1. Introduction
Potato late blight, caused by
Phytophthora infestans, is a significant disease worldwide that has a significant economic impact on potato crops, which is estimated to be over US
$5.45 million per year. Such impact is due to the capacity of the agent to destroy the plants and tubers rapidly and to propagate to large areas during the growing season thanks to the production of secondary inoculum [
1,
2]. There is, therefore, the need to detect the disease for avoiding its rapid propagation [
3]. Field scouting and continuous preventive pesticide applications have been adopted to prevent yield losses, but these strategies are either time-consuming or expensive, and they are not environmentally friendly. An alternative is to use imagery having the appropriate temporal and spatial scale to detect the disease at early stages by sensing the changes in chemical and physical properties of the crop [
4]. Indeed, late blight causes chlorosis that can be detected in the visible region of the spectra (400 to 700 nm) [
5]. The disease can also change the leaf structure, which is detectable in the near-infrared spectral region (700 to 1300 nm) [
6,
7].
One particular spectral region in the visible domain is the red region, which corresponds to one of the two chlorophyll absorption bands [
8]. Another spectra domain is the red-edge region, a transition zone from the red to the near-infrared region, which is usually located between 660 to 780 nm [
9]. Such a transition zone is due to the maximal chlorophyll absorbance in the red wavelength and the high reflectance in the near-infrared wavelengths because of multiple scatterings of the near-infrared radiation by the leaf mesophyll [
10]. According to Pu et al. [
11], two optical parameters related to the red-edge region can be defined: the red-well position (RWP or R0) and the red-edge inflection point (REP). RWP is the wavelength in the red region between 660 and 680 nm that corresponds to the minimum reflectance because of maximum chlorophyll absorption. REP is the wavelength corresponding to the inflection point of the spectral curve between the red and near-infrared spectral domains. At this wavelength, the reflectance slope in the red-edge region is maximal [
12]. Changes in the RWP and REP can be related to the presence of potato late blight. Indeed, this disease is caused by a hemibiotrophic pathogen that affects the leaf chemical composition, specifically its chlorophyll content, and induces changes in the internal leaf structure that can be detected using the near-infrared reflectance [
13,
14]. Shifts of the REP to longer or shorter wavelengths have already been related to changes in the chemical and morphological plant status [
15].
In the literature, there are several studies that related band reflectances or vegetation indices to late blight occurrence in potato or tomato crops [
16,
17,
18,
19,
20,
21,
22], but none of them tested the use of RWP and REP to detect the disease. All of them but Fernández et al. [
21] and Gold et al. [
22] were based on one or two dates of experimentations without reporting the moment of inoculation, and thus they were not able to determine the time after inoculation when the disease could be detected with spectral information. This study aims to improve the understanding of the spectral changes of RWP and REP at the leaf and canopy levels due to late blight disease on potato plants from the moment of inoculation. In particular, we modeled the changes in RWP and REP at the leaf and canopy levels as a function of the days post-inoculation (DPI). We also developed a Support Vector Machine (SVM) algorithm [
23] to sort healthy and infected leaves or plants by using two types of input features: i) the wavelengths corresponding to RWP or REP; and (ii) the reflectances of the red and red-edge bands of the Micasense
® Dual-X camera (Micasense, Inc., Seattle, WA, USA). The results of this study will be used to test UAV images acquired with this camera for detecting late blight disease over potato fields.
2. Materials and Methods
2.1. Experiment
The study used data that were acquired as part of the experiment, which is detailed in Fernández et al. [
21]. We will here summarize the main steps of the experiment. The experiment used two groups (healthy and infected) of eight Shepody potato plants grown in a walk-in growth chamber located at the Biotron facilities of the University of Western Ontario (London, ON, Canada). The chamber has a constant relative humidity of 65%, a photoperiod of 12 h at 23 °C, followed by a dark period of 12 h at 18 °C. The infected plants were inoculated by spraying at the flowering phenological stage 200 µL of a sporangia suspension of
Phytophthora infestans at the concentration of 1 × 10
5 sporangia mL
−1 on marked primary leaflets. A total of 101 leaflets were inoculated. To follow the spectral changes of each leaflet by acquiring spectra over the same leaflets, we marked the healthy and inoculated leaflets at the base of the petiole with a small piece of white tape which was numbered. Control (healthy) and infected leaflets were chosen from the middle and the top of the canopy so that they were larger than the leaf clip measurement area to avoid low reflectance measurements when the leaves were too small [
21]. Healthy plants were placed in a separate chamber to avoid cross-contamination.
Radiances between 350 and 2500 nm at 1.4 nm sampling intervals with a 25° field of view (FOV) bare fiber optic cable were measured at the leaf and canopy levels three hours before inoculation, until six days post-inoculation (DPI), with an ASD FieldSpec PRO FR spectroradiometer (ASD Inc., Boulder, CO, USA). Both the leaf and canopy spectra were acquired and calibrated using the RS3 version 6.4 software from ASD (ASD Inc., Boulder, CO, USA). The leaf-level reflectance spectra were acquired at the center of the marked leaflets of healthy and infected plants using a leaf clip, with a white Goretex (99% reflectance) and a black reference, attached to an ASD high-intensity probe equipped with a light source and a fiber optic cable. For each experiment day, we acquired a total of 101 spectra over the infected leaves and 124 spectra over the healthy leaves.
The canopy-level spectra were acquired on each plant by placing the 25° FOV bare fiber optic cable at 11 cm from the top of the canopy (
Figure 1). Plants were illuminated with two ASD halogen lamps (ASD Inc., Boulder, CO, USA), each of them angled at 45° from the horizontal line and located on each side of the plant. Canopy spectra were acquired over eight healthy and eight infected plants at each day of the experiment. For each plant, the canopy reflectance was measured four times after rotating the plant 90°. This allowed obtaining a total of 32 healthy and 32 infected canopy reflectance spectra at each day of the experiment. The measured plant area was 82.83 cm
2 according to Equation (1) as reported by Fernández et al. [
21]:
where
A = Measured plant area at the top of the plant canopy (in cm2);
FOV = field of view angle of the optic fiber (= 25°);
h = distance between the top of the canopy and the fiber optic sensor (= 11 cm).
2.2. Methodology
All data were processed using MATLAB R2019a (MathWorks, Inc., Natick, MA, USA). The spectra collected at the leaf and canopy levels from the healthy and infected plants were first cropped to the 400 to 900 nm range. Secondly, the raw spectra were converted to reflectance spectra using the ViewSpec Pro version 6.2.0 software (ASD Inc., Boulder, CO, USA). These reflectance spectra were then subjected to a Savitzky–Golay [
24] filtering method to reduce instrumental noise and to a multiplicative scatter correction (MSC) to remove additive and multiplicative scattering [
25]. The filtered reflectance spectra were used to compute the first-order derivative (FD) of the reflectance spectra that represents the signal change between two adjacent wavelengths [
26]. The FD is computed as follows [
27]:
where
FD (λ) = first-order derivative of the reflectance spectrum at wavelength λ;
R = reflectance at specified wavelengths λ + Δλ; or λ − Δλ;
λ = wavelength of the spectrum (nm);
Δλ; = difference between two successive wavelengths, i.e., 1.4 nm.
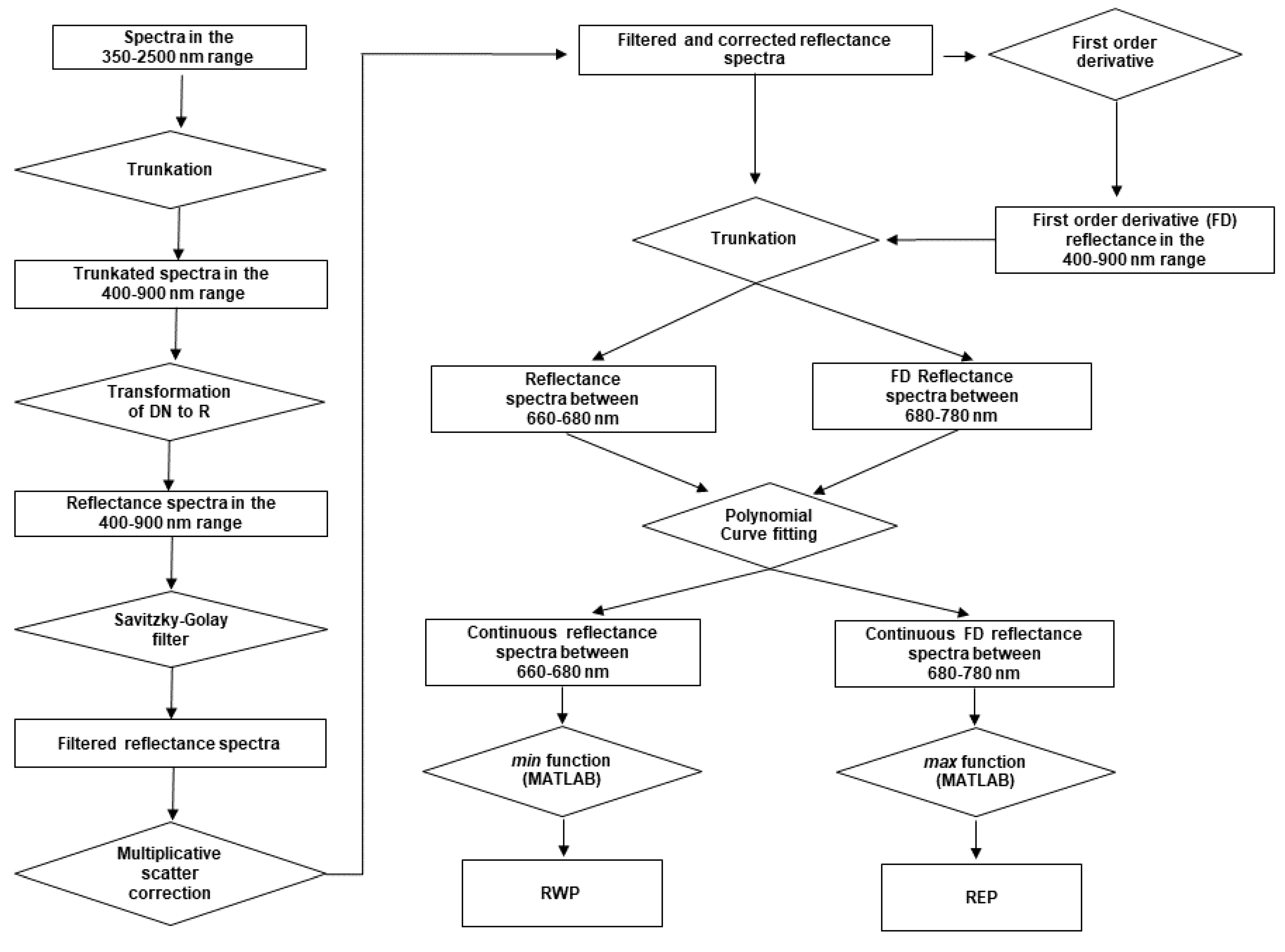
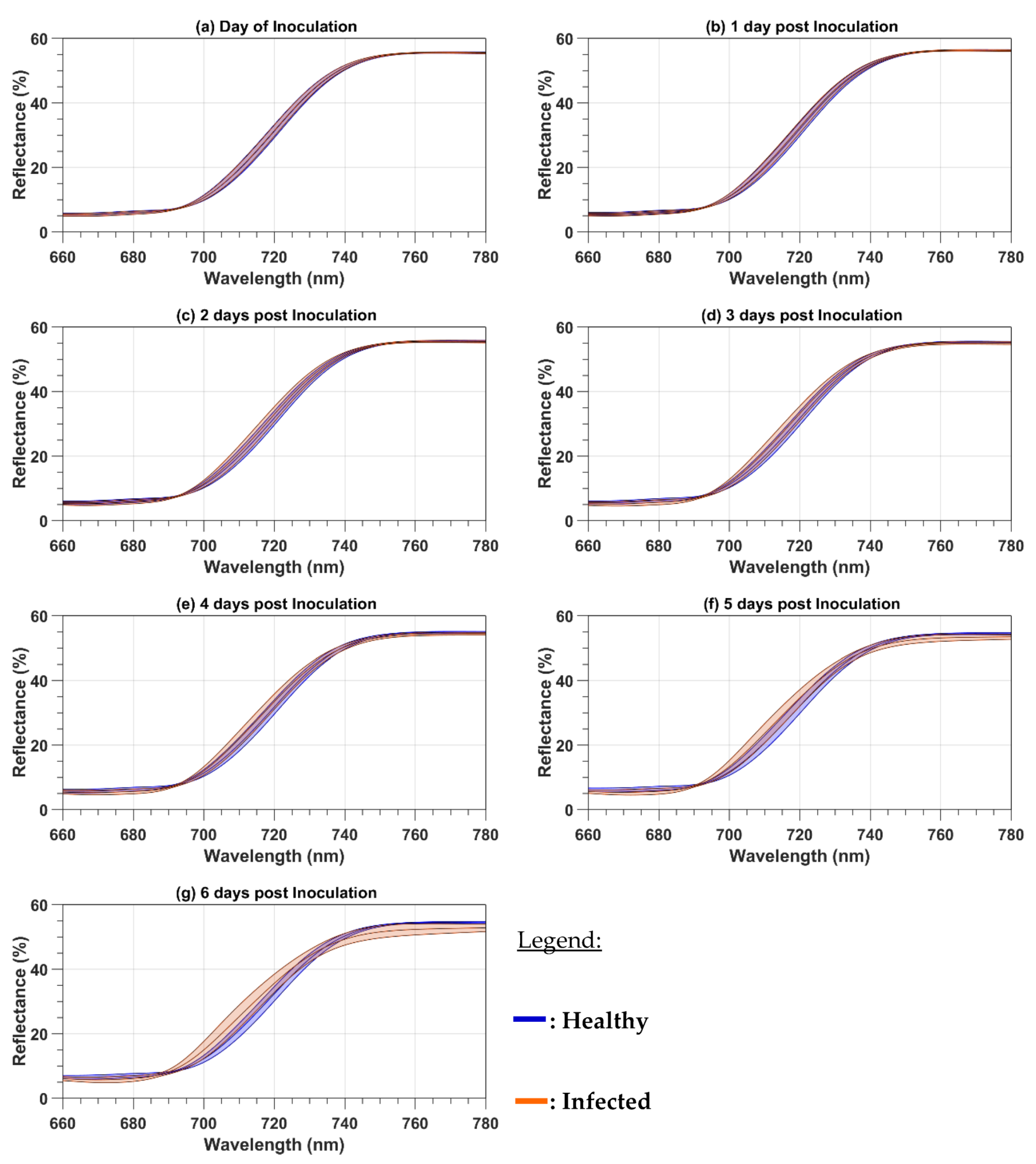
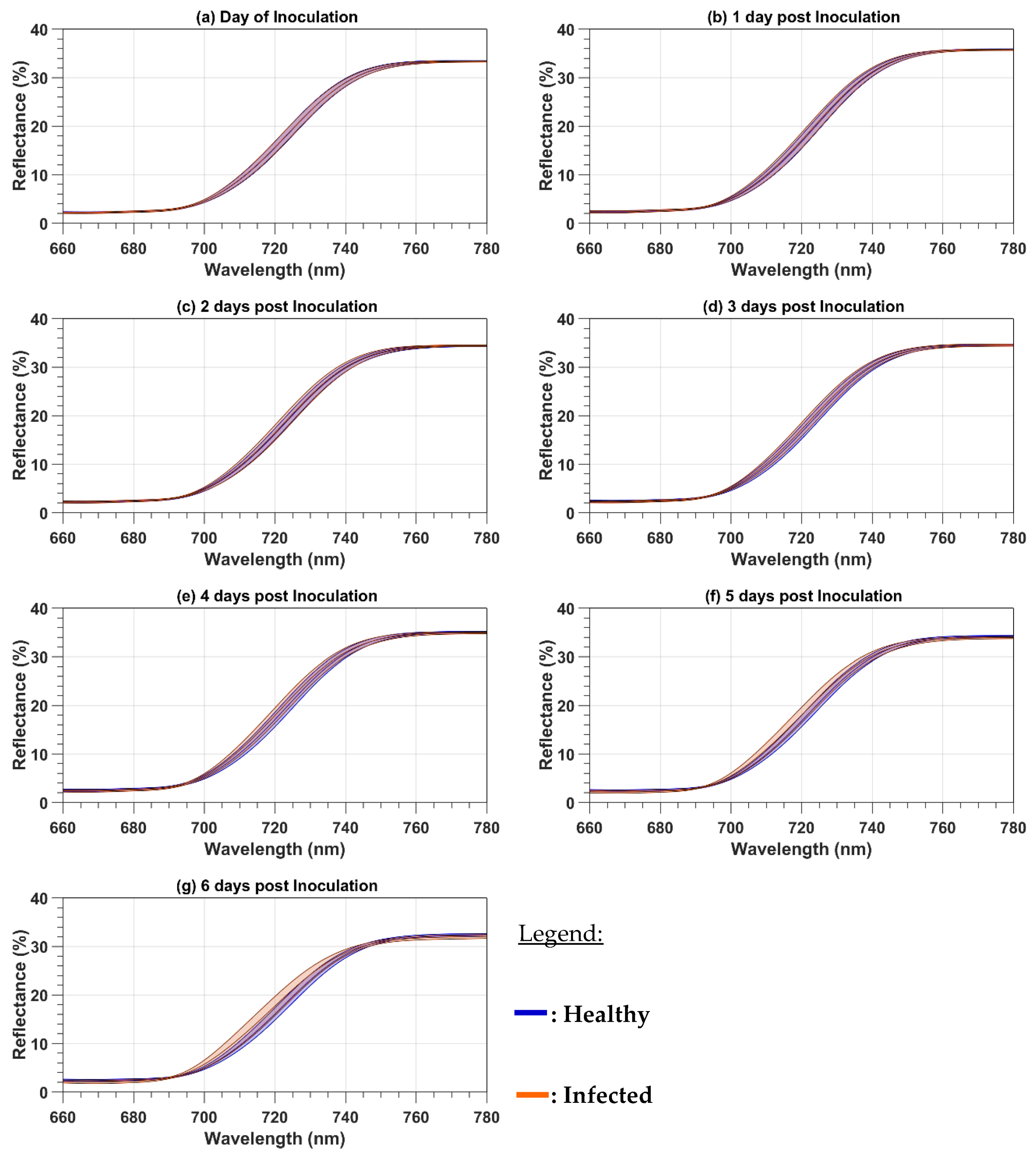
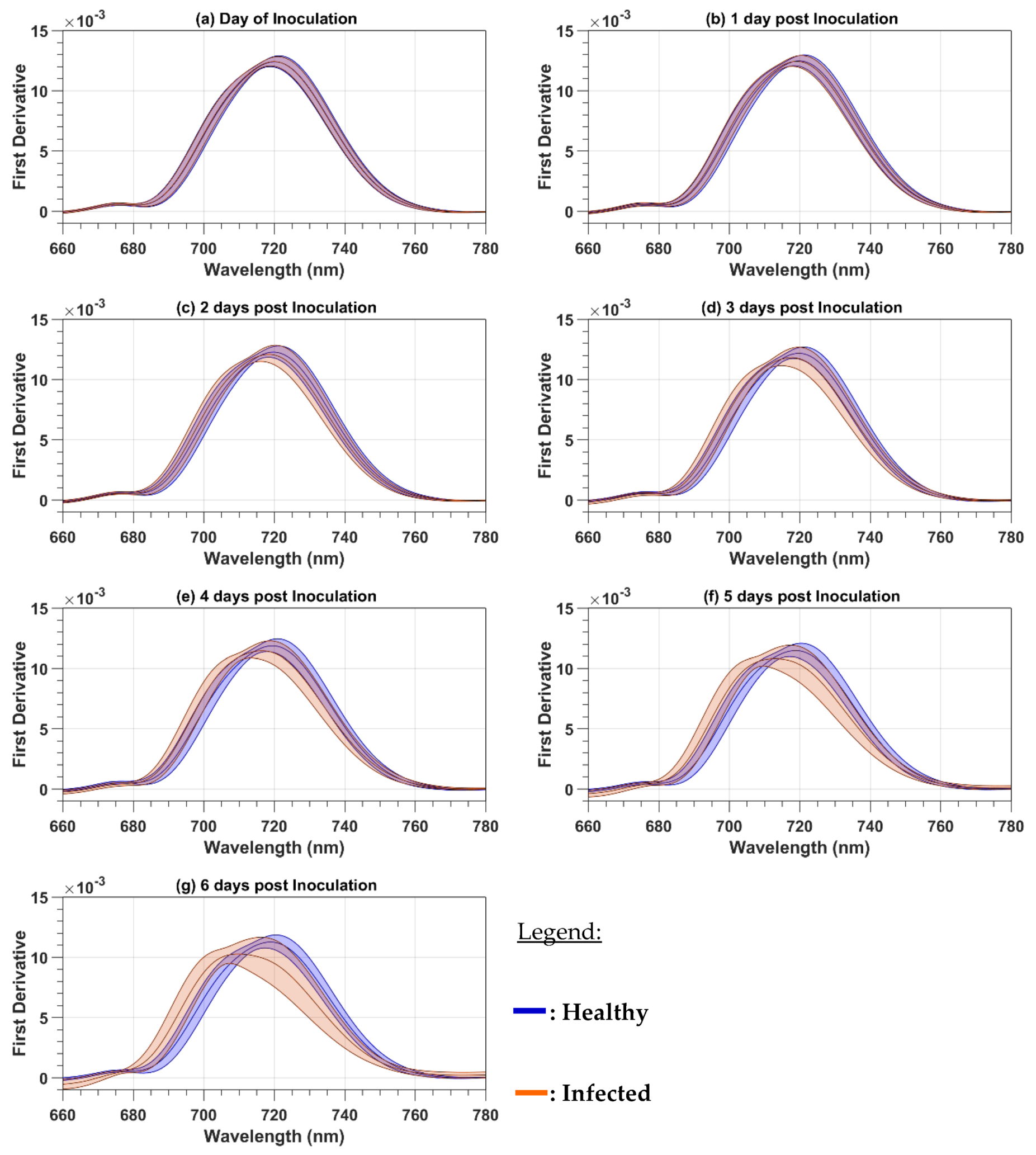
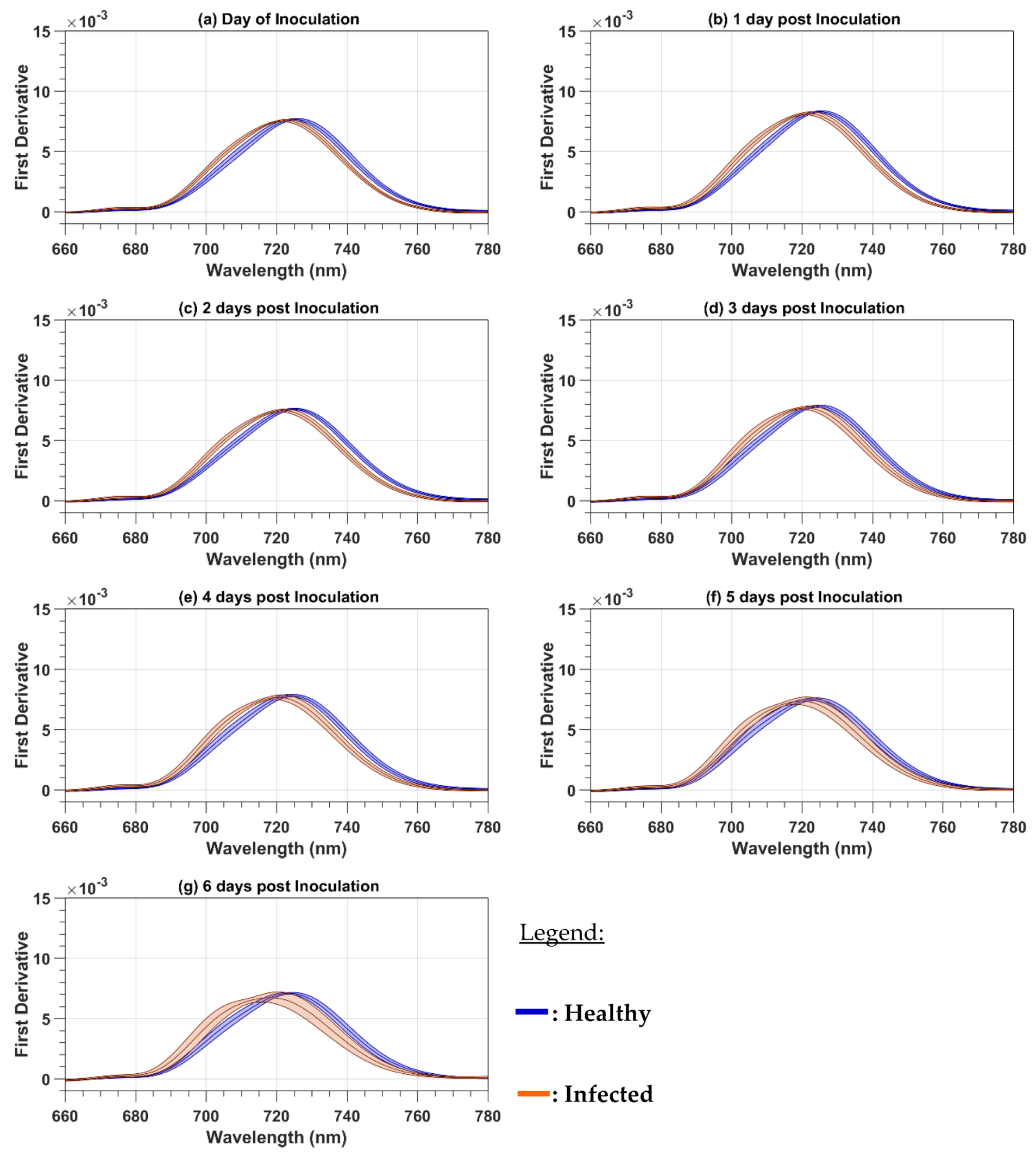
Because the paper focuses on the red and red-edge regions, both the reflectance and the first-order derivative spectra were cropped to the 660 to 780 nm spectral range. The mean cropped spectra were plotted to observe the spectral variations as a function of the DPI. The mean reflectance spectra were calculated as a simple average of all 124 (101) healthy (infected) leaf spectra and all the 32 healthy or infected canopy spectra. With the spectra acquired on both the leaf and canopy levels, two spectral parameters were computed for every day of evaluation from 0 to 6 DPI. The first one is the red-well point (RWP), which corresponds to the wavelengths having the minimum reflectance value in the red region [
11]. The methodology to compute the RWP involved three steps (
Figure 2). First, the reflectances in the red region, i.e., between 660 and 680 nm, of each spectrum were selected. Second, a curve was fitted to the reflectance values to make them continuous. Finally, since the RWP is the wavelength corresponding to the minimum reflectance value, it was determined by applying the
min function of MATLAB R2019a (MathWorks, Inc., Natick, MA, USA) to the fitted curve to retrieve the minimum reflectance and then the corresponding wavelength. The second parameter is the red-edge point (REP), which is the wavelength of the inflection point of the reflectance spectra in the red-edge region, i.e., between 680 and 780 nm [
28]. REP is, therefore, the wavelength corresponding to the maximum value of the FD spectra. This value was determined as follows (
Figure 2). First, the FD spectra were truncated to the red-edge region, and a curve was fitted over this spectral range to make the FD data continuous. The maximum of the fitted curve was then determined by using the
max function of MATLAB R2019a. For both REP and RWP, the curve has a parabolic shape that is modeled using a second-order polynomial function.
With the computed RWP and REP wavelengths for each DPI, a
t-test means comparison was used to analyze the difference of the RWP and REP values between healthy and infected cases as a function of the DPI. A
t-test mean comparison usually assumes that the data have a normal distribution and that the variance is equal, but Posten (1984) [
29] showed that such a
t-test is highly robust to deviations from the normality assumption. The normality assumption for the RWP and REP data distribution was visually assessed through histograms graphed with both the leaf- and canopy-level values. The equal variance assumption was tested with a two-sample F-test (
vartest2 function of MATLAB R2019a), which returned an acceptance of the null hypothesis. The
t-test was computed using RWP and REP values from 124 and 101 healthy and infected leaves. At the canopy level, the
t-test was computed using for RWP and REP data from 32 healthy and 32 infected spectral measurements.
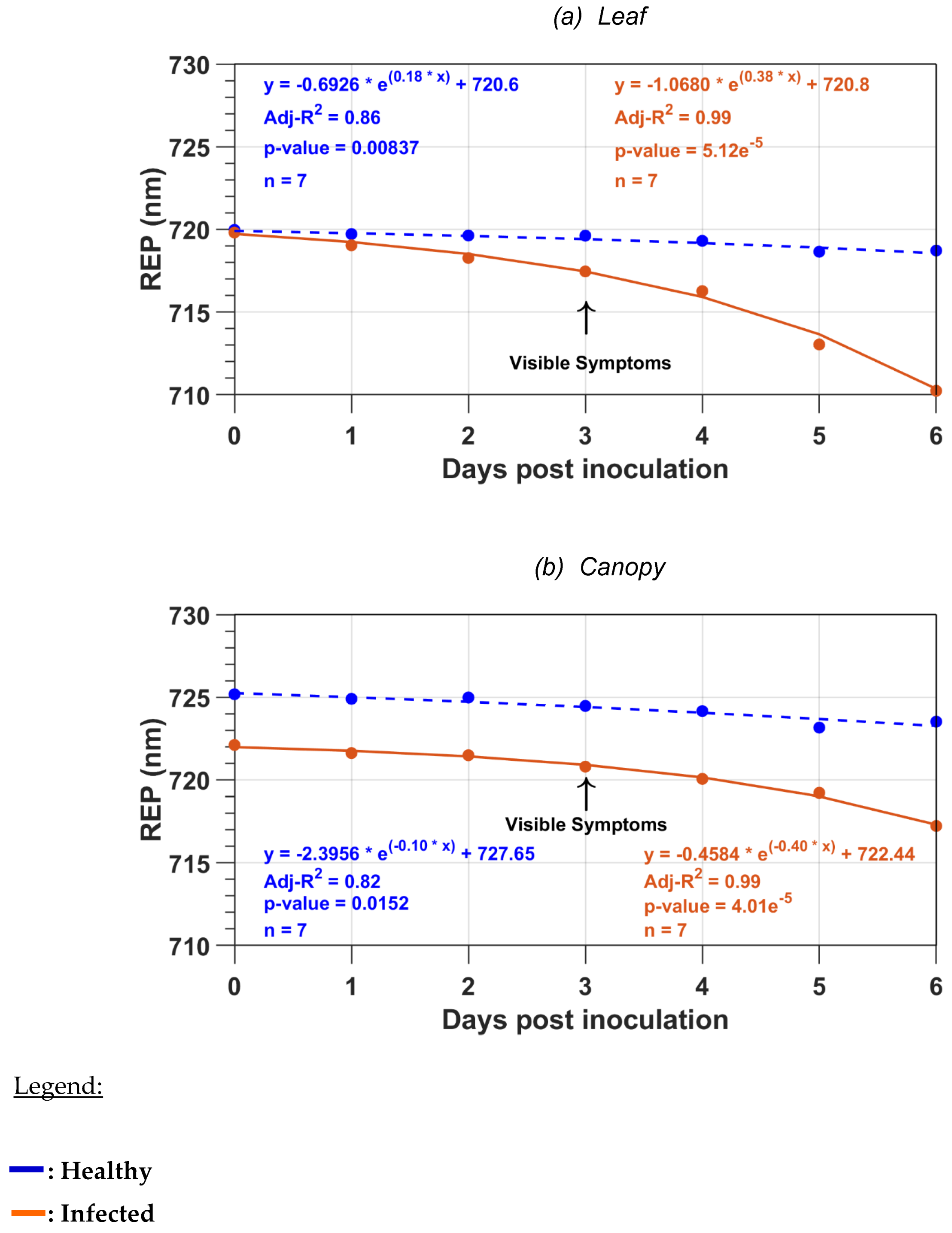
Regression models were used to model the RWP or REP wavelength variations as a function of the DPI. The applied regression methods assume that the data have a normal distribution, which was visually assessed using histogram plots such as for the t-test mean comparison. For both relationships, we tested both the linear and exponential models. The best-explained variance and the lowest sum of the square errors were obtained with a linear model in the case of RWP and an exponential model in the case of REP. The regression model was estimated on a rather small number of observations (n = 7), but each observation was a mean value of more than 100 leaf measurements or 32 canopy measurements for each DPI. Also, the regression parameters were significant, which is an indication that there was no type II error in the models due to the low sample size.
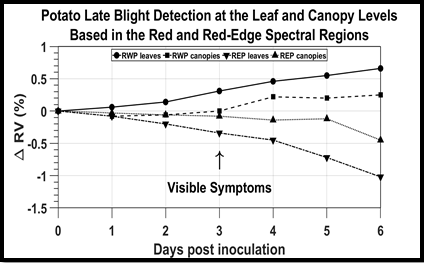
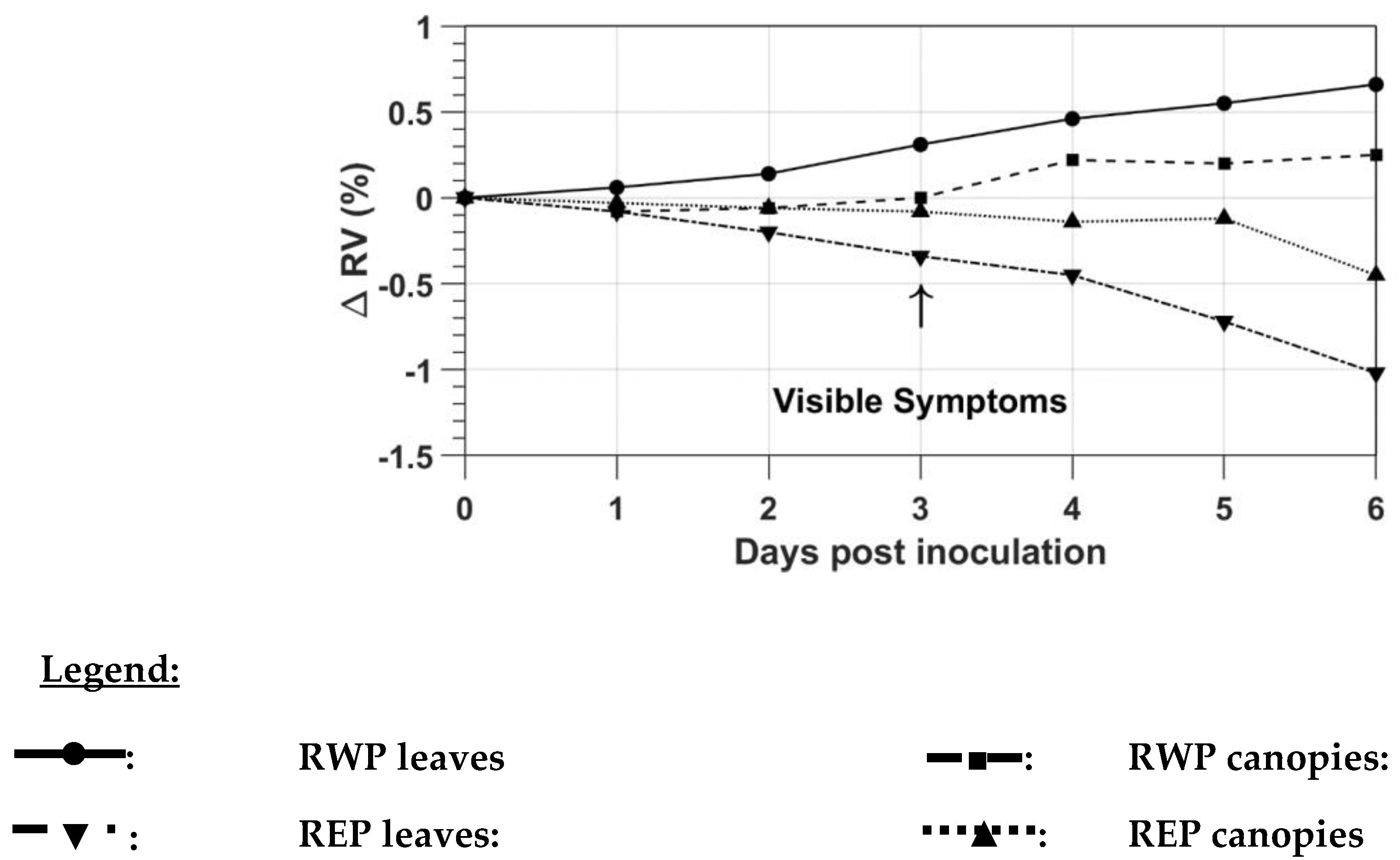
The variations of RWP and REP were also described by the ratio variations (ΔRV), which were computed with the mean daily RWP and REP wavelengths, following Fernández et al. [
21], as follows:
where
ΔRV = ratio variation of RWP or REP with respect to the healthy case reported to the day of inoculation (in %);
Vi = RWP or REP wavelength of the infected leaves or plants (nm);
Vh = RWP or REP wavelength of the healthy leaves or plants (nm);
Vi,o = RWP or REP wavelength of the infected leaves or plants on the day of inoculation (nm);
Vh,o = RWP or REP wavelength of the healthy leaves or plants on the day of inoculation (nm).
Finally, to test whether the RWP, REP, or spectra in the red and red-edge regions sorted healthy and infected leaves or plants, an SVM algorithm embedded in the MATLAB Statistics and Machine Learning Toolbox™ was applied to RWP and REP wavelengths, and the reflectances at 668, 705, 717 and 740 nm were measured at the leaf and canopy levels. These four wavelengths correspond to the central wavelengths of the red and red-edge bands of the Micasense® Dual-X camera (Micasense, Inc., Seattle, WA, USA) that will be calibrated in field conditions for monitoring the presence of potato late blight from red and red-edge UAV images. The SVM algorithm was calibrated using cross-validation and a 10 K-fold division to avoid overfitting.
4. Discussion
In this study, we analyzed the changes in the raw and first-order derivative reflectance spectra in the red and red-edge regions induced by late blight disease on potato leaves and plants. In our experiment, the pre-symptomatology period has the same length as the one of Gold et al. [
22] but a shorter length than those observed by Schumann [
30]. Our study and Gold et al. [
22] used data acquired in a growth chamber that provides optimal conditions for disease development. By contrast, Schumann [
30] reported field results, where the development of the disease depends on weather conditions and can take more time to be developed. For both the raw and first-order derivative spectra, the disease induces chlorosis that produces a blue shift. At the plant level, the first-order derivative reflectance spectra were more sensitive to the disease development than the raw reflectance spectra, given that the blue shift was observed at 2 DPI before the symptoms were visible with the first-order derivative reflectance spectra but only at 4 DPI after the symptoms were visible with the raw reflectance spectra. The reason for the high sensitivity of first-order derivative spectra is probably because first-order derivatives suppress the spectral effect of other organic components present in leaves, like lignin and secondary pigments [
28], therefore improving the sensitivity to chlorosis. We also observed an increase in the raw reflectance values in the red-edge region that was determined by Carter and Knapp [
31] as being an early indicator of stress.
The raw and first-order derivative reflectance spectra were used to derive two main parameters of the red and red-edge region: the RWP and REP wavelengths. The REP wavelength was shown to be more sensitive to the disease than the RWP. As a result, the modeling of the variation as a function of DPI produced models with higher R
2 values with the REP values than with the RWP values. The blue shift we observed with REP is in agreement with several studies [
32,
33,
34] that related this blue shift to chlorosis and plant stress. Our result with the RWP is not in agreement with Liu et al. [
35], who reported a high sensitivity of RWP to chlorophyll content. Results of Liu et al. [
35] were based on simulations, while our results are based on measurements. Another difference with this study is that Liu et al. [
35] used an inverted Gaussian model to estimate RWP while we searched for the minimum reflectance value of a quadratic curve.
Following Dutta et al. [
18] and Fernández et al. [
21], we also computed the percentual variations of the ratio between the infected and healthy cases (ΔRV). Our ratio variations computed with the RWP and REP values at the leaf and canopy levels (−1.02% to 0.66%) are lower than the NDVI-based ratio variations computed by Dutta et al. [
18] (−24% to −60%) with AWiFS and MODIS data over four potato sites infected with late blight, probably because for Dutta et al. [
18] the variation was computed over a longer period (24 days between the first and last evaluations) compared to our period (6 days between the first and last evaluations). The variations observed for both the RWP and REP wavelengths were also lower than those computed by Fernández et al. [
21] with the simple ratio [
36] and the red-edge chlorophyll index [
37] on the same dataset as the one used in this study. In this study, the variations are computed based on single wavelength positions (RWP and REP), while those of Fernández et al. [
21] use vegetation index values that are computed using reflectance in multiple bands.
We applied an SVM classifier to RWP or REP wavelengths as well as to reflectance values at the four red and red-edge wavelengths of the Micasense camera to sort healthy and infected leaves or plants. SVM classifiers were already determined to be highly performant even in the case of limited samples [
38]. The classifiers applied to RWP or REP were less performant than the ones applied to the reflectances of the four Micasense bands. At the canopy level, both the overall classification accuracy (93%) and the sensitivity (0.94) were high with both the healthy and infected plants at 0 DPI when using REP (
Table 6). As shown in
Table 2, there was a significant (at
p < 0.05) difference of the REP values between the healthy and infected groups. Such difference may be due to a difference in leaf area index or leaf angle distribution between both groups of plants. This difference was not quantified in the analysis as we did not measure the leaf area indices nor the leaf angle distributions. Differences in leaf area indices or leaf angle distributions have already been shown to influence REP [
32,
39]. Using the reflectances at the four red and red-edge wavelengths of the Micasense camera, at the leaf level (
Table 7), the overall classification accuracy and the sensitivities for the infected leaves increased when the first symptoms were visible at 3 DPI. The quadratic SVM could sort healthy and infected leaves, particularly when the symptoms became apparent. The highest classification accuracy (93.33%) and the highest sensitivity (0.90) for the infected case were obtained at 6 DPI once the disease was well established. This accuracy was on the same order of magnitude as the one (93.0%) obtained at 4 DPI by Römer et al. [
40] who applied an SVM classifier to ultraviolet-induced fluorescence data acquired between 370 to 800 nm for classifying healthy leaves and leaves infected with
Puccinia triticia in the case of winter wheat (
Triticum aestivum). It was higher than the one obtained with a Partial Least Square Discriminant Analysis (PLS-DA) method for sorting leaves infected with potato late blight (89.77% [
21] and 65–72.73% [
41]). At the canopy level, the highest classification accuracy was obtained at 5 DPI (89.06%,). It was higher than the one (85.93%) obtained by Fernández et al. [
21] who applied PLS-DA to the 400–900 reflectance spectra to classify healthy and infected plants.
Finally, a factor that needs to be considered as having a potential influence on the classification results at the canopy level is related to the measured area on each plant. As explained in the Materials and Methods, the measured area on each healthy or infected plant was 82.83 cm
2, and the total plant area was 490.87 cm
2. Hence, only 16.87% of the entire canopy area was measured. This measured area is lower than the one of Zhang et al. [
16], who acquired spectra on a measured area of 5658 cm
2 on tomato plants infected with late blight and of Bienkowski et al. [
42] who collected spectral data on a measured area of 1520 cm
2. However, both studies did not report the percentage of the plant area; the measurements were done in field conditions. Our spectral data were collected on plants grown in pots inside a walk-in chamber. Therefore, the influence of factors such as illumination and canopy plant geometry might be lower in our case than in the case of the aforementioned studies.
5. Conclusions
In this study, we analyzed the changes in the raw and first-order derivative reflectance spectra in the red and red-edge regions induced by late blight disease on potato leaves and plants. At the leaf level, with the disease development, both types of spectra had a blue spectral shift due to the chlorosis induced by the disease. The blue shift appeared earlier in the first-order derivative reflectance spectra than in the raw reflectance spectra. We also observed an increase in the raw leaf reflectance values in the red-edge region that can be an early indicator of stress. At the canopy level, the spectra were less sensitive to disease development than the leaf spectra. Such as for the leaf level, the first-order derivative spectra were also better than the raw reflectance spectra to detect the disease. Both types of spectra were used to compute two main parameters of the red and red-edge region: the RWP and REP wavelengths. The REP wavelength was shown to be more sensitive to the disease than the RWP for both the leaf and canopy measurements. The models of the variations as a function of DPI had higher R
2 values with REP than with RWP. Following Dutta et al. [
18] and Fernández et al. [
21], we also computed the percent variations of the ratio between the infected and healthy cases (ΔRV). The variations observed for both the RWP and REP wavelengths were lower than those of [
18] and [
21]. To sort healthy and infected leaves or plants, we applied an SVM classifier to RWP or REP wavelengths as well as to reflectance at four selected red and red-edge bands of the Micasense
® Dual-X camera. The SVM classifier applied to these reflectances gave higher overall accuracies that the SVM classifier applied to the REP and RWP values. The SVM classifier applied to reflectances of four Micasense
® Dual-X camera bands in the red and red-edge regions was able to sort healthy and infected cases with both leaf and canopy measurements, reaching an overall classification accuracy of 89.33% at 3 DPI when symptoms were visible for the first time with the leaf measurements and of 89.06% at 5 DPI, i.e., two days after the symptoms became apparent, with the canopy measurements.
Our results were obtained on Shepody cultivar potato plants. Further work is needed to test the method over other potato cultivars. This is especially important for the canopy-level results because the canopy-level measurements are influenced by the canopy geometry, which highly depends on the cultivar. Future research under controlled environments should investigate whether the size of the canopy areas considered for the spectral acquisition has an influence on the capacity of red and red-edge regions to detect late blight over potato plants. Additionally, the results were obtained in a walk-in chamber that had a controlled environment. Further work is needed to test the methodology in real field conditions. The study used point measurements, and there is the need to test the methodology on UAV imagery acquired over real field conditions. For field condition testing, it is important to determine the optimal spatial and temporal resolutions of the UAV images to be able to effectively monitor the disease occurrence in the field, as it is critical to get the information about the disease on its onset to have proper disease management strategies. While the results of this study are quite promising, they were acquired on a limited number of plants. Further work is needed to test the method of broad sampling. In this study, we only tested two metrics, REP and RWP, but other spectral metrics that can detect crop diseases can be considered, such as those already tested by past studies working on various crop diseases (
Table 9).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}