
An Economic Cost/Benefit Tool to Assess Bee Pollinator Conservation, Pollination Strategies, and Sustainable Policies: A Lowbush Blueberry Case Study



Abstract
1. Introduction
1.1. Bee Pollinator Decline—A Significant Concern
1.2. Economics of Pollination—Not Thoroughly Studied
1.3. Sustainable Pollinator or Pollination Protection Policies—A Global Perspective
1.4. Lowbush Blueberry—A Unique Native North American Wild Crop
1.5. Goals and Objectives for Lowbush Blueberry Sustainable Pollination
- (1)
- Determine that both native bees and managed honey bees contribute, in part, to fruit set;
- (2)
- Determine the relationship between honey bee hive stocking density and honey bee forager populations in the field;
- (3)
- Estimate the relationship between fruit set and yield;
- (4)
- Propose, formulate, and use an “Economic Pollinator Level” (EPL) metric for evaluating future conservation and pollination policies and strategies using case study examples;
- (5)
- Conduct Monte Carlo simulations on the economic uncertainty or risk in lowbush blueberry pollination by way of using the EPL to test pollination tactics.
2. Materials and Methods
2.1. Lowbush Blueberry Sites Sampled
2.2. Statistical, Theoretical, and Monte Carlo Analyses
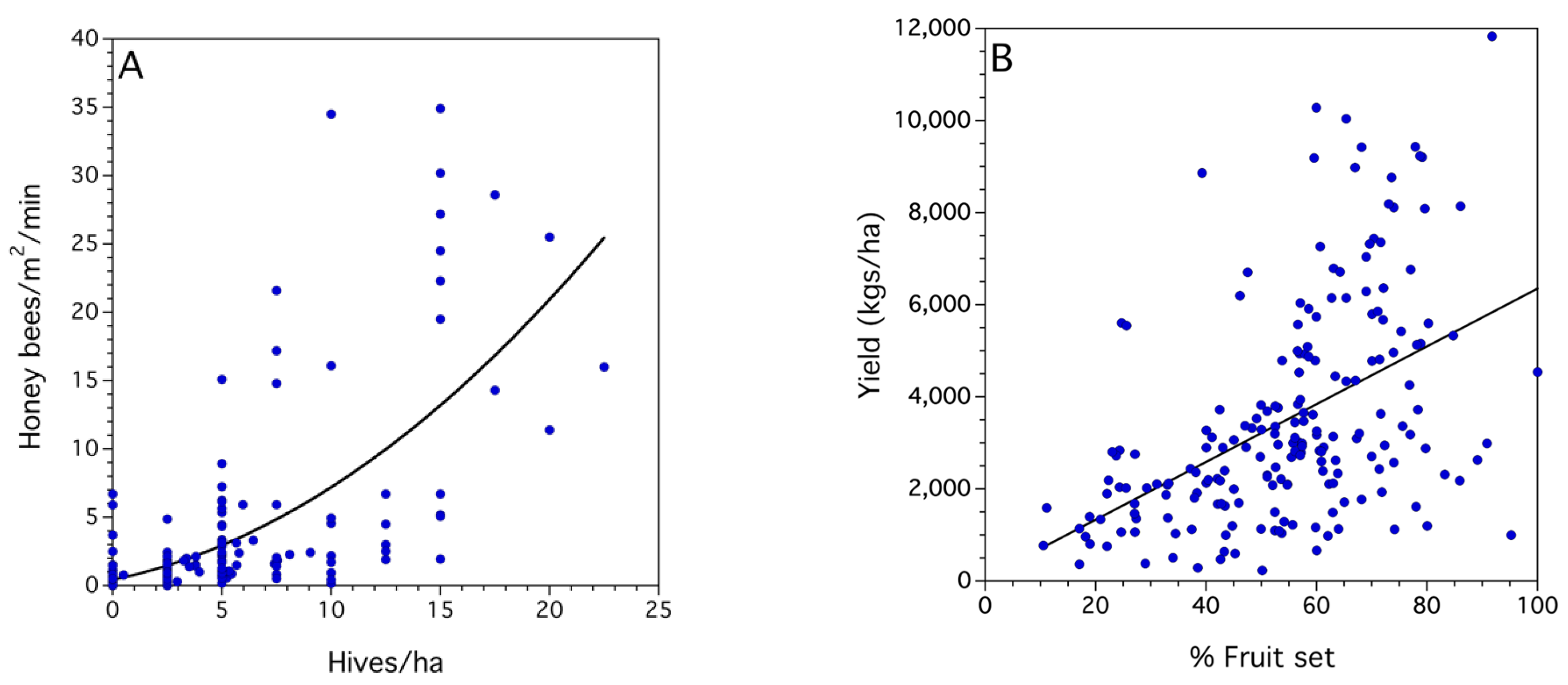
2.2.1. Modeling Relationships between Bee Activity Density, Hive Stocking Density, Proportion Fruit Set, and Yield
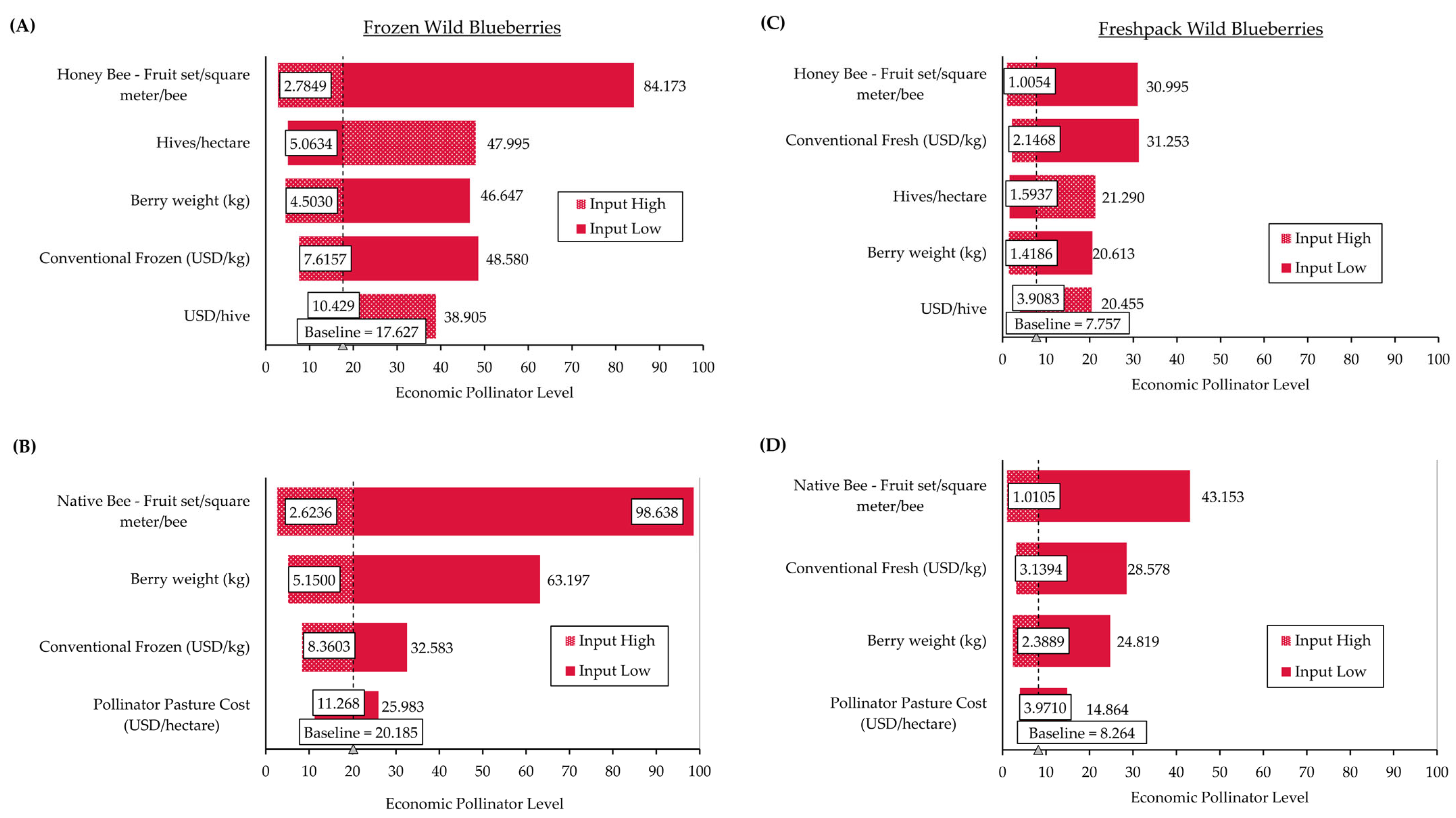
2.2.2. Development of an Economic Pollinator Level
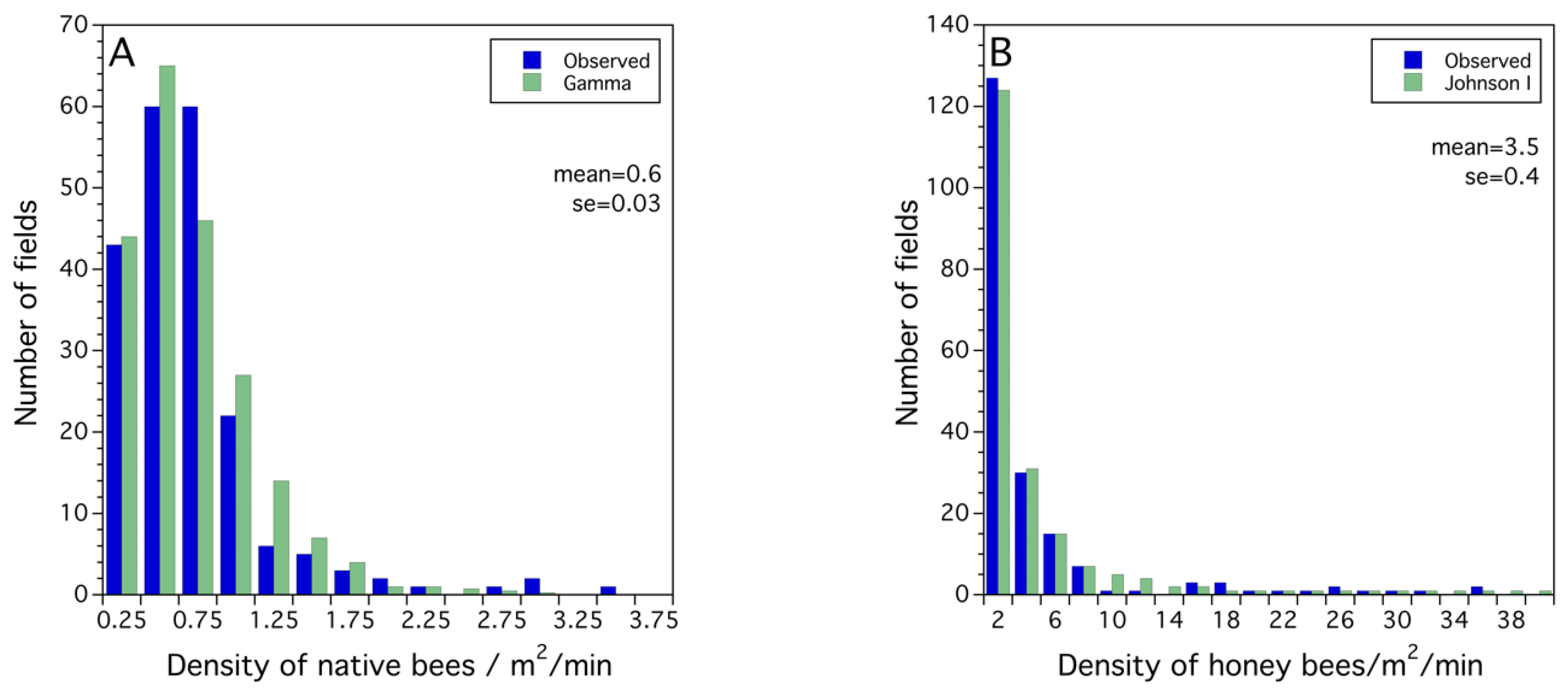
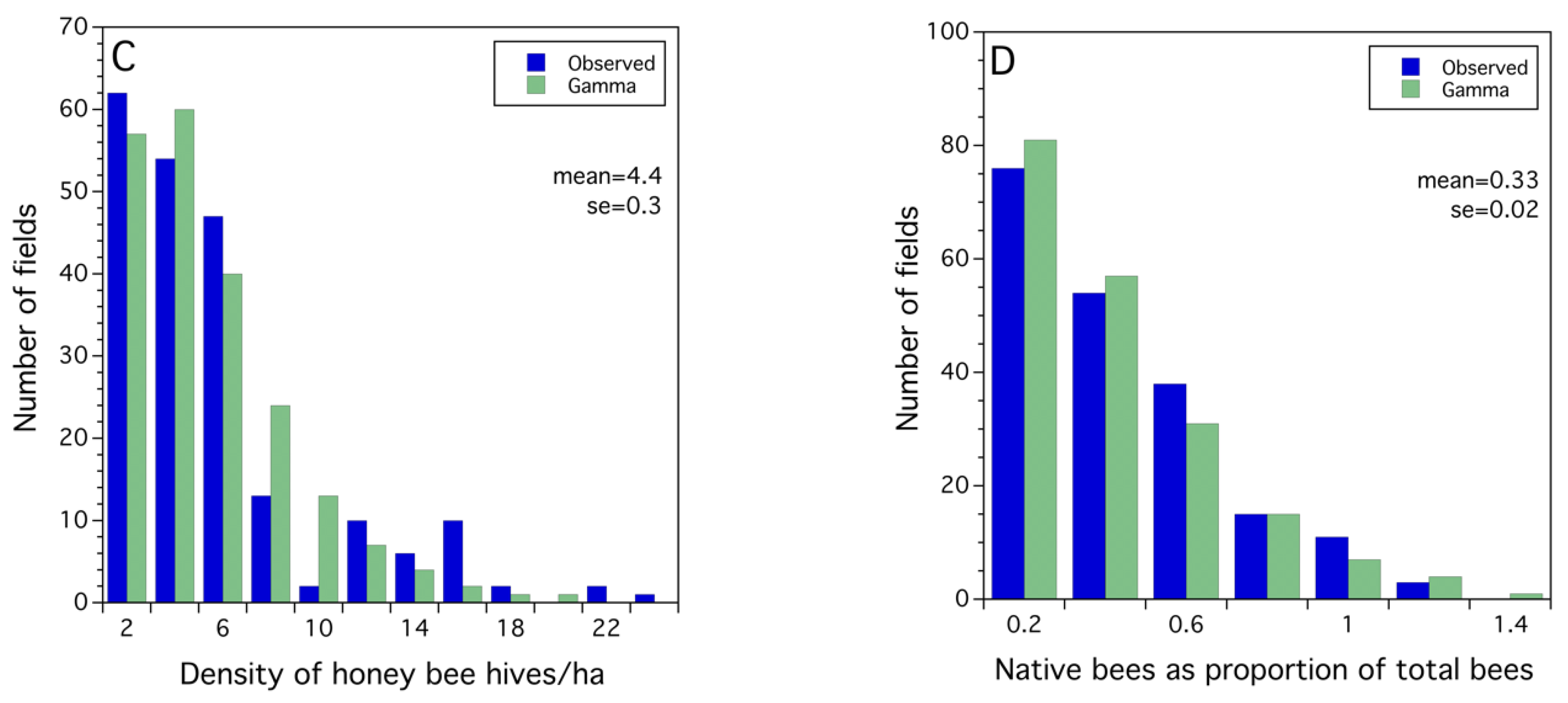
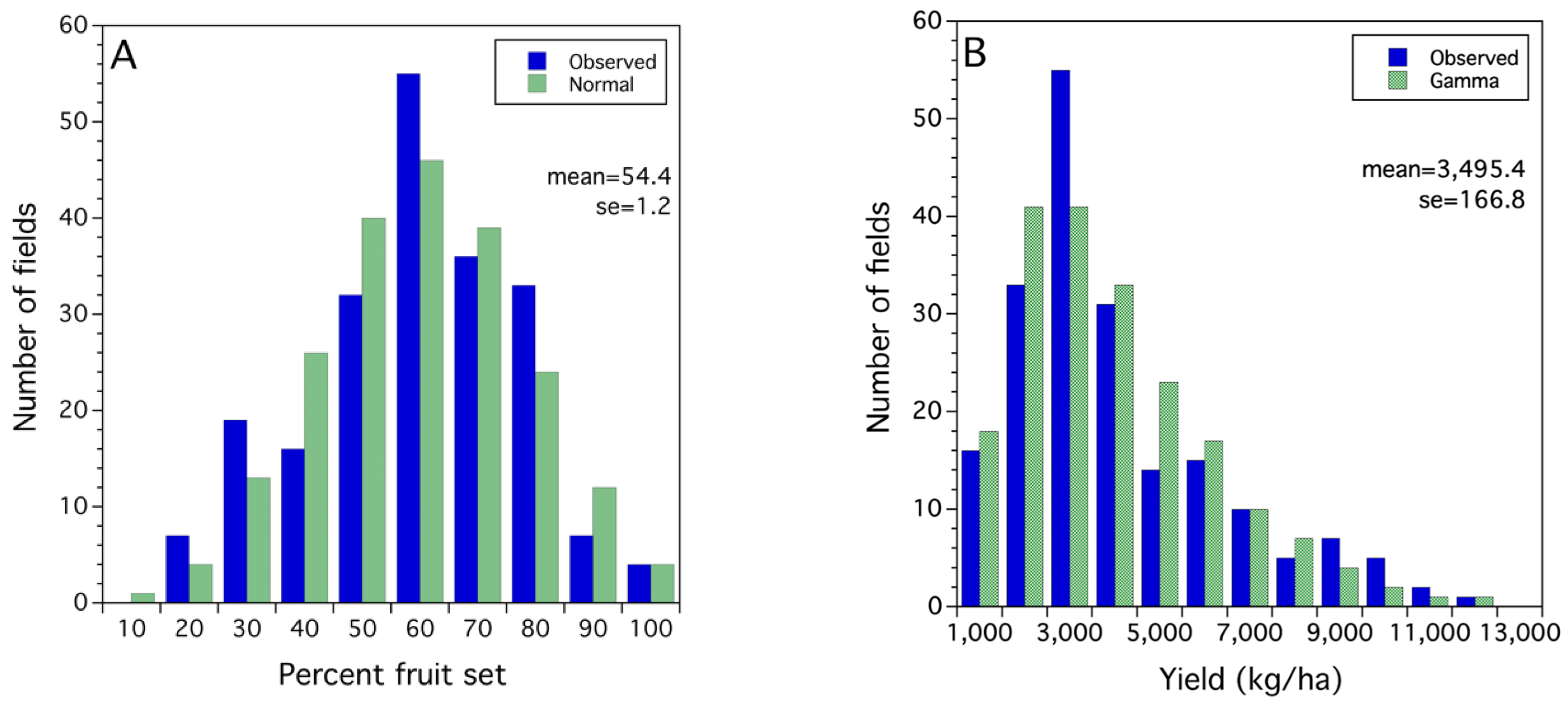
2.2.3. Probability Modeling—Fitting Probability Density Functions to Observed Frequency Distributions
2.2.4. Monte Carlo Simulation of Economic Uncertainty or Risk of Lowbush Blueberry Pollination and Yield

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Economic Pollinator Level Variables | Pollination System | Years | Data Source | Producer Price Index Adjust for Inflation [73] | Function Form for Probability Distribution Fit |
|---|---|---|---|---|---|
| (1) Monetary cost of pollination (C) | Native Bee Pasture | 2002–2023 | Various (Table A2) | Various (Table A2) | Various (Table A2) |
| Rented Honey Bees | 2003–2022 | Cooperating producer [26] | Lumber and plywood | Uniform | |
| (2) Value of wild blueberries (V) | |||||
| Native Bee Pasture and Rented Honey Bees | 2002–2021 | USDA, NASS [74] | Frozen fruits/vegis | Extreme value |
| Same as above | Same | Same | Fresh blueberries | Laplace |
| (3) Number of flowers set per m2 per bee (P) | Native Bee Pasture and Rented Honey Bees | 2005, 2013, and 2015 | [26,59,61] | Not used | Gamma Gamma |
| (4) Lowbush blueberry single berry weight (B) | Native Bee Pasture and Rented Honey Bees | 2006, 2007, and 2011 | [59,61,71] | Not used | Weibull |
3. Results
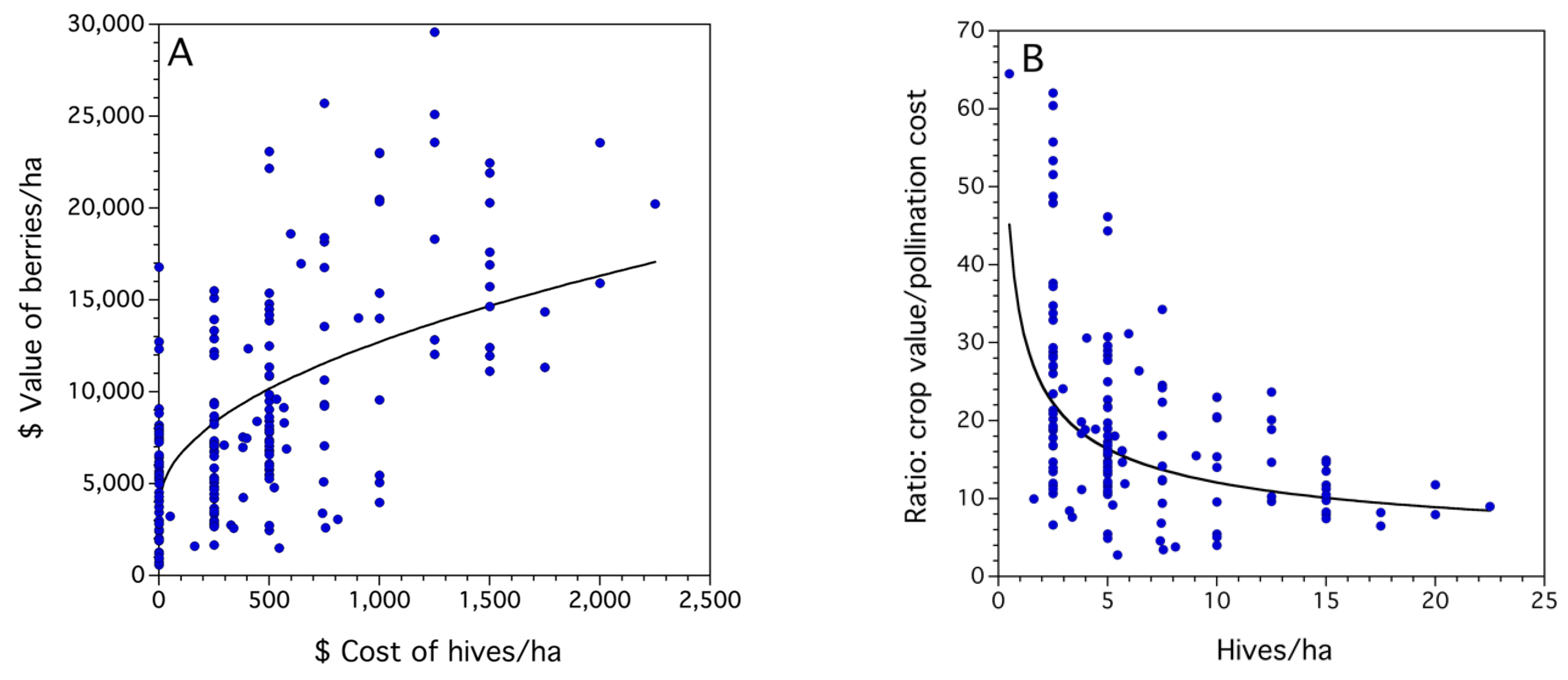
3.1. Frequency Distributions of Sampled Bee and Crop Pollination and Production Metrics
3.2. Statistical Models for Bee, Pollination and Crop Production Metrics
3.3. Case Studies Demonstrating Use of Economic Pollinator Level Model
3.3.1. Case Study 1: Native Bee Economic Pollinator Level—A Tool for Economic and Ecological Sustainability
- C =
- Annual cost of a pollinator planting = USD 17,999/hectare (ha) (see Appendix A, Table A1), therefore cost/m2 = USD 17,999/ha divided by 10,000 m2/ha = USD 1.80/m2.
- V =
- Value of fruit/kg, USD 1.50/kg (2018–2021 average inflation adjusted price or real price for frozen lowbush blueberries in Maine) versus an average real price of USD 3.10/kg for fresh lowbush blueberries over the same time period [72,74]. However, conventional road side prices for fresh fruit can be USD 5–6/kg, and fresh organic fruit is USD 10–14/kg [61].
- P =
- Flowers set per native bee/m2 = 22.9 flowers/stem × 738.4 stems/m2 = 16,909.4 flowers/m2 [61] and 10% fruit set/native bee/m2 [61], therefore 0.1 × 16,909.4 = 1690.9 fruits assuming a dropped proportion of 0.364 (median) are dropped [71], then 1690.9 × (1 − 0.364) = 1075.4 flowers/m2 set by a bee that become marketable fruit. However, lowbush blueberry plants and stems within plants do not initiate floral opening of all the flowers at the start of the bloom period [71]. Flowers open in an asymptotic sigmoidal pattern up until peak bloom [69], after which the number of flowers decreases due to flower stigma viability and successful pollination [70]. Flowers are usually only viable for 5–7 days [71]. Therefore, the distributed bloom over the bloom period and a floral viability of 6.5 days after opening over a typical 26-day bloom period means that bees do not have the total available number of flowers per m2 at any one point in time. The number of flowers that can be set by a single bee per day over the 26-day bloom period is a linear approximation derived by the average number of flowers available for visitation by the bee per day. Based upon these bloom dynamics, the number of flowers per m2 available for a bee to transform into marketable fruit after fruit drop is accounted for (see above) and is the number of flowers/m2 divided by 6.5. This is equivalent to the integral of a Gaussian bloom distribution over time (days) resulting in the total flower × days divided by the 6.5-day residence time of a viable flower. This results in a daily flower availability to foraging bees. This factor determines the daily flowers available for visitation and thus, pollination per day, given the distributed bloom and a viability duration of 6.5 days once a flower is open. Therefore, in this case study, 1075.4 flowers/m2/6.5 = 165.5 flowers/m2.
- B =
- Weight of one marketable berry is 0.0004 kg.
- The equation becomes:This equation could be used to economically justify planting a pollinator planting from a bee activity density perspective, as a before and after comparison.EPL = 18.13 native bees/m2
3.3.2. Case Study 2: Honey Bee Economic Pollinator Level—A Tool for Economic Sustainability
- C =
- Annual cost of deploying 5 hives/hectare (ha) × USD 150/hive (inflation adjusted 2003 to 2022 average price) = USD 750/ha therefore cost/m2 = USD 750/ha divided by 10,000 m2/ha = USD 0.075/m2.
- V =
- P =
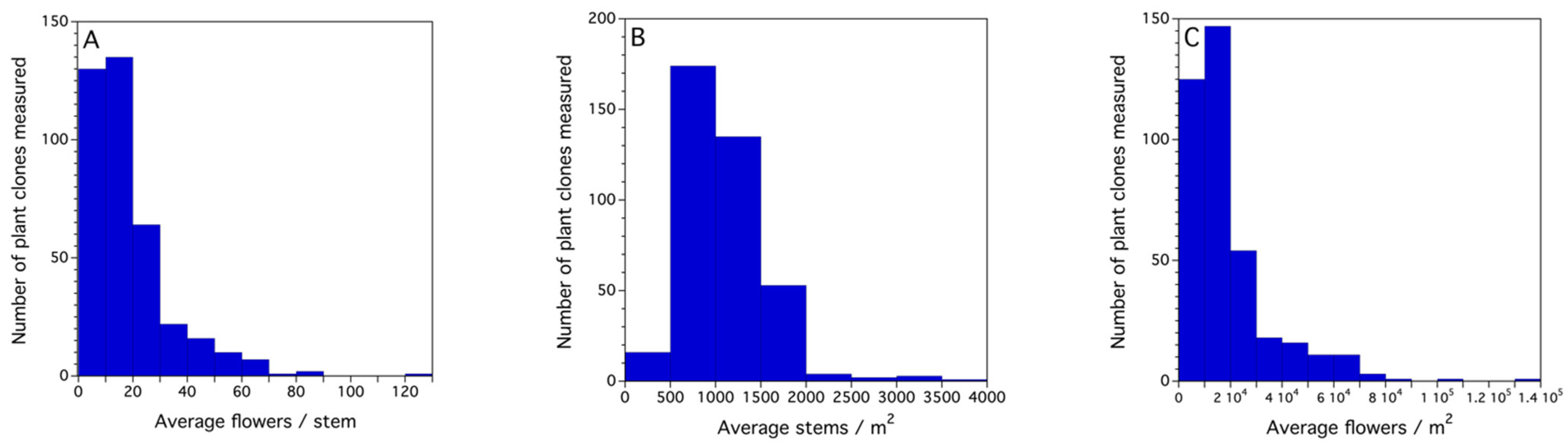
- Flowers set per native bee/m2 = as in case study 1, 22.9 flowers/stem × 738.4 stems/m2 = 16,909.4 flowers /m2. However, for honey bees, only a 1% fruit set/ bee/m2 is expected [62], therefore 0.01 × 16,909.4 = 169.1 fruits, assuming a proportion of 0.364 (median) are dropped as in case study 1, thus 169.1 × (1 − 0.364) = 107.5 flowers/m2. This number of flowers/m2 is divided by 6.5. This factor determines the daily flowers available for visitation and thus, pollination per day, given the distributed bloom and the viability duration of 5 days once a flower is open. Therefore, in this case study, 107.5 flowers/m2/6.5 = 16.5 flowers/m2 that will end up as marketable fruit at the end of the season. The observed frequency distributions of flowers/stem (Figure A1A), stems/m2 (Figure A1B), and the derived estimate of flowers/m2 (Figure A1C) are in Appendix A.
- B =
- Weight of one marketable berry is 0.0004 kg.
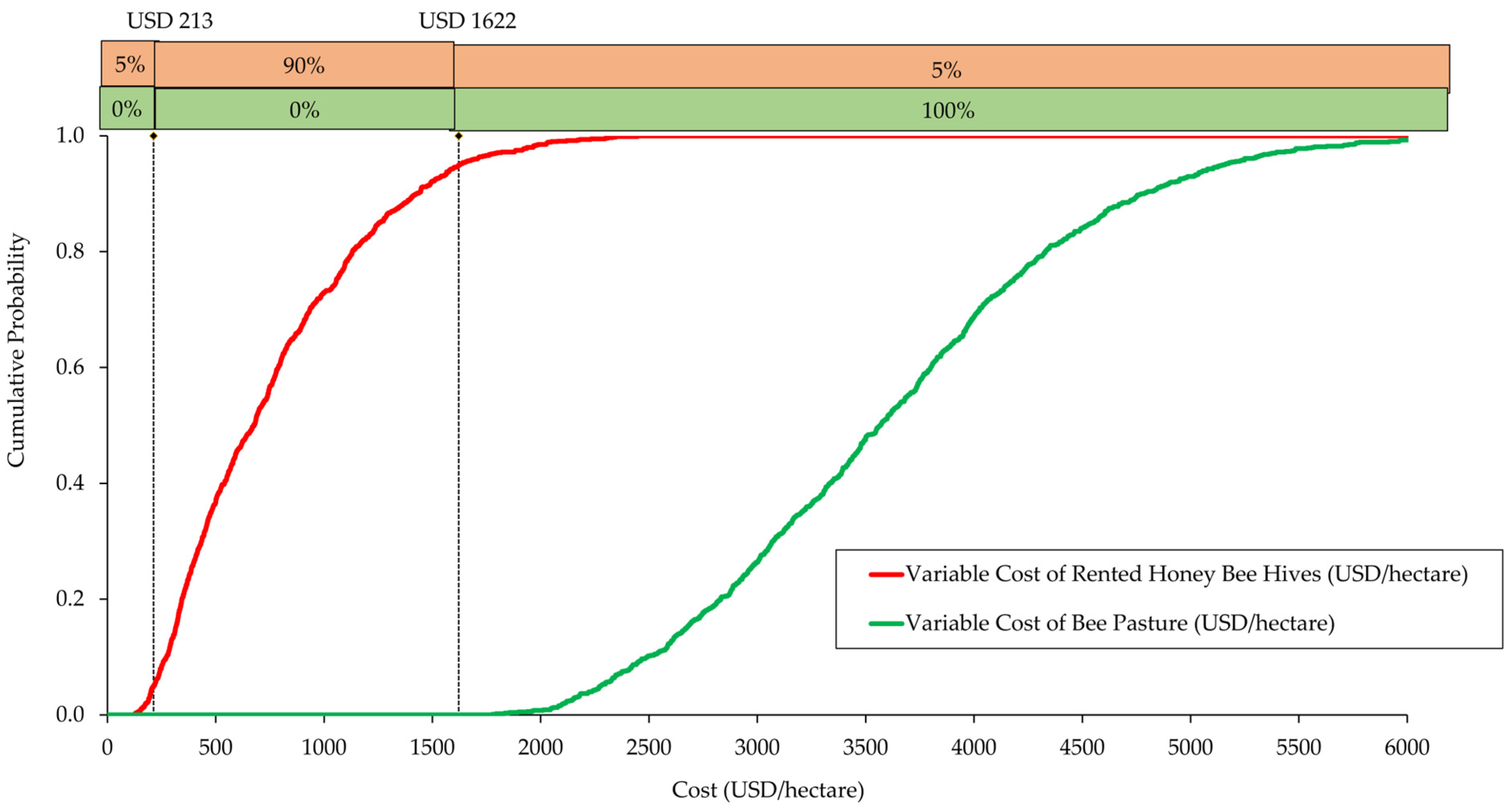
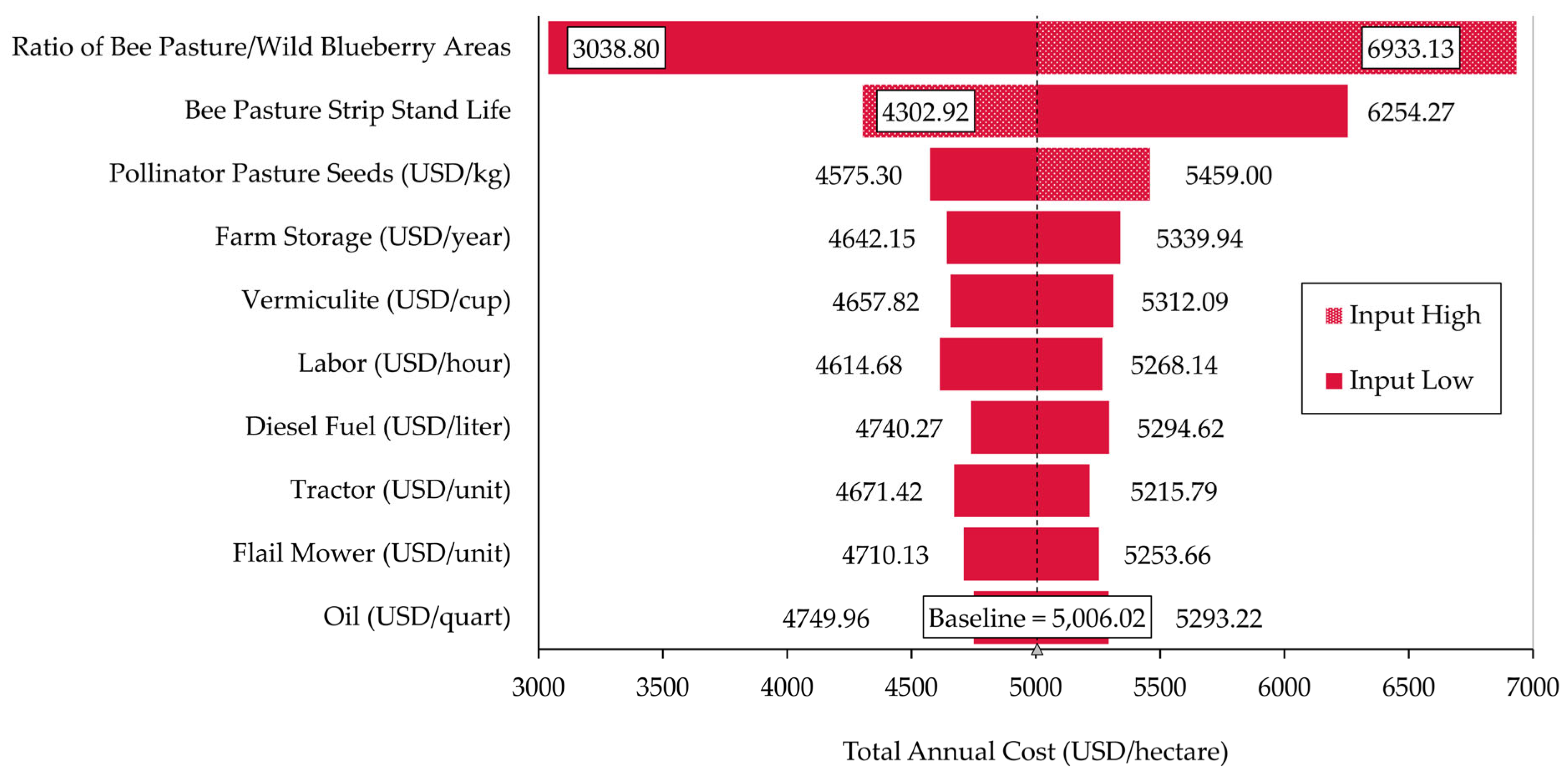
3.4. Monte Carlo Economic Simulations
4. Discussion
4.1. Interplay between Honey Bees and Native Bees in Lowbush Blueberry
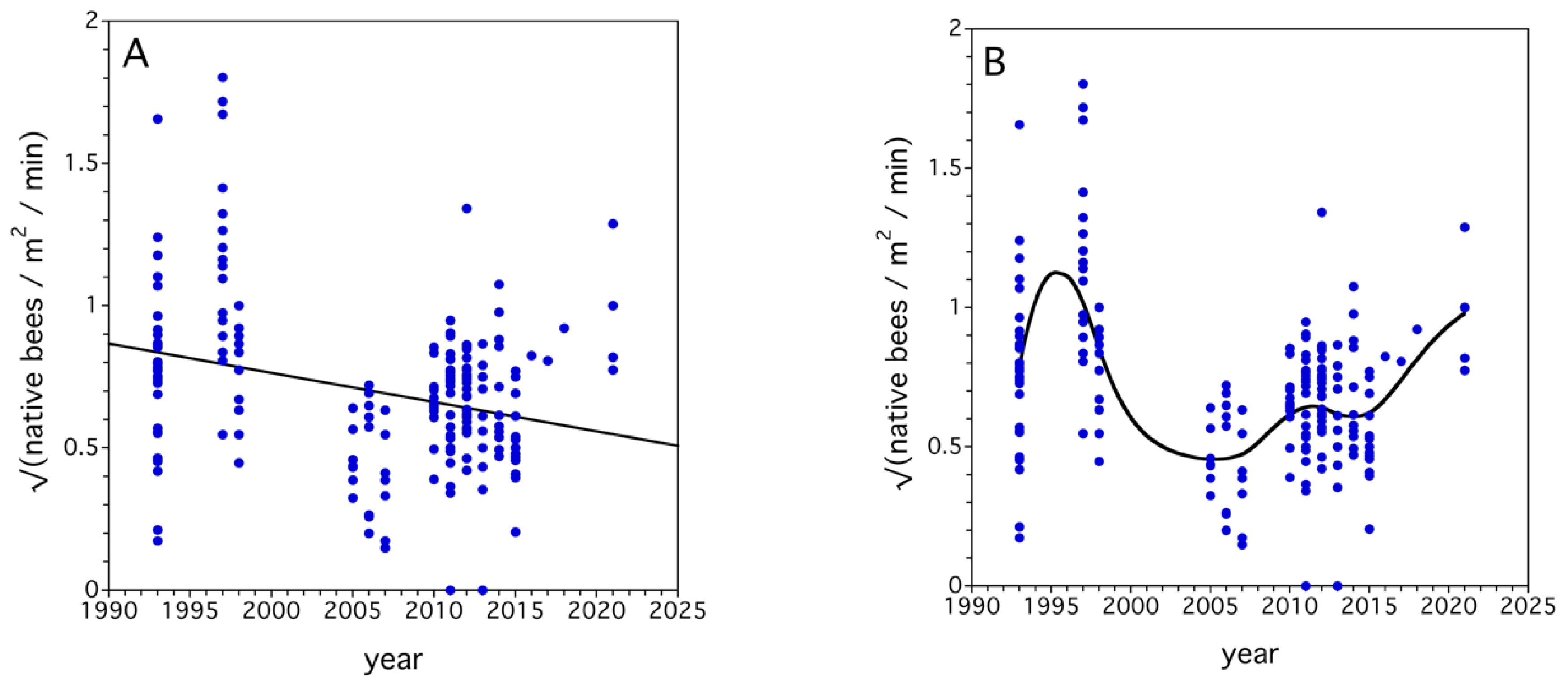
4.2. Long-Term Trends in Native Bee Communities
4.3. Conservation of Honey Bees and Native Bees
4.4. Tools for Pollination Strategies
4.5. Using Predictive Pollination Tools to Guide Pollinator Public Policy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Annual Pollination Pasture Budget | Scale: | 0.067 | Hectares | 5.953 | Year Stand Life |
| Characteristic/Costs | Total | Per Hectare | Per Square Meter | ||
| Number of Acres of Pasture | 0.164 | - | - | ||
| Number of Hectares of Pasture | 0.067 | - | - | ||
| Annual Operating Expenses (USD) | |||||
| Lime | 0 | 0 | 0 | ||
| Seedlings | 0 | 0 | 0 | ||
| Seeds | 77 | 1160.28 | 0.116 | ||
| Seed Amendments | 12 | 184.49 | 0.018 | ||
| Labor | |||||
| Establishment Year 1 | 50 | 745.30 | 0.075 | ||
| Ensure Establishment Years 1 to 3 | 8 | 114.74 | 0.011 | ||
| Annual | 41 | 614.72 | 0.061 | ||
| Diesel Fuel | |||||
| Establishment Year 1 | 10 | 147.52 | 0.015 | ||
| Ensure Establishment Years 1 to 3 | 0.40 | 6.06 | 0.001 | ||
| Annual | 0 | 0 | 0 | ||
| Oil | 3 | 38.10 | 0.004 | ||
| Maintenance and Upkeep | 111 | 1673.65 | 0.167 | ||
| Utilities | 0 | 0 | 0 | ||
| Total Operating Expenses | 312 | 4684.86 | 0.468 | ||
| Annual Ownership Expenses (USD) | |||||
| Depreciation and Interest | |||||
| Buildings and Structures | 116 | 1746.74 | 0.175 | ||
| Bee Pasture Equipment | 120 | 1803.84 | 0.180 | ||
| Repair and Management | 16 | 239.57 | 0.024 | ||
| Tractors and Vehicles | 422 | 6341.48 | 0.634 | ||
| Land | 17 | 252.03 | 0.025 | ||
| Interest on Loans | 0 | 0 | 0 | ||
| Insurance | 0 | 0 | 0 | ||
| Taxes | 195 | 2930.90 | 0.293 | ||
| Total Ownership Expenses | 886 | 13,314.57 | 1.331 | ||
| Total Annual Cost (USD) | 1197 | 17,999.43 | 1.800 |
| Native Bee Pasture Cost Line Items | Data Source(s) | Years | Producer Price Index Used to Adjust for Inflation [74] | Function Form for Probability Distribution Fit |
|---|---|---|---|---|
| Variable Costs | ||||
| Seeds (wildflower mix) | Neal 2019 [109]; OPN Seed [110]; Wong 2017 [111] | 2003–2022 | Hay, hayseeds and oilseeds | Triangular |
| Vermiculite | U.S. Geological Survey, National Minerals Information Center [112] | 2002–2022 | Nonmetallic mineral products | Triangular |
| Labor | U.S. Economic Research Service [113] | 2002–2022 | Already adjusted for inflation | Pareto |
| Diesel Fuel | USDA, ARS [114]; Hoshide et al., 2006 [115]; Asare et al., 2017 [26] | 2002–2023 | No. 2 diesel fuel | Triangular |
| Oil | Asare et al., 2017 [26] | 2002–2022 | Finished lubricants | Uniform |
| Maintenance and Upkeep | Adjusted in proportion to total capital used | 2002–2022 | Repair and maintenance services (partial) | Uniform |
| Fixed Costs | ||||
| Depreciation | ||||
| Equipment Storage | Asare et al., 2017 [26] | 2002–2022 | Farm service buildings and other prefab./portable | Triangular |
| Tiller (for tractor) | University of Minnesota Extension [116] | 2002–2022 | Farm plows, harrows, rollers, pulverizers, etc., and attachments | Uniform |
| Landscaping Rake | Venturini et al., 2017 [31] | 2002–2022 | All other farm machinery | Uniform |
| Paint bucket (seed) | Venturini et al., 2017 [31] | 2002–2022 | All other farm machinery | Uniform |
| Culti-Packer | Venturini et al., 2017 [31] | 2002–2022 | Farm plows, harrows, rollers, pulverizers, etc., and attachments | Uniform |
| Flail Mower (tractor) | Asare et al., 2017 [26] | 2002–2022 | Farm dairy equipment, sprayers and dusters, farm blowers, and attachments | Triangular |
| Hand Clippers | Venturini et al., 2017 [31] | 2002–2022 | All other farm machinery | Uniform |
| Computer | Asare et al., 2017 [26] | 2002–2022 | Personal computers and workstations | Uniform |
| Tractor | University of Minnesota Extension [116] | 2002–2022 | Total tractors | Triangular |
| Land | U.S. Economic Research Service [117] | 2002–2020 | Already adjusted for inflation | Triangular |
| Property Tax | Maine State Government [118] | 2012–2021 | Not adjusted since no PPI available | Triangular |


References
- Pettis, J.S.; Delaplane, K.S. Coordinated responses to honey bee decline in the USA. Apidologie 2010, 41, 256–263. [Google Scholar] [CrossRef]
- Tepedino, V.J.; Durham, S.; Cameron, S.A.; Goodell, K. Documenting bee decline or squandering scarce resources. Conserv. Biol. 2015, 29, 280–282. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Soroye, P.; Newbold, T.; Kerr, J. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Mathiasson, M.E.; Rehan, S.M. Wild bee declines linked to plant-pollinator network changes and plant species introductions. Insect Conserv. Divers. 2020, 13, 595–605. [Google Scholar] [CrossRef]
- Cariveau, D.P.; Winfree, R. Causes of variation in wild bee responses to anthropogenic drivers. Curr. Opin. Insect Sci. 2015, 10, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Scheper, J.; Reemer, M.; van Kats, R.; Ozinga, W.A.; van der Linden, G.T.; Schaminée, J.H.; Siepel, H.; Kleijn, D. Museum specimens reveal loss of pollen host plants as key factor driving wild bee decline in The Netherlands. Proc. Natl. Acad. Sci. USA 2014, 111, 17552–17557. [Google Scholar] [CrossRef] [PubMed]
- Zayed, A. Bee genetics and conservation. Apidologie 2009, 40, 237–262. [Google Scholar] [CrossRef]
- LeBuhn, G.; Luna, J.V. Pollinator decline: What do we know about the drivers of solitary bee declines? Curr. Opin. Insect Sci. 2021, 46, 106–111. [Google Scholar] [CrossRef]
- Jacobson, M.M.; Tucker, E.M.; Mathiasson, M.E.; Rehan, S.M. Decline of bumble bees in northeastern North America, with special focus on Bombus terricola. Biol. Conserv. 2018, 217, 437–445. [Google Scholar] [CrossRef]
- Bowers, M.A. Bumble Bee Colonization, Extinction, and Reproduction in Subalpine Meadows in Northeastern Utah: Ecological Archives E066-001. Ecology 1985, 66, 914–927. [Google Scholar] [CrossRef]
- Buchmann, S.; Ascher, J.S. The plight of pollinating bees. Bee World 2005, 86, 71–74. [Google Scholar] [CrossRef]
- Zattara, E.E.; Aizen, M.A. Worldwide occurrence records suggest a global decline in bee species richness. One Earth 2021, 4, 114–123. [Google Scholar] [CrossRef]
- Colla, S.R.; Ascher, J.S.; Arduser, M.; Cane, J.; Deyrup, M.; Droege, S.; Gibbs, J.; Griswold, T.; Hall, H.G.; Henne, C.; et al. Documenting persistence of most eastern North American bee species (Hymenoptera: Apoidea: Anthophila) to 1990–2009. J. Kans. Entomol. Soc. 2012, 85, 14–22. [Google Scholar] [CrossRef]
- Hamblin, A.L.; Youngsteadt, E.; Frank, S.D. Wild bee abundance declines with urban warming, regardless of floral density. Urban Ecosyst. 2018, 21, 419–428. [Google Scholar] [CrossRef]
- Lebuhn, G.; Droege, S.; Connor, E.F.; Gemmill-Herren, B.; Potts, S.G.; Minckley, R.L.; Griswold, T.; Jean, R.; Kula, E.; Roubik, D.W.; et al. Detecting insect pollinator declines on regional and global scales. Conserv. Biol. 2013, 27, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Roubik, D.W. Ups and downs in pollinator populations: When is there a decline? Conserv. Ecol. 2001, 5. Available online: https://www.jstor.org/stable/26271795 (accessed on 2 November 2023). [CrossRef]
- Decourtye, A.; Alaux, C.; Le Conte, Y.; Henry, M. Toward the protection of bees and pollination under global change: Present and future perspectives in a challenging applied science. Curr. Opin. Insect Sci. 2019, 35, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.R.; Zheng, H.; Gallant, A.L.; Iovanna, R.; Carlson, B.L.; Smart, M.D.; Hyberg, S. Past role and future outlook of the Conservation Reserve Program for supporting honey bees in the Great Plains. Proc. Natl. Acad. Sci. USA 2018, 115, 629–7634. [Google Scholar] [CrossRef]
- Winfree, R.; Gross, B.J.; Kremen, C. Valuing pollination services to agriculture. Ecol. Econ. 2011, 71, 80–88. [Google Scholar] [CrossRef]
- Carreck, N.; Williams, I. The economic value of bees in the UK. Bee World 1998, 79, 115–123. [Google Scholar] [CrossRef]
- Southwick, E.E.; Southwick, L., Jr. Estimating the economic value of honey bees (Hymenoptera: Apidae) as agricultural pollinators in the United States. J. Econ. Entomol. 1992, 85, 621–633. [Google Scholar] [CrossRef]
- Calderone, N.W. Insect pollinated crops, insect pollinators and US agriculture: Trend analysis of aggregate data for the period 1992–2009. PLoS ONE 2012, 7, e37235. [Google Scholar] [CrossRef] [PubMed]
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. Bioscience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Aizen, M.A.; Klein, A.M.; Cunningham, S.A.; Harder, L.D. Global growth and stability of agricultural yield decrease with pollinator dependence. Proc. Natl. Acad. Sci. USA 2011, 108, 5909–5914. [Google Scholar] [CrossRef] [PubMed]
- Asare, E.; Hoshide, A.K.; Drummond, F.A.; Chen, X.; Criner, G.K. Economic risk of bee pollination in Maine wild blueberry, Vaccinium angustifolium Aiton. J. Econ. Entomol. 2017, 110, 1980–1992. [Google Scholar] [CrossRef] [PubMed]
- Hoshide, A.K.; Drummond, F.A.; Stevens, T.H.; Venturini, E.M.; Hanes, S.P.; Sylvia, M.M.; Loftin, C.S.; Yarborough, D.E.; Averill, A.L. What is the value of wild bee pollination for wild blueberries and cranberries and who values it? Environments 2018, 5, 98. [Google Scholar] [CrossRef]
- Khalifa, S.A.; Elshafiey, E.H.; Shetaia, A.A.; El-Wahed, A.A.A.; Algethami, A.F.; Musharraf, S.G.; AlAjmi, M.F.; Zhao, C.; Masry, S.H.; Abdel-Daim, M.M.; et al. Overview of bee pollination and its economic value for crop production. Insects 2021, 12, 688. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Lippert, C.; Feuerbacher, A.; Narjes, M. Revisiting the economic valuation of agricultural losses due to large-scale changes in pollinator populations. Ecol. Econ. 2021, 180, 106860. [Google Scholar] [CrossRef]
- Venturini, E.M.; Drummond, F.A.; Hoshide, A.K.; Dibble, A.C.; Stack, L.B. Pollination reservoirs in Maine lowbush blueberry (Ericales: Ericaceae). J. Econ. Entomol. 2017, 110, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Kleijn, D.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.; Dainese, M.; Drummond, F.A.; Entling, M.; Ganser, D.; et al. A quantitative synthesis reveals flower plantings underpinning ecological intensification through enhanced pest control services and identifies drivers of increased crop pollination services. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Egan, P.A.; Dicks, L.V.; Hokkanen, H.M.; Stenberg, J.A. Delivering integrated pest and pollinator management (IPPM). Trends Plant Sci. 2020, 25, 577–589. [Google Scholar] [CrossRef] [PubMed]
- McDougall, R.; DiPaola, A.; Blaauw, B.; Nielsen, A.L. Managing orchard groundcover to reduce pollinator foraging post-bloom. Pest Manag. Sci. 2021, 77, 3554–3560. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodivers. Conserv. 2019, 28, 1343–1360. [Google Scholar] [CrossRef]
- Papanikolaou, A.D.; Kühn, I.; Frenzel, M.; Schweiger, O. Semi-natural habitats mitigate the effects of temperature rise on wild bees. J. Appl. Ecol. 2017, 54, 527–536. [Google Scholar] [CrossRef]
- Klaus, F.; Tscharntke, T.; Bischoff, G.; Grass, I. Floral resource diversification promotes solitary bee reproduction and may offset insecticide effects–evidence from a semi-field experiment. Ecol. Lett. 2021, 24, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Kline, O.; Joshi, N.K. Mitigating the effects of habitat loss on solitary bees in agricultural ecosystems. Agriculture 2020, 10, 115. [Google Scholar] [CrossRef]
- Durant, J.L.; Otto, C.R. Feeling the sting? Addressing land-use changes can mitigate bee declines. Land Use Policy 2019, 87, 104005. [Google Scholar] [CrossRef]
- Senapathi, D.; Biesmeijer, J.C.; Breeze, T.D.; Kleijn, D.; Potts, S.G.; Carvalheiro, L.G. Pollinator conservation—The difference between managing for pollination services and preserving pollinator diversity. Curr. Opin. Insect Sci. 2015, 12, 93–101. [Google Scholar] [CrossRef]
- Hall, D.M.; Steiner, R. Policy content analysis: Qualitative method for analyzing sub-national insect pollinator legislation. MethodsX 2020, 7, 100787. [Google Scholar] [CrossRef]
- Hipólito, J.; Coutinho, J.; Mahlmann, T.; Santana, T.B.R.; Magnusson, W.E. Legislation and pollination: Recommendations for policymakers and scientists. Perspect. Ecol. Conserv. 2021, 19, 1–9. [Google Scholar] [CrossRef]
- Faichnie, R.; Breeze, T.D.; Senapathi, D.; Garratt, M.P.; Potts, S.G. Scales matter: Maximising the effectiveness of interventions for pollinators and pollination. Adv. Ecol. Res. 2021, 64, 105–147. [Google Scholar] [CrossRef]
- Maderson, S.; Wynne-Jones, S. Beekeepers’ knowledges and participation in pollinator conservation policy. J. Rural Stud. 2016, 45, 88–98. [Google Scholar] [CrossRef]
- Baldock, K.C.R. Opportunities and threats for pollinator conservation in global towns and cities. Curr. Opin. Insect Sci. 2020, 38, 63–71. [Google Scholar] [CrossRef]
- Breeze, T.D.; Gallai, N.; Garibaldi, L.A.; Li, X.S. Economic measures of pollination services: Shortcomings and future directions. Trends Ecol. Evol. 2016, 31, 927–939. [Google Scholar] [CrossRef]
- Porto, R.G.; de Almeida, R.F.; Cruz-Neto, O.; Tabarelli, M.; Viana, B.F.; Peres, C.A.; Lopes, A.V. Pollination ecosystem services: A comprehensive review of economic values, research funding and policy actions. Food Secur. 2020, 12, 1425–1442. [Google Scholar] [CrossRef]
- United Nations (U.N.). The 17 Goals: Sustainable Development Goals. Department of Economic and Social Affairs, Sustainable Development. Available online: https://sdgs.un.org/goals (accessed on 16 January 2024).
- Cole, L.J.; Kleijn, D.; Dicks, L.V.; Stout, J.C.; Potts, S.G.; Albrecht, M.; Balzan, M.V.; Bartomeus, I.; Bebeli, P.J.; Bevk, D.; et al. A critical analysis of the potential for EU common agricultural policy measures to support wild pollinators on farmland. J. Appl. Ecol. 2020, 57, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Bommarco, R.; Felicioli, A.; Fischer, M.; Jokinen, P.; Kleijn, D.; Klein, A.M.; Kunin, W.E.; Neumann, P.; et al. Developing European conservation and mitigation tools for pollination services: Approaches of the STEP (Status and Trends of European Pollinators) project. J. Apic. Res. 2011, 50, 152–164. [Google Scholar] [CrossRef]
- Dicks, L.V.; Baude, M.; Roberts, S.P.; Phillips, J.; Green, M.; Carvell, C. How much flower-rich habitat is enough for wild pollinators? Answering a key policy question with incomplete knowledge. Ecol. Entomol. 2015, 40, 22–35. [Google Scholar] [CrossRef]
- Potts, S.G.; Ngo, H.T.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; Vanbergen, A. The assessment report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on pollinators, pollination and food production. In IPBES (2016): Summary for Policymakers of the Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination and Food Production; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016; 36p, ISBN 978-92-807-3568-0. [Google Scholar]
- Williams, I.H. Insect pollination and crop production: A European perspective. In Pollinating Bees–The Conservation Link between Agriculture and Nature; Kevan, P., Imperatriz, F.V.L., Eds.; Ministry of the Environment: Brasilia, Brazil, 2002; pp. 59–65. Available online: https://grist.org/wp-content/uploads/2012/01/livro_02_willians.pdf (accessed on 8 January 2024).
- Dicks, L.V.; Viana, B.; Bommarco, R.; Brosi, B.; Arizmendi, M.; Cunningham, S.A.; Galetto, L.; Hill, R.; Lopes, A.V.; Pires, C.; et al. Ten policies for pollinators. Science 2016, 354, 975–976. [Google Scholar] [CrossRef] [PubMed]
- Rose, T.; Kremen, C.; Thrupp, A. Policy Analysis Paper: Policy Mainstreaming of Biodiversity and Ecosystem Services with a Focus on Pollination; FAO: Rome, Italy, 2014; Available online: https://escholarship.org/uc/item/4p8419st (accessed on 2 November 2023).
- Bloom, E.H.; Graham, K.K.; Haan, N.L.; Heck, A.R.; Gut, L.J.; Landis, D.A.; Milbrath, M.O.; Quinlan, G.M.; Wilson, J.K.; Zhang, Y.; et al. Responding to the US national pollinator plan: A case study in Michigan. Front. Ecol. Environ. 2022, 20, 84–92. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Pérez-Méndez, N.; Garratt, M.P.; Gemmill-Herren, B.; Miguez, F.E.; Dicks, L.V. Policies for ecological intensification of crop production. Trends Ecol. Evol. 2019, 34, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Yarborough, D.E. Establishment and management of the cultivated lowbush blueberry (Vaccinium angustifolium). Internat. J. Fruit Sci. 2012, 12, 14–22. [Google Scholar] [CrossRef]
- Bushmann, S.A.; Drummond, F.A. Analysis of Pollination Services Provided by Wild and Managed Bees (Apoidea) in Wild Blueberry (Vaccinium angustifolium Aiton) Production in Maine, USA, with a Literature Review. Agronomy 2020, 10, 1413. [Google Scholar] [CrossRef]
- Drummond, F.A. Wild Blueberry Annual Research Reports; University of Maine: Orono, ME, USA, 2024. [Google Scholar]
- Qu, H.; Drummond, F.A. Simulation-based modeling of wild blueberry pollination. Comput. Electron. Agric. 2018, 144, 94–101. [Google Scholar] [CrossRef]
- SAS Institute. JMP(R), Version 16; SAS Institute Inc.: Cary, NC, USA, 2022.
- Palisade. Guide to Using @Risk: Risk Analysis and Simulation Add-In for Microsoft Excel; Version 7.5; Palisade Corporation: New Field, NY, USA, 2016. [Google Scholar]
- Hardaker, J.B.; Huirne, R.B.M.; Anderson, J.R.; Lien, G. Coping with Risk in Agriculture, 3rd ed.; CAB International Publishing: Oxfordshire, UK, 2015; pp. 1–289. [Google Scholar]
- Hummel, R.M.; Claassen, E.A.; Wolfinger, R.D. JMP for Mixed Models; SAS Institute Incorporated; SAS Campus Drive: Cary, NC, USA, 2021; pp. 1–243. Available online: https://www.jmp.com/en_us/whitepapers/book-chapters/jmp-for-mixed-models.html (accessed on 5 April 2024).
- Boker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. General linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Box, G.E.; Jenkins, G.M.; Reinsel, G.C.; Ljung, G.M. Time Series Analysis: Forecasting and Control, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; 712p, ISBN 978-1-118-67502-1. [Google Scholar]
- Stern, V.M.; Smith, R.F.; Van Den Bosch, R.; Hagen, K.S. The integrated control concept. Hilgardia 1959, 29, 81–101. [Google Scholar] [CrossRef]
- Pedigo, K.P.; Hutchins, S.H.; Higley, L.G. Economic injury levels in theory and practice. Annu. Rev. Entomol. 1986, 31, 341–368. [Google Scholar] [CrossRef]
- Javorek, S.K.; MacKenzie, K.E.; Kloet, S.P.V. Comparative Pollination Effectiveness among Bees (Hymenoptera: Apoidea) on Lowbush Blueberry (Ericaceae: Vaccinium angustifolium). Ann. Entomol. Soc. Am. 2002, 95, 345–351. [Google Scholar] [CrossRef]
- Drummond, F.A. Reproductive biology of wild blueberry (Vaccinium angustifolium Aiton). Agriculture 2019, 9, 69. [Google Scholar] [CrossRef]
- U.S. Bureau of Labor Statistics. Data Tools, Data Retrieval Tools, Series Report. Available online: https://data.bls.gov/cgi-bin/srgate (accessed on 10 November 2023).
- International Monetary Fund. Producer Price Index Manual: Theory and Practice. Manuals and Guides. 2004. Available online: https://www.imf.org/en/Publications/Manuals-Guides/Issues/2016/12/30/Producer-Price-Index-Manual-Theory-and-Practice-16966 (accessed on 7 December 2023).
- U.S. Department of Agriculture, National Agricultural Statistics Service. Quick Stats. Available online: https://quickstats.nass.usda.gov/results/A2524626-1096-3EF6-A6D7-CF842A803783 (accessed on 10 November 2023).
- Langemeier, M. Trends in General Inflation and Farm Input Prices. Center for Commercial Agriculture, Purdue University. 2022. Available online: https://ag.purdue.edu/commercialag/home/wp-content/uploads/2022/04/20220429_Langemeier-TrendsInGeneralInflationAndFarmInputPrices.pdf (accessed on 7 December 2023).
- David, A. Freedman. Statistical Models: Theory and Practice; Cambridge University Press: Cambridge, UK, 2009; pp. 1–458. ISBN 978-1-139-47731-4. [Google Scholar]
- Cutler, G.C.; Nams, V.O.; Craig, P.; Sproule, J.M.; Sheffield, C.S. Wild bee pollinator communities of lowbush blueberry fields: Spatial and temporal trends. Basic Appl. Ecol. 2015, 16, 73–85. [Google Scholar] [CrossRef]
- Eaton, L.J.; Murray, J.E. Relationships of pollinator numbers in blueberry fields to fruit development and yields. Acta Hortic. 1997, 446, 181–188. [Google Scholar] [CrossRef]
- Eaton, L.J.; Nams, V.O. Honeybee stocking numbers and wild blueberry production in Nova Scotia. Can. J. Plant Sci. 2012, 92, 1305–1310. [Google Scholar] [CrossRef]
- Yarborough, D.; Drummond, F.; Annis, S.; D’Appollonio, J. Maine Wild blueberry systems analysis. Acta Hort. 2017, 1180, 151–160. [Google Scholar] [CrossRef]
- Boulanger, L.W. Blueberry pollination and solitary bees. Maine Farm Res. 1964, 12, 5–11. [Google Scholar]
- Desjardins, E.; de Oliveira, D. Commercial bumble bee Bombus impatiens (Hymenoptera: Apidae) as a pollinator in lowbush blueberry (Ericale: Ericaceae) fields. J. Econ. Entomol. 2006, 99, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.; Hill, R.; Settele, J.; et al. Safeguarding Pollinators and Their Values to Human Well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef]
- Evans, J.D.; Chen, Y. Colony collapse disorder and honey bee health. In Honey Bee Medicine for the Veterinary Practitioner, 1st ed.; Kane, T.R., Faux, C.M., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2021; Chapter 19; pp. 229–234. ISBN 9781119583417. [Google Scholar] [CrossRef]
- Kerr, J.T.; Pindar, A.; Galpern, P.; Packer, L.; Potts, S.G.; Roberts, S.M.; Rasmont, P.; Schweiger, O.; Colla, S.R.; Richardson, L.L. Climate Change Impacts on Bumblebees Converge Across Continents. Science 2015, 349, 177–180. [Google Scholar] [CrossRef]
- Maine Bee Atlas. Available online: https://mainebumblebeeatlas.umf.maine.edu (accessed on 16 January 2024).
- Scheper, J.A. Promoting wild bees in European agricultural landscapes. Alterra Sci. Contrib. 2015, 47, 1–176. Available online: https://www.wur.nl/upload_mm/c/b/a/a738b550-9f10-48ba-aa17-ef1755804127_Scheper_2015_PhD%20thesis_Promoting%20wild%20bees%20in%20European%20agricultural%20landscapes.pdf (accessed on 5 January 2024).
- Isaacs, R.; Williams, N.; Ellis, J.; Pitts-Singer, T.L.; Bommarco, R.; Vaughan, M. Integrated crop pollination: Combining strategies to ensure stable and sustainable yields of pollination-dependent crops. Basic Appl. Ecol. 2017, 22, 44–60. [Google Scholar] [CrossRef]
- Cortés-Rivas, B.; Monzón, V.H.; Rego, J.O.; Mesquita-Neto, J.N. Pollination by native bees achieves high fruit quantity and quality of highbush blueberry: A sustainable alternative to managed pollinators. Front. Sustain. Food Syst. 2023, 7, 1142623. [Google Scholar] [CrossRef]
- Cooperative Extension Wild Blueberries. Available online: https://extension.umaine.edu/blueberries/ (accessed on 15 January 2024).
- Fanning, P.; Collins, J. 2023 Insect Pest Management Guide: Insects. University of Maine Cooperative Extension, Maine Wild Blueberries. Available online: https://extension.umaine.edu/blueberries/wp-content/uploads/sites/41/2022/02/2022-insecticide-chart.pdf (accessed on 13 January 2024).
- Markuson, J. Integrated Pest Pollinator Management. In Proceedings of the NRCS Bee Conservation Presentation, 2020 Wild Blueberry Conference, Bangor, ME, USA, 22 February 2020; Available online: https://extension.umaine.edu/blueberries/integrated-pest-and-pollinator-management-ippm/ (accessed on 13 December 2023).
- Blaauw, B.R.; Isaacs, R. Flower plantings increase wild bee abundance and the pollination services provided to a pollination-dependent crop. J. Appl. Ecol. 2014, 51, 890–898. [Google Scholar] [CrossRef]
- Tuell, J.K.; Isaacs, R. Weather during bloom affects pollination and yield of highbush blueberry. J. Econ. Entomol. 2010, 103, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Hanes, S.P.; Collum, K.K.; Hoshide, A.K.; Asare, E. Grower perceptions of native pollinators and pollination strategies in the lowbush blueberry industry. Renew. Agric. Food Syst. 2015, 30, 124–131. [Google Scholar] [CrossRef]
- Hanes, S.P.; Collum, K.K.; Drummond, F.A.; Hoshide, A.K. Assessing Wild Pollinators in Conventional Agriculture: A Case Study from Maine’s Blueberry Industry. Hum. Ecol. Rev. 2018, 24, 97–113. [Google Scholar] [CrossRef]
- Gemmill-Herren, B.; Garibaldi, L.A.; Kremen, C.; Ngo, H.T. Building effective policies to conserve pollinators: Translating knowledge into policy. Curr. Opin. Insect Sci. 2021, 46, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, A.A.; Epstein, G.B.; Colla, S.R. Understanding public and stakeholder attitudes in pollinator conservation policy development. Environ. Sci. Policy 2020, 111, 27–34. [Google Scholar] [CrossRef]
- Galetto, L.; Aizen, M.A.; Arizmendi, M.D.C.; Freitas, B.M.; Garibaldi, L.A.; Giannini, T.C.; Lopes, A.V.; Santo, M.M.D.E.; Maués, M.M.; Nates-Parra, G.; et al. Risks and opportunities associated with pollinators’ conservation and management of pollination services in Latin America. Ecol. Austral 2022, 32, 55–76. [Google Scholar] [CrossRef]
- Nalepa, R.; Colla, S.R. Toward a wild pollinator strategy for Canada: Expert-recommended solutions and policy levers. FACETS 2023, 8, 1–18. [Google Scholar] [CrossRef]
- Vanegas, M. The Silent Beehive. Ecol. Law Q. 2017, 44, 311–342. [Google Scholar] [CrossRef]
- Giovanetti, M.; Bortolotti, L. Pollinators and policy: The intersecting path of various actors across an evolving CAP. Renew. Agric. Food Syst. 2023, 38, e27. [Google Scholar] [CrossRef]
- Ratamäki, O.; Jokinen, P.; Sørensen, P.B.; Breeze, T.D.; Potts, S. A multilevel analysis on pollination-related policies. Ecosyst. Serv. 2015, 14, 133–143. [Google Scholar] [CrossRef]
- Steele, D.J.; Baldock, K.C.R.; Breeze, T.D.; Brown, M.J.F.; Carvell, C.; Dicks, L.V.; Garratt, M.P.; Norman, H.; Potts, S.G.; Senapathi, D.; et al. Management and Drivers of Change of Pollinating Insects and Pollination Services. National Pollinator Strategy: For Bees and Other Pollinators in England, Evidence Statements and Summary of Evidence; Project Report; Department for Environment, Food and Rural Affairs: London, UK, 2019; pp. 1–91. Available online: https://nrl.northumbria.ac.uk/id/eprint/42117/ (accessed on 7 January 2024).
- Hall, D.M.; Steiner, R. Insect pollinator conservation policy innovations at subnational levels: Lessons for lawmakers. Environ. Sci. Policy 2019, 93, 118–128. [Google Scholar] [CrossRef]
- Whitney, K.S.; Stringer, B.B. Evaluation of US state pollinator plans using 3 evidence-based policymaking frameworks. bioRxiv 2021. [Google Scholar] [CrossRef]
- Drummond, F.A. Factors that affect yield in wild blueberry, Vaccinium angustifolium Aiton. Agric. Res. Technol. 2019, 22, 556212. [Google Scholar] [CrossRef]
- Drummond, F.A. Wild blueberry fruit drop: A consequence of seed set? Special Issue: Pollinator Diversity and Pollination in Agricultural Systems. Agronomy 2020, 10, 939. [Google Scholar] [CrossRef]
- Neal, C. University of New Hampshire Cooperative Extension. Planting for Pollinators: Establishing a Wildflower Meadow from Seed. Fact Sheet. 2019. Available online: https://extension.unh.edu/sites/default/files/migrated_unmanaged_files/Resource007652_Rep11219.pdf (accessed on 10 November 2023).
- OPN Seed. Eastern Great Lakes Native Pollinator Seed Mix. Available online: https://www.opnseed.com/products/eastern-great-lakes-native-pollinator-mix?variant=40552977694918 (accessed on 10 November 2023).
- Wong, D. Peace Region Forage Seed Association. Historical Grass and Legume Seed Prices: 1970 to 2016. White Paper, 2017; pp. 1–4. Available online: http://www.peaceforageseed.ca/pdf/markets_2013_onwards/2017/Historical_prices_to_2016.pdf (accessed on 10 November 2023).
- U.S. Geological Survey, National Minerals Information Center. Vermiculite–Historical Statistics (Data Series 140). 2020. Available online: https://www.usgs.gov/media/files/vermiculite-historical-statistics-data-series-140 (accessed on 10 November 2023).
- U.S. Department of Agriculture, Economic Research Service. Real Wages for U.S. Nonsupervisory Farm and Nonfarm Workers, 1990–2022. Available online: https://www.ers.usda.gov/webdocs/charts/63462/wagerates2022linesbar_d.html?v=5868.1 (accessed on 10 November 2023).
- U.S. Department of Agriculture, Agricultural Marketing Service. Historical Diesel Fuel Prices, Annual Average U.S. On-Highway Diesel Fuel Prices ($/gallon). Available online: https://agtransport.usda.gov/Fuel/Historical-Diesel-Fuel-Prices/u2kh-s8ke (accessed on 10 November 2023).
- Hoshide, A.K.; Dalton, T.J.; Smith, S.N. Profitability of coupled potato and dairy farms in Maine. Renew. Agric. Food Sys. 2006, 21, 261–272. [Google Scholar] [CrossRef]
- Lazarus, W.F. Machinery Cost Estimates (2004, 2008, 2012, 2013, 2021, and 2023). University of Minnesota Extension. Available online: https://wlazarus.cfans.umn.edu/william-f-lazarus-farm-machinery-management (accessed on 10 November 2023).
- U.S. Department of Agriculture, Economic Research Service. Average U.S. Farm Real Estate Value, Nominal and Real (Inflation Adjusted), 1970–2020. Available online: https://www.ers.usda.gov/webdocs/charts/55910/farmrealestatevalue2020_d.html?v=9097.6 (accessed on 10 November 2023).
- Maine Revenue Services. Estimated Full Value Tax Rates (2012–2021). Available online: https://www.maine.gov/revenue/sites/maine.gov.revenue/files/inline-files/fullvaluerates.pdf (accessed on 10 November 2023).








| Model a and r2 | Effects | DF b (Num, Den) | F Ratio | p-Values c | Coefficients d |
|---|---|---|---|---|---|
| 1. Fruit set (%) | Study—random | <0.0001 | |||
| r2 = 0.42 | Year (Study)—random | <0.0001 | |||
| Native bee density—fixed | 1, 206 | 11.191 | 0.0002 | 2.676 | |
| Honey bee density—fixed | 1, 206 | 21.779 | <0.0001 | 0.185 | |
| 2. Yield (kg/ha) | Study—random | 0.511 | |||
| r2 = 0.52 | Year (Study)—random | 0.028 | |||
| % Fruit set—fixed | 1, 11,870 | 83.039 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drummond, F.A.; Hoshide, A.K. An Economic Cost/Benefit Tool to Assess Bee Pollinator Conservation, Pollination Strategies, and Sustainable Policies: A Lowbush Blueberry Case Study. Sustainability 2024, 16, 3242. https://doi.org/10.3390/su16083242
Drummond FA, Hoshide AK. An Economic Cost/Benefit Tool to Assess Bee Pollinator Conservation, Pollination Strategies, and Sustainable Policies: A Lowbush Blueberry Case Study. Sustainability. 2024; 16(8):3242. https://doi.org/10.3390/su16083242
Chicago/Turabian StyleDrummond, Francis A., and Aaron Kinyu Hoshide. 2024. "An Economic Cost/Benefit Tool to Assess Bee Pollinator Conservation, Pollination Strategies, and Sustainable Policies: A Lowbush Blueberry Case Study" Sustainability 16, no. 8: 3242. https://doi.org/10.3390/su16083242
APA StyleDrummond, F. A., & Hoshide, A. K. (2024). An Economic Cost/Benefit Tool to Assess Bee Pollinator Conservation, Pollination Strategies, and Sustainable Policies: A Lowbush Blueberry Case Study. Sustainability, 16(8), 3242. https://doi.org/10.3390/su16083242