Sub-Daily Temperature Heterogeneity in a Side Channel and the Influence on Habitat Suitability of Freshwater Fish

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
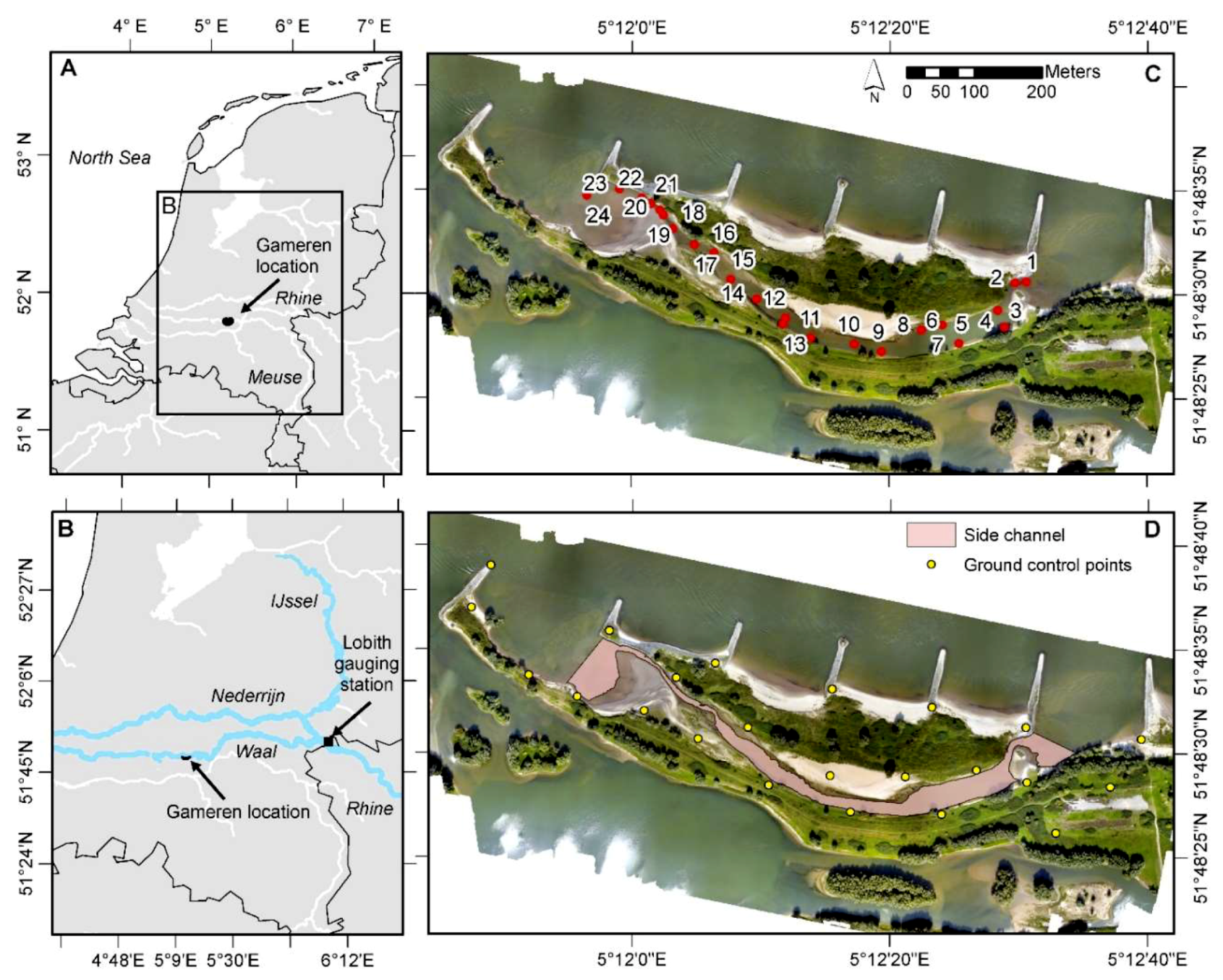
2.1. Study Area
2.2. Thermal Iimagery
2.3. In-Situ Measurements
2.4. Pre-Processing and Accuracy Assessment
2.5. Spatiotemporal Temperature Changes and Habitat Suitability
2.5.1. Water Temperature Variation
2.5.2. Added Value of Using Thermal Imagery to Predict the Habitat Suitability
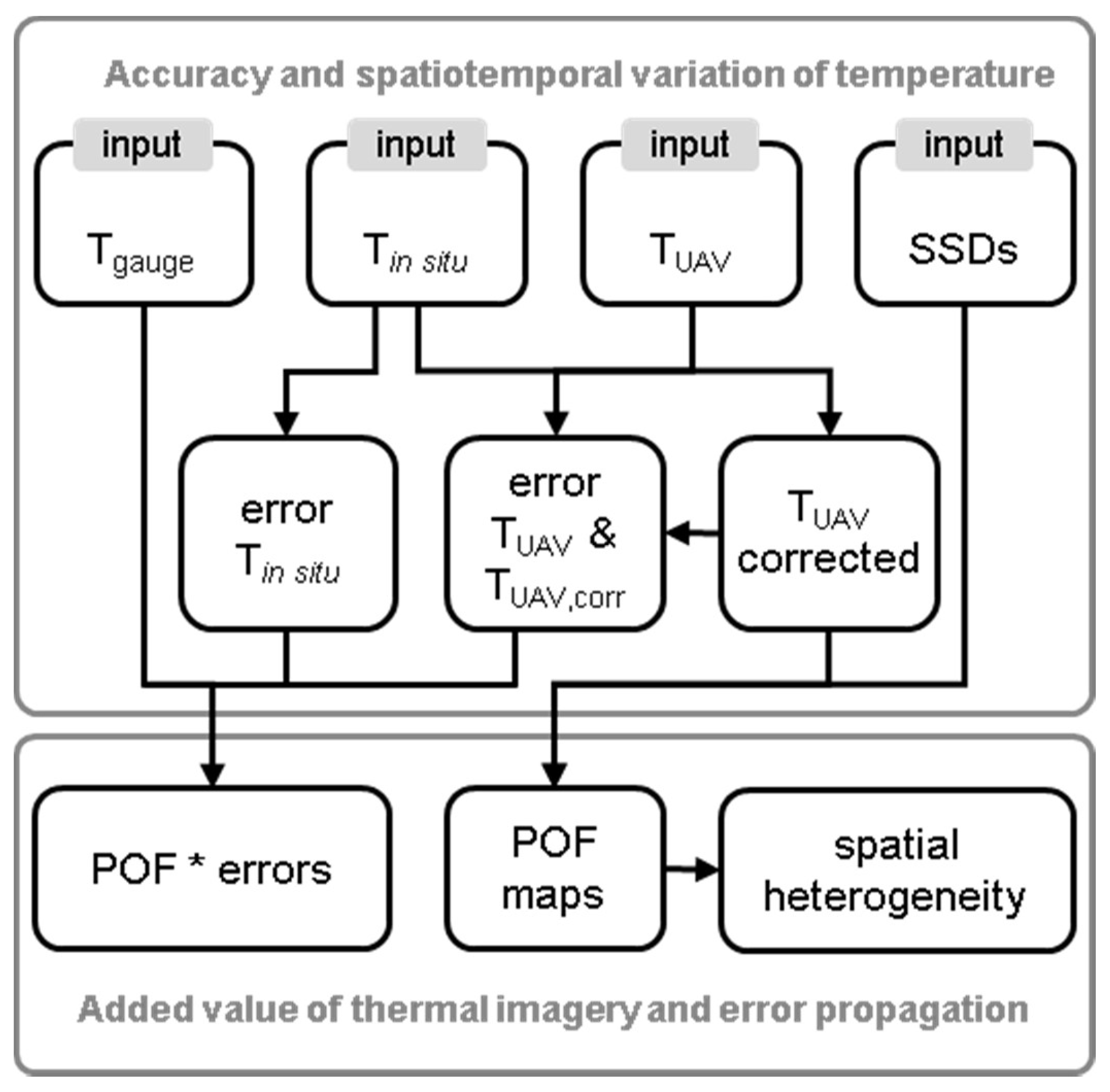
2.5.3. Temperature Error Propagation
3. Results
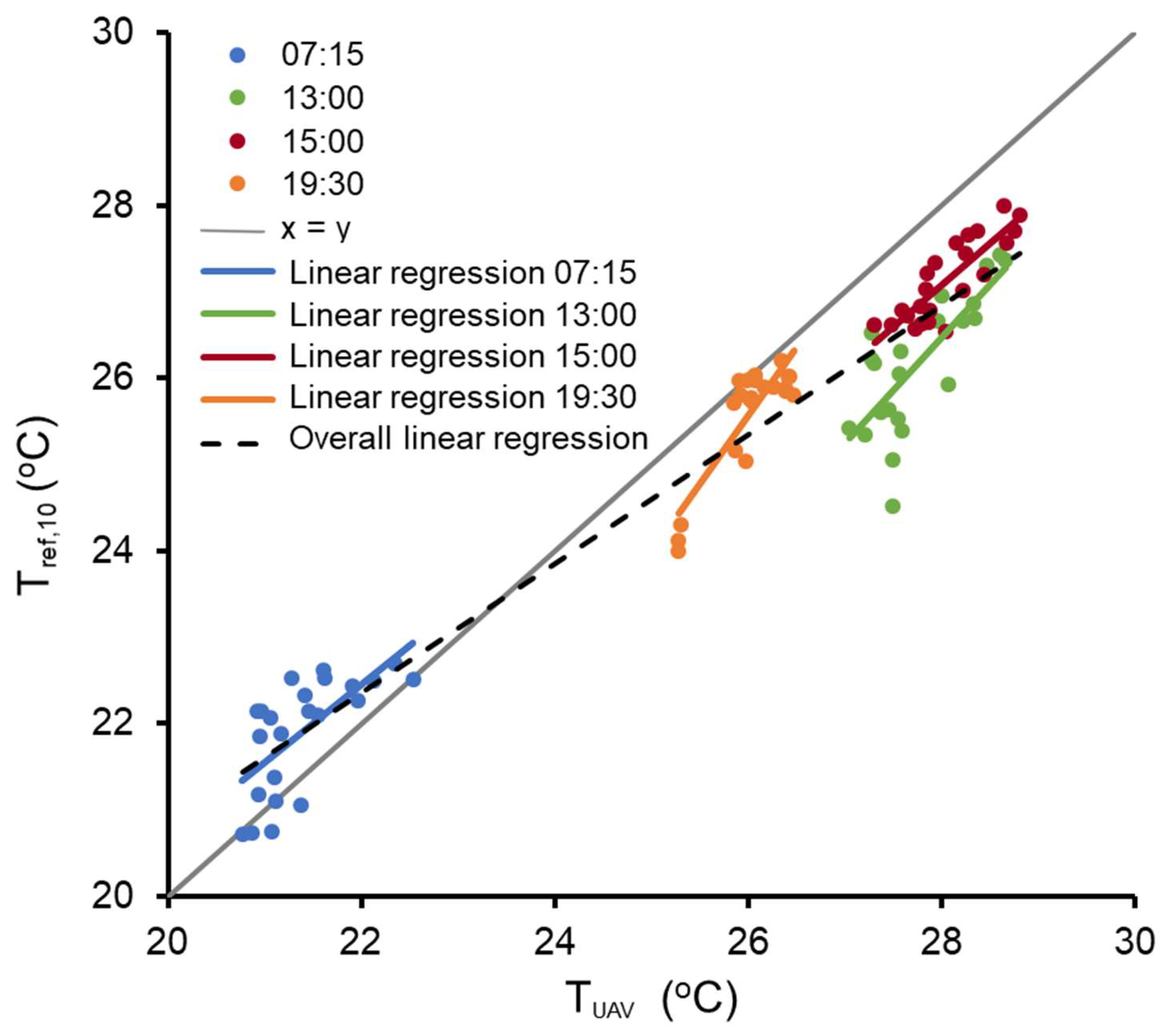
3.1. Thermal Imagery Accuracy Assessment
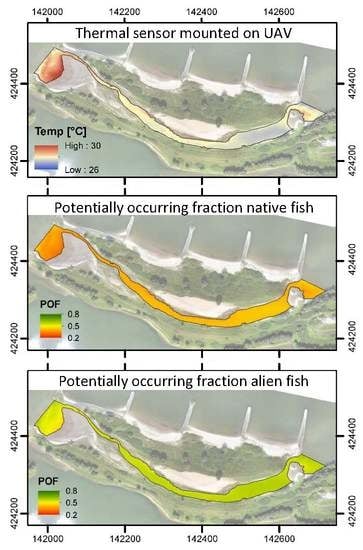
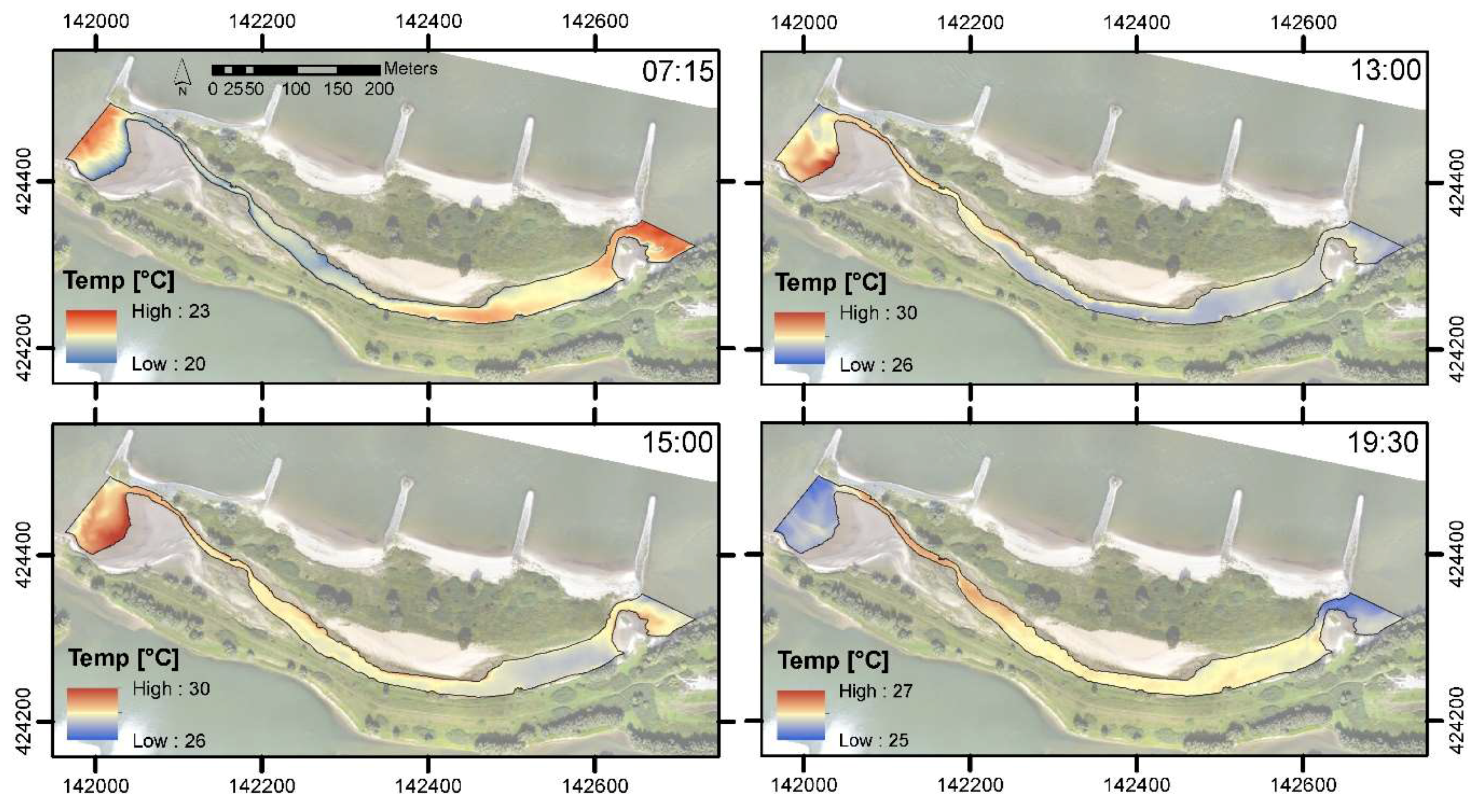
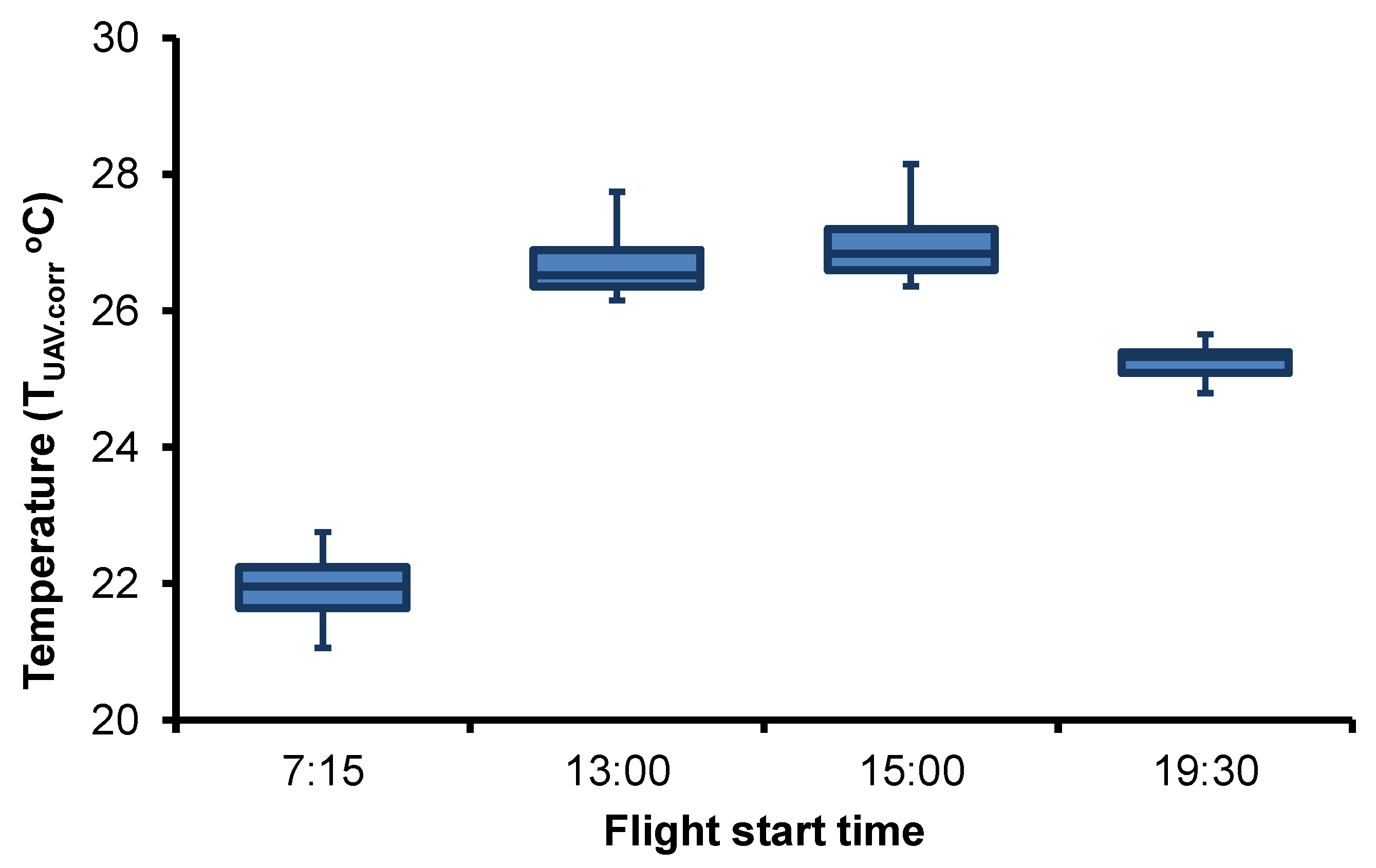
3.2. Spatiotemporal Variation of Temperature in the Side Channel
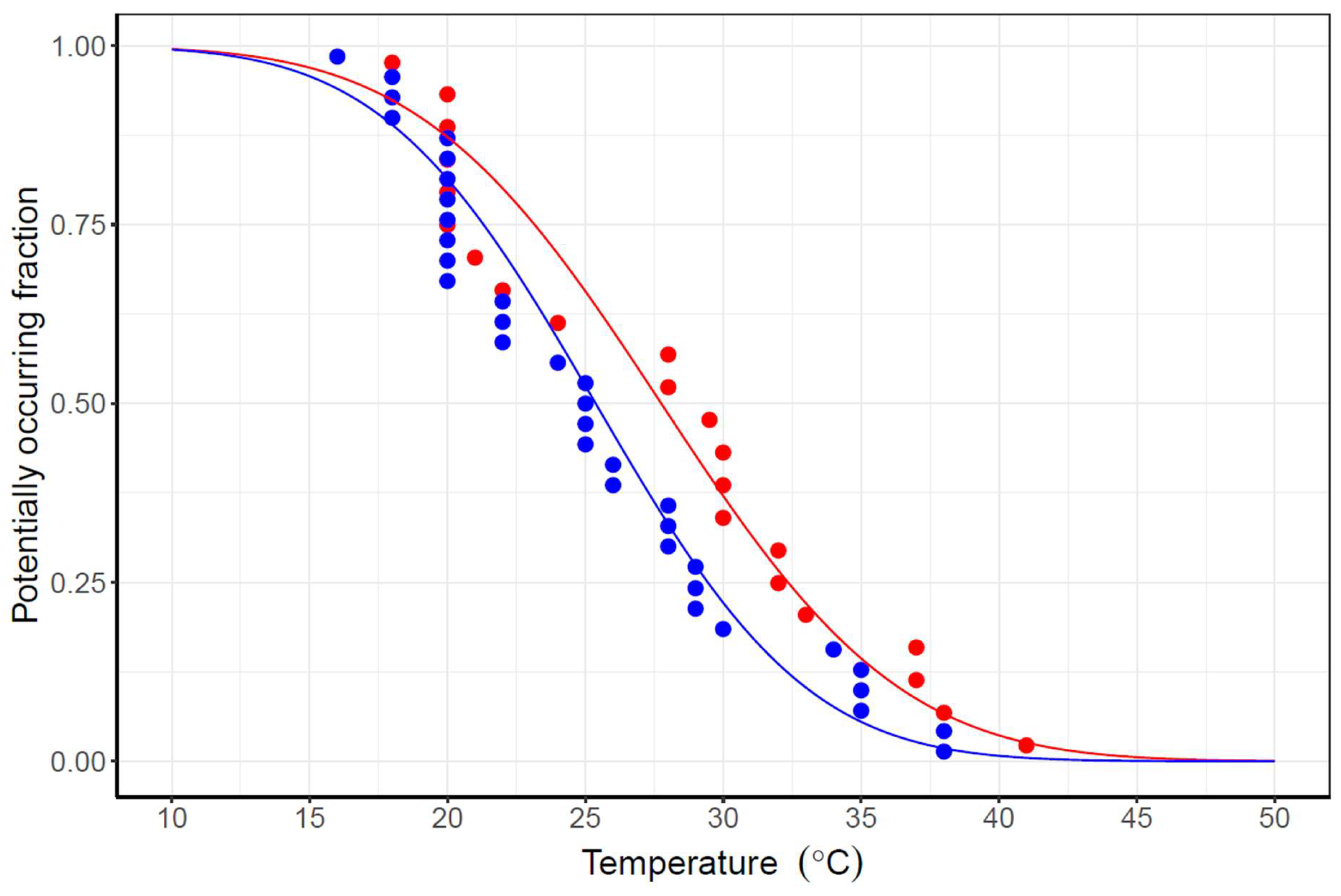
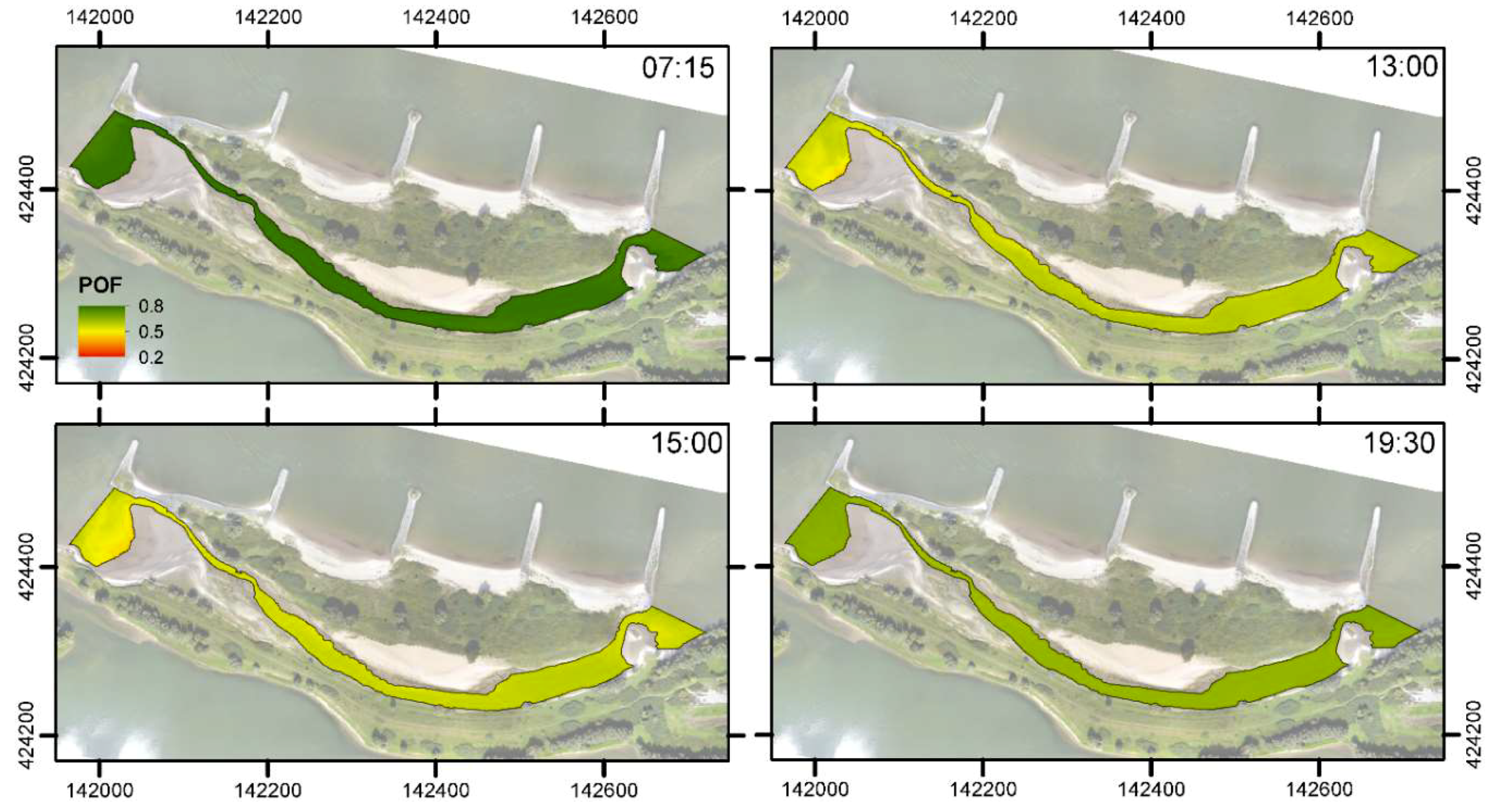
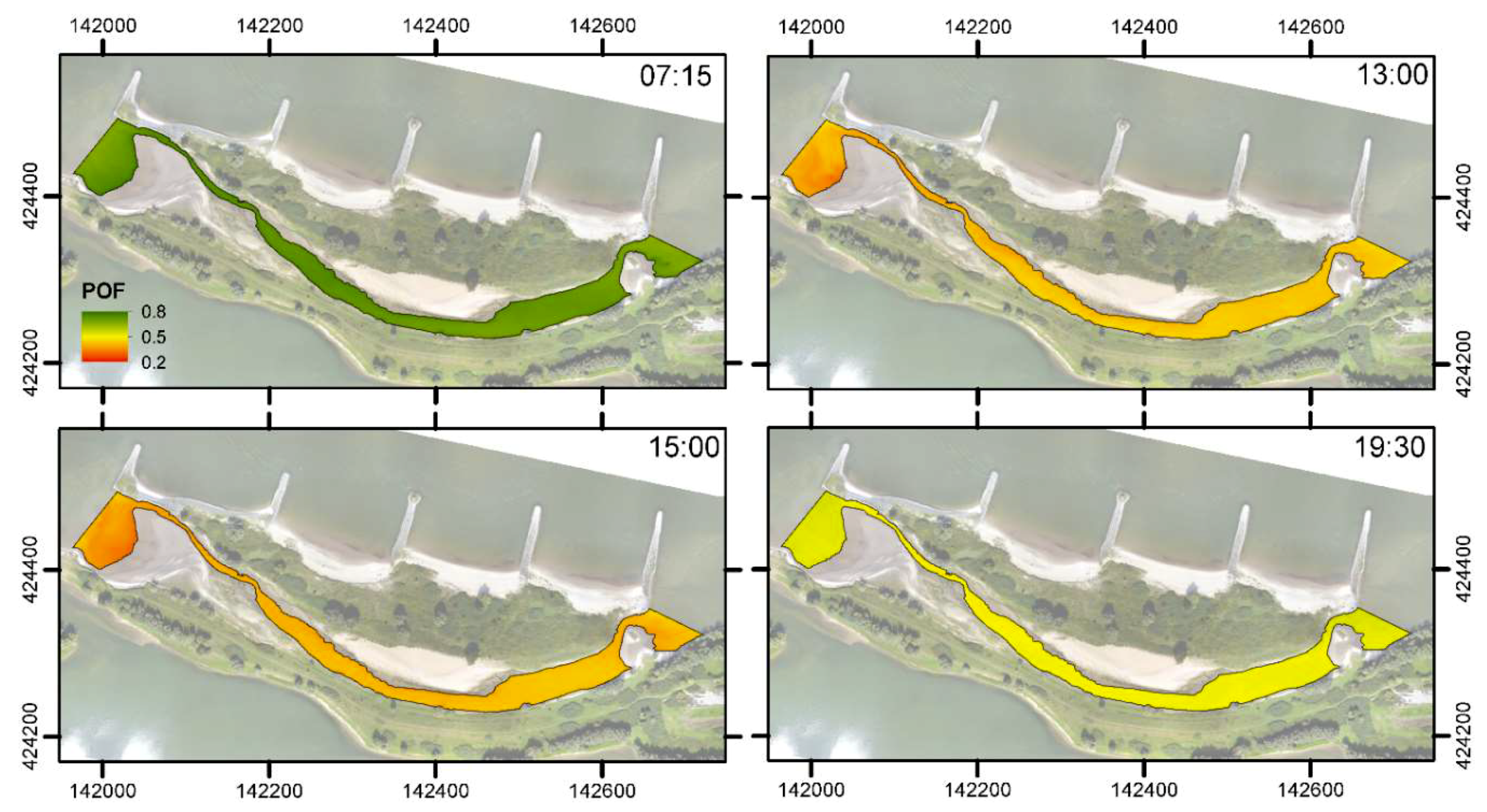
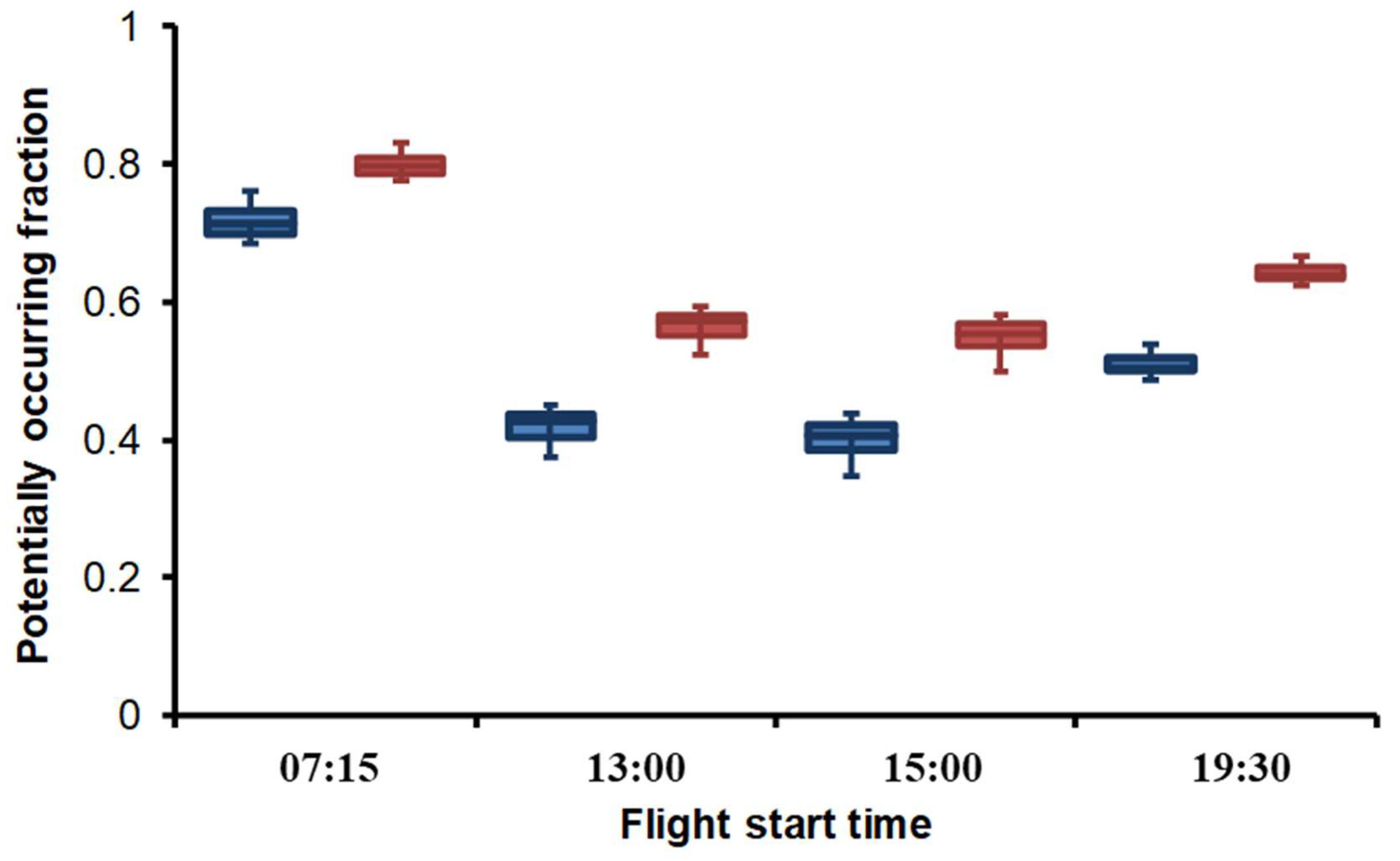
3.3. Habitat Suitability
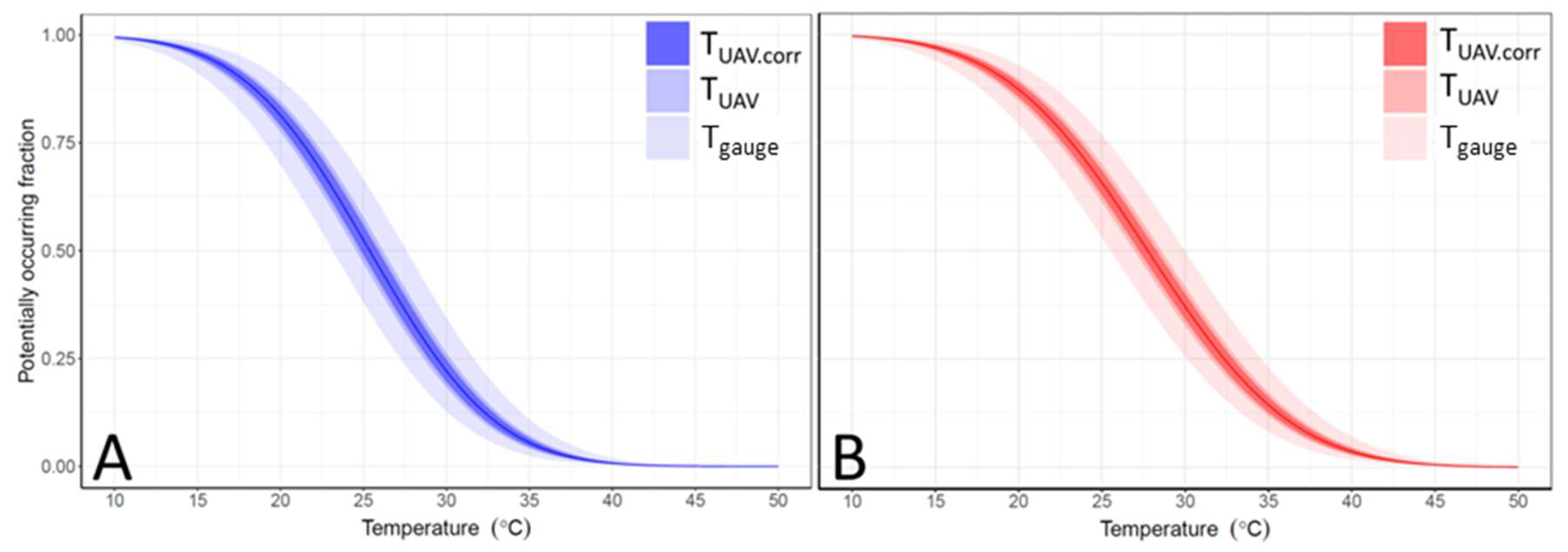
3.4. Temperature Error Propagation to Habitat Suitability
4. Discussion
4.1. Accuracy of Thermal Imagery to Estimate Water Temperature
4.2. Spatiotemporal Variation of Water Temperature and Habitat Suitability
4.3. Added Value of Thermal Imagery to Estimate Water Temperature and Habitat Suitability
4.4. Recommendations for Side Channels as Floodplain Restoration Measure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- Burgmer, T.; Hillebrand, H.; Pfenninger, M. Effects of climate-drive temperature changes on the diversity of freshwater macroinvertebrates. Oecologia 2007, 151, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, H. Potential effects of global warming on northern European freshwater fish and fisheries. Fish. Manag. Ecol. 1996, 3, 59–71. [Google Scholar] [CrossRef]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities - the role of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar] [CrossRef]
- Van Vliet, M.T.H.; Franssen, W.H.P.; Yearsley, J.R.; Ludwig, F.; Haddeland, I.; Lettenmaier, D.P.; Kabat, P. Global river discharge and water temperature under climate change. Global Environ. Chang. 2013, 23, 450–464. [Google Scholar] [CrossRef]
- Souchon, Y.; Tissot, L. Synthesis of the thermal tolerances of the common freshwater fish species in large Western Europe rivers. Knowl. Manag. Aquat. Ec. 2012, 405, 03. [Google Scholar] [CrossRef]
- Koopman, K.R.; Collas, F.P.L.; Van der Velde, G.; Verberk, W.C.E.P. Oxygen can limit heat tolerance in freshwater gastropods: Differences between gill and lung breathers. Hydrobiologia 2016, 763, 301–312. [Google Scholar] [CrossRef]
- Verberk, W.C.E.P.; Leuven, R.S.E.W.; Van der Velde, G.; Gabel, F. Thermal limits in native and alien freshwater peracarid Crustacea: The role of habitat use and oxygen limitation. Funct. Ecol. 2018, 32, 926–936. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the effects of climate change on aquatic invasive species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef]
- Leuven, R.S.E.W.; Hendriks, A.J.; Huijbregts, M.A.J.; Lenders, H.J.R.; Matthews, J.; Van der Velde, G. Differences in sensitivity of native and exotic fish species to changes in river temperature. Curr. Zool. 2011, 57, 852–862. [Google Scholar] [CrossRef]
- Verbrugge, L.N.H.; Schipper, A.M.; Huijbregts, M.A.J.; Van der Velde, G.; Leuven, R.S.E.W. Sensitivity of native and non-native mollusc species to changing river water temperature and salinity. Biol. Invasions 2012, 14, 1187–1199. [Google Scholar] [CrossRef]
- Collas, F.P.L.; Buijse, A.D.; Hendriks, A.J.; Van der Velde, G.; Leuven, R.S.E.W. Sensitivity of European freshwater bivalve species to climate related environmental factors. Ecosphere 2018, 9, e02184. [Google Scholar] [CrossRef]
- Rahel, F.J. Homogenization of fish faunas across the United States. Science 2000, 288, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Leprieur, F.; Beauchard, O.; Blanchet, S.; Oberdorff, T.; Brosse, S. Fish invasions in the world’s river systems: When natural processes are blurred by human activities. PLoS Biol. 2008, 6, 404–410. [Google Scholar] [CrossRef]
- Leuven, R.S.E.W.; Van der Velde, G.; Baijens, I.; Snijders, J.; Van der Zwart, C.; Lenders, H.J.R.; Bij de Vaate, A. The river Rhine: A global highway for dispersal of aquatic invasive species. Biol. Invasions 2009, 11, 1989–2008. [Google Scholar] [CrossRef]
- De Vries, P.; Tamis, J.E.; Murk, A.J.; Smit, M.G.D. Development and application of a species sensitivity distribution for temperature-induced mortality in the aquatic environment. Environ. Toxicol. Chem. 2009, 27, 2591–2598. [Google Scholar] [CrossRef] [PubMed]
- Del Signore, A.; Hendriks, A.J.; Lenders, H.J.R.; Leuven, R.S.E.W.; Breure, A.M. Development and application of the SSD approach in scientific case studies for Ecological Risk Assessment. Environ. Toxicol. Chem. 2016, 35, 2149–2161. [Google Scholar] [CrossRef]
- Posthuma, L.; Suter II, G.W.; Traas, T.P. Species Sensitivity Distributions in Ecotoxicology; Lewis Publishers: Boca Raton, FL, USA, 2002. [Google Scholar]
- Koopman, K.R.; Collas, F.P.L.; Breure, A.M.; Lenders, H.J.R.; Van der Velde, G.; Leuven, R.S.E.W. Predicting effects of ship-induced changes in flow velocity on native and alien molluscs in the littoral zone of lowland rivers. Aquat. Invasions 2018, 13, 481–490. [Google Scholar] [CrossRef]
- Collas, F.P.L.; Koopman, K.R.; Hendriks, A.J.; Van der Velde, G.; Verbrugge, L.N.H.; Leuven, R.S.E.W. Effects of desiccation on native and non-native molluscs in rivers. Freshw. Biol. 2014, 59, 41–55. [Google Scholar] [CrossRef]
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Vatland, S.J.; Gresswell, R.E.; Poole, G.C. Quantifying stream thermal regimes at multiple scales: Combining thermal infrared imagery and stationary stream temperature data in a novel modelling framework. Water Resour. Res. 2014, 51, 31–46. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Liss, W.J.; Frissell, C.A. Cold water patches in warm streams: Physicochemical characteristics and the influence of shading. J. Am. Water Resour. Assoc. 2003, 39, 355–368. [Google Scholar] [CrossRef]
- Matthews, K.R.; Berg, N.H.; Azuma, D.L.; Lambert, T.R. Cool water formation and trout habitat use in a deep pool in the Sierra Nevada, California. Trans. Am. Fish. Soc. 1994, 123, 549–564. [Google Scholar] [CrossRef]
- Dzara, J.R.; Neilson, B.T.; Null, S.E. Quantifying thermal refugia connectivity by combining temperature modeling, distributed temperature sensing, and thermal infrared imaging. Hydrol. Earth Syst. Sci. 2019, 23, 2965–2982. [Google Scholar] [CrossRef]
- Boyd, M.; Kasper, B. Analytical Methods for Dynamic Open Channel Heat and Mass Transfer; Technical report; Department of Environmental Quality: Portland, OR, USA, 2017.
- Handcock, R.N.; Torgersen, C.E.; Cherkauer, K.A.; Gillespie, A.R.; Tockner, K.; Faux, R.N.; Tan, J. Thermal infrared remote sensing of water temperature in riverine landscapes. In Fluvial Remote Sensing for Science and Management; John Wiley & Sons: New York, NY, USA, 2012; pp. 85–113. [Google Scholar]
- Turner, D.; Lucieer, A.; Malenovský, Z.; King, D.H.; Robinson, S.A. Spatial co-registration of ultra-high resolution visible, multispectral and thermal images acquired with a micro-UAV over Antarctic moss beds. Remote Sens. 2014, 6, 4003–4024. [Google Scholar] [CrossRef]
- Norman, J.M.; Becker, F. Terminology in thermal infrared remote sensing of natural surfaces. Int. J. Remote Sens. 1995, 12, 159–173. [Google Scholar] [CrossRef]
- Anderson, J.M.; Wilson, S.B. The physical basis of current infrared remote-sensing techniques and the interpretation of data from aerial surveys. Int. J. Remote Sens. 1984, 5, 1–18. [Google Scholar] [CrossRef]
- DeBell, L.; Anderson, K.; Brazier, R.E.; King, N.; Jones, L. Water resource management at catchment scales using lightweight UAVs: Current capabilities and future perspectives. J. Unmanned Veh. Syst. 2015, 4, 7–30. [Google Scholar] [CrossRef]
- Van Iersel, W.; Straatsma, M.; Middelkoop, H.; Addink, E. Multitemporal Classification of River Floodplain Vegetation Using Time Series of UAV Images. Remote Sens. 2018, 10, 1144. [Google Scholar] [CrossRef]
- Näsi, R.; Honkavaara, E.; Lyytikäinen-Saarenmaa, P.; Blomqvist, M.; Litkey, P.; Hakala, T.; Holopainen, M. Using UAV-based photogrammetry and hyperspectral imaging for mapping bark beetle damage at tree-level. Remote Sens. 2015, 7, 15467–15493. [Google Scholar] [CrossRef]
- Khan, S.; Aragão, L.; Iriarte, J. A UAV–lidar system to map Amazonian rainforest and its ancient landscape transformations. Int. J. Remote Sens. 2017, 38, 2313–2330. [Google Scholar] [CrossRef]
- Nishar, A.; Richards, S.; Breen, D.; Robertson, J.; Breen, B. Thermal infrared imaging of geothermal environments by UAV (unmanned aerial vehicle). J. Unmanned Veh. Syst. 2016, 4, 136–145. [Google Scholar] [CrossRef]
- SenseFly Drones for Professionals, Mapping & Photogrammetry, Flight Planning & Control Software. Available online: https://www.sensefly.com/home.html (accessed on 12 December 2017).
- SenseFly Our Camera Payloads & Accessories. Available online: https://www.sensefly.com/drones/accessories.html (accessed on 25 January 2018).
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2015. Available online: https://www.R-project.org/ (accessed on 25 January 2018).
- Hijmans, R.J.; Van Etten, J.; Cheng, J.; Mattiuzzi, M.; Sumner, M.; Greenberg, J.A.; Perpinan Lamigueiro, O.; Bevan, A.; Racine, E.B.; Shortridge, A.; et al. Package Raster - Geographic Data Analysis and Modelling. Available online: https://cran.r-project.org/web/packages/raster/raster.pdf (accessed on 23 January 2018).
- Cressie, N. Statistics for Spatial Data; John Wiley: New York, NY, USA, 1993. [Google Scholar]
- Torgersen, C.E.; Faux, R.N.; McIntosh, B.A.; Poage, N.J.; Norton, D.J. Airborne thermal remote sensing for water temperature assessment in rivers and streams. Remote Sens. Environ. 2001, 76, 386–398. [Google Scholar] [CrossRef]
- Fullerton, A.H.; Torgersen, C.E.; Lawler, J.J.; Faux, R.N.; Steel, E.A.; Beechie, T.J.; Leibowitz, S.G. Rethinking the longitudinal stream temperature paradigm: Region-wide comparison of thermal infrared imagery reveals unexpected complexity of river temperatures. Hydrol. Process. 2015, 29, 4719–4737. [Google Scholar] [CrossRef]
- Dugdale, S.J.; Bergeron, N.E.; St-Hilaire, A. Spatial distribution of thermal refuges analysed in relation to riverscape hydromorphology using airborne thermal infrared imagery. Remote Sens. Environ. 2015, 160, 43–55. [Google Scholar] [CrossRef]
- Handcock, R.N.; Gillespie, A.R.; Cherkauer, K.A.; Kay, J.E.; Burges, S.J.; Kampf, S.K. Accuracy and uncertainty of thermal-infrared remote sensing of stream temperatures at multiple spatial scales. Remote Sens. Environ. 2006, 100, 427–440. [Google Scholar] [CrossRef]
- Henderson, B.G.; Theiler, J.; Villeneuve, P. The polarized emissivity of a wind-roughened sea surface: A Monte Carlo model. Remote Sens. Environ. 2003, 88, 453–467. [Google Scholar] [CrossRef]
- Prakash, P. Thermal remote sensing: Concepts, issues and applications. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2000, 33, 239–243. [Google Scholar]
- Yousef, Y.A.; McLellon, W.M.; Zebuth, H.H. Changes in phosphorus concentrations due to mixing by motorboats in shallow lakes. Water Res. 1980, 14, 841–852. [Google Scholar] [CrossRef]
- Becker, A.; Kirchesch, V.; Baumert, H.Z.; Fischer, H.; Schöl, A. Modelling the effects of thermal stratification on the oxygen budget of an impounded river. River Res. Appl. 2010, 26, 572–588. [Google Scholar] [CrossRef]
- Engel, F.; Fischer, H. Effect of thermal stratification on phytoplankton and nutrient dynamics in a regulated river (Saar, Germany). River Res. Appl. 2017, 33, 135–146. [Google Scholar] [CrossRef]
- Clough, S.; Ladle, M. Diel migration and site fidelity in a stream-dwelling cyprinid, Leuciscus leuciscus. J. Fish. Biol. 1997, 50, 1117–1119. [Google Scholar] [CrossRef]
- Borcherding, J.; Bauerfeld, M.; Hintzen, D.; Neumann, D. Lateral migrations of fishes between floodplain lakes and their drainage channels at the Lower Rhine: Diel and seasonal aspects. J. Fish. Biol. 2002, 61, 1154–1170. [Google Scholar] [CrossRef]
- Hohausová, E.; Copp, G.H.; Jankovský, P. Movement of fish between a river and its backwater: Diel activity and relation to environmental gradients. Ecol. Freshw. Fish. 2003, 12, 107–117. [Google Scholar] [CrossRef]
- Collas, F.P.L.; Buijse, A.D.; Van den Heuvel, L.; Van Kessel, N.; Schoor, M.M.; Eerden, H.; Leuven, R.S.E.W. Longitudinal training dams mitigate effects of shipping on environmental conditions and fish density in the littoral zones of the river Rhine. Sci. Total Environ. 2018b, 619–620, 1183–1193. [Google Scholar] [CrossRef]
- Schiemer, F.; Keckeis, H.; Kamler, E. The early life history stages of riverine fish: Ecophysiological and environmental bottlenecks. Comp. Biochem. Phys. A 2003, 133, 439–449. [Google Scholar] [CrossRef]
- Trinci, G.; Harvey, G.L.; Henshaw, A.J.; Bertoldi, W.; Hölker, F. Life in turbulent flow interactions between hydrodynamics and aquatic organisms in rivers. WIREs Water 2017, 4, e1213. [Google Scholar] [CrossRef]
- Armstrong, J.B.; Schindler, D.E.; Ruff, C.P.; Brooks, G.T.; Bentley, K.E.; Torgersen, C.E. Diel horizontal migration in streams: Juvenile fish exploit spatial heterogeneity in thermal and trophic resources. Ecology 2013, 94, 2066–2075. [Google Scholar] [CrossRef]
- Nunn, A.D.; Cowx, I.G.; Frear, P.A.; Harvey, J.P. Is water temperature an adequate predictor of recruitment success in cyprinid fish populations in lowland rivers? Freshw. Biol. 2003, 48, 579–588. [Google Scholar] [CrossRef]
- Nienhuis, P.H.; Buijse, A.D.; Leuven, R.S.E.W.; Smits, A.J.M.; De Nooij, R.J.W.; Samborska, E.M. Ecological rehabilitation of the lowland basin of the river Rhine (NW Europe). Hydrobiologia 2002, 478, 53–72. [Google Scholar] [CrossRef]
- Van Stokkom, H.T.C.; Smits, A.J.M.; Leuven, R.S.E.W. Flood defense in the Netherlands: A new era, a new approach. Water Int. 2005, 30, 76–87. [Google Scholar] [CrossRef]
- Rijke, J.; Van Herk, S.; Zevenbergen, C.; Ashley, R. Room for the River: Delivering integrated river basin management in the Netherlands. Int. J. River Basin Manag. 2012, 10, 369–382. [Google Scholar] [CrossRef]
- Straatsma, M.W.; Bloecker, A.M.; Lenders, H.J.R.; Leuven, R.S.E.W.; Kleinhans, M.G. Biodiversity recovery following delta-wide measures for flood risk reduction. Sci. Adv. 2017, 3, e1602762. [Google Scholar] [CrossRef]
- Cucherousset, J.; Olden, J.D. Ecological Impacts of Non-native Freshwater Fishes. Fisheries 2011, 36, 215–230. [Google Scholar] [CrossRef]
- Sinokrot, B.A.; Gulliver, J.S. In-stream flow impact on river water temperatures. J. Hydraul. Res. 2000, 38, 339–349. [Google Scholar] [CrossRef]
- Kurylyk, B.L.; MacQuarrie, K.T.B.; Li, T. Preserving, augmenting, and creating cold-water thermal refugia in rivers: Concepts derived from research on the Miramichi River, New Brunswick (Canada). Ecohydrology 2015, 8, 1095–1108. [Google Scholar] [CrossRef]
- Christidis, N.; Jones, G.S.; Stott, P.A. Dramatically increasing chance of extremely hot summers since the 2003 European heatwave. Nat. Clim. Chang. 2015, 5, 46–50. [Google Scholar] [CrossRef]
- Baptist, M.J.; Penning, W.E.; Duel, H.; Smits, A.J.M.; Geerling, G.W.; Van der Lee, G.E.M.; Van Alphen, J.S.L. Assessment of the effects of cyclic floodplain rejuvenation on flood levels and biodiversity along the Rhine River. River Res. Appl. 2004, 20, 285–297. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UAV Flights | |
| Flight duration | 15 min |
| Flying altitude | 130 m |
| Flight start times | 07:15, 13:00, 15:00, 19:30 |
| ThermoMAP sensor [37] | |
| Ground resolution | 25 cm × 25 cm |
| Radiometric sensitivity | 7–15 µm |
| Max. response at | 10 µm |
| Temperature resolution | 0.1 °C |
| Temperature range | −40 to +160 °C |
| Temperature calibration | Automatic, in-flight |
| Output format | TIFF images |
| Weight of sensor | ~ 134 g |
| Flight Time | Average Tref.10 (°C) | MAE (SD) + (°C) | Regression Equation | R2 | UAV RMSE † (°C) | LOOCV RMSE ‡ (°C) |
|---|---|---|---|---|---|---|
| 07:15 | 21.90 | 0.58 (0.40) | Tref.10 = 0.9025 TUAV + 2.5880 | 0.45 | 0.49 | 0.38 |
| 13:00 | 26.19 | 1.58 (0.52) | Tref.10 = 1.2218 TUAV + 7.7372 | 0.57 | 0.73 | 0.51 |
| 15:00 | 27.13 | 0.92 (0.26) | Tref.10 = 0.9595 TUAV + 0.2136 | 0.71 | 0.36 | 0.29 |
| 19:30 | 25.61 | 0.42 (0.37) | Tref.10 = 1.5732 TUAV – 14.331 | 0.74 | 0.49 | 0.21 |
| Overall | 24.94 | 0.81 (0.60) | Tref.10 = 0.7469 TUAV + 5.9301 | 0.93 | 0.53 | 0.34 |
| Tref.50 | Tref.50 = 0.8085 Tref.10 + 4.2448 | 0.87 | 1.31 | 0.46 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collas, F.P.L.; van Iersel, W.K.; Straatsma, M.W.; Buijse, A.D.; Leuven, R.S.E.W. Sub-Daily Temperature Heterogeneity in a Side Channel and the Influence on Habitat Suitability of Freshwater Fish. Remote Sens. 2019, 11, 2367. https://doi.org/10.3390/rs11202367
Collas FPL, van Iersel WK, Straatsma MW, Buijse AD, Leuven RSEW. Sub-Daily Temperature Heterogeneity in a Side Channel and the Influence on Habitat Suitability of Freshwater Fish. Remote Sensing. 2019; 11(20):2367. https://doi.org/10.3390/rs11202367
Chicago/Turabian StyleCollas, Frank P.L., Wimala K. van Iersel, Menno W. Straatsma, Anthonie D. Buijse, and Rob S.E.W. Leuven. 2019. "Sub-Daily Temperature Heterogeneity in a Side Channel and the Influence on Habitat Suitability of Freshwater Fish" Remote Sensing 11, no. 20: 2367. https://doi.org/10.3390/rs11202367
APA StyleCollas, F. P. L., van Iersel, W. K., Straatsma, M. W., Buijse, A. D., & Leuven, R. S. E. W. (2019). Sub-Daily Temperature Heterogeneity in a Side Channel and the Influence on Habitat Suitability of Freshwater Fish. Remote Sensing, 11(20), 2367. https://doi.org/10.3390/rs11202367