
Characterization of Fish Spawning Grounds near the Likouala-Aux-Herbes River, Lac Tele Community Reserve (LTCR), Republic of Congo, for Sustainable Wetland Management



Abstract
1. Introduction
2. Materials and Methods
- -
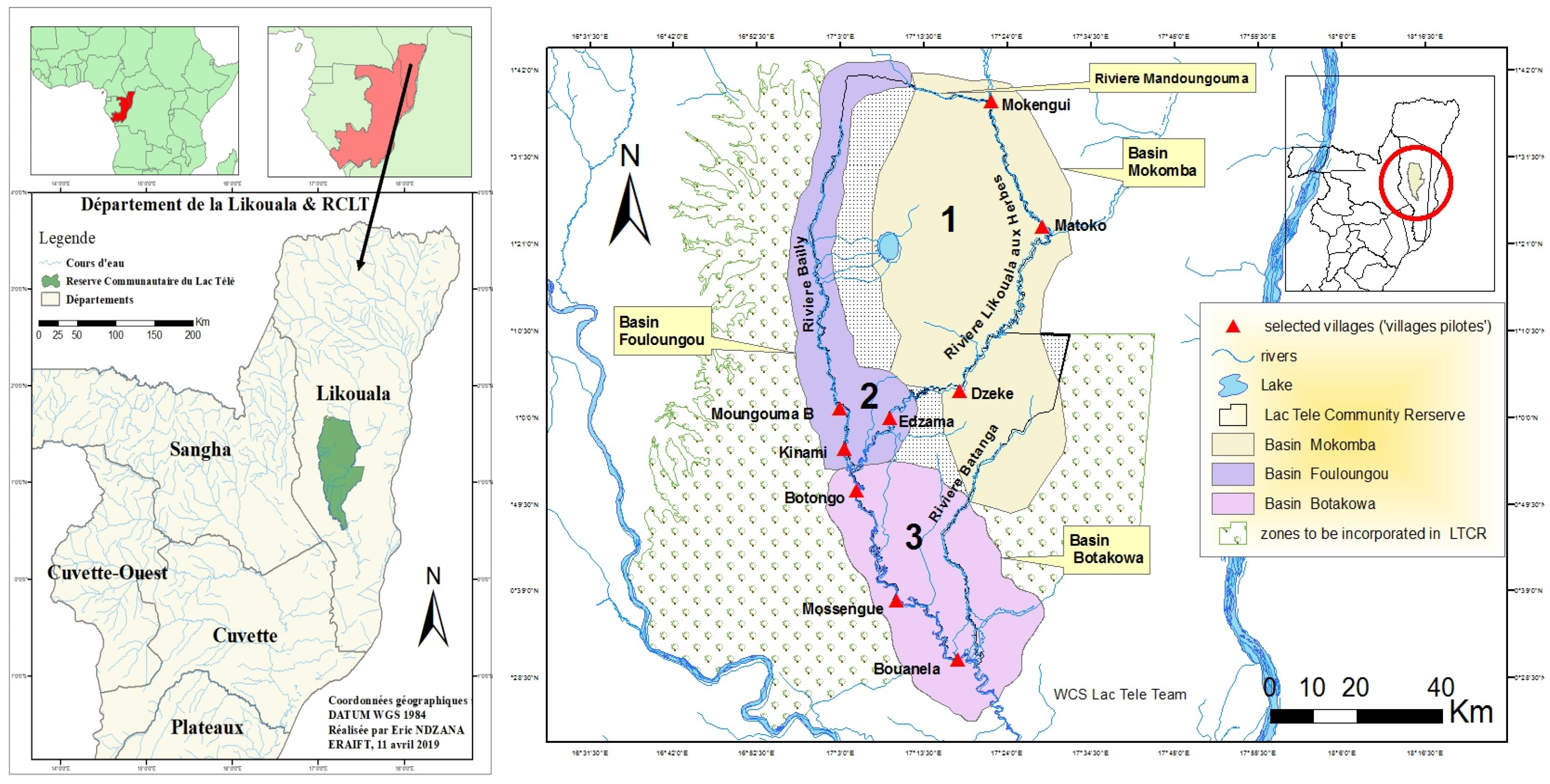
- 1st station: upstream of the village (northern limit of the spawning grounds) (i).
- -
- 2nd station: village area (near the main port of the village) (ii).
- -
- 3rd station: downstream of the village (southern limit, of the h spawning grounds) (iii).

- Changes: the mode of space occupation remained the same between the two years (initial state).
- Modification: the mode of occupation of space has changed from one class to another, but remained in the same category.
- Conversion: the mode of occupation of the space of a class has converted to another class in a different category (example: Gallery forest became herbaceous or tree savannah). To quantify land use changes, the annual expansion rate method was used. It consisted of calculating the annual expansion rate using the formula proposed by the FAO in 1996, as follows: T = S2 − S1; where T = rate, S1 = area of class in the first year, and S2 = area of class in the second year. Regarding the value of S (S in unit), if S is a positive value, there was an increase in the area of the classes during the period analyzed, and if S is a negative value, there was a loss of area in the classes between the two periods. A value close to zero means that no change in the classes was noted between the two periods [21].
3. Results
3.1. Descption of Differents Results
3.1.1. Spatiotemporal Dynamics of Land Use in the Likouala-aux-Herbes Watershed between 1986, 1999, and 2019
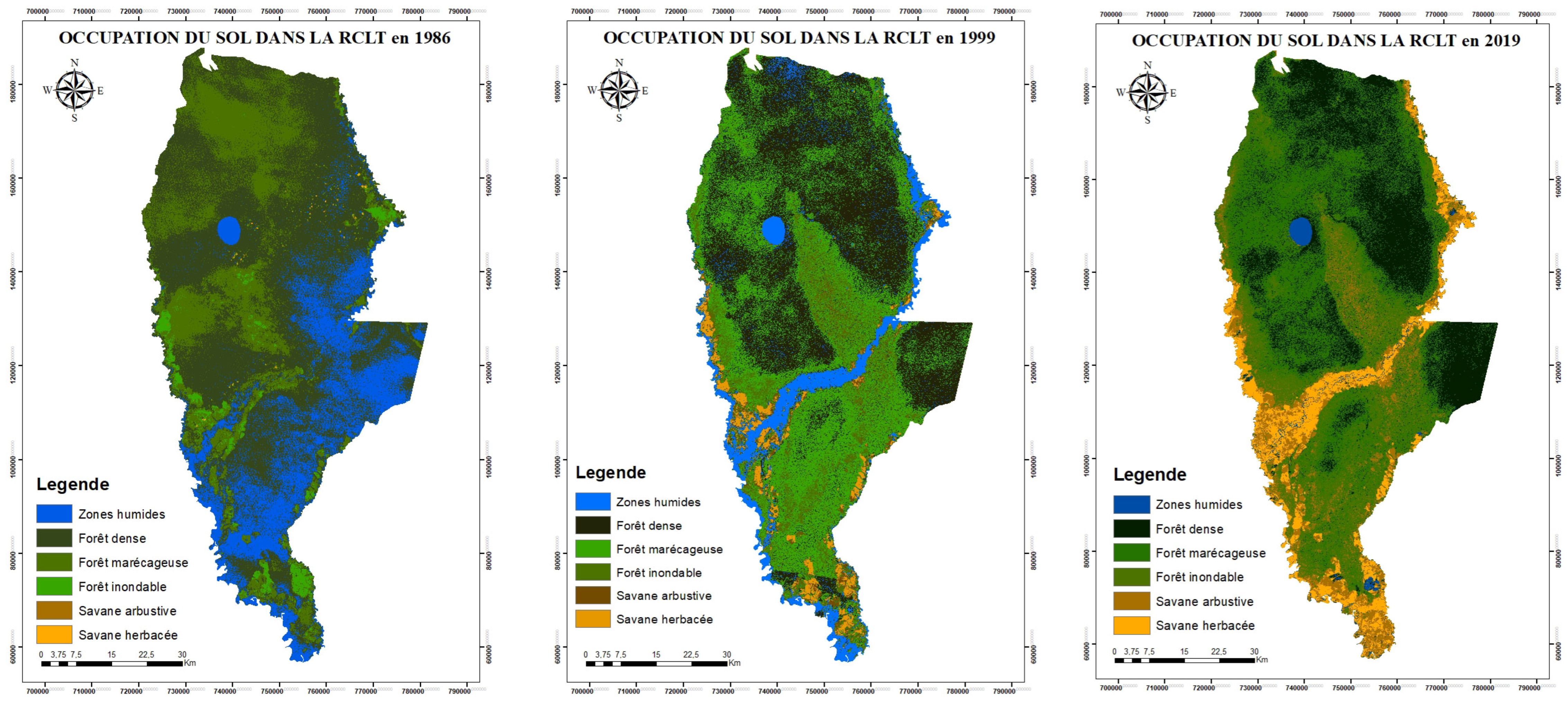
- The distribution of land use (Figure 2) classes between 1986, 1999, and 2019 shows a relative conversion of forest (dense, swampy, floodable, and gallery) of the LTCR environment into savannah (herbaceous and shrubby).
- We observe a predominance of dense and swampy forests and a savannah dynamic around the watershed of the Likouala-aux-Herbes River.
- According to the analysis shown in 1986 (Table 1), the forest class was divided into dense forest, covering 53.4% (241,021.98 ha) of the LTCR; swamp forest, comprising 24% (108,334.26 ha) of the LTCR; and floodable forest, covering 2.5% (11,442.78 ha) of the area. Of the total (79.5%), in 1999, this progressed to 85%, comprised of 39.02% dense forest (176,187.78 ha), 34% swamp forest (153,325.44 ha), and 12.14% floodable forest (54,846.81 ha).
- During the same period, the shrub and herbaceous savannah classes increased from 0.018% in 1986 to 5% of the area of the LTCR in 1999 (Table 2), from 642.06 ha to 25,403.04 ha, respectively, and increasing to 17.8% in 2019, with 9.4% of shrub savannah (42,444.9 ha) and 8.4% (38,128.77 ha) of herbaceous savannah.
- The wetlands class occupied 20% of the LTCR in 1986 (90,012.06 ha), 10% in 1999 (41,690.07 ha), and 1% (4701.42 ha) in 2019.
3.1.2. Characterization of the Ichthyological Diversity of the Likouala-Aux-Herbes River
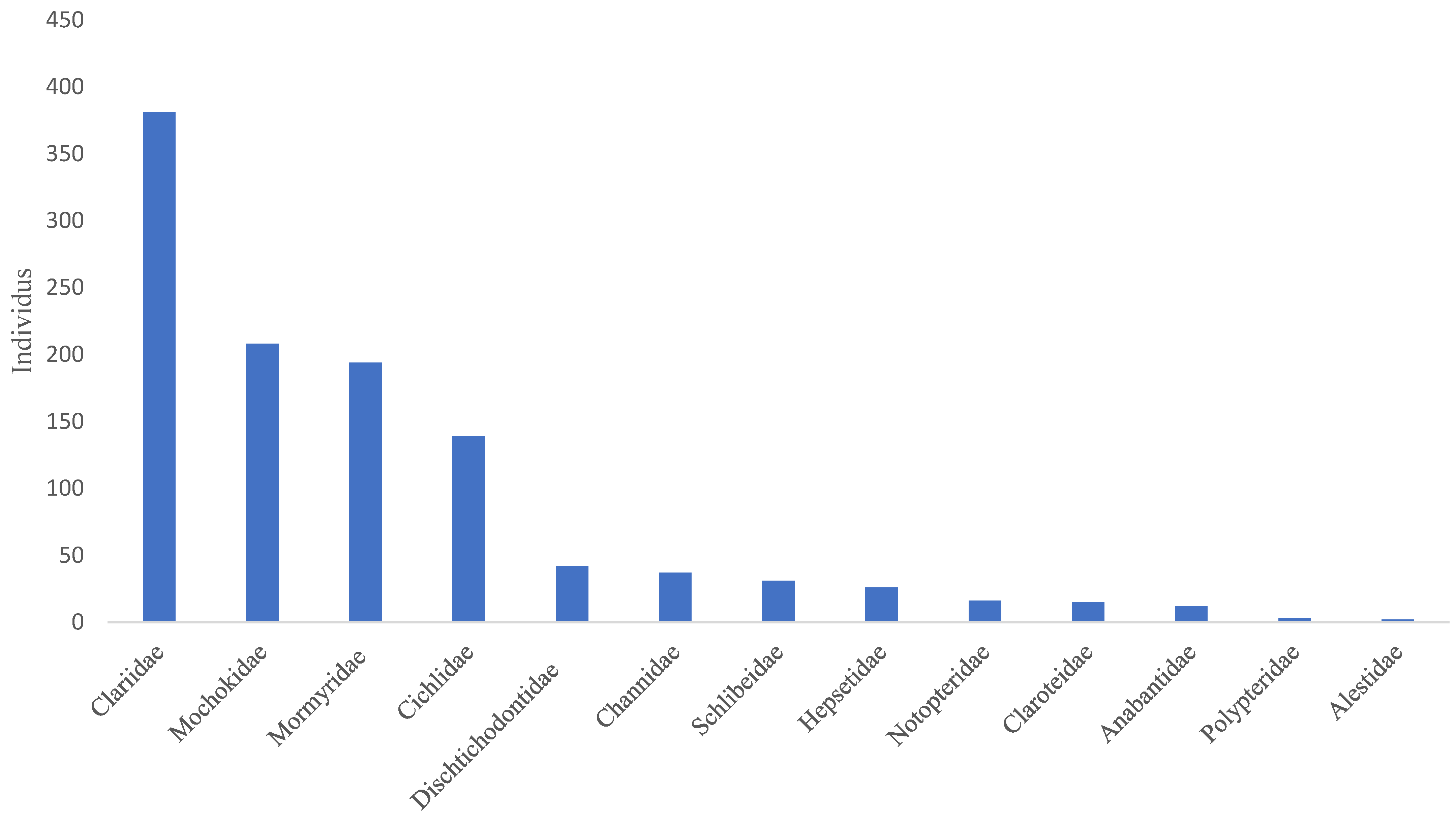
- The Clariidae family was the most abundant, with 381 specimens, representing 34.07%, followed by the Mochokidae family, with 208 individuals, or 19.67%. The Bouanela and Mossengue stations contain the largest numbers of individuals, representing respectively 20.61% and 31.1%, while the Moungouma-Bailly and Dzeke stations recorded the lowest numbers of 8 and 28 individuals, i.e., 0.72% and 2.53% (Figure 3 and Table 4 and Table 5).
- The presence of Alestidae (Component 2, Figure 3) is correlated with temperature peaks, namely 28.79 ± 1.7 °C for the Botongo station and 28.13 ± 1.17 °C for the Epena station (Table 5). As for the Mormyridae, the highest diversity indices in this case, 0.6229 for Mossengue and 0.5993 for Botongo, were recorded at the stations with the highest temperature peaks.
- The results of the principal component analysis (PCA) regarding the spatial distribution of the captured species made it possible to select the environmental and biotic variables which best express the links that exist between species and the parameters of the various aquatic habitats.
- Overall, considering all the stations, the environmental variables which influenced the distribution of the numbers of fish species recorded during the study period are the temperature, the width of the wetted section, the depth, the flow rate, and the composition of the different substrates encountered in the stations.
- reveals that the 13 families of fish species recorded in the different pilot villages were distributed according to a combination of the physicochemical parameters and nature of the substrate. Thus, following axis I (Component 1, Figure 4), the Cichlidae, Channidae, Claroteidae, and Mochokidae samples presented more affinity with the width of the wetted section of the medium, the depth, the flow, and the substrate of a clay–muddy nature.
- The highest specific richness values, namely (4 taxa) for Cichlidae and (3 taxa) for Mochokidae, were recorded in the Botongo and Mossengue stations, whose substrate composition is mud–sandy and exhibits the highest average widths.
- In axis II of the PCA (Component 2), the distribution of the Alestidae and Mormyridae families seems to be influenced by the presence of sand, foliage, and macrophyte type substrates, coupled with the increase in temperature of the water. The presence of Alestidae, is correlated with temperature peaks, namely 28.79 ± 1.7 °C for the Botongo station and 28.13 ± 1.17 °C for the Epena station. As for the Mormyridae samples, the highest diversity indices, 0.6229 for Mossengue and 0.5993 for Botongo, were recorded in the stations whose temperature peaks are among the highest.
3.2. Figures, Tables, and Schemes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Translation | Areas (ha) | Expansion Rate | |||
|---|---|---|---|---|---|
| CLASSES | CLASSES | 1986 | 1999 | Hectares (ha) | Percentage (%) |
| Zones humides | Wetlands | 90,012.06 | 41,690.07 | −48,321.99 | 53.68390636 |
| Forêt dense | Dense forest | 241,021.98 | 176,187.78 | −64,834.2 | 26.89970433 |
| Forêt marécageuse | Swamp forest | 108,334.26 | 153,325.44 | 44,991.18 | 41.52996476 |
| Forêt inondable | Floodable forest | 11,442.78 | 54,846.81 | 43,404.03 | 379.3136808 |
| Savane arbustive | Shrub savannah | 378 | 11,488.32 | 11,110.32 | 2,939.238095 |
| Savane herbacée | Herbaceous savannah | 264.06 | 13914.72 | 13,650.66 | 5169.529652 |
| Translation | Areas (ha) | Expansion Rate | |||
|---|---|---|---|---|---|
| CLASSES | CLASSES | 1999 | 2019 | Hectares (ha) | Percentage (%) |
| Zones humides | Wetlands | 41,690.07 | 4,701.42 | -36,988.65 | 88.72292611 |
| Forêt dense | Dense forest | 176,187.78 | 119,643.84 | -56,543.94 | 32.0929976 |
| Forêt marécageuse | Swamp forest | 153,325.44 | 152,263.26 | -1,062.18 | 0.692761749 |
| Forêt inondable | Floodable forest | 54,846.81 | 94,270.95 | 39,424.14 | 71.88046123 |
| Savane arbustive | Shrub savannah | 11,488.32 | 42,444.9 | 30,956.58 | 269.4613312 |
| Savane herbacée | Herbaceous savannah | 13,914.72 | 38,128.77 | 24,214.05 | 174.0175153 |
| Polypteridae Polypterus delhezi (Boulenger, 1899) Mormyridae Petrocephalus ballayi (Sauvage, 1883) Petrocephalus microphthalmus (Pellegrin, 1908) Mormyrops boulengeri (Pellegrin, 1900) Marcusenius moorii (Günther, 1867) Marcusenius sp Gnatonemus petersii (Günther, 1862) Hippopotamyrus weeksii (Boulenger, 1902) Notopteridae Papyrogranus afer (Günther, 1868) Xenomystus nigri (Günther, 1868) Hepsetidae Hepsetus odoe (Bloch, 1794) Alestidae Alestes macrophthalmus (Günther, 1867) Distichodontidae Ichthyoborus ornatus (Boulenger, 1899) Distichodus noboli (Boulenger, 1899) Distichodus altus (Boulenger, 1899) Xenocharax spilurus (Günther, 1867) Anabantidae Microctenopoma nanum,(Guichenot, 1859) Ctenopoma kingsleyae Günther, 1896 | Claroteidae Chrysichthys sp. Chrysichthys cranchii (Burchell, 1822) Parauchenoglanis punctatus. (Sauvage, 1879) Parailia occidentalis (Pellegrin, 1901) Schilbe marmoratus Boulenger, 1911) Schilbe grenfelli (Boulenger, 1900) Clariidae Clarias buthupogon (Sauvage, 1879) Clarias spp. (Sauvage, 1879) Mochokidae Synodontis nigriventris (David, 1936) Synodontis camelopardalis (Poll, 1971) Synodontis falvitaeniatus (Boulenger, 1919) Channidae Parachanna obscura (Günther, 1861) Cichlidae Hemichromis bimaculatus (Gill, 1863) Hemichromis elongatus (Guichenot, 1859) Tilapia congica, (Gill, 1863) Tylochromis lateralis (Sauvage, 1879) |



| VILLAGES/FAMILIES | MOKENGUI | EPENA | DZEKE | MOUNGOUMA BAILLY | BOTONGO | MOSSENGUE | BOUANELA | TOTAL |
|---|---|---|---|---|---|---|---|---|
| Alestidae | 1 | 1 | 2 | |||||
| Anabantidae | 12 | 12 | ||||||
| Channidae | 1 | 2 | 14 | 20 | 37 | |||
| Cichlidae | 6 | 11 | 34 | 88 | 139 | |||
| Clariidae | 206 | 32 | 18 | 3 | 45 | 59 | 18 | 381 |
| Claroteidae | 13 | 1 | 1 | 15 | ||||
| Dischtichodontidae | 6 | 13 | 23 | 42 | ||||
| Hepsetidae | 3 | 9 | 14 | 26 | ||||
| Mochokidae | 3 | 22 | 100 | 83 | 208 | |||
| Mormyridae | 1 | 64 | 5 | 55 | 51 | 18 | 194 | |
| Notopteridae | 8 | 8 | 16 | |||||
| Polypteridae | 3 | 3 | ||||||
| Schlibeidae | 2 | 4 | 25 | 31 | ||||
| TOTAL | 207 | 108 | 28 | 8 | 183 | 344 | 228 | 1106 |
| 18.1 | 9.76 | 2.53 | 0.72 | 16.54 | 31.1 | 20.61 | 100.00 | |
| Indices of diversity (H’) | ||||||||
| Taxa (N) | 2 | 6 | 4 | 2 | 11 | 12 | 6 | |
| Individus (N) | 207 | 108 | 28 | 8 | 183 | 344 | 228 | |
| (H’) | 0.0132 | 0.455 | 0.4223 | 0.287 | 0.8477 | 0.9118 | 0.5964 | |
| Parameters/Stations | Mokengui | Epena | Dzeke | Moungouma-Bailly | Botongo | Mossengue | Bouanela |
|---|---|---|---|---|---|---|---|
| Width (m) | 42.32 ± 21.4 | 42.32 ± 19.23 | 42.30 ± 19.2 | 40.01 ± 21.28 | 43.15 ± 20.99 | 42.36 ± 21.55 | 45.36 ± 23.01 |
| Depth (m) | 3.43 ± 0.73 | 3.43 ± 0.73 | 3.43 ± 0.73 | 3.25 ± 0.75 | 3.26 ±0.73 | 3.24 ± 0.76 | 3.30 ± 0.78 |
| Transparency-Secchi (cm) | 88.70 ± 12.94 | 66.56 ± 8.50 | 66.51 ± 8.65 | 64.74 ± 10.15 | 65.69 ± 10.57 | 66.76 ± 11.63 | 68.48 ± 13.76 |
| Speed (m/s) | 0.23 ± 0.08 | 0.23 ± 0.08 | 0.23 ± 0.08 | 0.20 ± 0.08 | 0.2 ± 0.09 | 0.19 ± 0.1 | 0.16 ± 0.1 |
| Flow (m3/s) | 24.04 ± 16.78 | 24.04 ±15.54 | 24.03 ± 15.52 | 17.81 ± 13.08 | 19 ± 13.56 | 17.98 ± 14.05 | 17.30 ± 14.52 |
| Temperature (°C) | 25.82 ± 2.61 | 28.13 ± 1.17 | 28.11 ± 1.88 | 28.86 ± 1.62 | 28.79 ± 1.7 | 28.64 ± 1.87 | 28.69 ± 2.21 |
| pH | 4.2 ± 0.42 | 4.53 ± 0.14 | 4.53 ± 0.19 | 4.48 ± 0.16 | 4.46 ± 0.16 | 4.42 ± 0.15 | 4.35 ± 0.13 |
| Conductivity at 25 °C (μS/cm) | 90.40 ± 27.65 | 68.16 ±23.45 | 68.12 ± 23.46 | 61.49 ± 22.58 | 57.65 ± 21.05 | 58.52 ± 21.75 | 58.20 ± 24.50 |
| Water color observed with the naked eye | Very high yellow (golden) | High yellow (golden) | Average yellow (bright) | Average yellow (bright) | Average yellow (bright) | Average yellow (almost clear) | Average yellow (almost clear) |
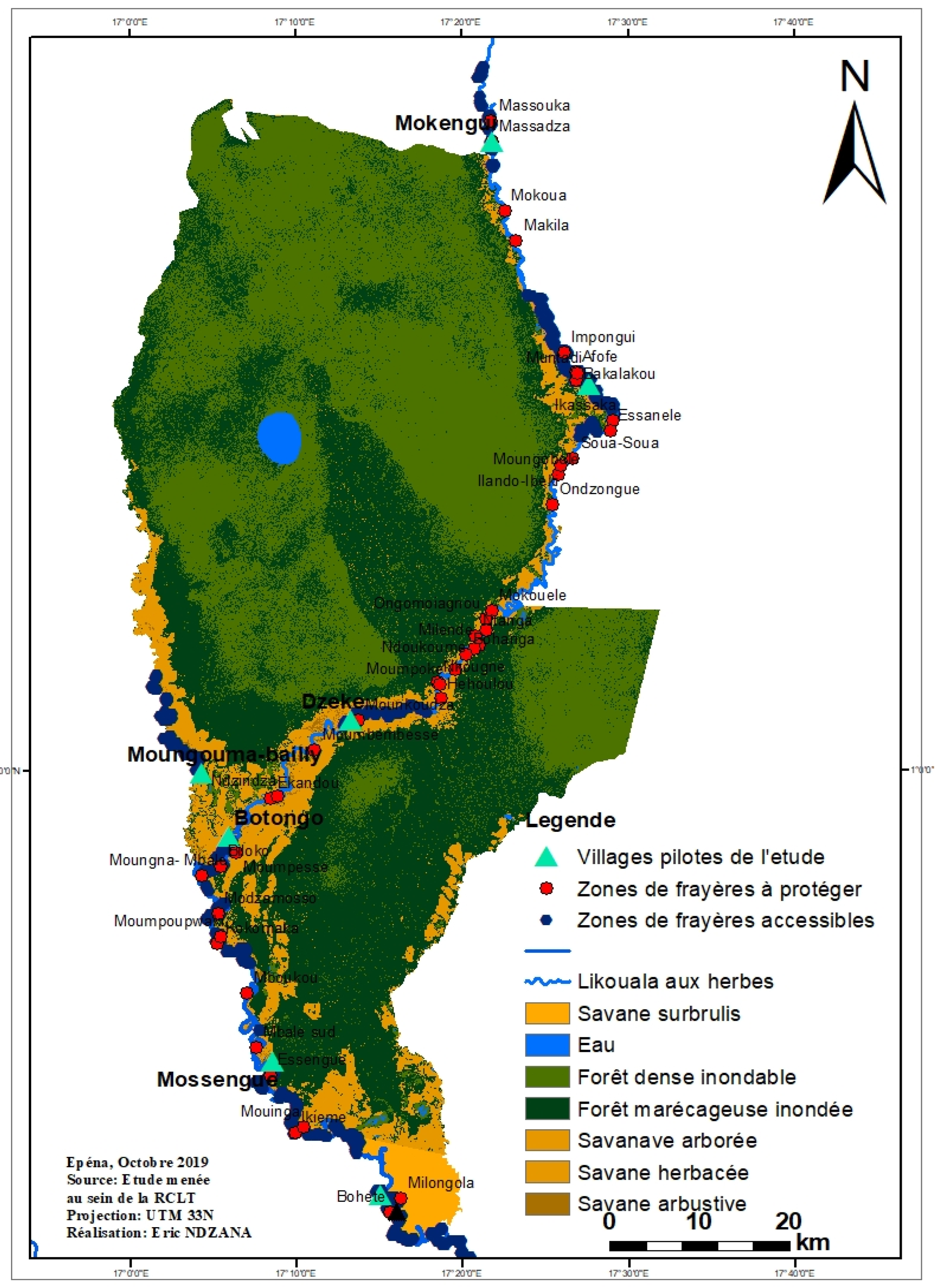
| Land cover classes | Gallery forest | Gallery forest, flooded grassy savannah | River (course bed and banks) | River, flooded grassy savannah, gallery forest | River, backwaters | River, flooded grassy savannah, backwaters | River, flooded grassy savannah, backwaters |
| Substrates of spawning grounds | Mud, sand, clay, mud and silt | Mud, sand, clay, sandy mud, foliage and macrophytes | Mud, sand, clay, sandy mud, foliage and macrophytes | Mud, sand, foliage and macrophytes mud | Mud, sand | Foliage and macrophytes mud, mud, sand, sandy silt | Sand, silt, Sand, silt and silt, foliage and macrophytes |
| Other information on the surrounding vegetation observed in the spawning area | Variety of forest species with a dominance of the species with local name “Môtôlô” | Variety of forest species and grassy formations with a dominance of grasses | Variety of partially burned forest formations with a dominance of helophytes | Variety of grassy formations with a dominance of helophytes Hydrophytes and submerged macrophytes | Variety of savannah formations with a dominance helophytes and floating hydrophytes (reeds) | Variety of partially burned savannah formations with a dominance Macrophytes Helophytes and floating hydrophytes (reeds) | Variety of partially burned savannah formations with a dominance macrophytes helophytes floating hydrophytes, (reeds) |
| Accessible spawning grounds | 11 | 28 | 16 | 17 | 24 | 28 | 27 |
| Sum of water areas of spawning grounds in (m2) | 497,998.07 | 2,512,671.414 | 502,085.049 | 848,104.1616 | 1,559,800.586 | 1,094,650.936 | 977,408.9037 |
| Sum of water areas of spawning grounds in (ha) | 4.9 | 25.12 | 5.02 | 8.4 | 15.59 | 10.94 | 9.7 |
4. Discussion
5. Conclusions
- To obtain primary knowledge of the dynamics of land use around the basin of the Likouala-aux-Herbes River in the LTCR. The LTCR is deteriorating under the probable effect of climate change, but also, and above all, due to human actions, which each year burn the floodplain and the gallery forest, degrading the potential spawning grounds;
- To obtain the distribution dynamics of fish species populations in the LTCR during the period from July to September 2019;
- To locate and characterize the 204 spawning grounds of the Likouala-aux-Herbes fisheries.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centre National de la Statistique et des Etudes Economiques (CNSEE); ICF International. Enquête Démographique Et De Santé du Congo (EDSC-II) 2011–2012; Ministère de l’Économie, des Finances, du Plan, du Portefeuille Public et de l’Intégration: Brazzaville, Congo, 2013; 412p. [Google Scholar]
- Mazenot, G. 1966. L’occupation du Bassin de la Likouala-Mossaka. 1909–1914. Cahiers d’Etudes Africaines 6, 213. Available online: https://www.google.com/url?sa=t&source=web&rct=j&opi=89978449&url=https://www.persee.fr/doc/cea_0008-0055_1966_num_6_22_3067&ved=2ahUKEwj546z8uLKFAxWWU6QEHWp8B6sQFnoECA8QAQ&usg=AOvVaw195lfC16GEb86xQQrjyyVY (accessed on 30 January 2024).
- Pouyaud, B. La Likouala Aux Herbes À Epéna Pk 530, À Botouali Pk 75; Office de la Recherche Scientifique et Technique d’Outre-Mer, Service Hydrologique, Centre de Brazzaville: Brazzaville, Congo, 1970; 30p. [Google Scholar]
- Pouyaud, B.; Barilly, A. Le Bassin De La Sangha: La Sangha À Ouesso, Le Dja À Fort-Soufflay; ORSTOM: Brazzaville, Congo, 1971; 25p. [Google Scholar]
- N’Kaloulou, B. Dynamique Paysanne Et Développement Rural Au Congo; L’Harmattan: Paris, France, 1984; 200p. [Google Scholar]
- Ashley, B.; Robin, A. Ecoregion 539: Upper Congo Rapids in Freshwater Ecoregion of the World; Conservation Science Program; WWF-US: Washington, DC, USA, 2008. [Google Scholar]
- Bokika, J. La Conservation Communautaire Dans Le Territoire De Bolobo: Forces Et Faiblesses [Revue De Primatologie [En Ligne]]. OpenEdition Journals. 2013. Available online: https://journals.openedition.org/primatologie/1434 (accessed on 1 October 2023).
- FAO. La Situation Mondiale De La Pêche Et De L’aquaculture: Gestion De L’effort De Pêche; FAO: Rome, Italy, 2008; 155p. [Google Scholar]
- FAO. Directives Techniques Pour Une Pêche Responsable: Les Dimensions Humaines De L’approche Écosystémique Des Pêches; FAO: Rome, Italy, 2010; 156p. [Google Scholar]
- FAO. La Situation Mondiale De La Pêche Et De L’aquaculture: Atteindre Les Objectifs Du Développement Durable; Rome CC BY-NC-SA 3.0 IGO; FAO: Rome, Italy, 2018; 301p. [Google Scholar]
- Welcomme, R.L. Review of the present state of knowledge of fish stocks and fisheries of African Rivers. Proc. Int. River Symp. Can. Spec. Publ. Fish. Aquat. Sci. 1989, 105, 515–532. [Google Scholar]
- Zebaze Togouet, S.H.; Njine, T.; Kemka, N.; Foto Menbohan, S.; Niyitegeka, D.; Ngassam, P.; Boutin, C. Composition et distribution spatio-temporelle des protozoaires ciliés dans un petit lac hyper trophique du Cameroun (Afrique Centrale). Rev. Sci. Eau 2006, 19, 151–162. [Google Scholar]
- Lévêque, C.; Paugy, D.; Teugels, G.G.; Musée Royal de l’Afrique Centrale (Eds.) Faune des poissons d’eaux douces et saumâtres d’Afrique de l’Ouest. In The Fresh and Brackish Water Fishes of West Africa; Musée Royal de l’Afrique Centrale [u.a.]; Publications Scientifiques du Muséum: Paris, France, 1990. [Google Scholar]
- Poll, M.; Gosse, J.P. Généra Des Poissons d’Eau Douce De l’Afrique; Académie Royale de Belgique: Bruxelles, Belgium, 1995. [Google Scholar]
- Lévêque, C.D. La pêche. In Les Poissons Des Eaux Continentales Africaines: Diversité, Ecologie, Utilisation par l’Homme; Lévêque, C., Paugy, D., Eds.; IDR: Paris, France, 1999; pp. 385–424. [Google Scholar]
- Paugy, D.; Lévêque, C.; Teugels, G.G. Poissons d’Eaux Douces et Saumâtres de l’Afrique de l’Ouest (Tome I et II); IRD, MNHN et MRAC: Paris, France, 2003. [Google Scholar]
- Fermon, Y.; Friel, J.; Ng, H.H.; De Weirdt, D. Les Mochokidae. In Faune des Poissons des Eaux Douces de Basse Guinée; IRD; MNHN; MRAC; Faune et Flore Tropicales; 42, Paris, France; 2007; pp. 698–752. Available online: https://fishbase.mnhn.fr/References/FBRefSummary.php?id=81251&database=FB&lang=farsi (accessed on 22 December 2023).
- Stiassny ML, J.; Teugels, G.G.; Hopkins, C.D. Poissons d’eaux douces et saumâtres de basse Guinée, ouest de l’Afrique centrale. In The Fresh and Brackish Water Fishes of Lower Guinea, West-Central Africa; Publications Scientifiques du Muséum: Paris, France, 2007; Volume 1. [Google Scholar]
- Teugels, G.; Mélanie, L.J.; Hopkins, C.D. Poissons D’eaux Douces et Saumâtres de Basse Guinée, Ouest de l’Afrique Centrale, Paris; Coll. Faune et Flore 42; Musée Royal de l’Afrique Centrale, Tervuren, IRD et MNHN: Paris, France, 2007; pp. 512–530. [Google Scholar]
- Apostolopoulos, D.; Nikolakopoulos, K. A review and meta-analysis of remote sensing data, GIS methods, materials and indices used for monitoring the coastline evolution over the last twenty years. Eur. J. Remote Sens. 2021, 54, 240–265. [Google Scholar] [CrossRef]
- Lindquist, E.J.; D’Annunzio, R.; Gerrand, A.; Achard, F.; Beuchle, R.; Brink, A.; Eva, H.D.; Mayaux, P.; Stibig, H.-J.; FAO et CCR. Changement d’utilisation des terres forestières mondiales 1990–2005. In Étude FAO: Forêts No 169, Organisation des Nations Unies pour L’alimentation et L’agriculture et Centre Commun de Recherchede la Commission Européenne; FAO: Rome, Italy, 2012. [Google Scholar]
- Ba, T.; Seck, D. Dynamique De L’Occupation des Sols, Cartographie Des CLPA, Des Zones De Pêche Et Mise En Place D’Un Système D’Information Geographique; Centre de Suivi Ecologique et USAID/COMFISH Project, Senegal; University of Rhode Island: Narragansett, RI, USA, 2012; 66p. [Google Scholar]
- Koumoi, Z.; Alassane, A.; Djangbedja, M.; Boukpessi, T.; Kouya, A.-E. Date Dynamique spatio-temporelle de l’occupation du sol dans le centre-togo. In Ahoho—Revue de Géographie du LARDYMES; Université de Lomé: Lomé, Togo, 2013; Volume 7, pp. 163–172. [Google Scholar]
- Matunguru, J. Perceptions de la Surexploitation Halieutique et des Stratégies de Gestion de la Pêche par les Pêcheurs du lac Edouard, Parc National des Virunga: Cas des Pêcheries de Vitshumbi, Kyavinyonge, Lunyasenge et Kisaka. (République Démocratique du Congo) [DESS en Aménagement et Gestion Intégrés des Forêts et Territoires Tropicaux]. ERAIFT/UNESCO. 2015. Available online: https://fr.scribd.com/document/663947269/Joseph-MASIRIKA-MATUNGURU-version-Finale (accessed on 22 December 2023).
- Badahoui, A.; Fiogbe, E.D.; Boko, M. Les causes de la dégradation du lac Ahémé et ses chenaux. Int. J. Biol. Chem. Sci. 2010, 4, 882–897. [Google Scholar] [CrossRef]
- Bergh, M.O.; Sowman, M. Impact Socio-Économiques Et Environnementaux De La Pêche Illégale, Non Déclarée Et Non Régle Mentaire (IUU) En Afrique, Bureau Interafricain Des Ressources Animales; Union Africaine: Addis Ababa, Ethiopia, 2016; 11p. [Google Scholar]
- Mamonekene, V. Les ressources halieutiques de la Réserve communautaire du Lac Télé Likouala aux Herbes. Diversité et exploitation. Rapp. De Consult. 2006, 25, 30. [Google Scholar]
- Mutambue, M.J.-C.E.S.; Twagirashyaka, F.; Mobongo, R.; Bakabana, P.; Mangonga, T. Plan d’aménagement et de gestion des zones humides de la réserve communautaire du lac Télé, République du Congo (PAGZHRCLT), 4-5MICHA J.-C. 2.017–Wetland Management Plan, a Hope for Sustainable Artisanal Fisheries in Protected Areas (PA)? Quel avenir pour les Aires Protégées (AP) aux zones humides oubliées en Afrique continentale. Tropicultura 2016, 35, 235–236. [Google Scholar]
- Hounsounou, L.C. Importance et Place Socioéconomique de Pêche à la Senne de Plage Dans les Moyens D’existence des Communautés de Pêche au Bénin, Cotonou 2011, 65p. Available online: https://www.researchgate.net/publication/346258010_Etude_socio-economique_de_la_peche_dans_la_partie_Sud-Ouest_du_lac_Albert_Ituri_RD_Congo (accessed on 14 November 2023).
- Hounsounou, L.C.; Sohou, Z.; Akouchou, G. Pêche à la senne de plage au Benin et durabilité des ressources halieutiques. J. Rech. Sci. Univ. Togo Série A 2013, 15, 1–13. [Google Scholar]
- Moutsambote, J.M. Rapport de Mission Botanique Effectuée au lac Télé (District d’Epéna, Likouala) 08–27 June 1992; Laboratoire de Botanique, CERVE: Brazzaville, Congo, 1992; 6p. [Google Scholar]
- Byanikiro, R.M.; Nsila, J.N.; Busanga, A.K.; Ulyelali-Patho, J.; Micha, J.C. Caractérisation et écologie des peuplements des mormyridae dans la réserve de Yoko (Kisangani, RD Congo). Int. J. Biol. Chem. Sci. 2017, 11, 967–999. [Google Scholar] [CrossRef][Green Version]
- Kamfu, P.; Bekeli, N.M.; Pigneur, L.-M.; Lomema, T.; Micha, J.-C. Environmental variables structuring Labeo species (Pisces, Cyprinidae) in Malebo Pool, Congo River. Rev. Sci. Tech. Forêt Et Environ. Du Bassin Du Congo 2011, 1, 46–55. [Google Scholar] [CrossRef][Green Version]
- Tomedi Eyango, M.; Efole ETBlé, M.C.; Ndzana, B.E.B.; Songmo, B.L.; Nyamsi Tchatcho, N.L.; Mikolasek, O.; Tchoumbougnang, F. Rizipisciculture: Voie D’intensification Écologique des Systèmes Piscicoles Extensifs. F.Tech&Doc. Vul.: 18-23-Volume Spécial; CRO, Centre de Recherche Océanographique d’Abidjan: Abidjan, Côte d’Ivoire, 2015. [Google Scholar]
- Chikou, A. Etude de la Démographie et de L’exploitation Halieutique de Six Espèces de Poissons Chats (Teleostei, Siluriformes) Dans le Delta de l’Ouémé au Bénin. [Thèse de Doctorat, Université de Liège]. 2006. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkozje))/reference/referencespapers.aspx?referenceid=2482975 (accessed on 1 October 2023).
- Kowozogono, R.K.; Ngbolua, K.N.; Lusasi, W.S.; Inkoto, C.L.; Zwa, T.G.; Iteku, J.B. Inventaire systématique des poissons frais vendus dans le marché Central de Yakoma (Province du Nord Ubangi) en République démocratique du Congo. Rev. Marocaine Des Sci. Agron. Vétérinaires 2021, 9, 730–736. Available online: https://www.agrimaroc.org/index.php/Actes_IAVH2/article/view/969/1265 (accessed on 1 October 2023).
- Omasombo Tshonda, J. République Démocratique Du Congo, MAI-NDOMBE Mosaïque De Peuples Établie Sur Un Patrimoine naturel. Africa Museum. 2019. Available online: https://www.africamuseum.be/sites/default/files/media/docs/research/publications/rmca/online/monographies-provinces/vol12_Mai_Ndombe.pdf (accessed on 1 October 2023).
- Attingli, A.H.; Ahouansou Montcho, S.; Vissin, E.W.; Zinsou, L.H.; Laleye, P.A. Influence des engins et techniques de pêche sur l’abondance relative des espèces dans la Basse Vallee de l’Ouémé au Benin. Afr. Crop Sci. J. 2017, 25, 47. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndzana Biloa, E.B.; Mamonekene, V.; Micha, J.-C. Characterization of Fish Spawning Grounds near the Likouala-Aux-Herbes River, Lac Tele Community Reserve (LTCR), Republic of Congo, for Sustainable Wetland Management. Sustainability 2024, 16, 3353. https://doi.org/10.3390/su16083353
Ndzana Biloa EB, Mamonekene V, Micha J-C. Characterization of Fish Spawning Grounds near the Likouala-Aux-Herbes River, Lac Tele Community Reserve (LTCR), Republic of Congo, for Sustainable Wetland Management. Sustainability. 2024; 16(8):3353. https://doi.org/10.3390/su16083353
Chicago/Turabian StyleNdzana Biloa, Eric Bertin, Victor Mamonekene, and Jean-Claude Micha. 2024. "Characterization of Fish Spawning Grounds near the Likouala-Aux-Herbes River, Lac Tele Community Reserve (LTCR), Republic of Congo, for Sustainable Wetland Management" Sustainability 16, no. 8: 3353. https://doi.org/10.3390/su16083353
APA StyleNdzana Biloa, E. B., Mamonekene, V., & Micha, J.-C. (2024). Characterization of Fish Spawning Grounds near the Likouala-Aux-Herbes River, Lac Tele Community Reserve (LTCR), Republic of Congo, for Sustainable Wetland Management. Sustainability, 16(8), 3353. https://doi.org/10.3390/su16083353