
A New Direction for the Green, Environmentally Friendly and Sustainable Bioproduction of Aminobenzoic Acid and Its Derivatives
 and
and
Abstract
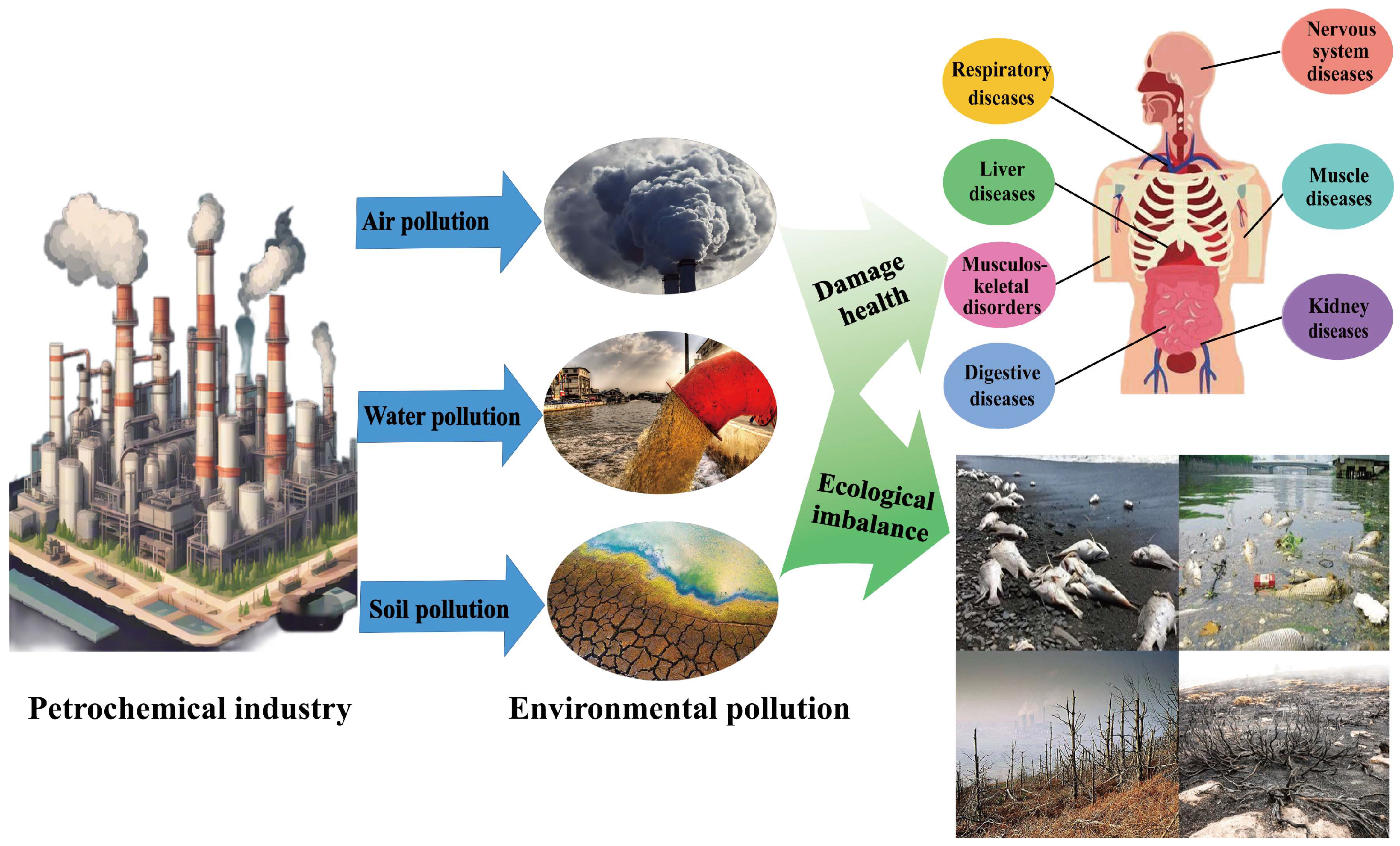
:1. Introduction
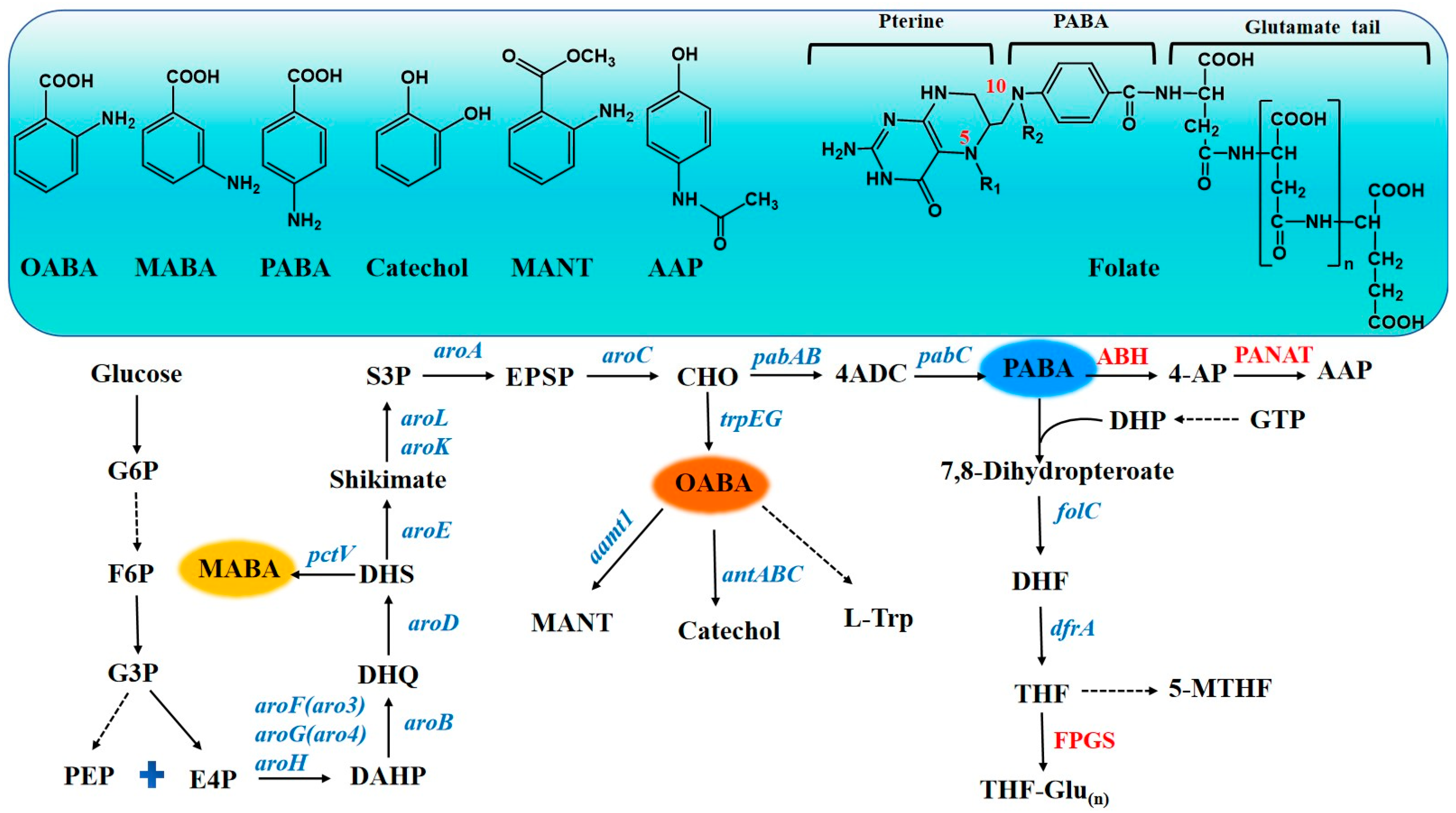
2. Synthesis Pathway
2.1. OABA and Its Derivatives
2.2. MABA
2.3. PABA and Its Derivatives
3. Biosynthesis of OABA and Its Derivatives
3.1. OABA
3.2. Methyl Anthranilate
3.3. Catechol
4. Biosynthesis of MABA
5. Biosynthesis of PABA and Its Derivatives
5.1. PABA
5.2. Folate
5.3. Acetaminophen
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lu, Y.; Xu, W.; Sun, H.; Jin, J.; Liu, H.; Jin, S.; Wang, D.; Guo, M. Single-crystal and molecular structures of six hydrogen-bonding 3D supramolecular salts from 2-aminobenzoic acid, 3-aminobenzoic acid, 4-aminobenzoic acid, and acidic components. J. Mol. Struct. 2019, 1178, 639–654. [Google Scholar] [CrossRef]
- Walsh, C.T.; Haynes, S.W.; Ames, B.D. Aminobenzoates as building blocks for natural product assembly lines. Nat. Prod. Rep. 2012, 29, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Wiklunda, P.; Bergman, J. The Chemistry of Anthranilic Acid. Curr. Org. Synth. 2006, 3, 379–402. [Google Scholar] [CrossRef]
- Zhang, Y.; Anaya-Sanchez, A.; Portnoy Daniel, A. para-Aminobenzoic Acid Biosynthesis Is Required for Listeria monocytogenes Growth and Pathogenesis. Infect. Immun. 2022, 90, e00207–e00222. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Guo, Z.; Wang, Y.; Xiao, Y.; Bao, Y. Solubility study of m-aminobenzoic acid form III in different mono-solvents by thermodynamic analysis and molecular simulation. J. Mol. Liq. 2022, 354, 118871. [Google Scholar] [CrossRef]
- Kasap, E.Y.; Grishin, D.V. To a Question on the Mechanism of the Antimicrobial Action of Ortho-Benzoic Sulfimide. Pharmaceuticals 2020, 13, 461. [Google Scholar] [CrossRef]
- Fernandez-Cabezon, L.; Rosich, I.B.B.; Kozaeva, E.; Gurdo, N.; Nikel, P.I. Dynamic flux regulation for high-titer anthranilate production by plasmid-free, conditionally-auxotrophic strains of Pseudomonas putida. Metab. Eng. 2022, 73, 11–25. [Google Scholar] [CrossRef]
- Kuepper, J.; Dickler, J.; Biggel, M.; Behnken, S.; Jager, G.; Wierckx, N.; Blank, L.M. Metabolic Engineering of Pseudomonas putida KT2440 to Produce Anthranilate from Glucose. Front. Microbiol. 2015, 6, 1310. [Google Scholar] [CrossRef]
- Zhang, J.; Qu, Q.; Chen, X.B. Assessing the sustainable safety practices based on human behavior factors: An application to Chinese petrochemical industry. Environ. Sci. Pollut. Res. Int. 2022, 29, 44618–44637. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, Y.; Zou, H. Transformation of pollution control and green development: Evidence from China’s chemical industry. J. Environ. Manag. 2020, 275, 111246. [Google Scholar] [CrossRef]
- Liu, W.; Yao, H.; Xu, W.; Liu, G.; Wang, X.; Tu, Y.; Shi, P.; Yu, N.; Li, A.; Wei, S. Suspect screening and risk assessment of pollutants in the wastewater from a chemical industry park in China. Environ. Pollut. 2020, 263, 114493. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, Z.; Cao, H.; Xu, M.; Xu, L. Concentrations, speciation, and ecological risk of heavy metals in the sediment of the Songhua River in an urban area with petrochemical industries. Chemosphere 2019, 219, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Relic, D.; Sakan, S.; Andelkovic, I.; Popovic, A.; Dordevic, D. Pollution and Health Risk Assessments of Potentially Toxic Elements in Soil and Sediment Samples in a Petrochemical Industry and Surrounding Area. Molecules 2019, 24, 2139. [Google Scholar] [CrossRef] [PubMed]
- Dantas, G.; Gorne, I.; da Silva, C.M.; Arbilla, G. Benzene, Toluene, Ethylbenzene and Xylene (BTEX) Concentrations in Urban Areas Impacted by Chemical and Petrochemical Industrial Emissions. Bull. Environ. Contam. Toxicol. 2022, 108, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, J.H. Recent Research Progress in the Microbial Production of Aromatic Compounds Derived from L-Tryptophan. J. Life Sci. 2020, 30, 919–929. [Google Scholar] [CrossRef]
- Noda, S.; Kondo, A. Recent Advances in Microbial Production of Aromatic Chemicals and Derivatives. Trends Biotechnol. 2017, 35, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Wang, Z.; Wang, B.; Hou, B.; Cheng, J.; Bai, T.; Zhang, Y.; Wang, W.; Yan, L.; Zhang, J. Expanding the application of tryptophan: Industrial biomanufacturing of tryptophan derivatives. Front. Microbiol. 2023, 14, 1099098. [Google Scholar] [CrossRef]
- Averesch, N.J.H.; Kromer, J.O. Metabolic Engineering of the Shikimate Pathway for Production of Aromatics and Derived Compounds-Present and Future Strain Construction Strategies. Front. Bioeng. Biotechnol. 2018, 6, 32. [Google Scholar] [CrossRef]
- Walter, T.; Al Medani, N.; Burgardt, A.; Cankar, K.; Ferrer, L.; Kerbs, A.; Lee, J.H.; Mindt, M.; Risse, J.M.; Wendisch, V.F. Fermentative N-Methylanthranilate Production by Engineered Corynebacterium glutamicum. Microorganisms 2020, 8, 866. [Google Scholar] [CrossRef]
- Upadhyay, P.; Lali, A. Engineered Pseudomonas putida for biosynthesis of catechol from lignin-derived model compounds and biomass hydrolysate. Prep. Biochem. Biotechnol. 2022, 52, 80–88. [Google Scholar] [CrossRef]
- Zhang, H.; Stephanopoulos, G. Co-culture engineering for microbial biosynthesis of 3-amino-benzoic acid in Escherichia coli. Biotechnol. J. 2016, 11, 981–987. [Google Scholar] [CrossRef]
- Averesch, N.J.; Winter, G.; Kromer, J.O. Production of para-aminobenzoic acid from different carbon-sources in engineered Saccharomyces cerevisiae. Microb. Cell Factories 2016, 15, 89. [Google Scholar] [CrossRef]
- Koma, D.; Yamanaka, H.; Moriyoshi, K.; Sakai, K.; Masuda, T.; Sato, Y.; Toida, K.; Ohmoto, T. Production of P-aminobenzoic acid by metabolically engineered Escherichia coli. Biosci. Biotechnol. Biochem. 2014, 78, 350–357. [Google Scholar] [CrossRef]
- Luo, Z.W.; Cho, J.S.; Lee, S.Y. Microbial production of methyl anthranilate, a grape flavor compound. Proc. Natl. Acad. Sci. USA 2019, 116, 10749–10756. [Google Scholar] [CrossRef]
- Mutz, M.; Bruning, V.; Brusseler, C.; Muller, M.F.; Noack, S.; Marienhagen, J. Metabolic engineering of Corynebacterium glutamicum for the production of anthranilate from glucose and xylose. Microb. Biotechnol. 2024, 17, e14388. [Google Scholar] [CrossRef]
- Li, Z.; Lu, Y.; Wang, X.; Vekaria, A.; Jiang, M.; Zhang, H. Enhancing anthranilic acid biosynthesis using biosensor-assisted cell selection and in situ product removal. Biochem. Eng. J. 2020, 162, 107722. [Google Scholar] [CrossRef]
- Kuivanen, J.; Kannisto, M.; Mojzita, D.; Rischer, H.; Toivari, M.; Jantti, J. Engineering of Saccharomyces cerevisiae for anthranilate and methyl anthranilate production. Microb. Cell Factories 2021, 20, 34. [Google Scholar] [CrossRef]
- Wu, S.; Chen, W.; Lu, S.; Zhang, H.; Yin, L. Metabolic Engineering of Shikimic Acid Biosynthesis Pathway for the Production of Shikimic Acid and Its Branched Products in Microorganisms: Advances and Prospects. Molecules 2022, 27, 4779. [Google Scholar] [CrossRef]
- Kubota, T.; Watanabe, A.; Suda, M.; Kogure, T.; Hiraga, K.; Inui, M. Production of para-aminobenzoate by genetically engineered Corynebacterium glutamicum and non-biological formation of an N-glucosyl byproduct. Metab. Eng. 2016, 38, 322–330. [Google Scholar] [CrossRef]
- Dong, L.H.; Liu, Y.J. Catalytic Mechanism of Pyridoxal 5′-Phosphate-Dependent Aminodeoxychorismate Lyase: A Computational QM/MM Study. J. Chem. Inf. Model. 2023, 63, 1313–1322. [Google Scholar] [CrossRef]
- Hou, F.; Xian, M.; Huang, W. De novo biosynthesis and whole-cell catalytic production of paracetamol on a gram scale in Escherichia coli. Green Chem. 2021, 23, 8280–8289. [Google Scholar] [CrossRef]
- Gorelova, V.; Bastien, O.; De Clerck, O.; Lespinats, S.; Rebeille, F.; Van Der Straeten, D. Evolution of folate biosynthesis and metabolism across algae and land plant lineages. Sci. Rep. 2019, 9, 5731. [Google Scholar] [CrossRef]
- Homayouni Rad, A.; Yari Khosroushahi, A.; Khalili, M.; Jafarzadeh, S. Folate bio-fortification of yoghurt and fermented milk: A review. Dairy Sci. Technol. 2016, 96, 427–441. [Google Scholar] [CrossRef]
- Lu, C.; Liu, Y.; Li, J.; Liu, L.; Du, G. Engineering of Biosynthesis Pathway and NADPH Supply for Improved L-5-Methyltetrahydrofolate Production by Lactococcus lactis. J. Microbiol. Biotechnol. 2019, 31, 154–162. [Google Scholar] [CrossRef]
- Inglis, J.J.; Criado, G.; Andrews, M.; Feldmann, M.; Williams, R.O.; Selley, M.L. The anti-allergic drug, N-(3′,4′-dimethoxycinnamonyl) anthranilic acid, exhibits potent anti-inflammatory and analgesic properties in arthritis. Rheumatology 2007, 46, 1428–1432. [Google Scholar] [CrossRef]
- Li, X.H.; Kim, S.K.; Lee, J.H. Anti-biofilm effects of anthranilate on a broad range of bacteria. Sci. Rep. 2017, 7, 8604. [Google Scholar] [CrossRef]
- Muszynska, B.; Lojewski, M.; Rojowski, J.; Opoka, W.; Sulkowska-Ziaja, K. Natural products of relevance in the prevention and supportive treatment of depression. Psychiatr. Pol. 2015, 49, 435–453. [Google Scholar] [CrossRef]
- Prasher, P.; Sharma, M. Medicinal chemistry of anthranilic acid derivatives: A mini review. Drug Dev. Res. 2021, 82, 945–958. [Google Scholar] [CrossRef]
- Shaw, C.; Hess, M.; Weimer, B.C. Microbial-Derived Tryptophan Metabolites and Their Role in Neurological Disease: Anthranilic Acid and Anthranilic Acid Derivatives. Microorganisms 2023, 11, 1825. [Google Scholar] [CrossRef]
- Kim, H.J.; Seo, S.Y.; Park, H.S.; Ko, J.Y.; Choi, S.S.; Lee, S.J.; Kim, E.S. Engineered Escherichia coli cell factory for anthranilate over-production. Front. Microbiol. 2023, 14, 1081221. [Google Scholar] [CrossRef]
- Walsh, C.T.; Haynes, S.W.; Ames, B.D.; Gao, X.; Tang, Y. Short pathways to complexity generation: Fungal peptidyl alkaloid multicyclic scaffolds from anthranilate building blocks. ACS Chem. Biol. 2013, 8, 1366–1382. [Google Scholar] [CrossRef] [PubMed]
- Balderas-Hernandez, V.E.; Sabido-Ramos, A.; Silva, P.; Cabrera-Valladares, N.; Hernandez-Chavez, G.; Baez-Viveros, J.L.; Martinez, A.; Bolivar, F.; Gosset, G. Metabolic engineering for improving anthranilate synthesis from glucose in Escherichia coli. Microb. Cell Factories 2009, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Balderas-Hernández, V.E.; Treviño-Quintanilla, L.G.; Hernández-Chávez, G.; Martinez, A.; Bolívar, F.; Gosset, G. Catechol biosynthesis from glucose in Escherichia coli anthranilate-overproducer strains by heterologous expression of anthranilate 1,2-dioxygenase from Pseudomonas aeruginosa PAO1. Microb. Cell Factories 2014, 13, 136. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Chen, X.; Wang, J.; Sun, X.; Dong, S.; Li, Y.; Yan, Y.; Wang, J.; Yuan, Q. Design and construction of an artificial pathway for biosynthesis of acetaminophen in Escherichia coli. Metab. Eng. 2021, 68, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, Y.; Li, J.; Liu, L.; Du, G.; Chen, J. Systems metabolic engineering of Bacillus subtilis for efficient biosynthesis of 5-methyltetrahydrofolate. Biotechnol. Bioeng. 2020, 117, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Schillert, L.; Wirtz, D.; Weber, N.; Schaller, F.; Striegel, L.; Schmitt-Kopplin, P.; Rychlik, M. Metabolic folate profiling as a function of time during cultivation suggests potential C2-metabolism in Saccharomyces cerevisiae. Front. Nutr. 2022, 9, 984094. [Google Scholar] [CrossRef] [PubMed]
- Kuepper, J.; Otto, M.; Dickler, J.; Behnken, S.; Magnus, J.; Jager, G.; Blank, L.M.; Wierckx, N. Adaptive laboratory evolution of Pseudomonas putida and Corynebacterium glutamicum to enhance anthranilate tolerance. Microbiology 2020, 166, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; De Luca, V. The biosynthesis and regulation of biosynthesis of Concord grape fruit esters, including ‘foxy’ methylanthranilate. Plant J. 2005, 44, 606–619. [Google Scholar] [CrossRef]
- Kollner, T.G.; Lenk, C.; Zhao, N.; Seidl-Adams, I.; Gershenzon, J.; Chen, F.; Degenhardt, J. Herbivore-induced SABATH methyltransferases of maize that methylate anthranilic acid using s-adenosyl-L-methionine. Plant Physiol. 2010, 153, 1795–1807. [Google Scholar] [CrossRef]
- Song, G.; Wu, F.; Peng, Y.; Jiang, X.; Wang, Q. High-Level Production of Catechol from Glucose by Engineered Escherichia coli. Fermentation 2022, 8, 344. [Google Scholar] [CrossRef]
- Pugh, S.; McKenna, R.; Osman, M.; Thompson, B.; Nielsen, D.R. Rational engineering of a novel pathway for producing the aromatic compounds p-hydroxybenzoate, protocatechuate, and catechol in Escherichia coli. Process Biochem. 2014, 49, 1843–1850. [Google Scholar] [CrossRef]
- Wang, S.; Ma, Y.; Wang, Y.; Jiao, M.; Luo, X.; Cui, M. One-step electrodeposition of poly(m-aminobenzoic acid) membrane decorated with peptide for antifouling biosensing of Immunoglobulin E. Colloids Surf. B Biointerfaces 2020, 186, 110706. [Google Scholar] [CrossRef] [PubMed]
- Jokić, B.M.; Džunuzović, E.S.; Grgur, B.N.; Jugović, B.Z.; Trišovic, T.L.; Stevanović, J.S.; Gvozdenović, M.M. The influence of m-aminobenzoic acid on electrochemical synthesis and behavior of poly(aniline-co-(m-aminobenzoic acid). J. Polym. Res. 2017, 24, 146. [Google Scholar] [CrossRef]
- Hirayama, A.; Eguchi, T.; Kudo, F. A single PLP-dependent enzyme PctV catalyzes the transformation of 3-dehydroshikimate into 3-aminobenzoate in the biosynthesis of pactamycin. Chembiochem 2013, 14, 1198–1203. [Google Scholar] [CrossRef]
- Hirayama, A.; Miyanaga, A.; Kudo, F.; Eguchi, T. Mechanism-Based Trapping of the Quinonoid Intermediate by Using the K276R Mutant of PLP-Dependent 3-Aminobenzoate Synthase PctV in the Biosynthesis of Pactamycin. Chembiochem 2015, 16, 2484–2490. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, L.; Deng, Z. Functional characterization of the first two actinomycete 4-amino-4-deoxychorismate lyase genes. Microbiology 2009, 155, 2450–2459. [Google Scholar] [CrossRef]
- Lonshakov, D.V.; Sheremet’ev, S.V.; Belosludtseva, E.M.; Korovkin, S.A.; Semchenko, A.V.; Katlinskij, A.V. Synthesis of 4-aminobenzoic acid esters of polyethylene glycol and their use for pegylation of therapeutic proteins. RSC Adv. 2015, 5, 42903–42909. [Google Scholar] [CrossRef]
- Zaib Un, N.; Akhtar, T. para-Aminobenzoic Acid-A Substrate of Immense Significance. Mini-Rev. Org. Chem. 2020, 17, 686–700. [Google Scholar] [CrossRef]
- Michael, R.; Richards, E.; Xing, D.K.L. The effect of p-aminobenzoic acid on the uptake of thymidine and uracil by Escherichia coli. Int. J. Pharm. 1995, 116, 217–221. [Google Scholar] [CrossRef]
- Laborda, P.; Zhao, Y.; Ling, J.; Hou, R.; Liu, F. Production of Antifungal p-Aminobenzoic Acid in Lysobacter antibioticus OH13. J. Agric. Food Chem. 2018, 66, 630–636. [Google Scholar] [CrossRef]
- Kołton, A.; Długosz-Grochowska, O.; Wojciechowska, R.; Czaja, M. Biosynthesis Regulation of Folates and Phenols in Plants. Sci. Hortic. 2022, 291, 110561. [Google Scholar] [CrossRef]
- Saubade, F.; Hemery, Y.M.; Guyot, J.P.; Humblot, C. Lactic acid fermentation as a tool for increasing the folate content of foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3894–3910. [Google Scholar] [CrossRef]
- Khiralla, G.; El-Tarras, A.; Elhariry, H. Dihydrofolate Reductase Thermosensitive (ts-dfrA) Mutant Induces Dihydrofolate Overproduction by Lactobacillus plantarum. Food Biotechnol. 2019, 33, 17–33. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, X.; Liu, Y.; Liu, L.; Li, J.; Du, G.; Chen, J. Synthetic biology-driven microbial production of folates: Advances and perspectives. Bioresour. Technol. 2021, 324, 124624. [Google Scholar] [CrossRef]
- Sybesma, W.; Starrenburg, M.; Tijsseling, L.; Hoefnagel, M.H.N.; Hugenholtz, J. Effects of Cultivation Conditions on Folate Production by Lactic Acid Bacteria. Appl. Environ. Microbiol. 2003, 69, 4542–4548. [Google Scholar] [CrossRef]
- Patring, J.D.M.; Hjortmo, S.B.; Jastrebova, J.A.; Svensson, U.K.; Andlid, T.A.; Jägerstad, I.M. Characterization and quantification of folates produced by yeast strains isolated from kefir granules. Eur. Food Res. Technol. 2006, 223, 633–637. [Google Scholar] [CrossRef]
- Serrano-Amatriain, C.; Ledesma-Amaro, R.; Lopez-Nicolas, R.; Ros, G.; Jimenez, A.; Revuelta, J.L. Folic Acid Production by Engineered Ashbya gossypii. Metab. Eng. 2016, 38, 473–482. [Google Scholar] [CrossRef]
- Blancquaert, D.; Storozhenko, S.; Loizeau, K.; De Steur, H.; De Brouwer, V.; Viaene, J.; Ravanel, S.; Rébeillé, F.; Lambert, W.; Van Der Straeten, D. Folates and Folic Acid: From Fundamental Research Toward Sustainable Health. Crit. Rev. Plant Sci. 2010, 29, 14–35. [Google Scholar] [CrossRef]
- Raffa, R.B.; Pergolizzi, J.V., Jr.; Taylor, R., Jr.; Decker, J.F.; Patrick, J.T. Acetaminophen (paracetamol) oral absorption and clinical influences. Pain Pract. 2014, 14, 668–677. [Google Scholar] [CrossRef]
- Jóźwiak-Bebenista, M.; Nowak, J.Z. Paracetamol: Mechanism of action, applications and safety concern. Acta Pol. Pharm. 2014, 71, 11–23. [Google Scholar]
- Bertolini, A.; Ferrari, A.; Ottani, A.; Guerzoni, S.; Tacchi, R.; Leone, S. Paracetamol: New vistas of an old drug. CNS Drug Rev. 2006, 12, 250–275. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.J.; Lee, E.W.; Bang, W.G.; Lee, C.K.; Kim, K.H.; Choi, I.G. Molecular characterization of a novel bacterial aryl acylamidase belonging to the amidase signature enzyme family. Mol. Cells 2010, 29, 485–492. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Product | Host | Carbon Source | Titer (g/L) | Time | Fermentation Mode | Engineered Strategy | Advantages | Disadvantages | Reference |
|---|---|---|---|---|---|---|---|---|---|
| OABA | C. glutamicum | Glucose and xylose | 5.9 | 32 h | Pulsed-fed-batch | 3-Deoxyarabinoheptulosonate-7 phase syntax and TrpEG were imported, and genes nagD and qsuD were removed | Engineering C. glutamicum with strong tolerance | Expensive and complex culture media | [25] |
| E. coli | Glucose | 4.0 | 60 h | Fed-batch | Destruction of pheA, tyrA, pabA, ubiC, entC, and trpR genes, overexpression of aroE and tktA genes | Higher yield | Complex strain transformation steps | [40] | |
| S. cerevisiae | D-glucose | 0.57 | 72 h | Batch | Disruption of TRP4, overexpression of GLN1, regulation of Aro4 and Trp2, and overexpression of TRP3 | Realize the production of OABA by S. cerevisiae | Low carbon flux | [27] | |
| MANT | S. cerevisiae | D-glucose | 0.41 | 72 h | Batch | Expression of anthranilic acid methyltransferase 1 from Medicago truncatula | Realize the production of MANT by S. cerevisiae | Low carbon flux | [27] |
| C. glutamicum | Glucose | 5.74 | 110 h | Fed-batch | Expression of pSH36HTc and pEKGH in C. glutamicum | The reaction is simpler and the cofactors are less. | High residual levels of precursor metabolites | [24] | |
| E. coli | Glucose | 4.47 | 72 h | Fed-batch | Expression of pBBR1GfbrAfbrEfbr and pTacT in E. coli | The reaction is simpler and the cofactors are less. | Low enzyme catalytic activity | [24] | |
| Catechol | E. coli | Glucose | 4.47 | 76 h | Fed-batch | Expression of 1,2-dioxygenase, DAHP synthase and transketolase. in E. coli | Low cultivation cost | Lower glucose consumption capacity of the strain | [43] |
| MABA | E. coli | Glucose | 0.048 | 6 d | Batch | Coupling of 3AB synthase PctV with engineered shikimate pathway | The modular nature of co culture engineering allows for rapid identification of specific enzymes or optimal strains | Low activity of MABA synthase PctV | [21] |
| PABA | E. coli | Glycerol | 0.84 | 48 h | Batch | Strengthening the shikimate pathway and overexpressing PABA synthase | Few by-products | The enzyme promiscuity is not beneficial to the production | [44] |
| S. cerevisiae | Glycerol-ethanol | 0.22 | 78 h | Fed-batch | Overexpression of ABZ1 and ABZ2 genes in wine yeast AWRI1631 and QA23 | Low process cost | High glycerol will have a negative impact on PABA titer | [22] | |
| C. glutamicum | Glucose | 43 | 48 h | Test-tube scale culture | Introducing pabAB from C. callunae and pabC from X. bovienii into strains overexpressing the shikimate pathway | High PABA production titer | folP deficiency can lead to malnutrition in bacterial strains | [29] | |
| Folate | Bacillus subtilis | Glucose | 0.003 | 15 h | Fed-batch | Replacing yitJ with metF, knocking out purU, overexpressing dfrA, folC, pab, folE, and yciA, inhibiting thyA, pheA, trpE, and panB genes | Production cost reduction | Low yield | [45] |
| S. cerevisiae | Glucose | 0.13 | 20 h | Batch | SIDA (Stable Isotope Dilution Analysis) determination and Molecular Network (MN) Analysis | Increasing of yield | C2-metabolic mechanism unclear | [46] | |
| Acetaminophen | E. coli | Glycerol | 0.12 | 48 h | Batch | Expression of p-AP N-acetyltransferase | Implementation of producing AAP with a simple carbon source | High by-products | [44] |
| E. coli | Glucose | 0.94 | 24 h | Fed-batch | Heterologous expression of enzymes from five different microbial sources, modification of ABH and PANAT enzymes | The increase in AAP yield, no need for auxiliary factors | Medium with complex composition | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, S.; Zeng, R.; Wang, B.; Zhang, S.; Cheng, J.; Zhang, J. A New Direction for the Green, Environmentally Friendly and Sustainable Bioproduction of Aminobenzoic Acid and Its Derivatives. Sustainability 2024, 16, 3052. https://doi.org/10.3390/su16073052
Xiao S, Zeng R, Wang B, Zhang S, Cheng J, Zhang J. A New Direction for the Green, Environmentally Friendly and Sustainable Bioproduction of Aminobenzoic Acid and Its Derivatives. Sustainability. 2024; 16(7):3052. https://doi.org/10.3390/su16073052
Chicago/Turabian StyleXiao, Shujian, Rumei Zeng, Bangxu Wang, Suyi Zhang, Jie Cheng, and Jiamin Zhang. 2024. "A New Direction for the Green, Environmentally Friendly and Sustainable Bioproduction of Aminobenzoic Acid and Its Derivatives" Sustainability 16, no. 7: 3052. https://doi.org/10.3390/su16073052