. [Google Scholar] [CrossRef] [PubMed]
- Barrett, C.B. Overcoming global food security challenges through science and solidarity. Am. J. Agric. Econ. 2021, 103, 422–447. [Google Scholar] [CrossRef]
- Mrabet, R. Sustainable Agriculture for Food and Nutritional Security; Academic Press: Cambridge, MA, USA, 2023; pp. 25–90. [Google Scholar] [CrossRef]
- Li, B.; Sun, C.; Li, J.; Gao, C. Targeted genome-modification tools and their advanced applications in crop breeding. Nat. Rev. Genet. 2024, 1–20. [Google Scholar] [CrossRef]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef]
- Mueller, N.; Flachs, A. Domestication, crop breeding, and genetic modification are fundamentally different processes: Implications for seed sovereignty and agrobiodiversity. Agric. Hum. Values 2021, 39, 455–472. [Google Scholar] [CrossRef]
- Acharjee, S.; Higgins, T.J. Genetic Engineering of Grain Legumes: Their Potential for Sustainable Agriculture and Food and Nutritional Security. In Genetic Enhancement in Major Food Legumes; Springer: Cham, Switzerland, 2021; pp. 91–121. [Google Scholar]
- Gulevich, A.A.; Baranova, E.N.; Polyakov, V.Y.; Kharchenko, P.N. Ultrastructural evaluation of possible results of cell selection of Medicago sativa for NaCl. Russ. Agric. Sci. 2010, 36, 100–104. [Google Scholar] [CrossRef]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Tittonell, P.; Scopel, E.; Andrieu, N.; Posthumus, H.; Mapfumo, P.; Corbeels, M.; van Halsema, G.E.; Lahmar, R.; Lugandu, S.; Rakotoarisoa, J. Agroecology-based aggradation-conservation agriculture (ABACO): Targeting innovations to combat soil degradation and food insecurity in semi-arid Africa. Field Crops Res. 2012, 132, 168–174. [Google Scholar] [CrossRef]
- Rosegrant, M.W.; Koo, J.; Cenacchi, N.; Ringler, C.; Robertson, R.D.; Fisher, M.; Cox, C.M.; Garrett, K.; Perez, N.D.; Sabbagh, P. Food Security in a World of Natural Resource Scarcity: The Role of Agricultural Technologies; International Food Policy Research Institute: Washington, DC, USA, 2014. [Google Scholar]
- Andrews-Speed, P.; Bleischwitz, R.; Boersma, T.; Johnson, C.; Kemp, G.; Vandeveer, S. The Global Resource Nexus. The Struggles for Land, Energy, Water, Minerals; Transatlantic Academy: Washington, DC, USA, 2013. [Google Scholar]
- Smýkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Varshney, R.K. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef]
- Samantara, K.; Bohra, A.; Mohapatra, S.R.; Prihatini, R.; Asibe, F.; Singh, L.; Varshney, R.K. Breeding more crops in less time: A perspective on speed breeding. Biology 2022, 11, 275. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Sharma, A.; Sarkar, P.K. Conventional and emerging processing techniques for the post-harvest reduction of antinutrients in edible legumes. Appl. Food Res. 2022, 2, 100112. [Google Scholar] [CrossRef]
- Popoola, J.O.; Aworunse, O.S.; Ojuederie, O.B.; Adewale, B.D.; Ajani, O.C.; Oyatomi, O.A.; Obembe, O.O. The exploitation of orphan legumes for food, income, and nutrition security in Sub-Saharan Africa. Front. Plant Sci. 2022, 13, 782140. [Google Scholar] [CrossRef] [PubMed]
- Shamsutdinov, Z.S.; Shamsutdinov, N.Z. Biogeocenotic principles and methods of environmental restoration of desert pasture ecosystems in Central Asia. Arid Ecosyst. 2012, 2, 139–149. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, X.; Liu, X.; Li, Y.; Luo, Y.; Lian, J.; Duan, Y. Research in desertification mechanism and reversion techniques promotes sustainable restoration of degraded ecosystem and agro-pastoral development. Bull. Chin. Acad. Sci. Chin. Version 2019, 34, 832–840. [Google Scholar]
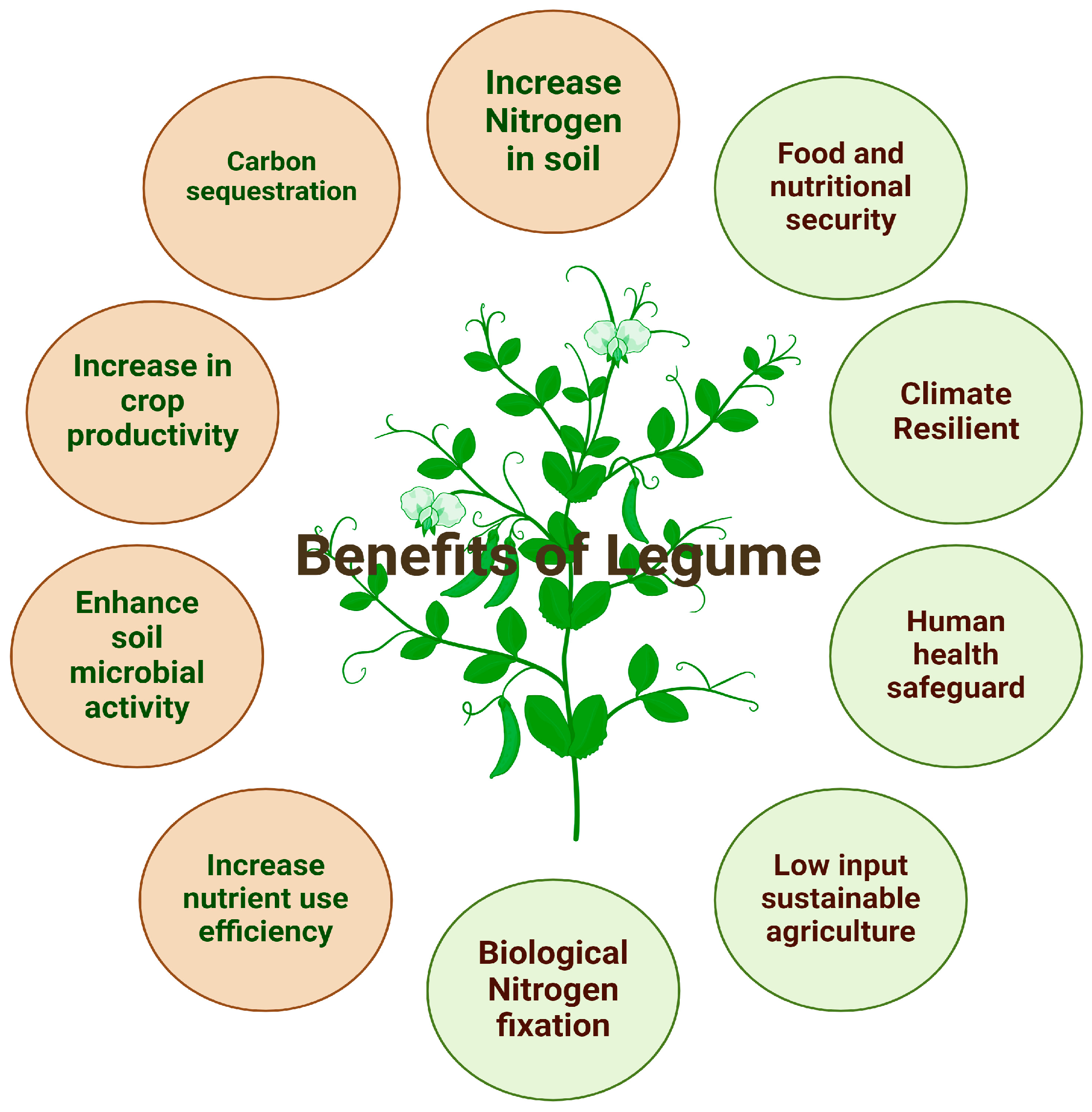
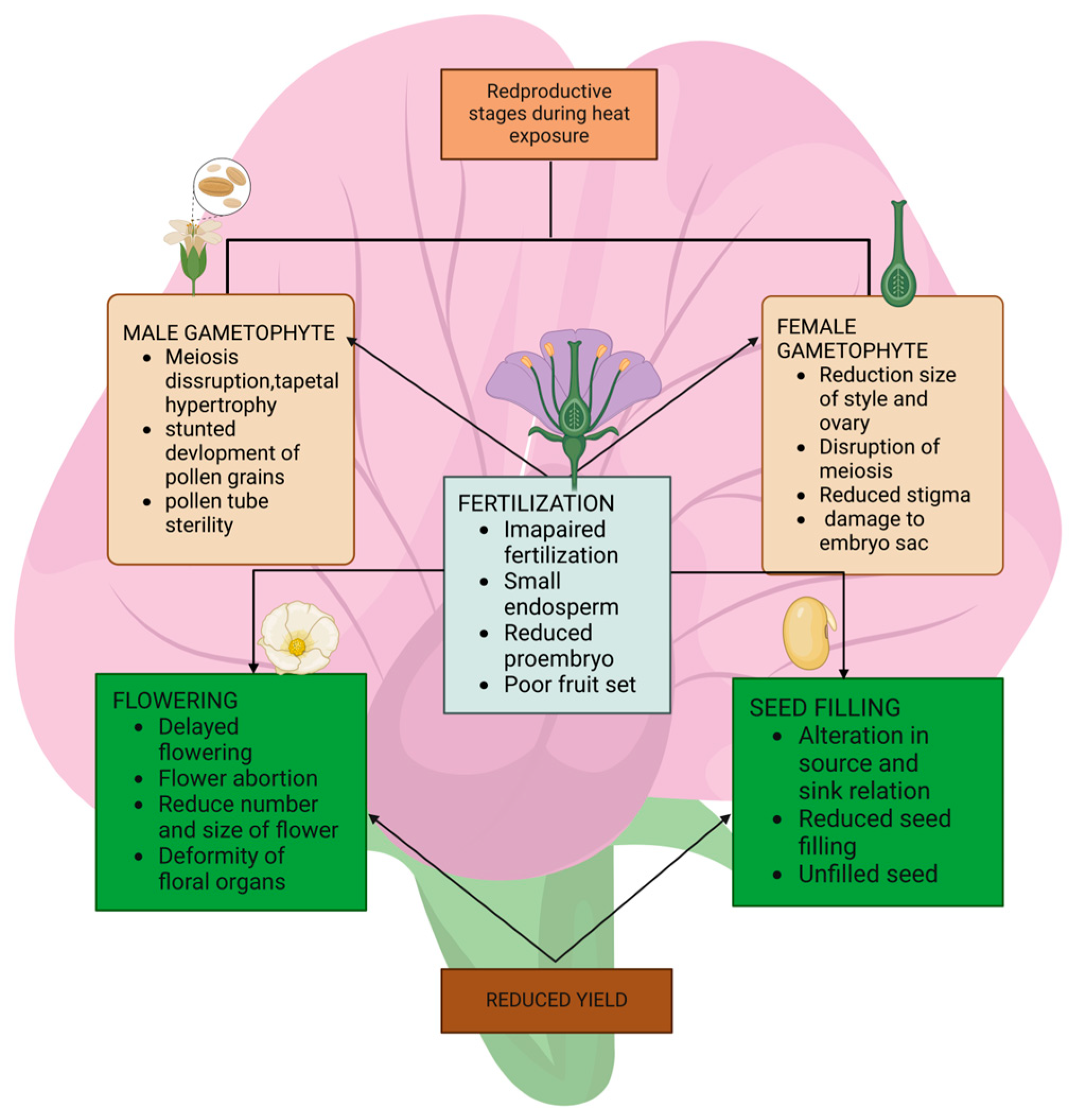
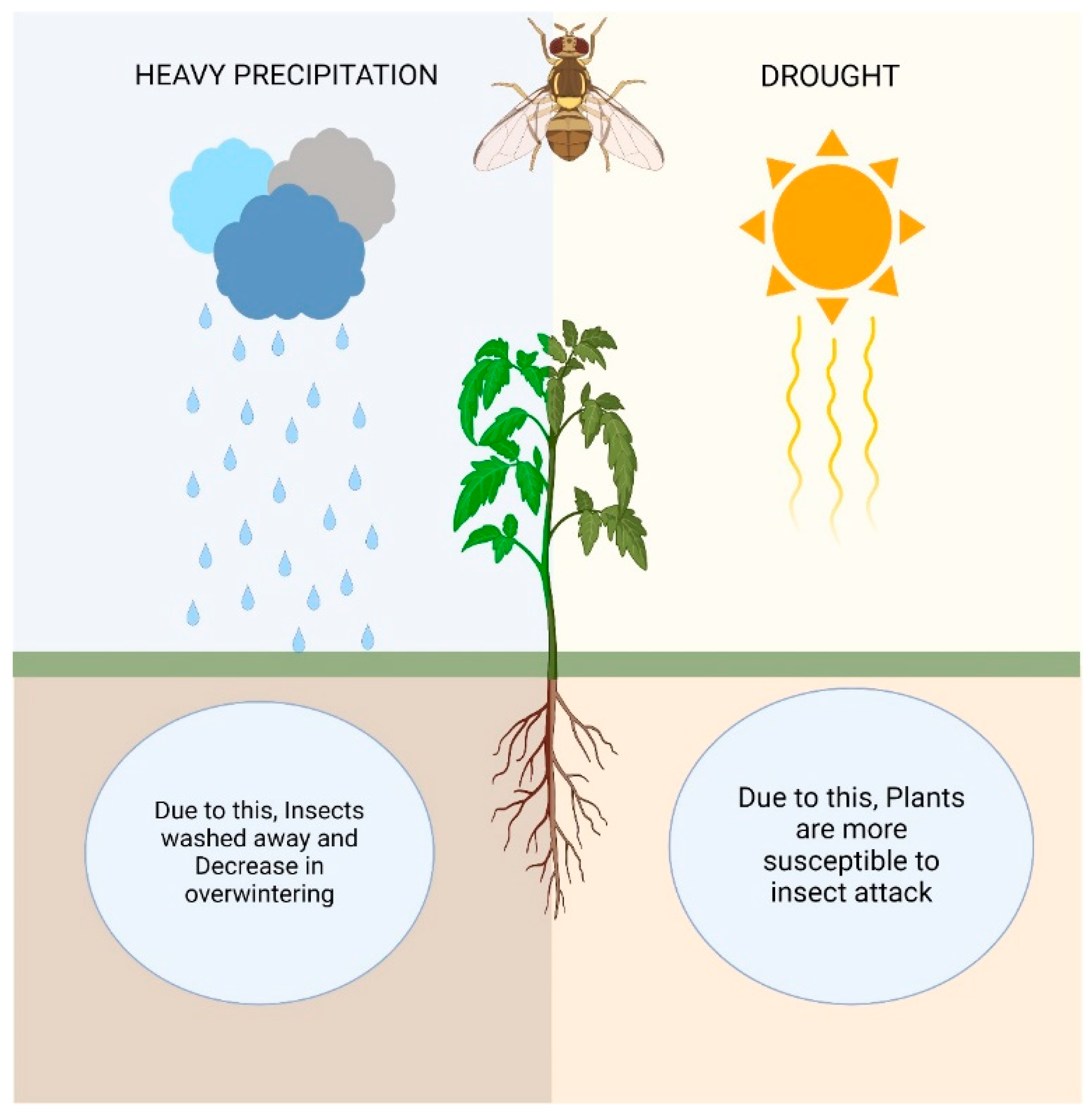
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).