Monitoring Mega-Crown Leaf Turnover from Space

 ,
,  ,
,  ,
,
Abstract
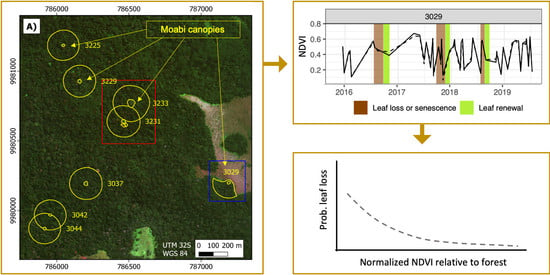
:
1. Introduction
2. Materials and Methods
2.1. Data
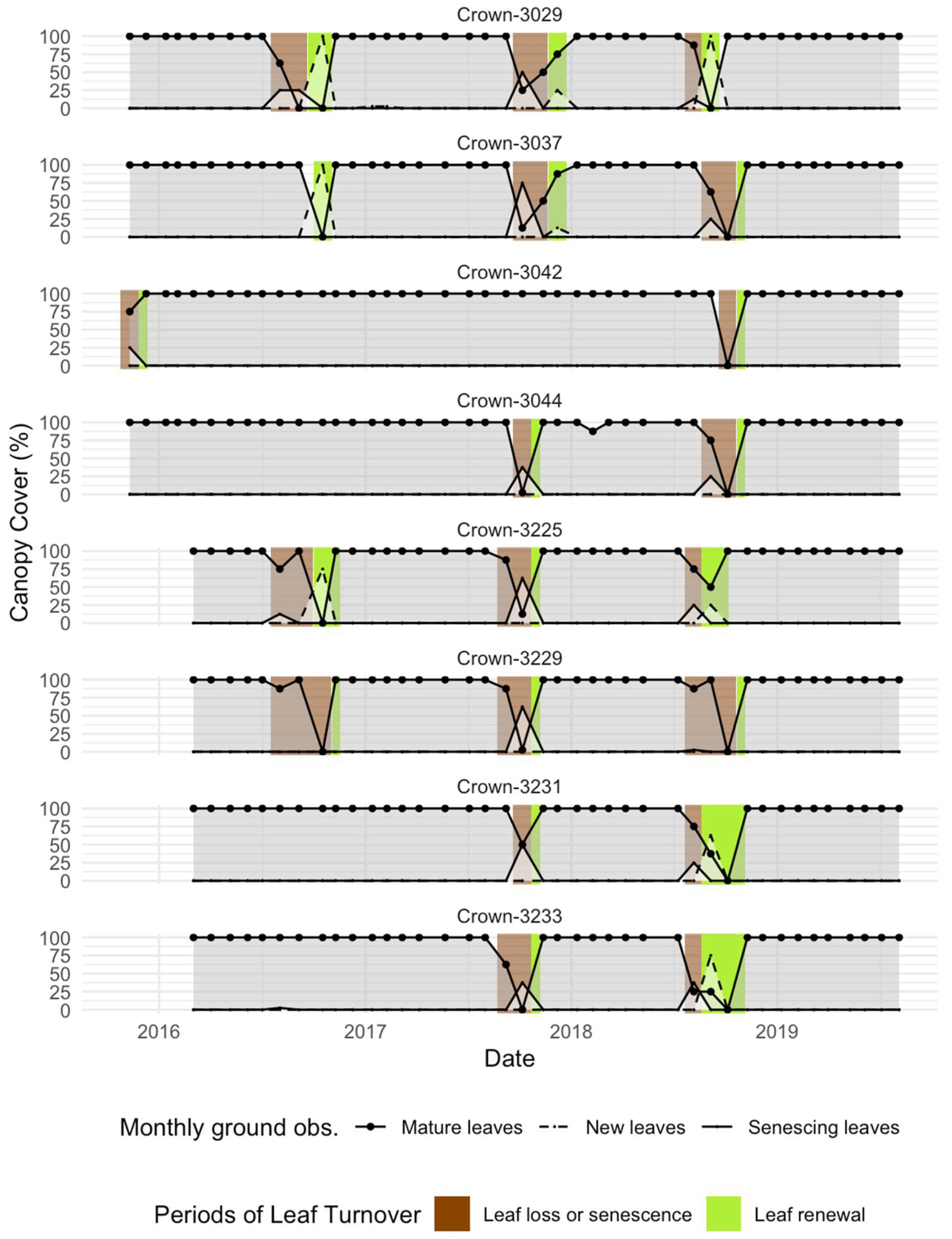
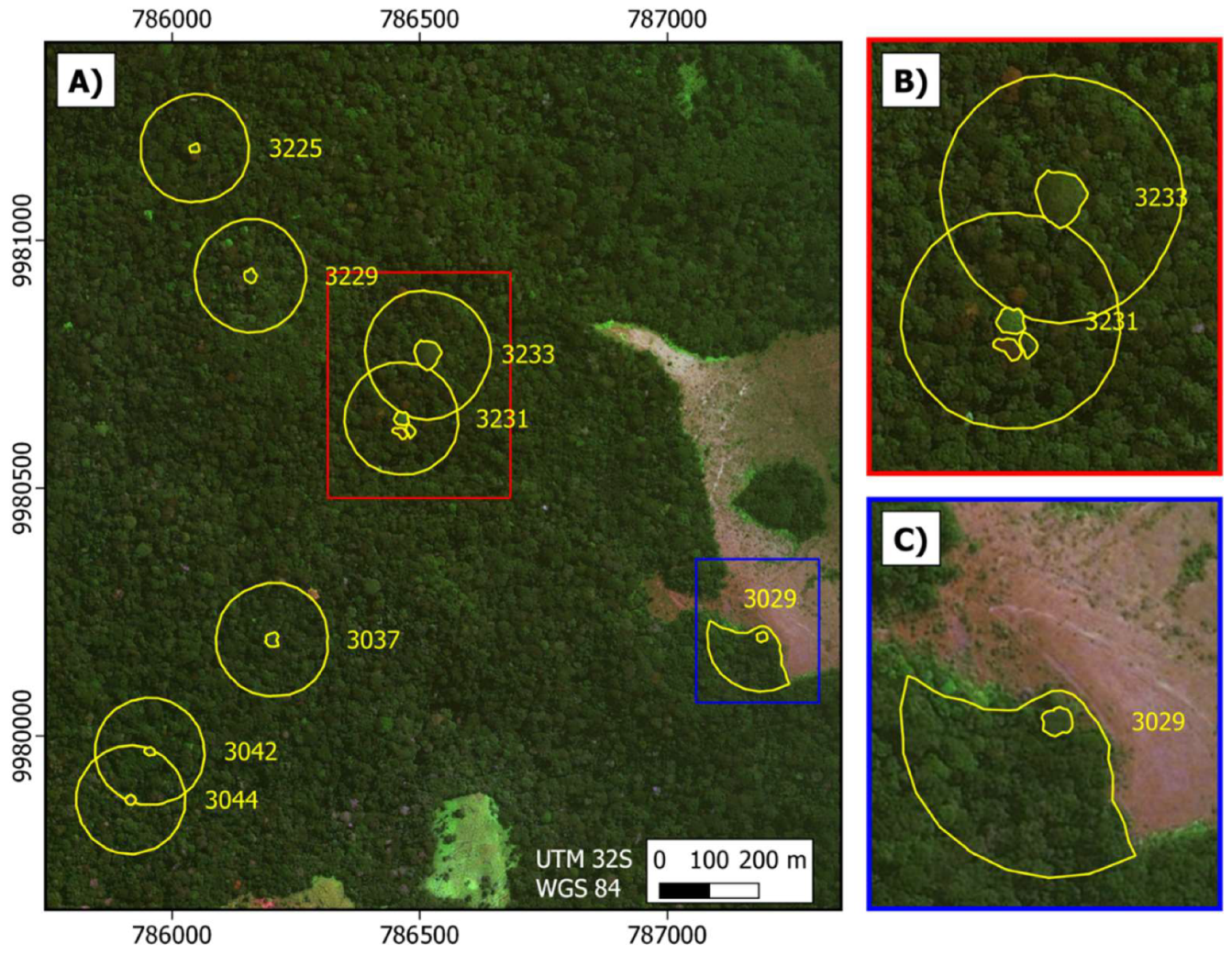
2.1.1. Ground Observations of Moabi Phenology at Lopé NP
2.1.2. Satellite Observations of Moabi Phenology at Lopé NP
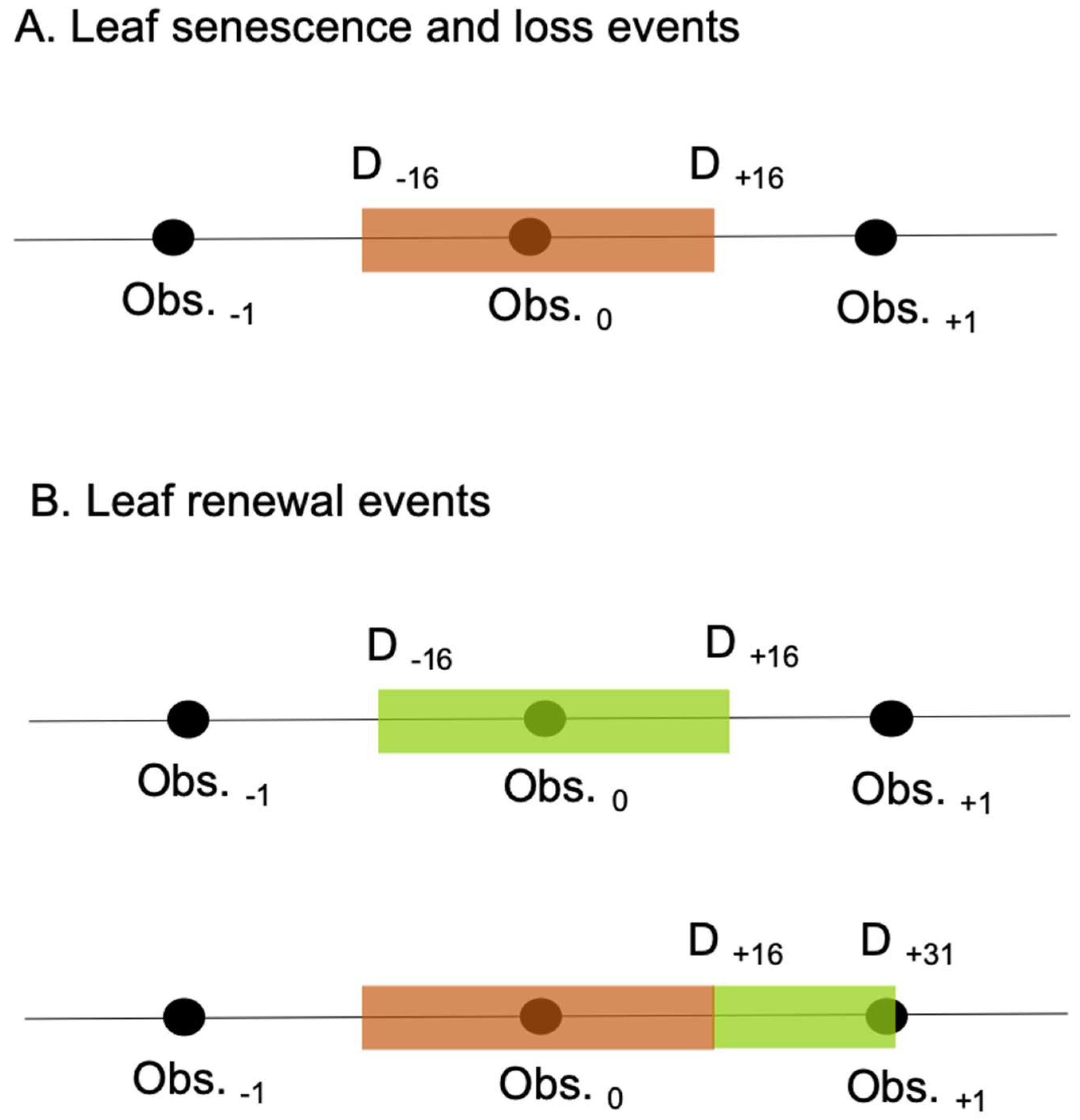
2.2. Analyses
- Model 1: Leaf senescence and loss event ~ VV (canopy) + (1|TreeID) + (1|Year)
- Model 2: Leaf senescence and loss event ~ VV (normalised canopy) + (1|TreeID) + (1|Year)
- Model 3: Leaf senescence and loss event ~ VH (canopy) + (1|TreeID) + (1|Year)
- Model 4: Leaf senescence and loss event ~ VH (normalised canopy) + (1|TreeID) + (1|Year)
- Model 5: Leaf senescence and loss event ~ NDVI (canopy) + (1|TreeID) + (1|Year)
- Model 6: Leaf senescence and loss event ~ NDVI (normalised canopy) + (1|TreeID) + (1|Year)
- Model 7: Leaf senescence and loss event ~ GLI (canopy) + (1|TreeID) + (1|Year)
- Model 8: Leaf senescence and loss event ~ GLI (normalised canopy) + (1|TreeID) + (1|Year)
- Model 9: Leaf renewal event ~ NDVI (canopy) + (1|TreeID) + (1|Year)
- Model 10: Leaf renewal event ~ NDVI (normalised canopy) + (1|TreeID) + (1|Year)
- Model 11: Leaf renewal event ~ GLI (canopy) + (1|TreeID) + (1|Year)
- Model 12: Leaf renewal event ~ GLI (normalised canopy) + (1|TreeID) + (1|Year)
3. Results
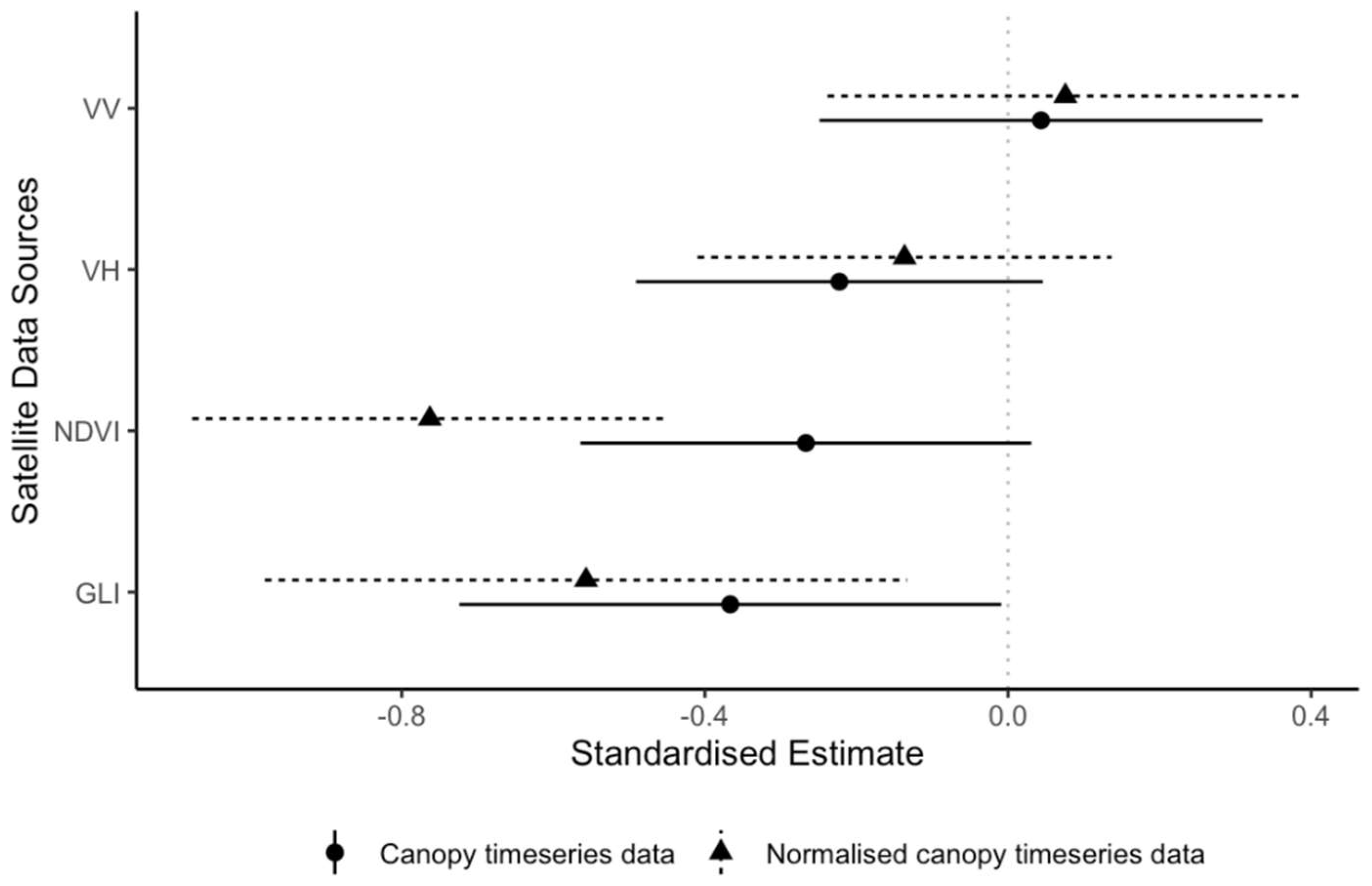
3.1. Leaf Senescence and Loss
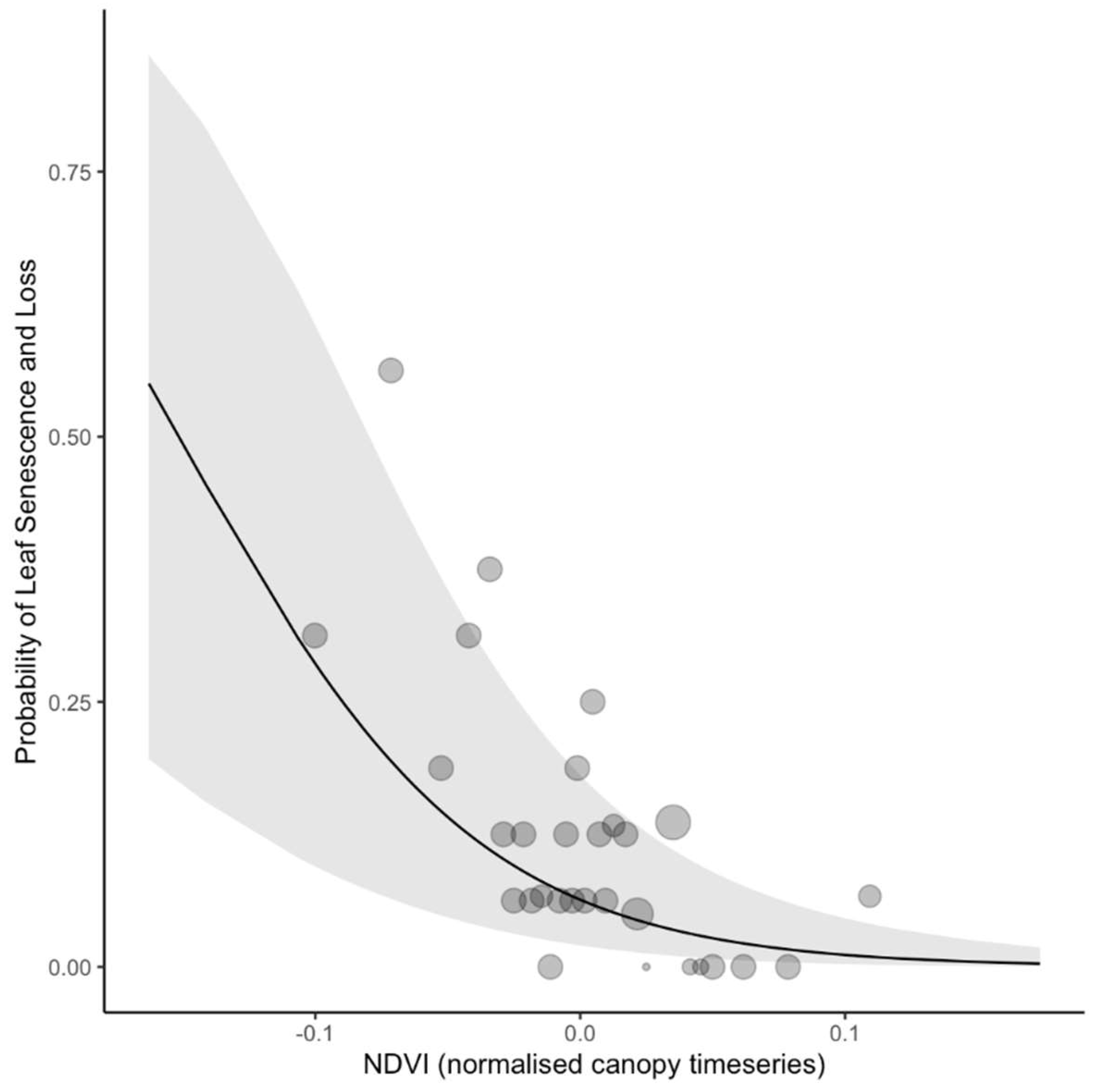
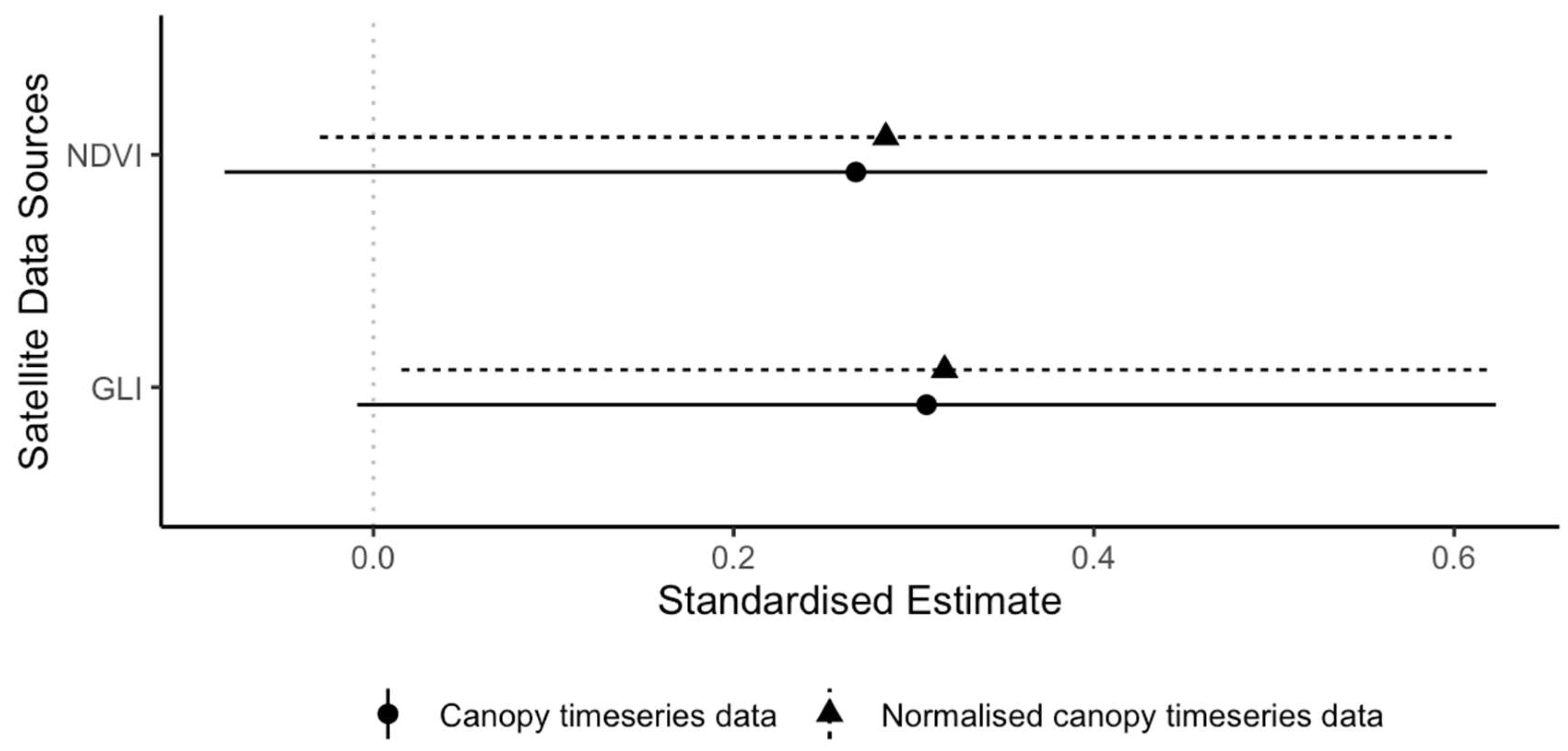
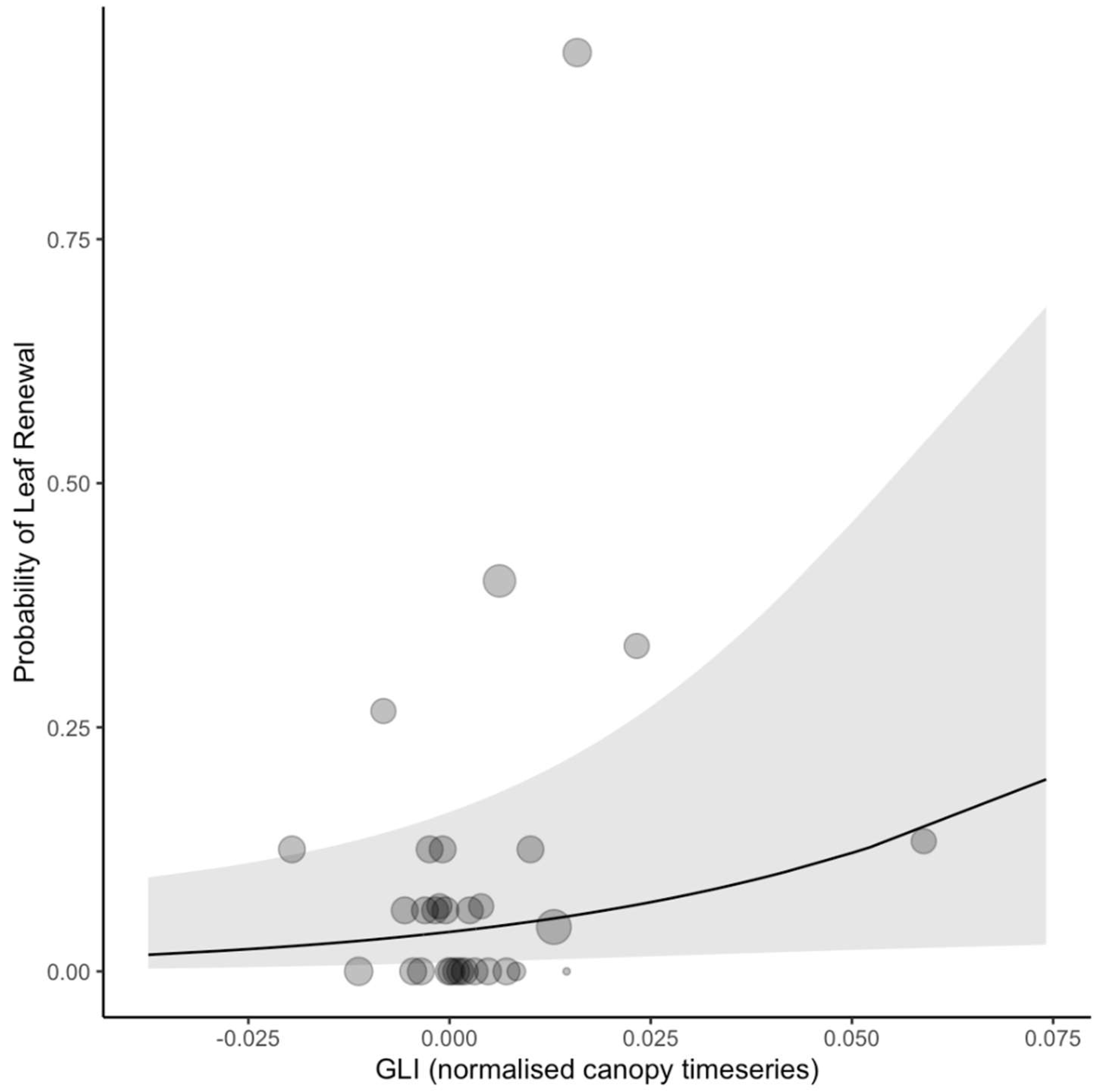
3.2. Leaf Renewal
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Abernethy, K.; Bush, E.R.; Forget, P.M.; Mendoza, I.; Morellato, L.P.C. Current issues in tropical phenology: A synthesis. Biotropica 2018, 50, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G.B.; Doney, S.C. Climate, ecosystems, and planetary futures: The challenge to predict life in Earth system models. Science 2018, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchard, E.T.A. The tropical forest carbon cycle and climate change. Nature 2018, 559, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.A.; Exbrayat, J.F.; Van Der Velde, I.R.; Feng, L.; Williams, M. The decadal state of the terrestrial carbon cycle: Global retrievals of terrestrial carbon allocation, pools, and residence times. Proc. Natl. Acad. Sci. USA 2016, 113, 1285–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Kobayashi, H.; Stark, S.C.; Meng, R.; Guan, K.; Tran, N.N.; Gao, S.; Yang, W.; Restrepo-Coupe, N.; Miura, T.; et al. Biological processes dominate seasonality of remotely sensed canopy greenness in an Amazon evergreen forest. New Phytol. 2018, 217, 1507–1520. [Google Scholar] [CrossRef] [Green Version]
- Morton, D.C.; Nagol, J.; Carabajal, C.C.; Rosette, J.; Palace, M.; Cook, B.D.; Vermote, E.F.; Harding, D.J.; North, P.R.J. Amazon forests maintain consistent canopy structure and greenness during the dry season. Nature 2014, 506, 221. [Google Scholar] [CrossRef]
- Huete, A.R.; Saleska, S.R. Remote sensing of tropical forest phenology: Issues and controversies. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2010, 38, 539–541. [Google Scholar]
- Albert, L.P.; Restrepo-Coupe, N.; Smith, M.N.; Wu, J.; Chavana-Bryant, C.; Prohaska, N.; Taylor, T.C.; Martins, G.A.; Ciais, P.; Mao, J.; et al. Cryptic phenology in plants: Case studies, implications, and recommendations. Glob. Chang. Biol. 2019, 25, 3591–3608. [Google Scholar] [CrossRef]
- Park, J.Y.; Muller-Landau, H.C.; Lichstein, J.W.; Rifai, S.W.; Dandois, J.P.; Bohlman, S.A. Quantifying leaf phenology of individual trees and species in a tropical forest using unmanned aerial vehicle (UAV) images. Remote Sens. 2019, 11, 1534. [Google Scholar] [CrossRef] [Green Version]
- Alberton, B.; Almeida, J.; Helm, R.; Torres, R.; da, S.; Menzel, A.; Morellato, L.P.C. Using phenological cameras to track the green up in a cerrado savanna and its on-the-ground validation. Ecol. Inform. 2014, 19, 62–70. [Google Scholar] [CrossRef]
- Nagai, S.; Ichie, T.; Yoneyama, A.; Kobayashi, H.; Inoue, T.; Ishii, R.; Suzuki, R.; Itioka, T. Usability of time-lapse digital camera images to detect characteristics of tree phenology in a tropical rainforest. Ecol. Inform. 2016, 32, 91–106. [Google Scholar] [CrossRef]
- Louppe, D. Baillonella toxisperma Pierre. [Internet] Record from PROTA4U. In PROTA (Plant Resources of Tropical Africa / Ressources végétales de l’Afrique tropicale). 2005. Available online: https://www.prota4u.org/database/protav8.asp?h=M10,M14,M15,M16,M18,M20,M23,M25,M26,M27,M36,M5,M6,M7,M8,M9&t=moabi,Moabi&p=Baillonella+toxisperma#AdulterationsAndSubstitutes (accessed on 28 January 2020).
- Malhi, Y.; Jackson, T.; Bentley, L.P.; Lau, A.; Shenkin, A.; Herold, M.; Calders, K.; Bartholomeus, H.; Disney, M.I. New perspectives on the ecology of tree structure and tree communities through terrestrial laser scanning. Interface Focus 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plenderleith, K.; Brown, N. Moabi (Baillonella toxisperma). In The Key Non-Timber Forest Products of Central Africa: State of the Knowledge; Clark, L.E., Sunderland, T.C.H., Eds.; USAID, Bureau for Africa, Office of Sustainable Development: Washington, DC, USA, 2004; pp. 141–162. [Google Scholar]
- Bush, E.R. Tropical Phenology in a Time of Change. 2018. Available online: https://dspace.stir.ac.uk/handle/1893/29325#.XjpjvyMRVPZ (accessed on 27 January 2020).
- Philippon, N.; Cornu, G.; Monteil, L.; Gond, V.; Moron, V.; Pergaud, J.; Sèze, G.; Bigot, S.; Camberlin, P.; Doumenge, C.; et al. The light-deficient climates of western Central African evergreen forests. Environ. Res. Lett. 2019, 14, 034007. [Google Scholar] [CrossRef]
- Bush, E.R.; Jeffery, K.; Bunnefeld, N.; Tutin, C.; Musgrave, R.; Moussavou, G.; Mihindou, V.; Malhi, Y.; Lehmann, D.; Ndong, J.E.; et al. Ground data confirm warming and drying are at a critical level for forest survival in western equatorial Africa. PeerJ Prepr. 2019. [Google Scholar] [CrossRef]
- Tutin, C.E.G. Gorillas and their food plants in the Lope Reserve, Gabon. In Chorology, Taxonomy and Ecology of the Floras of Africa and Madagascar; Kew Publication: London, UK, 1998; pp. 227–243. [Google Scholar]
- Bush, E.R.; Abernethy, K.A.; Jeffery, K.; Tutin, C.; White, L.; Dimoto, E.; Dikangadissi, J.T.; Jump, A.S.; Bunnefeld, N. Fourier analysis to detect phenological cycles using tropical field data and simulations. Methods Ecol. Evol. 2017, 8, 530–540. [Google Scholar] [CrossRef]
- Bush, E.R.; Bunnefeld, N.; Dimoto, N.; Dikangadissi, J.T.; Jeffery, K.; Tutin, C.; White, L.; Abernethy, K.A. Towards effective monitoring of tropical phenology: Maximising returns and reducing uncertainty in long-term studies. Biotropica 2018, 50, 455–464. [Google Scholar] [CrossRef]
- Myneni, R.; Knyazikhin, Y.; Park, T. MCD15A2H MODIS/Terra+Aqua Leaf Area Index/FPAR 8-day L4 Global 500m SIN Grid V006. NASA EOSDIS L. Process. DAAC 2015. Available online: http://doi.org/10.5067/MODIS/MCD15A3H.006 (accessed on 28 January 2020).
- Knyazikhin, Y.; Glassy, J.; Privette, J.L.; Tian, Y.; Lotsch, A.; Zhang, Y.; Wang, Y.; Morisette, J.T.; Votava, P.; Myneni, R.B.; et al. MODIS Leaf Area Index (LAI) And Fraction Of Photosynthetically Active Radiation Absorbed By Vegetation (FPAR) Product (MOD15) Algorithm Theoretical Basis Document. 1999. Available online: https://modis.gsfc.nasa.gov/data/atbd/atbd_mod15.pdf (accessed on 28 January 2020).
- Wang, Q.; Adiku, S.; Tenhunen, J.; Granier, A. On the relationship of NDVI with leaf area index in a deciduous forest site. Remote Sens. Environ. 2005, 94, 244–255. [Google Scholar] [CrossRef]
- Richardson, A.D. Tracking seasonal rhythms of plants in diverse ecosystems with digital camera imagery. New Phytol. 2019, 222, 1742–1750. [Google Scholar] [CrossRef] [Green Version]
- Alberton, B.; Torres, S.; Cancian, L.F.; Borges, B.D.; Almeida, J.; Mariano, G.C.; Patricia, L.; Morellato, C. Introducing digital cameras to monitor plant phenology in the tropics: Applications for conservation. Perspect. Ecol. Conserv. 2017, 15, 82–90. [Google Scholar] [CrossRef]
- Hunt, E.R.; Doraiswamy, P.C.; McMurtrey, J.E.; Daughtry, C.S.T.; Perry, E.M.; Akhmedov, B. A visible band index for remote sensing leaf chlorophyll content at the Canopy scale. Int. J. Appl. Earth Obs. Geoinf. 2012, 21, 103–112. [Google Scholar] [CrossRef] [Green Version]
- De Moura, Y.M.; Galvão, L.S.; Hilker, T.; Wu, J.; Saleska, S.; do Amaral, C.H.; Nelson, B.W.; Lopes, A.P.; Wiedeman, K.K.; Prohaska, N.; et al. Spectral analysis of amazon canopy phenology during the dry season using a tower hyperspectral camera and modis observations. ISPRS J. Photogramm. Remote Sens. 2017, 131, 52–64. [Google Scholar] [CrossRef]
- Louhaichi, M.; Borman, M.M.; Johnson, D.E. Spatially located platform and aerial photography for documentation of grazing impacts on wheat. Geocarto Int. 2001, 16, 65–70. [Google Scholar] [CrossRef]
- Macfarlane, C.; Ogden, G.N. Automated estimation of foliage cover in forest understorey from digital nadir images. Methods Ecol. Evol. 2012, 3, 405–415. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant remote sensing vegetation indices: A review of developments and applications. J. Sensors 2017, 2017, 1353691. [Google Scholar] [CrossRef] [Green Version]
- Morcillo-Pallarés, P.; Rivera-Caicedo, J.P.; Belda, S.; De Grave, C.; Burriel, H.; Moreno, J.; Verrelst, J. Quantifying the Robustness of Vegetation Indices through Global Sensitivity Analysis of Homogeneous and Forest Leaf-Canopy Radiative Transfer Models. Remote Sens. 2019, 11, 2418. [Google Scholar] [CrossRef] [Green Version]
- Hunt, E.R.; Daughtry, C.S.T.; Eitel, J.U.H.; Long, D.S. Remote sensing leaf chlorophyll content using a visible band index. Agron. J. 2011, 103, 1090–1099. [Google Scholar] [CrossRef] [Green Version]
- Proisy, C.; Mougin, E.; Dufrêne, E.; Dantec, V. Le Monitoring seasonal changes of a mixed temperate forest using ERS SAR observations. IEEE Trans. Geosci. Remote Sens. 2000, 38, 540–552. [Google Scholar] [CrossRef]
- Tanase, M.A.; Villard, L.; Pitar, D.; Apostol, B.; Petrila, M.; Chivulescu, S.; Leca, S.; Borlaf-Mena, I.; Pascu, I.S.; Dobre, A.C.; et al. Synthetic aperture radar sensitivity to forest changes: A simulations-based study for the Romanian forests. Sci. Total Environ. 2019, 689, 1104–1114. [Google Scholar] [CrossRef]
- Ju, J.; Roy, D.P. The availability of cloud-free Landsat ETM+ data over the conterminous United States and globally. Remote Sens. Environ. 2008, 112, 1196–1211. [Google Scholar] [CrossRef]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P.; et al. Sentinel-2: ESA’s Optical High-Resolution Mission for GMES Operational Services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Torres, R.; Snoeij, P.; Geudtner, D.; Bibby, D.; Davidson, M.; Attema, E.; Potin, P.; Rommen, B.Ö.; Floury, N.; Brown, M.; et al. GMES Sentinel-1 mission. Remote Sens. Environ. 2012, 120, 9–24. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 27 January 2020).
- Kellner, J.R.; Albert, L.P.; Burley, J.T.; Cushman, K.C. The case for remote sensing of individual plants. Am. J. Bot. 2019, 106, 1139–1142. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Obs. | Canopy Time Series | Buffer Time Series | Normalized Canopy Time Series |
|---|---|---|---|---|
| Sentinel-1 VV | 837 | 0.18 (0.07) | 0.22 (0.04) | –0.04 (0.07) |
| Sentinel-1 VH | 837 | 0.04 (0.02) | 0.05 (0.01) | –0.01 (0.02) |
| Sentinel-2 NDVI | 477 | 0.44 (0.19) | 0.43 (0.18) | 0 (0.04) |
| Sentinel-2 GLI | 477 | 0.02 (0.03) | 0.01 (0.02) | 0 (0.01) |
| Model | Predictor | Est. | SE | Z | P |
|---|---|---|---|---|---|
| 1 | VV (canopy) | 0.04 | 0.15 | 0.29 | 0.77 |
| 2 | VV (normalized canopy) | 0.08 | 0.16 | 0.47 | 0.64 |
| 3 | VH (canopy) | −0.22 | 0.14 | −1.62 | 0.10 |
| 4 | VH (normalized canopy) | −0.14 | 0.14 | −0.98 | 0.33 |
| 5 | NDVI (canopy) | −0.27 | 0.15 | −1.76 | 0.08 |
| 6 | NDVI (normalized canopy) | −0.76 | 0.16 | −4.77 | <0.01 |
| 7 | GLI (canopy) | −0.37 | 0.18 | −2.01 | 0.04 |
| 8 | GLI (normalized canopy) | −0.56 | 0.22 | −2.58 | 0.01 |
| Model | Predictor | Est. | SE | Z | P |
|---|---|---|---|---|---|
| 9 | NDVI (canopy) | 0.27 | 0.18 | 1.50 | 0.13 |
| 10 | NDVI (normalized canopy) | 0.28 | 0.16 | 1.78 | 0.08 |
| 11 | GLI (canopy) | 0.31 | 0.16 | 1.90 | 0.06 |
| 12 | GLI (normalized canopy) | 0.32 | 0.15 | 2.06 | 0.04 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bush, E.R.; Mitchard, E.T.A.; Silva, T.S.F.; Dimoto, E.; Dimbonda, P.; Makaga, L.; Abernethy, K. Monitoring Mega-Crown Leaf Turnover from Space. Remote Sens. 2020, 12, 429. https://doi.org/10.3390/rs12030429
Bush ER, Mitchard ETA, Silva TSF, Dimoto E, Dimbonda P, Makaga L, Abernethy K. Monitoring Mega-Crown Leaf Turnover from Space. Remote Sensing. 2020; 12(3):429. https://doi.org/10.3390/rs12030429
Chicago/Turabian StyleBush, Emma R., Edward T. A. Mitchard, Thiago S. F. Silva, Edmond Dimoto, Pacôme Dimbonda, Loïc Makaga, and Katharine Abernethy. 2020. "Monitoring Mega-Crown Leaf Turnover from Space" Remote Sensing 12, no. 3: 429. https://doi.org/10.3390/rs12030429
APA StyleBush, E. R., Mitchard, E. T. A., Silva, T. S. F., Dimoto, E., Dimbonda, P., Makaga, L., & Abernethy, K. (2020). Monitoring Mega-Crown Leaf Turnover from Space. Remote Sensing, 12(3), 429. https://doi.org/10.3390/rs12030429