
Lactic Acid Bacteria in Sustainable Food Production


Abstract

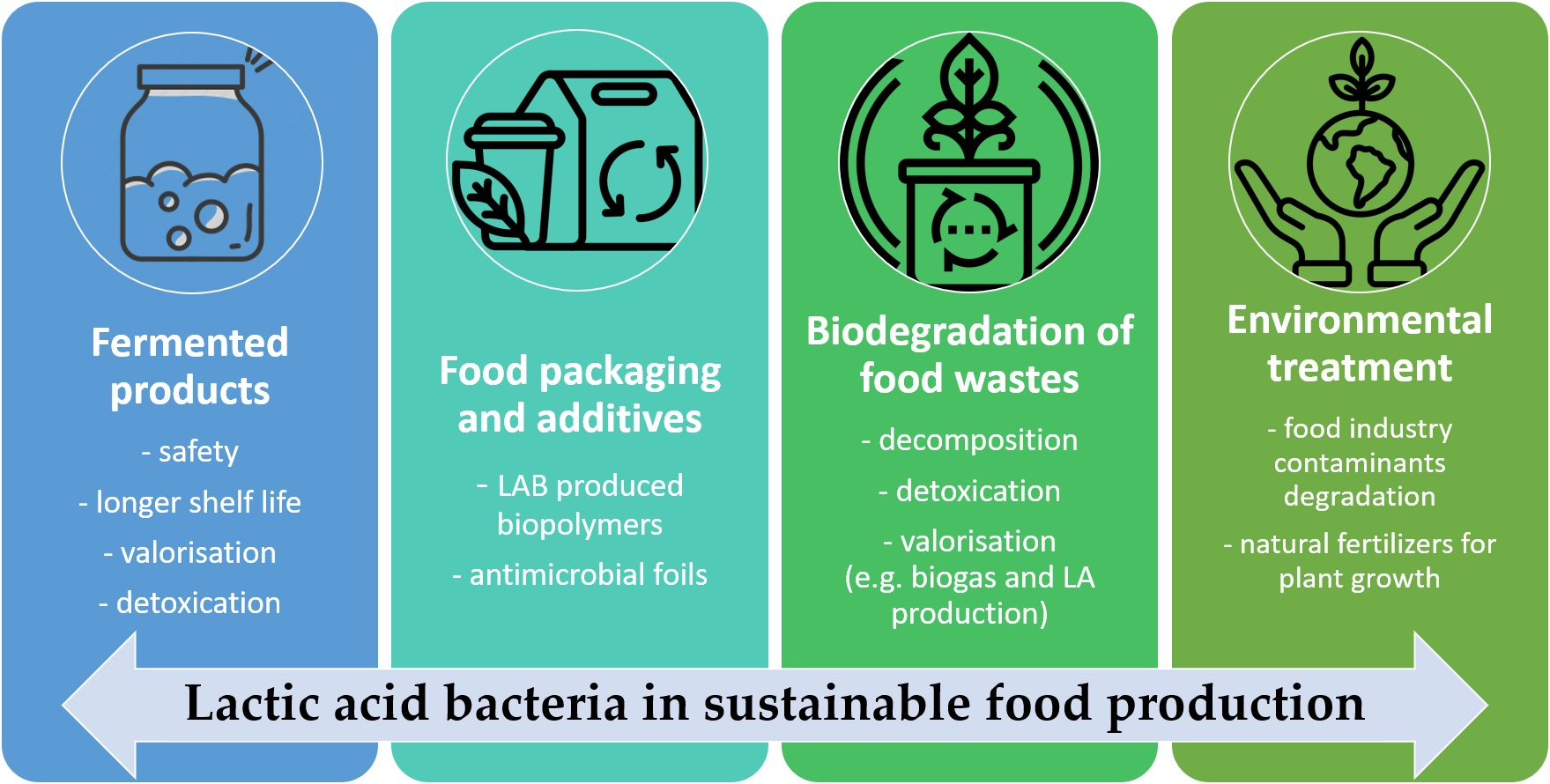{kind=link}
1. Introduction
2. The Role of Lactic Acid Bacteria in Food Fermentation
2.1. Traditional Applications of LAB in Food Fermentation
2.2. The Role of LAB in Enhancing the Shelf Life and Safety of Fermented Food Products
2.3. Enhancing Nutritional Value of Food through LAB Fermentation
2.4. LAB in Detoxification of Food Products
2.5. Innovative LAB Fermentation Methods Supporting Sustainable Food Production
3. Environmental Impact of Lactic Acid Bacteria
3.1. Biodegradation of Organic Waste
3.2. Biogas Production
3.3. Environmental Treatment and Restoration
3.4. Biopolymers Production
3.4.1. Polylactic Acid (PLA)
3.4.2. Polyhydroxyalkanoates (PHAs)
3.4.3. Exopolysaccharides (EPS)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | anaerobic digestion |
| ATCC | American Type Culture Collection |
| ATP | Adenosine triphosphate |
| ATPase | F0F1-Adenosine triphosphate Synthase |
| CH4 | Methane |
| CO2 | carbon dioxide |
| COD | chemical oxygen demand |
| DNA | Deoxyribonucleic acid |
| EPS | Exopolysaccharide |
| EU | European Union |
| FAO | The Food and Agriculture Organisation |
| FOS | Fructooligosaccharides |
| FW | food waste |
| HA | heterocyclic amines |
| LA | lactic acid |
| LAB | Lactic acid bacteria |
| MRS broth | De Man, Rogosa and Sharpe broth |
| MS | municipal sludge |
| NA | N-nitrosamine |
| NDMA | nitrosodimethylamine |
| NH3 | ammonia |
| PAH | polycyclic aromatic hydrocarbon |
| PHA | polyhydroxyalkanoate |
| PHV | polyhydroxyvalerate |
| PHVB | poly(3-hydroxybutyrate-co-3-hydroxyvalerate) |
| PLA | polylactic acid |
| ROS | reactive oxygen species |
| USD | The United States dolar |
References
- Fanzo, J.; Rudie, C.; Sigman, I.; Grinspoon, S.; Benton, T.G.; Brown, M.E.; Covic, N. Sustainable food systems and nutrition in the 21st century: A report from the 22 nd annual Harvard Nutrition Obesity Symposium. Am. J. Clin. Nutr. 2022, 115, 18–33. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H. El Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.; Kim, J.S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.J.; Kim, S.J. Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Xu, J.L.; Wang, Z. Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020; ISBN 9789811501692. [Google Scholar]
- Nicolescu, C.M.; Bumbac, M.; Buruleanu, C.L.; Popescu, E.C.; Stanescu, S.G.; Georgescu, A.A.; Toma, S.M. Biopolymers Produced by Lactic Acid Bacteria: Characterization and Food Application. Polymers 2023, 15, 1539. [Google Scholar] [CrossRef]
- Zapaśnik, A.; Sokołowska, B.; Bryła, M. Role of Lactic Acid Bacteria in Food Preservation and Safety. Foods 2022, 11, 1283. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Mitchell, R. Fermentation and pathogen control: A risk assessment approach. Int. J. Food Microbiol. 2002, 79, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Tucker, G. Pasteurization: Principles and Applications, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9780123849533. [Google Scholar]
- Tassou, C.C.; Panagou, E.Z.; Katsaboxakis, K.Z. Microbiological and physicochemical changes of naturally black olives fermented at different temperatures and NaCl levels in the brines. Food Microbiol. 2002, 19, 605–615. [Google Scholar] [CrossRef]
- Haque, A.; Richardson, R.K.; Morris, E.R. Effect of fermentation temperature on the rheology of set and stirred yogurt. Food Hydrocoll. 2001, 15, 593–602. [Google Scholar] [CrossRef]
- Chung, H.C.; Jeong, B.Y.; Han, G.D. Optimum conditions for combined application of Leuconostoc sp. and Saccharomyces sp. to sourdough. Food Sci. Biotechnol. 2011, 20, 1373–1379. [Google Scholar] [CrossRef]
- Wouters, D.; Bernaert, N.; Anno, N.; Van Droogenbroeck, B.; De Loose, M.; Van Bockstaele, E.; De Vuyst, L. Application’ and validation of autochthonous lactic acid bacteria starter cultures for controlled leek fermentations and their influence on the antioxidant properties of leek. Int. J. Food Microbiol. 2013, 165, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Motarjemi, Y. Impact of small scale fermentation technology on food safety in developing countries. Int. J. Food Microbiol. 2002, 75, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Wirawati, C.U.; Sudarwanto, M.B.; Lukman, D.W.; Wientarsih, I.; Srihanto, E.A. Diversity of lactic acid bacteria in dadih produced by either back-slopping or spontaneous fermentation from two different regions of West Sumatra, Indonesia. Vet. World 2019, 12, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, N.; Ananthanarayan, L. Use of the backslopping method for accelerated and nutritionally enriched idli fermentation. J. Sci. Food Agric. 2015, 95, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Budzyńska, A.; Grudlewska-Buda, K.; Wiktorczyk-Kapischke, N.; Andrzejewska, M.; Wałecka-Zacharska, E.; Gospodarek-Komkowska, E. Two Faces of Fermented Foods—The Benefits and Threats of Its Consumption. Front. Microbiol. 2022, 13, 845166. [Google Scholar] [CrossRef] [PubMed]
- García-Díez, J.; Saraiva, C. Use of starter cultures in foods from animal origin to improve their safety. Int. J. Environ. Res. Public Health 2021, 18, 2544. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of starter cultures on the safety of fermented meat products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef]
- Yu, Z.; Peng, C.; Kwok, L.Y.; Zhang, H. The bacterial diversity of spontaneously fermented dairy products collected in northeast asia. Foods 2021, 10, 2321. [Google Scholar] [CrossRef]
- Malo, P.M.; Urquhart, E.A. Fermented Foods: Use of Starter Cultures. In Encyclopedia of Food and Health; Academic Press: Cambridge, MA, USA, 2015; pp. 681–685. ISBN 9780123849533. [Google Scholar]
- Picon, A. Cheese microbial ecology and safety. In Global Cheesemaking Technology: Cheese Quality and Characteristics; Wiley-IEEE Press: Hoboken, NJ, USA, 2017; pp. 71–99. ISBN 9781119046165. [Google Scholar]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Aghababaie, M.; Khanahmadi, M.; Beheshti, M. Developing a kinetic model for co-culture of yogurt starter bacteria growth in pH controlled batch fermentation. J. Food Eng. 2015, 166, 72–79. [Google Scholar] [CrossRef]
- Frantzen, C.A.; Kleppen, H.P.; Holo, H. Lactococcus lactis diversity in undefined mixed dairy starter cultures as revealed by comparative genome analyses and targeted amplicon sequencing of epsD. Appl. Environ. Microbiol. 2018, 84, e02199-17. [Google Scholar] [CrossRef] [PubMed]
- Pavunc, A.L.; Beganović, J.; Kos, B.; Uroić, K.; Blàžić, M.; Šušković, J. Characterization and application of autochthonous starter cultures for fresh cheese production. Food Technol. Biotechnol. 2012, 50, 141–151. [Google Scholar]
- Xiong, T.; Li, X.; Guan, Q.; Peng, F.; Xie, M. Starter culture fermentation of Chinese sauerkraut: Growth, acidification and metabolic analyses. Food Control 2014, 41, 122–127. [Google Scholar] [CrossRef]
- Mun, S.Y.; Chang, H.C. Characterization of Weissella koreensis sk isolated from kimchi fermented at low temperature (Around 0 °C) based on complete genome sequence and corresponding phenotype. Microorganisms 2020, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; García-Vaquero, M. Bioactive Compounds from Fermented Food Products. In Novel Food Fermentation Technologies; Food Engineering Series; Springer: Berlin/Heidelberg, Germany, 2016; pp. 293–310. [Google Scholar]
- Ferranti, P.; Nitride, C.; Nicolai, M.A.; Mamone, G.; Picariello, G.; Bordoni, A.; Valli, V.; Di Nunzio, M.; Babini, E.; Marcolini, E.; et al. In vitro digestion of Bresaola proteins and release of potential bioactive peptides. Food Res. Int. 2014, 63, 157–169. [Google Scholar] [CrossRef]
- Di Cagno, R.; Chaves Lòpez, C.; Tofalo, R.; Gallo, G.; De Angelis, M.; Paparella, A.; Hammes, W.P.; Gobbetti, M. Comparison of the compositional, microbiological, biochemical and volatile profile characteristics of three Italian PDO fermented sausages. Meat Sci. 2008, 79, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Bis-Souza, C.V.; Penna, A.L.B.; da Silva Barretto, A.C. Applicability of potentially probiotic Lactobacillus casei in low-fat Italian type salami with added fructooligosaccharides: In vitro screening and technological evaluation. Meat Sci. 2020, 168, 108186. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Yang, X.; Zhang, K.; Zhao, Y.; Cheng, F.; Jin, Y. Influence of Lactobacillus helveticus ZF22 and TR1-1-3 strains on the aromatic flavor of fermented sausages. Front. Nutr. 2023, 9, 1058109. [Google Scholar] [CrossRef]
- Ammor, M.S.; Gueimonde, M.; Danielsen, M.; Zagorec, M.; Van Hoek, A.H.A.M.; De Los Reyes-Gavilán, C.G.; Mayo, B.; Margolles, A. Two different tetracycline resistance mechanisms, plasmid-carried tet(L) and chromosomally located transposon-associated tet(M), coexist in Lactobacillus sakei rits 9. Appl. Environ. Microbiol. 2008, 74, 1394–1401. [Google Scholar] [CrossRef]
- Mozaffari, S.; Abedi, K.A.D.; Garmaroudi, A.A. Assessing The Emergency Preparedness of The Educational Hospitals in Kurdistan’s University of Medical Sciences, using The Resilience Engineering Method In 2018. Prev. Nutr. Food Sci. 2021, 26, 476–484. [Google Scholar]
- Sionek, B.; Szydłowska, A.; Küçükgöz, K.; Kołożyn-Krajewska, D. Traditional and New Microorganisms in Lactic Acid Fermentation of Food. Fermentation 2023, 9, 1019. [Google Scholar] [CrossRef]
- Salvetti, E.; Fondi, M.; Fani, R.; Torriani, S.; Felis, G.E. Evolution of lactic acid bacteria in the order lactobacillales as depicted by analysis of glycolysis and pentose phosphate pathways. Syst. Appl. Microbiol. 2013, 36, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Šušković, J.; Kos, B.; Beganović, J.; Pavunc, A.L.; Habjanič, K.; Matoć, S. Antimicrobial activity—The most important property of probiotic and starter lactic acid bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Peetermans, A.; Foulquié-Moreno, M.R.; Thevelein, J.M. Mechanisms underlying lactic acid tolerance and its influence on lactic acid production in Saccharomyces cerevisiae. Microb. Cell 2021, 8, 111–130. [Google Scholar] [CrossRef]
- Kawazoe, N.; Kimata, Y.; Izawa, S. Acetic acid causes endoplasmic reticulum stress and induces the unfolded protein response in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1192. [Google Scholar] [CrossRef]
- Rallu, F.; Gruss, A.; Maguin, E. Lactococcus lactis and stress. Antonie Van Leeuwenhoek 1996, 70, 243–251. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ohishi, H.; Benno, Y. H+-ATPase activity in Bifidobacterium with special reference to acid tolerance. Int. J. Food Microbiol. 2004, 93, 109–113. [Google Scholar] [CrossRef]
- Lowe, D.P.; Arendt, E.K.; Soriano, A.M.; Ulmer, H.M. The influence of lactic acid bacteria on the quality of malt. J. Inst. Brew. 2005, 111, 42–50. [Google Scholar] [CrossRef]
- Zhang, C.; Brandt, M.J.; Schwab, C.; Gänzle, M.G. Propionic acid production by cofermentation of Lactobacillus buchneri and Lactobacillus diolivorans in sourdough. Food Microbiol. 2010, 27, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Johanningsmeier, S.D.; McFeeters, R.F. Metabolism of lactic acid in fermented cucumbers by Lactobacillus buchneri and related species, potential spoilage organisms in reduced salt fermentations. Food Microbiol. 2013, 35, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, Q.; Xu, J.; Sun, H.; Cheng, Q.; Xie, Y.; Wang, C.; Li, P.; Chen, C.; Zheng, Y. Effect of different organic acid additives on the fermentation quality and bacterial community of paper mulberry (Broussonetia papyrifera) silage. Front. Microbiol. 2022, 13, 1038549. [Google Scholar] [CrossRef] [PubMed]
- Punia Bangar, S.; Suri, S.; Trif, M.; Ozogul, F. Organic acids production from lactic acid bacteria: A preservation approach. Food Biosci. 2022, 46, 101615. [Google Scholar] [CrossRef]
- Sorrells, K.M.; Enigl, D.C. Effect of pH, acidulant, sodium chloride, and temperature on the growth of Listeria monocytogenes. J. Food Saf. 1990, 11, 31–37. [Google Scholar] [CrossRef]
- Al-Rousan, W.M.; Olaimat, A.N.; Osaili, T.M.; Al-Nabulsi, A.A.; Ajo, R.Y.; Holley, R.A. Use of acetic and citric acids to inhibit Escherichia coli O157: H7, Salmonella Typhimurium and Staphylococcus aureus in tabbouleh salad. Food Microb. 2018, 73, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Zhang, X.; Xiao, Y.; Sheng, Q.; Tu, L.; Chen, F.; Yan, Y.; Zheng, Y.; Wang, M. Interaction of acetic acid bacteria and lactic acid bacteria in multispecies solid-state fermentation of traditional Chinese cereal vinegar. Front. Microbiol. 2022, 13, 964855. [Google Scholar] [CrossRef] [PubMed]
- Özcelik, S.; Kuley, E.; Özogul, F. Formation of lactic, acetic, succinic, propionic, formic and butyric acid by lactic acid bacteria. LWT 2016, 73, 536–542. [Google Scholar] [CrossRef]
- Vollenweider, S.; Grassi, G.; König, I.; Puhan, Z. Purification and structural characterization of 3-hydroxypropionaldehyde and its derivatives. J. Agric. Food Chem. 2003, 51, 3287–3293. [Google Scholar] [CrossRef]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The antimicrobial compound reuterin (3-hydroxypropionaldehyde) induces oxidative stress via interaction with thiol groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Langa, S.; Martín-Cabrejas, I.; Montiel, R.; Landete, J.M.; Medina, M.; Arqués, J.L. Short communication: Combined antimicrobial activity of reuterin and diacetyl against foodborne pathogens. J. Dairy Sci. 2014, 97, 6116–6121. [Google Scholar] [CrossRef]
- Gómez-Torres, N.; Ávila, M.; Gaya, P.; Garde, S. Prevention of late blowing defect by reuterin produced in cheese by a Lactobacillus reuteri adjunct. Food Microbiol. 2014, 42, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Muthukumarasamy, P.; Holley, R.A. Survival of Escherichia coli O157:H7 in dry fermented sausages containing micro-encapsulated probiotic lactic acid bacteria. Food Microbiol. 2007, 24, 82–88. [Google Scholar] [CrossRef]
- Bachmann, H.; Pronk, J.T.; Kleerebezem, M.; Teusink, B. Evolutionary engineering to enhance starter culture performance in food fermentations. Curr. Opin. Biotechnol. 2015, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Grosu-Tudor, S.S.; Stancu, M.M.; Pelinescu, D.; Zamfir, M. Characterization of some bacteriocins produced by lactic acid bacteria isolated from fermented foods. World J. Microbiol. Biotechnol. 2014, 30, 2459–2469. [Google Scholar] [CrossRef]
- Devi, S.M.; Ramaswamy, A.M.; Halami, P.M. In situ production of pediocin PA-1 like bacteriocin by different genera of lactic acid bacteria in soymilk fermentation and evaluation of sensory properties of the fermented soy curd. J. Food Sci. Technol. 2014, 51, 3325–3332. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Lucena-Padrós, H.; Ruiz-Barba, J.L. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons. Food Microbiol. 2013, 33, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Lee, H.J.; Chang, H.C. Identification of the agent from Lactobacillus plantarum KFRI464 that enhances bacteriocin production by Leuconostoc citreum GJ7. J. Appl. Microbiol. 2007, 103, 2504–2515. [Google Scholar] [CrossRef]
- Abitayeva, G.K.; Urazova, M.S.; Abilkhadirov, A.S.; Sarmurzina, Z.S.; Shaikhin, S.M. Characterization of a new bacteriocin-like inhibitory peptide produced by Lactobacillus sakei B-RKM 0559. Biotechnol. Lett. 2021, 43, 2243–2257. [Google Scholar] [CrossRef]
- Fu, Y.; Zhao, D.; Wang, L.; Jiang, G.; Liu, X. A broad-spectrum novel bacteriocin produced by Lactobacillus sakei in Nanjing Steamed Roast Duck: Purification, antimicrobial characteristics, and antibacterial mechanisms. Food Biosci. 2022, 50, 101995. [Google Scholar] [CrossRef]
- Contessa, C.R.; de Souza, N.B.; Gonçalo, G.B.; de Moura, C.M.; da Rosa, G.S.; Moraes, C.C. Development of active packaging based on agar-agar incorporated with bacteriocin of lactobacillus sakei. Biomolecules 2021, 11, 1869. [Google Scholar] [CrossRef] [PubMed]
- Kiczorowski, P.; Kiczorowska, B.; Samolińska, W.; Szmigielski, M.; Winiarska-Mieczan, A. Effect of fermentation of chosen vegetables on the nutrient, mineral, and biocomponent profile in human and animal nutrition. Sci. Rep. 2022, 12, 13422. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Leeuwendaal, N.K.; Stanton, C.; O’toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough microbiome comparison and benefits. Microorganisms 2021, 9, 1355. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.H.; Huang, L.Y.; Terefe, N.S.; Augustin, M.A. Fermentation-based biotransformation of glucosinolates, phenolics and sugars in retorted broccoli puree by lactic acid bacteria. Food Chem. 2019, 286, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Mohammadi-Kouchesfahani, M.; Hamidi-Esfahani, Z.; Azizi, M.H. Isolation and identification of lactic acid bacteria with phytase activity from sourdough. Food Sci. Nutr. 2019, 7, 3700–3708. [Google Scholar] [CrossRef]
- Rosa-Sibakov, N.; Re, M.; Karsma, A.; Laitila, A.; Nordlund, E. Phytic Acid Reduction by Bioprocessing as a Tool to Improve the in Vitro Digestibility of Faba Bean Protein. J. Agric. Food Chem. 2018, 66, 10394–10399. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubbaiah, R.; Rajesh, S.K. Food Processing and Fermentation Studies on Reduction of Phytic Acid in Triticum Aestivum and Sorghum Bicolor (L.). J. Microbiol. Biotechnol. Food Sci. 2020, 10, 166–169. [Google Scholar] [CrossRef]
- Onipede, G.O.; Banwo, K.; Ogunremi, O.R.; Sanni, A.; Polytechnic, T.; State, O. Influence of starter culture lactic acid bacteria on the phytic acid content of sorghum-ogi (an indigenous cereal gruel). Ann. Food Sci. Technol. 2014, 15, 121–134. [Google Scholar]
- Scheers, N.; Rossander-Hulthen, L.; Torsdottir, I.; Sandberg, A.S. Increased iron bioavailability from lactic-fermented vegetables is likely an effect of promoting the formation of ferric iron (Fe3+). Eur. J. Nutr. 2016, 55, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lai, S.; Zhou, Z.; Yang, J.; Liu, H.; Zhong, Z.; Fu, H.; Ren, Z.; Shen, L.; Cao, S.; et al. Screening and evaluation of lactic acid bacteria with probiotic potential from local Holstein raw milk. Front. Microbiol. 2022, 13, 918774. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Torres, L.; Yépez, A.; Aznar, R.; Arahal, D.R. Genomic Insights Into Five Strains of Lactobacillus plantarum With Biotechnological Potential Isolated From chicha, a Traditional Maize-Based Fermented Beverage From Northwestern Argentina. Front. Microbiol. 2019, 10, 2232. [Google Scholar] [CrossRef] [PubMed]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Sarkar, D.; Shetty, K. Fermentation-based biotransformation of bioactive phenolics and volatile compounds from cashew apple juice by select lactic acid bacteria. Process Biochem. 2017, 59, 141–149. [Google Scholar] [CrossRef]
- Llamas-Arriba, M.G.; Hernández-Alcántara, A.M.; Mohedano, M.L.; Chiva, R.; Celador-Lera, L.; Velázquez, E.; Prieto, A.; Dueñas, M.T.; Tamame, M.; López, P. Lactic acid bacteria isolated from fermented doughs in Spain produce dextrans and riboflavin. Foods 2021, 10, 2004. [Google Scholar] [CrossRef]
- Juvonen, R.; Honkapää, K.; Maina, N.H.; Shi, Q.; Viljanen, K.; Maaheimo, H.; Virkki, L.; Tenkanen, M.; Lantto, R. The impact of fermentation with exopolysaccharide producing lactic acid bacteria on rheological, chemical and sensory properties of pureed carrots (Daucus carota L.). Int. J. Food Microbiol. 2015, 207, 109–118. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef] [PubMed]
- Pasolli, E.; De Filippis, F.; Mauriello, I.E.; Cumbo, F.; Walsh, A.M.; Leech, J.; Cotter, P.D.; Segata, N.; Ercolini, D. Large-scale genome-wide analysis links lactic acid bacteria from food with the gut microbiome. Nat. Commun. 2020, 11, 2610. [Google Scholar] [CrossRef] [PubMed]
- Mörschbächer, A.P.; Pappen, E.; Henriques, J.A.P.; Granada, C.E. Effects of probiotic supplementation on the gut microbiota composition of adults: A systematic review of randomized clinical trials. An. Acad. Bras. Cienc. 2023, 95, e20230037. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Xu, B.; Chen, C.; Li, P.; Luo, H. The function and mechanism of lactic acid bacteria in the reduction of toxic substances in food: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 5950–5963. [Google Scholar] [CrossRef] [PubMed]
- Chiocchetti, G.M.; Jadán-Piedra, C.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. Use of lactic acid bacteria and yeasts to reduce exposure to chemical food contaminants and toxicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kang, K.H.; Kim, S.H.; Lee, S.; Lee, S.H.; Ha, E.S.; Sung, N.J.; Kim, J.G.; Chung, M.J. Lactic acid bacteria directly degrade N-nitrosodimethylamine and increase the nitrite-scavenging ability in kimchi. Food Control 2017, 71, 101–109. [Google Scholar] [CrossRef]
- Nowak, A.; Kuberski, S.; Libudzisz, Z. Probiotic lactic acid bacteria detoxify N-nitrosodimethylamine. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2014, 31, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Liao, E.; Xu, Y.; Jiang, Q.; Xia, W. Effects of inoculating autochthonous starter cultures on N-nitrosodimethylamine and its precursors formation during fermentation of Chinese traditional fermented fish. Food Chem. 2019, 271, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.B.; Ohta, Y. Binding of Mutagens by Fractions of the Cell Wall Skeleton of Lactic Acid Bacteria on Mutagens. J. Dairy Sci. 1991, 74, 1477–1481. [Google Scholar] [CrossRef]
- Stidl, R.; Sontag, G.; Koller, V.; Knasmüller, S. Binding of heterocyclic aromatic amines by lactic acid bacteria: Results of a comprehensive screening trial. Mol. Nutr. Food Res. 2008, 52, 322–329. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Mozuriene, E.; Krungleviciute, V.; Novoslavskij, A.; Santini, A.; Rozentale, I.; Juodeikiene, G.; Cizeikiene, D. The impact of lactic acid bacteria with antimicrobial properties on biodegradation of polycyclic aromatic hydrocarbons and biogenic amines in cold smoked pork sausages. Food Control 2017, 71, 285–292. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Krungleviciute, V.; Pugajeva, I.; Zadeike, D.; Juodeikiene, G.; Cizeikiene, D. The Influence of Scalded Flour, Fermentation, and Plants Belonging to Lamiaceae Family on the Wheat Bread Quality and Acrylamide Content. J. Food Sci. 2018, 83, 1560–1568. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, F.; Qi, Y.; Dziugan, P.; Bai, F.; Walczak, P.; Zhang, B. Screening of Lactobacillus strains for their ability to bind Benzo(a)pyrene and the mechanism of the process. Food Chem. Toxicol. 2013, 59, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.R.; Yu, R.C.; Chou, C.C.; Huang, E.C. Determinations of the antimutagenic activities of several probiotic bifidobacteria under acidic and bile conditions against benzo[a]pyrene by a modified Ames test. Int. J. Food Microbiol. 2004, 93, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Niño, J.C.; Cavazos-Garduño, A.; Cantú-Cornelio, F.; Gonzalez-Cordova, A.F.; Vallejo-Cordoba, B.; Hernandez-Mendoza, A.; García, H.S. In vitro reduced availability of aflatoxin B1 and acrylamide by bonding interactions with teichoic acids from lactobacillus strains. LWT 2015, 64, 1334–1341. [Google Scholar] [CrossRef]
- Zhao, S.; Zhao, X.; Liu, Q.; Jiang, Y.; Li, Y.; Feng, W.; Xu, H.; Shao, M. Protective effect of Lactobacillus plantarum ATCC8014 on acrylamide-induced oxidative damage in rats. Appl. Biol. Chem. 2020, 63, 43. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, W.; Li, L.; Zhao, H.Y.; Sun, H.Y.; Meng, M.H.; Zhang, S.; Shao, M.L. Key role of peptidoglycan on acrylamide binding by lactic acid bacteria. Food Sci. Biotechnol. 2017, 26, 271–277. [Google Scholar] [CrossRef]
- Lahtinen, S.J.; Haskard, C.A.; Ouwenhand, A.C.; Salminen, S.J.; Ahokas, J.T. Binding of aflatoxin B1 to cell wall components of Lactobacillus rhamnosus strain GG. Food Addit. Contam. 2004, 21, 158–164. [Google Scholar] [CrossRef]
- Tohno, M.; Tanizawa, Y.; Sawada, H.; Sakamoto, M.; Ohkuma, M.; Kobayashi, H. A novel species of lactic acid bacteria, Ligilactobacillus pabuli sp. nov., isolated from alfalfa silage. Int. J. Syst. Evol. Microbiol. 2022, 72, 5587. [Google Scholar] [CrossRef]
- Chen, Y.S.; Wang, L.T.; Wu, Y.C.; Mori, K.; Tamura, T.; Chang, C.H.; Chang, Y.C.; Wu, H.C.; Yi, H.H.; Wang, P.Y. Leuconostoc litchii sp. Nov., a novel lactic acid bacterium isolated from lychee. Int. J. Syst. Evol. Microbiol. 2020, 70, 1585–1590. [Google Scholar] [CrossRef]
- Kouya, T.; Ishiyama, Y.; Ohashi, S.; Kumakubo, R.; Yamazaki, T.; Otaki, T. Philodulcilactobacillus myokoensis gen. nov., sp. nov., a fructophilic, acidophilic, and agar-phobic lactic acid bacterium isolated from fermented vegetable extracts. PLoS ONE 2023, 18, e0286677. [Google Scholar] [CrossRef]
- Terpou, A.; Mantzourani, I.; Galanis, A.; Kanellaki, M.; Bezirtzoglou, E.; Bekatorou, A.; Koutinas, A.A.; Plessas, S. Employment of l. Paracasei k5 as a novel potentially probiotic freeze-dried starter for feta-type cheese production. Microorganisms 2019, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Sarengaole, B.; Tsend-Ayush, C.; Bayinjirigala, J.; Menghebilige, B. Development technology of starter cultures using lactic acid bacteria isolated from fermented Camel milk with cholesterol lowering ability. Mong. J. Chem. 2022, 23, 38–50. [Google Scholar] [CrossRef]
- Garofalo, C.; Norici, A.; Mollo, L.; Osimani, A.; Aquilanti, L. Fermentation of Microalgal Biomass for Innovative Food Production. Microorganisms 2022, 10, 2069. [Google Scholar] [CrossRef] [PubMed]
- Parada, J.L.; De Caire, G.Z.; De Mulé, M.C.Z.; De Cano, M.M.S. Lactic acid bacteria growth promoters from Spirulina platensis. Int. J. Food Microbiol. 1998, 45, 225–228. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H.ü. Influence of incorporated Spirulina platensis on the growth of microflora and physicochemical properties of ayran as a functional food. Algal Res. 2019, 44, 101710. [Google Scholar] [CrossRef]
- Atallah, A.A.; Morsy, O.M.; Gemiel, D.G. Characterization of functional low-fat yogurt enriched with whey protein concentrate, Ca-caseinate and spirulina. Int. J. Food Prop. 2020, 23, 1678–1691. [Google Scholar] [CrossRef]
- Uwamahoro, H.P.; Li, F.; Timilsina, A.; Liu, B.; Wang, X.; Tian, Y. An Assessment of the Lactic Acid-Producing Potential of Bacterial Strains Isolated from Food Waste. Microbiol. Res. 2022, 13, 278–291. [Google Scholar] [CrossRef]
- Wolka, K.; Melaku, B. Exploring selected plant nutrient in compost prepared from food waste and cattle manure and its effect on soil properties and maize yield at Wondo Genet, Ethiopia. Environ. Syst. Res. 2015, 4, 9. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Esmail, G.A.; Arasu, M.V. Co-fermentation of food waste and municipal sludge from the Saudi Arabian environment to improve lactic acid production by Lactobacillus rhamnosus AW3 isolated from date processing waste. Sustainability 2020, 12, 6899. [Google Scholar] [CrossRef]
- Tran, Q.N.M.; Mimoto, H.; Nakasaki, K. Inoculation of lactic acid bacterium accelerates organic matter degradation during composting. Int. Biodeterior. Biodegrad. 2015, 104, 377–383. [Google Scholar] [CrossRef]
- Bühlmann, C.H.; Mickan, B.S.; Tait, S.; Batstone, D.J.; Bahri, P.A. Lactic acid production from food waste at an anaerobic digestion biorefinery: Effect of digestate recirculation and sucrose supplementation. Front. Bioeng. Biotechnol. 2023, 11, 1177739. [Google Scholar] [CrossRef]
- Sabater, C.; Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Margolles, A. Valorization of Vegetable Food Waste and By-Products Through Fermentation Processes. Front. Microbiol. 2020, 11, 581997. [Google Scholar] [CrossRef]
- Novik, G.; Meerovskaya, O.; Savich, V. Waste Degradation and Utilization by Lactic Acid Bacteria: Use of Lactic Acid Bacteria in Production of Food Additives, Bioenergy and Biogas. In Food Additives; IntechOpen: London, UK, 2017; ISBN 978-953-51-3490-9. [Google Scholar]
- Velikova, P.; Stoyanov, A.; Blagoeva, G.; Popova, L.; Petrov, K.; Gotcheva, V.; Angelov, A.; Petrova, P. Starch utilization routes in lactic acid bacteria: New insight by gene expression assay. Starch-Stärke 2016, 68, 953–960. [Google Scholar] [CrossRef]
- Tsapekos, P.; Alvarado-Morales, M.; Baladi, S.; Bosma, E.F.; Angelidaki, I. Fermentative Production of Lactic Acid as a Sustainable Approach to Valorize Household Bio-Waste. Front. Sustain. 2020, 1, 538783. [Google Scholar] [CrossRef]
- López-Gómez, J.P.; Pérez-Rivero, C.; Venus, J. Valorisation of solid biowastes: The lactic acid alternative. Process Biochem. 2020, 99, 222–235. [Google Scholar] [CrossRef]
- Song, L.; Yang, D.; Liu, R.; Liu, S.; Dai, L.; Dai, X. Microbial production of lactic acid from food waste: Latest advances, limits, and perspectives. Bioresour. Technol. 2022, 345, 126052. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Wang, Q.H.; Wang, X.M.; Ma, H.Z.; Liu, Y.Y. Process optimization for lactic acid production from kitchen waste. Harbin Gongye Daxue Xuebao/J. Harbin Inst. Technol. 2009, 41, 58–63. [Google Scholar]
- Dedenaro, G.; Costa, S.; Rugiero, I.; Pedrini, P.; Tamburini, E. Valorization of agri-food waste via fermentation: Production of L-lactic acid as a building block for the synthesis of biopolymers. Appl. Sci. 2016, 6, 379. [Google Scholar] [CrossRef]
- Kwan, T.H.; Hu, Y.; Lin, C.S.K. Valorisation of food waste via fungal hydrolysis and lactic acid fermentation with Lactobacillus casei Shirota. Bioresour. Technol. 2016, 217, 129–136. [Google Scholar] [CrossRef]
- Pleissner, D.; Demichelis, F.; Mariano, S.; Fiore, S.; Navarro Gutiérrez, I.M.; Schneider, R.; Venus, J. Direct production of lactic acid based on simultaneous saccharification and fermentation of mixed restaurant food waste. J. Clean. Prod. 2017, 143, 615–623. [Google Scholar] [CrossRef]
- Song, L.; Liu, S.; Liu, R.; Yang, D.; Dai, X. Direct lactic acid production from household food waste by lactic acid bacteria. Sci. Total Environ. 2022, 840, 156479. [Google Scholar] [CrossRef]
- Anagnostopoulou, C.; Kontogiannopoulos, K.N.; Gaspari, M.; Morlino, M.S.; Assimopoulou, A.N.; Kougias, P.G. Valorization of household food wastes to lactic acid production: A response surface methodology approach to optimize fermentation process. Chemosphere 2022, 296, 133871. [Google Scholar] [CrossRef]
- Chenebault, C.; Moscoviz, R.; Trably, E.; Escudié, R.; Percheron, B. Lactic acid production from food waste using a microbial consortium: Focus on key parameters for process upscaling and fermentation residues valorization. Bioresour. Technol. 2022, 354, 127230. [Google Scholar] [CrossRef]
- Asano, R.; Otawa, K.; Ozutsumi, Y.; Yamamoto, N.; Abdel-Mohsein, H.S.; Nakai, Y. Development and analysis of microbial characteristics of an acidulocomposting system for the treatment of garbage and cattle manure. J. Biosci. Bioeng. 2010, 110, 419–425. [Google Scholar] [CrossRef]
- Tran, Q.N.M.; Mimoto, H.; Koyama, M.; Nakasaki, K. Lactic acid bacteria modulate organic acid production during early stages of food waste composting. Sci. Total Environ. 2019, 687, 341–347. [Google Scholar] [CrossRef]
- Yang, S.Y.; Ji, K.S.; Baik, Y.H.; Kwak, W.S.; McCaskey, T.A. Lactic acid fermentation of food waste for swine feed. Bioresour. Technol. 2006, 97, 1858–1864. [Google Scholar] [CrossRef]
- Seo, J.-Y.; Song, I.-G.; Lee, D.-G.; Lee, K.-Y.; Kim, Y.-J. From food wastes into useful probiotics: Development of Lactic acid bacteria as useful probiotics for animal feed without antibiotics using food waste. J. Korea Org. Resour. Recycl. Assoc. 2006, 14, 112–120. [Google Scholar]
- Juodeikiene, G.; Cizeikiene, D.; Glasner, C.; Bartkiene, E.; Dikiy, A.; Shumilina, E.; Ilić, N.; Di Berardino, S.; Foncesa, C. Evaluation of the potential of utilizing lactic acid bacteria and dairy wastewaters for methane production. Energy Explor. Exploit. 2017, 35, 388–402. [Google Scholar] [CrossRef]
- Bohn, J.; Yüksel-Dadak, A.; Dröge, S.; König, H. Isolation of lactic acid-forming bacteria from biogas plants. J. Biotechnol. 2017, 244, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, P.; Steinigeweg, S.; Siefert, E.; Cypionka, H. Effect of Lactate and Starter Inoculum on Biogas Production from Fresh Maize and Maize Silage. Adv. Microbiol. 2017, 7, 358–376. [Google Scholar] [CrossRef]
- Menardo, S.; Balsari, P.; Tabacco, E.; Borreani, G. Effect of Conservation Time and the Addition of Lactic Acid Bacteria on the Biogas and Methane Production of Corn Stalk Silage. Bioenergy Res. 2015, 8, 1810–1823. [Google Scholar] [CrossRef]
- Vervaeren, H.; Hostyn, K.; Ghekiere, G.; Willems, B. Biological ensilage additives as pretreatment for maize to increase the biogas production. Renew. Energy 2010, 35, 2089–2093. [Google Scholar] [CrossRef]
- Vasmara, C.; Marchetti, R.; Carminati, D. Wastewater from the production of lactic acid bacteria as feedstock in anaerobic digestion. Energy 2021, 229, 120740. [Google Scholar] [CrossRef]
- Jung, S.J.; Kim, M.J.; Chae, S.W. Quality and functional characteristics of kimchi made with organically cultivated young Chinese cabbage (olgari-baechu). J. Ethn. Foods 2016, 3, 150–158. [Google Scholar] [CrossRef]
- Paladhi, A.G.; Joshi, J.T.; George, A.; Manohar, M.V.; Vallinayagam, S.; Malik, J.A. Lipase and lactic acid bacteria for biodegradation and bioremediation. In Microbes and Microbial Biotechnology for Green Remediation; Elsevier: Amsterdam, The Netherlands, 2022; pp. 265–286. [Google Scholar]
- Zhou, X.W.; Zhao, X.H. Susceptibility of nine organophosphorus pesticides in skimmed milk towards inoculated lactic acid bacteria and yogurt starters. J. Sci. Food Agric. 2015, 95, 260–266. [Google Scholar] [CrossRef]
- Ameen, F.A.; Hamdan, A.M.; El-Naggar, M.Y. Assessment of the heavy metal bioremediation efficiency of the novel marine lactic acid bacterium, Lactobacillus plantarum MF042018. Sci. Rep. 2020, 10, 314. [Google Scholar] [CrossRef]
- Syukur, S.; Yolanda, S.; Jamsari, J.; Fachrial, E. Isolation, antimicrobial activity and bioremediation of heavy metal cadmium (Cd) by using lactic acid bacteria from dadih origin lareh sago halaban, payakumbuh, west Sumatera, Indonesia. J. Chem. Pharm. Res. 2015, 7, 235–241. [Google Scholar]
- Hasan, M.S.; Islam, M.Z.; Liza, R.I.; Sarker, M.A.H.; Islam, M.A.; Harun-ur-Rashid, M. Novel Probiotic Lactic Acid Bacteria with In Vitro Bioremediation Potential of Toxic Lead and Cadmium. Curr. Microbiol. 2022, 79, 387. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Salminen, S.; Tahvonen, R. Rapid removal of lead and cadmium from water by specific lactic acid bacteria. Int. J. Food Microbiol. 2007, 114, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Nduko, J.M.; Taguchi, S. Microbial Production of Biodegradable Lactate-Based Polymers and Oligomeric Building Blocks From Renewable and Waste Resources. Front. Bioeng. Biotechnol. 2021, 8, 618077. [Google Scholar] [CrossRef]
- Taib, N.A.A.B.; Rahman, M.R.; Huda, D.; Kuok, K.K.; Hamdan, S.; Bin Bakri, M.K.; Bin Julaihi, M.R.M.; Khan, A. A review on poly lactic acid (PLA) as a biodegradable polymer. Polym. Bull. 2023, 80, 1179–1213. [Google Scholar] [CrossRef]
- Kwan, T.H.; Hu, Y.; Lin, C.S.K. Techno-economic analysis of a food waste valorisation process for lactic acid, lactide and poly(lactic acid) production. J. Clean. Prod. 2018, 181, 72–87. [Google Scholar] [CrossRef]
- Nilsen-Nygaard, J.; Fernández, E.N.; Radusin, T.; Rotabakk, B.T.; Sarfraz, J.; Sharmin, N.; Sivertsvik, M.; Sone, I.; Pettersen, M.K. Current status of biobased and biodegradable food packaging materials: Impact on food quality and effect of innovative processing technologies. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1333–1380. [Google Scholar] [CrossRef]
- Kumari, S.V.G.; Pakshirajan, K.; Pugazhenthi, G. Recent advances and future prospects of cellulose, starch, chitosan, polylactic acid and polyhydroxyalkanoates for sustainable food packaging applications. Int. J. Biol. Macromol. 2022, 221, 163–182. [Google Scholar] [CrossRef]
- Yildirim, S.; Röcker, B.; Pettersen, M.K.; Nilsen-Nygaard, J.; Ayhan, Z.; Rutkaite, R.; Radusin, T.; Suminska, P.; Marcos, B.; Coma, V. Active Packaging Applications for Food. Compr. Rev. Food Sci. Food Saf. 2018, 17, 165–199. [Google Scholar] [CrossRef]
- Huang, S.; Xue, Y.; Yu, B.; Wang, L.; Zhou, C.; Ma, Y. A Review of the Recent Developments in the Bioproduction of Polylactic Acid and Its Precursors Optically Pure Lactic Acids. Molecules 2021, 26, 6446. [Google Scholar] [CrossRef]
- Bosco, F.; Cirrincione, S.; Carletto, R.; Marmo, L.; Chiesa, F.; Mazzoli, R.; Pessione, E. Pha production from cheese whey and “scotta”: Comparison between a consortium and a pure culture of leuconostoc mesenteroides. Microorganisms 2021, 9, 2426. [Google Scholar] [CrossRef]
- Aslim, B.; Çalışkan, F.; Beyatlı, Y.; Gündüz, U. Poly-β-hydroxybutyrate production by lactic acid bacteria. FEMS Microbiol. Lett. 1998, 159, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, R.; Bosco, F.; Mizrahi, I.; Bayer, E.A.; Pessione, E. Towards lactic acid bacteria-based biorefineries. Biotechnol. Adv. 2014, 32, 1216–1236. [Google Scholar] [CrossRef] [PubMed]
- Vijayendra, S.V.N.; Shamala, T.R. Film forming microbial biopolymers for commercial applications-A review. Crit. Rev. Biotechnol. 2014, 34, 338–357. [Google Scholar] [CrossRef]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Moradi, M.; Guimarães, J.T.; Sahin, S. Current applications of exopolysaccharides from lactic acid bacteria in the development of food active edible packaging. Curr. Opin. Food Sci. 2021, 40, 33–39. [Google Scholar] [CrossRef]
- Torino, M.I.; de Valdez, G.F.; Mozzi, F. Biopolymers from lactic acid bacteria. Novel applications in foods and beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef]
- Ghasemlou, M.; Khodaiyan, F.; Oromiehie, A. Physical, mechanical, barrier, and thermal properties of polyol-plasticized biodegradable edible film made from kefiran. Carbohydr. Polym. 2011, 84, 477–483. [Google Scholar] [CrossRef]
- Hassan, A.N.; Corredig, M.; Frank, J.F.; Elsoda, M. Microstructure and rheology of an acid-coagulated cheese (Karish) made with an exopolysaccharide-producing Streptococcus thermophilus strain and its exopolysaccharide non-producing genetic variant. J. Dairy Res. 2004, 71, 116–120. [Google Scholar] [CrossRef]
- Low, D.; Ahlgren, J.A.; Horne, D.; McMahon, D.J.; Oberg, C.J.; Broadbent, J.R. Role of Streptococcus thermophilus MR-1C capsular exopolysaccharide in cheese moisture retention? Appl. Environ. Microbiol. 1998, 64, 2147–2151. [Google Scholar] [CrossRef]
- Lynch, K.M.; McSweeney, P.L.H.; Arendt, E.K.; Uniacke-Lowe, T.; Galle, S.; Coffey, A. Isolation and characterisation of exopolysaccharide-producing Weissella and Lactobacillus and their application as adjunct cultures in Cheddar cheese. Int. Dairy J. 2014, 34, 125–134. [Google Scholar] [CrossRef]
- Costa, N.E.; Hannon, J.A.; Guinee, T.P.; Auty, M.A.E.; McSweeney, P.L.H.; Beresford, T.P. Effect of exopolysaccharide produced by isogenic strains of Lactococcus lactis on half-fat Cheddar cheese. J. Dairy Sci. 2010, 93, 3469–3486. [Google Scholar] [CrossRef] [PubMed]
- Dertli, E.; Toker, O.S.; Durak, M.Z.; Yilmaz, M.T.; Tatlisu, N.B.; Sagdic, O.; Cankurt, H. Development of a fermented ice-cream as influenced by in situ exopolysaccharide production: Rheological, molecular, microstructural and sensory characterization. Carbohydr. Polym. 2016, 136, 427–440. [Google Scholar] [CrossRef]
- Li, D.; Li, J.; Zhao, F.; Wang, G.; Qin, Q.; Hao, Y. The influence of fermentation condition on production and molecular mass of EPS produced by Streptococcus thermophilus 05-34 in milk-based medium. Food Chem. 2016, 197, 367–372. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rachwał, K.; Gustaw, K. Lactic Acid Bacteria in Sustainable Food Production. Sustainability 2024, 16, 3362. https://doi.org/10.3390/su16083362
Rachwał K, Gustaw K. Lactic Acid Bacteria in Sustainable Food Production. Sustainability. 2024; 16(8):3362. https://doi.org/10.3390/su16083362
Chicago/Turabian StyleRachwał, Kamila, and Klaudia Gustaw. 2024. "Lactic Acid Bacteria in Sustainable Food Production" Sustainability 16, no. 8: 3362. https://doi.org/10.3390/su16083362
APA StyleRachwał, K., & Gustaw, K. (2024). Lactic Acid Bacteria in Sustainable Food Production. Sustainability, 16(8), 3362. https://doi.org/10.3390/su16083362