
Spatiotemporal Changes and Driver Analysis of Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
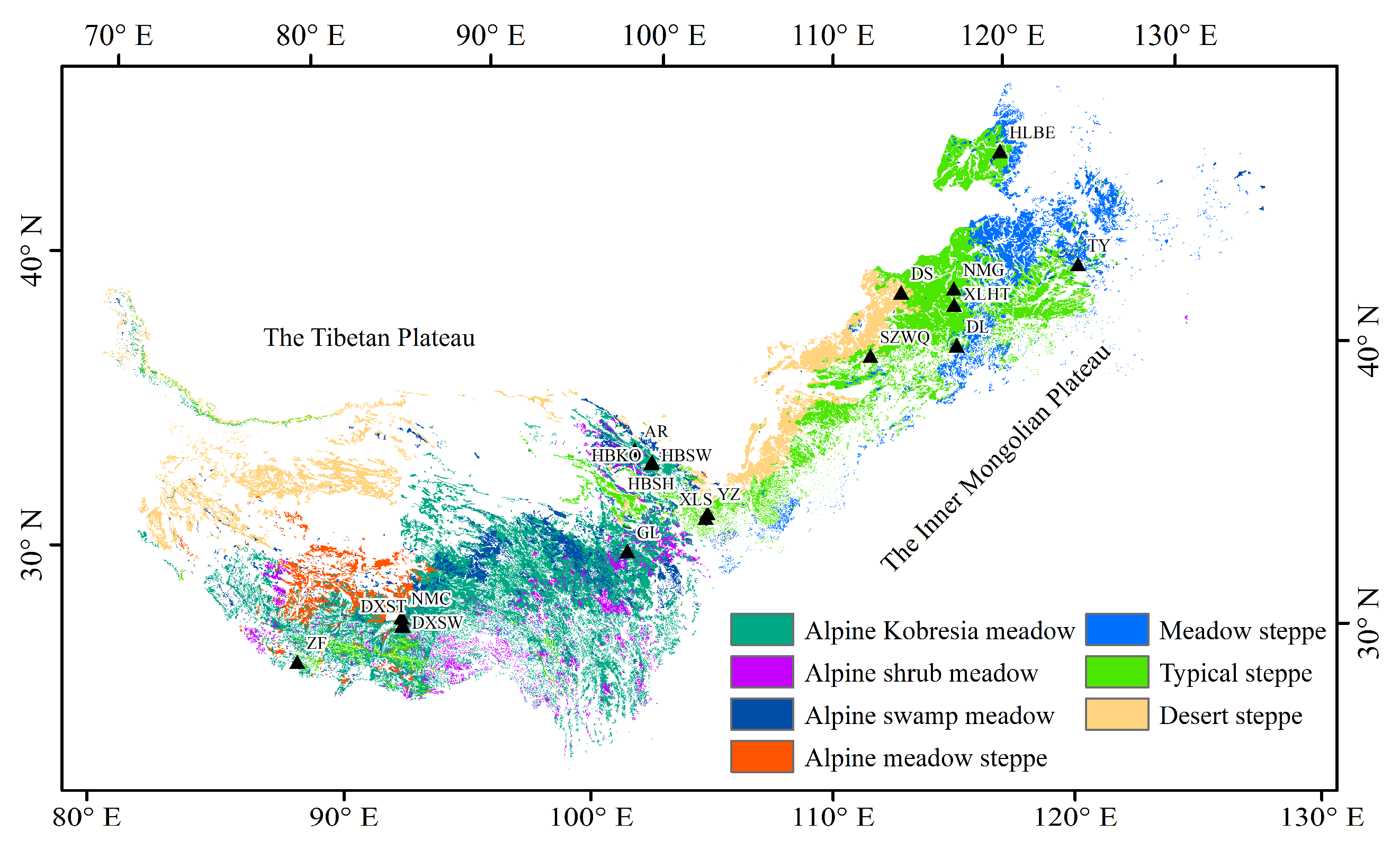
2.1. Study Area
2.2. Data
2.2.1. Flux and Meteorological Data
2.2.2. Remote Sensing Data
2.2.3. Soil Data
2.3. Model
2.3.1. Deep Learning Model: Geoman
2.3.2. Model Training and Evaluation
3. Results
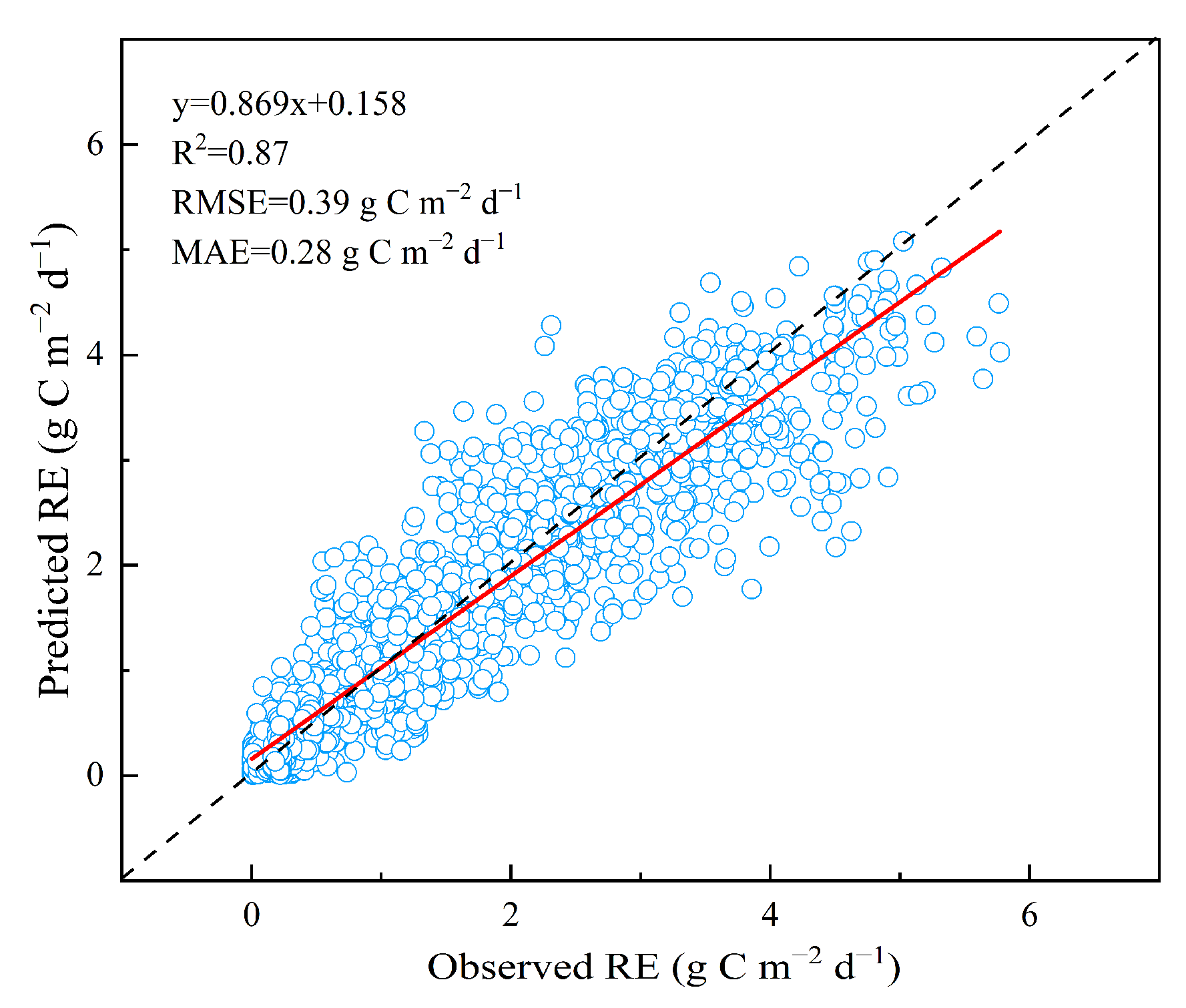
3.1. The Performance of Geoman at the Site Scale
3.1.1. Model Evaluation
3.1.2. Relative Importance of Environmental Factors
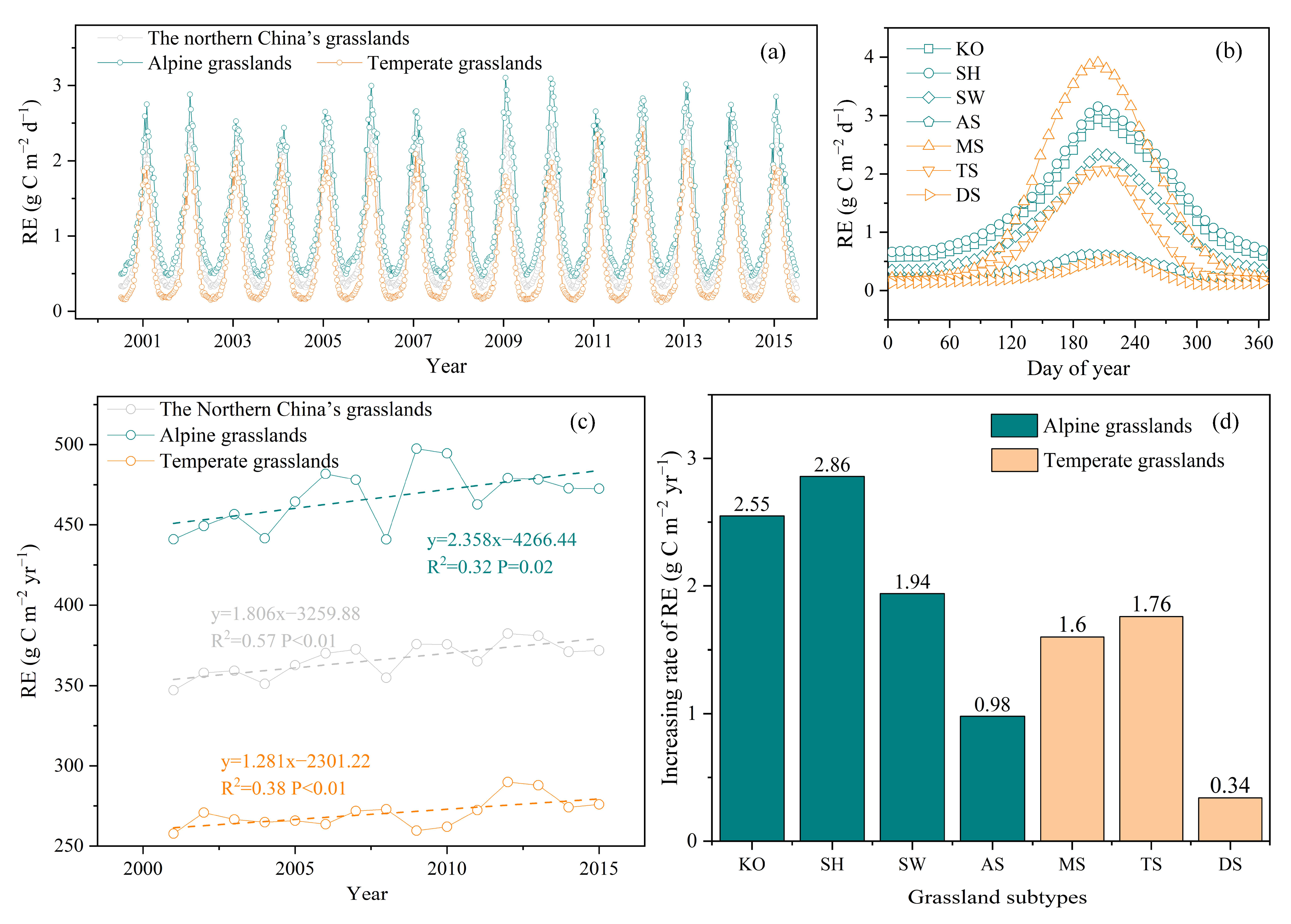
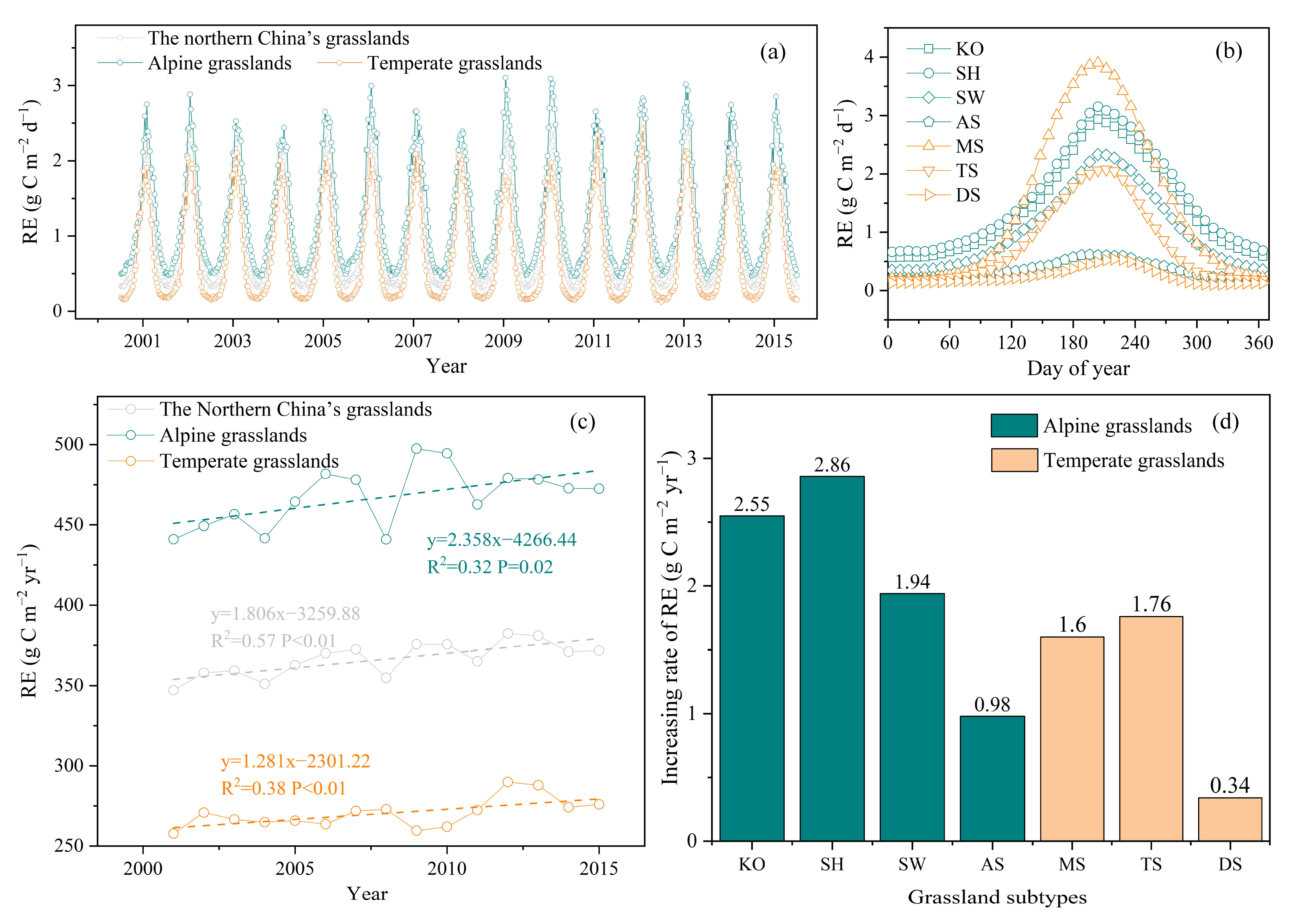
3.2. Spatiotemporal Changes in RE in Northern China’s Grasslands
3.2.1. Spatial Patterns of RE in Northern China’s Grasslands
3.2.2. Temporal Variation for RE in Northern China’s Grasslands
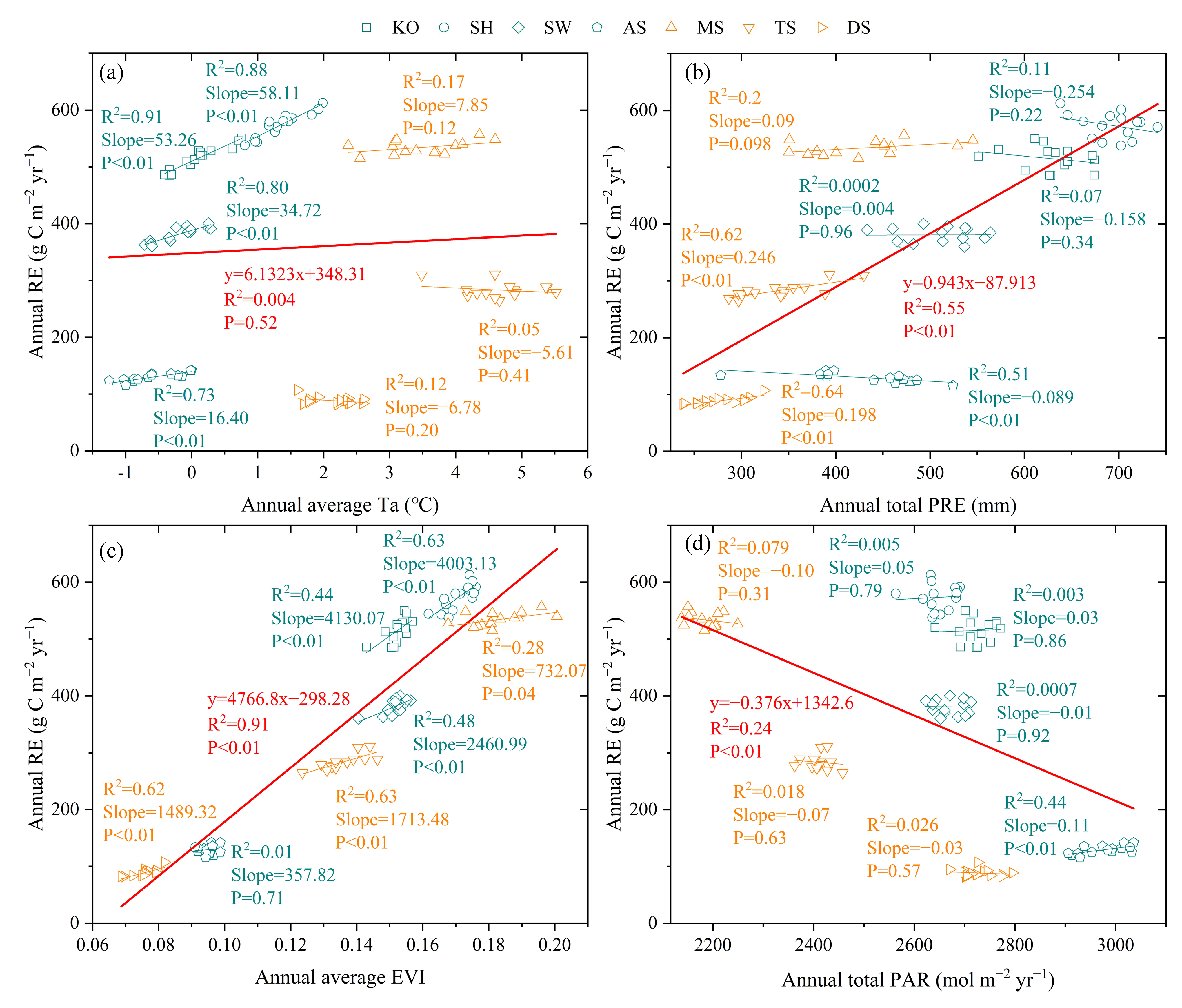
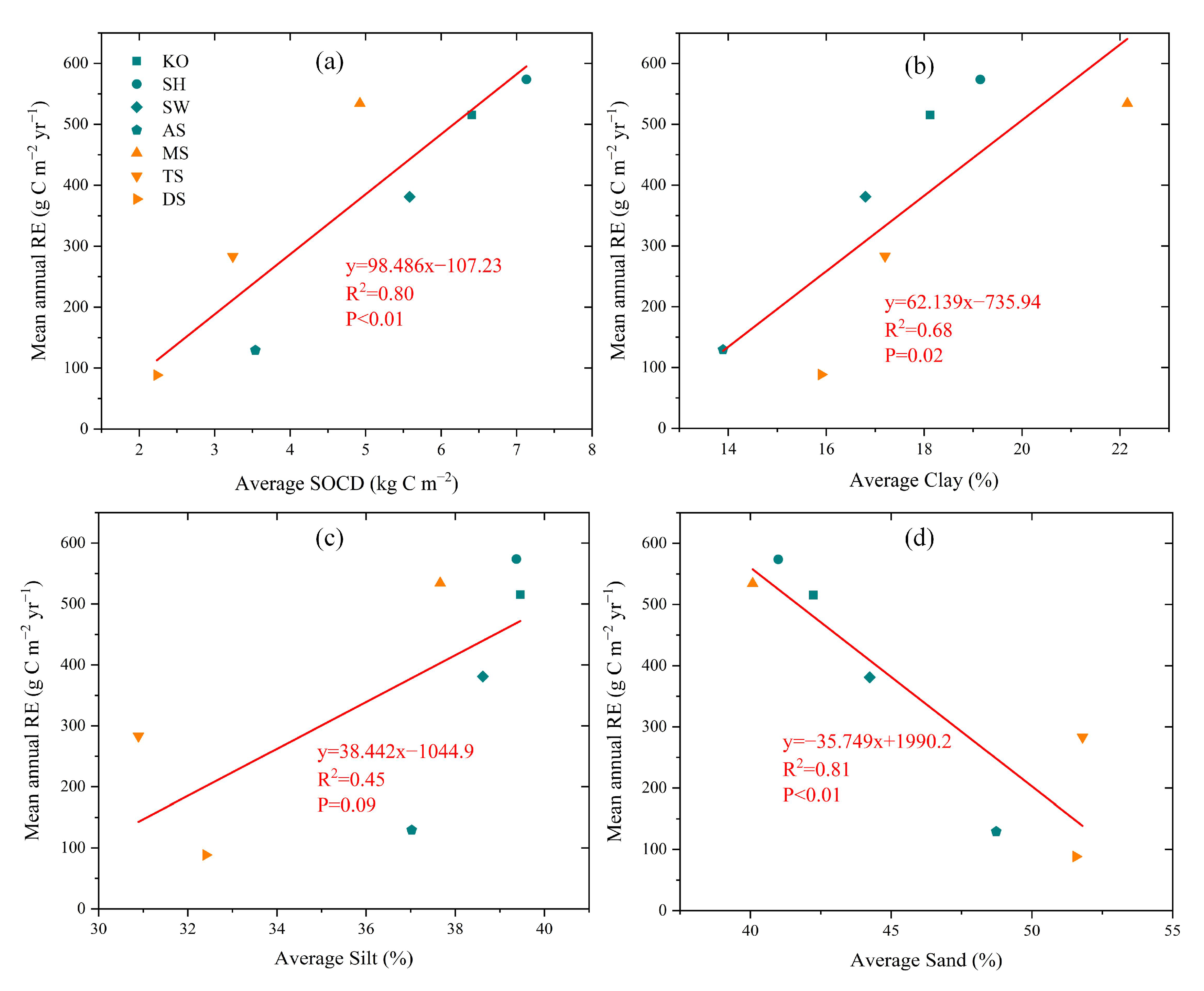
3.3. Environmental Effects on RE Spatiotemporal Variation
3.3.1. Environment Effects on RE Seasonal Dynamics
3.3.2. Environment Effects on RE Interannual Dynamics
3.3.3. Environment Effects on RE Spatial Variation
4. Discussion
4.1. Model Comparison
4.2. Climatic and Biotic Control over RE
4.3. Limitations and Prospects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Migliavacca, M.; Reichstein, M.; Richardson, A.D.; Colombo, R.; Sutton, M.A.; Lasslop, G.; Tomelleri, E.; Wohlfahrt, G.; Carvalhais, N.; Cescatti, A.; et al. Semiempirical modeling of abiotic and biotic factors controlling ecosystem respiration across eddy covariance sites. Glob. Change Biol. 2011, 17, 390–409. [Google Scholar] [CrossRef]
- Tang, X.; Zhou, Y.; Li, H.; Yao, L.; Ding, Z.; Ma, M.; Yu, P. Remotely monitoring ecosystem respiration from various grasslands along a large-scale east–west transect across northern China. Carbon Balance Manag. 2020, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Q. Terrestrial carbon-cycle feedback to climate warming. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 683–712. [Google Scholar] [CrossRef] [Green Version]
- Wohlfahrt, G.; Anderson-Dunn, M.; Bahn, M.; Balzarolo, M.; Berninger, F.; Campbell, C.; Carrara, A.; Cescatti, A.; Christensen, T.; Dore, S.; et al. Biotic, Abiotic, and Management Controls on the Net Ecosystem CO2 Exchange of European Mountain Grassland Ecosystems. Ecosystems 2008, 11, 1338–1351. [Google Scholar] [CrossRef]
- Zheng, C.; Tang, X.; Gu, Q.; Wang, T.; Wei, J.; Song, L.; Ma, M. Climatic anomaly and its impact on vegetation phenology, carbon sequestration and water-use efficiency at a humid temperate forest. J. Hydrol. 2018, 565, 150–159. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Margolis, H.A.; Cescatti, A.; Richardson, A.D.; Altaf Arain, M.; Arneth, A.; Bernhofer, C.; Bonal, D.; Chen, J.; et al. Global patterns of land-atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. J. Geophys. Res. Biogeosci. 2011, 116, G3. [Google Scholar] [CrossRef] [Green Version]
- Ai, J.; Jia, G.; Epstein, H.E.; Wang, H.; Zhang, A.; Hu, Y. MODIS-Based Estimates of Global Terrestrial Ecosystem Respiration. J. Geophys. Res. Biogeosci. 2018, 123, 326–352. [Google Scholar] [CrossRef]
- Li, W.; Ciais, P.; Wang, Y.; Yin, Y.; Peng, S.; Zhu, Z.; Bastos, A.; Yue, C.; Ballantyne, A.P.; Broquet, G.; et al. Recent Changes in Global Photosynthesis and Terrestrial Ecosystem Respiration Constrained From Multiple Observations. Geophys. Res. Lett. 2018, 45, 1058–1068. [Google Scholar] [CrossRef]
- Niu, B.; Zhang, X.; Piao, S.; Janssens, I.A.; Fu, G.; He, Y.; Zhang, Y.; Shi, P.; Dai, E.; Yu, C.; et al. Warming homogenizes apparent temperature sensitivity of ecosystem respiration. Sci. Adv. 2021, 7, eabc7358. [Google Scholar] [CrossRef]
- Huang, N.; Wang, L.; Song, X.-P.; Black, T.A.; Jassal, R.S.; Myneni, R.B.; Wu, C.; Wang, L.; Song, W.; Ji, D.; et al. Spatial and temporal variations in global soil respiration and their relationships with climate and land cover. Sci. Adv. 2020, 6, eabb8508. [Google Scholar] [CrossRef]
- Bond-Lamberty, B. New Techniques and Data for Understanding the Global Soil Respiration Flux. Earth’s Future 2018, 6, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Ge, R.; He, H.; Ren, X.; Zhang, L.; Li, P.; Zeng, N.; Yu, G.; Zhang, L.; Yu, S.-Y.; Zhang, F.; et al. A Satellite-Based Model for Simulating Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands. Remote Sens. 2018, 10, 149. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Norby, R.J.; Ledford, J.; Weltzin, J.F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Glob. Change Biol. 2007, 13, 2411–2424. [Google Scholar] [CrossRef]
- Reichstein, M.; Rey, A.; Freibauer, A.; Tenhunen, J.; Valentini, R.; Banza, J.; Casals, P.; Cheng, Y.; Grünzweig, J.M.; Irvine, J.; et al. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Glob. Biogeochem. Cycles 2003, 17, 4. [Google Scholar] [CrossRef]
- McCulley, R.L.; Burke, I.C.; Nelson, J.A.; Lauenroth, W.K.; Knapp, A.K.; Kelly, E.F. Regional Patterns in Carbon Cycling Across the Great Plains of North America. Ecosystems 2005, 8, 106–121. [Google Scholar] [CrossRef]
- Jägermeyr, J.; Gerten, D.; Lucht, W.; Hostert, P.; Migliavacca, M.; Nemani, R. A high-resolution approach to estimating ecosystem respiration at continental scales using operational satellite data. Glob. Change Biol. 2014, 20, 1191–1210. [Google Scholar] [CrossRef]
- Luo, Y.; Ahlström, A.; Allison, S.D.; Batjes, N.H.; Brovkin, V.; Carvalhais, N.; Chappell, A.; Ciais, P.; Davidson, E.; Finzi, A.; et al. Toward more realistic projections of soil carbon dynamics by Earth system models. Glob. Biogeochem. Cycles 2016, 30, 40–56. [Google Scholar] [CrossRef]
- Yuan, W.; Luo, Y.; Li, X.; Liu, S.; Yu, G.; Zhou, T.; Bahn, M.; Black, A.; Desai, A.; Cescatti, A.; et al. Redefinition and global estimation of basal ecosystem respiration rate. Glob. Biogeochem. Cycles 2011, 25, 4. [Google Scholar] [CrossRef]
- Huang, N.; Niu, Z.; Zhan, Y.; Xu, S.; Tappert, M.C.; Wu, C.; Huang, W.; Gao, S.; Hou, X.; Cai, D. Relationships between soil respiration and photosynthesis-related spectral vegetation indices in two cropland ecosystems. Agric. For. Meteorol. 2012, 160, 80–89. [Google Scholar] [CrossRef]
- Lei, J.S.; Guo, X.; Zeng, Y.F.; Zhou, J.Z.; Gao, Q.; Yang, Y.F. Temporal changes in global soil respiration since 1987. Nat. Commun. 2021, 12, 403. [Google Scholar] [CrossRef]
- Dilustro, J.J.; Collins, B.; Duncan, L.; Crawford, C. Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests. For. Ecol. Manag. 2005, 204, 87–97. [Google Scholar] [CrossRef]
- Lohila, A.; Aurela, M.; Regina, K.; Laurila, T. Soil and total ecosystem respiration in agricultural fields: Effect of soil and crop type. Plant Soil 2003, 251, 303–317. [Google Scholar] [CrossRef]
- Franzluebbers, K.; Franzluebbers, A.J.; Jawson, M.D. Environmental controls on soil and whole-ecosystem respiration from a tallgrass prairie. Soil Sci. Soc. Am. J. 2002, 66, 254–262. [Google Scholar]
- Janssens, I.A.; Lankreijer, H.; Matteucci, G.; Kowalski, A.S.; Buchmann, N.; Epron, D.; Pilegaard, K.; Kutsch, W.; Longdoz, B.; Grünwald, T.; et al. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Glob. Change Biol. 2002, 7, 269–278. [Google Scholar] [CrossRef]
- Kätterer, T.; Reichstein, M.; Andrén, O.; Lomander, A. Temperature dependence of organic matter decomposition: A critical review using literature data analyzed with different models. Biol. Fertil. Soils 1998, 27, 258–262. [Google Scholar] [CrossRef]
- Hashimoto, S.; Carvalhais, N.; Ito, A.; Migliavacca, M.; Nishina, K.; Reichstein, M. Global spatiotemporal distribution of soil respiration modeled using a global database. Biogeosciences 2015, 12, 4121–4132. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Wang, Q.; Han, X.; Wan, S.; Li, L. Temporal and spatial variability and controls of soil respiration in a temperate steppe in northern China. Glob. Biogeochem. Cycles 2010, 24, 2. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Change Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- LaFleur, P.M.; Roulet, N.T.; Bubier, J.L.; Frolking, S.; Moore, T.R. Interannual variability in the peatland-atmosphere carbon dioxide exchange at an ombrotrophic bog. Glob. Biogeochem. Cycles 2003, 17, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Zhang, K.; Bagheri, M.; Burken, J.G.; Gu, A.; Li, B.; Ma, X.; Marrone, B.L.; Ren, Z.J.; Schrier, J.; et al. Machine Learning: New Ideas and Tools in Environmental Science and Engineering. Environ. Sci. Technol. 2021, 55, 12741–12754. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Bondeau, A. Towards global empirical upscaling of FLUXNET eddy covariance observations: Validation of a model tree ensemble approach using a biosphere model. Biogeosciences 2009, 6, 2001–2013. [Google Scholar] [CrossRef] [Green Version]
- Dou, X.; Yang, Y. Evapotranspiration estimation using four different machine learning approaches in different terrestrial ecosystems. Comput. Electron. Agric. 2018, 148, 95–106. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, G.; Hellwich, O.; Chen, C.; Zhang, W.; Xie, M.; He, H.; Shi, H.; Wang, Y. A framework for estimating actual evapotranspiration at weather stations without flux observations by combining data from MODIS and flux towers through a machine learning approach. J. Hydrol. 2021, 603, 127047. [Google Scholar] [CrossRef]
- Zhu, X.; He, H.; Ma, M.; Ren, X.; Zhang, L.; Zhang, F.; Li, Y.; Shi, P.; Chen, S.; Wang, Y.; et al. Estimating Ecosystem Respiration in the Grasslands of Northern China Using Machine Learning: Model Evaluation and Comparison. Sustainability 2020, 12, 2099. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Dafflon, B.; Tran, A.P.; Falco, N.; Hubbard, S.S. A deep learning hybrid predictive modeling (HPM) approach for estimating evapotranspiration and ecosystem respiration. Hydrol. Earth Syst. Sci. 2021, 25, 6041–6066. [Google Scholar] [CrossRef]
- Tramontana, G.; Jung, M.; Schwalm, C.R.; Ichii, K.; Camps-Valls, G.; Ráduly, B.; Reichstein, M.; Arain, M.A.; Cescatti, A.; Kiely, G.; et al. Predicting carbon dioxide and energy fluxes across global FLUXNET sites with regression algorithms. Biogeosciences 2016, 13, 4291–4313. [Google Scholar] [CrossRef] [Green Version]
- Kaba, K.; Sarıgül, M.; Avcı, M.; Kandırmaz, H.M. Estimation of daily global solar radiation using deep learning model. Energy 2018, 162, 126–135. [Google Scholar] [CrossRef]
- Roy, B. Optimum machine learning algorithm selection for forecasting vegetation indices: MODIS NDVI & EVI. Remote Sens. Appl. Soc. Environ. 2021, 23, 100582. [Google Scholar] [CrossRef]
- Lou, J.; Xu, G.; Wang, Z.; Yang, Z.; Ni, S. Multi-Year NDVI Values as Indicator of the Relationship between Spatiotemporal Vegetation Dynamics and Environmental Factors in the Qaidam Basin, China. Remote Sens. 2021, 13, 1240. [Google Scholar] [CrossRef]
- Kerkech, M.; Hafiane, A.; Canals, R. Deep leaning approach with colorimetric spaces and vegetation indices for vine diseases detection in UAV images. Comput. Electron. Agric. 2018, 155, 237–243. [Google Scholar] [CrossRef]
- Arrieta, A.B.; Díaz-Rodríguez, N.; Del Ser, J.; Bennetot, A.; Tabik, S.; Barbado, A.; Garcia, S.; Gil-Lopez, S.; Molina, D.; Benjamins, R.; et al. Explainable Artificial Intelligence (XAI): Concepts, taxonomies, opportunities and challenges toward responsible AI. Inf. Fusion 2020, 58, 82–115. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, S.; Mithal, V.; Polatkan, G.; Ramanath, R. An Attentive Survey of Attention Models. ACM Trans. Intell. Syst. Technol. 2021, 12, 1–32. [Google Scholar] [CrossRef]
- Reichstein, M.; Camps-Valls, G.; Stevens, B.; Jung, M.; Denzler, J.; Carvalhais, N.; Prabhat. Deep learning and process understanding for data-driven Earth system science. Nature 2019, 566, 195–204. [Google Scholar] [CrossRef]
- Liang, Y.; Ke, S.; Zhang, J.; Yi, X.; Zheng, Y. Geoman: Multi-level attention networks for geo-sensory time series prediction. In Proceedings of the Twenty-Seventh International Joint Conference on Artificial Intelligence, Stockholm, Sweden, 13–19 July 2018. [Google Scholar]
- Vaswani, A.; Shazeer, N.; Parmar, N.; Uszkoreit, J.; Jones, L.; Gomez, A.N.; Kaiser, L.; Polosukhin, I. Attention is all you need. In Proceedings of the 31st International Conference on Neural Information Processing Systems, Long Beach, CA, USA, 4–9 December 2017. [Google Scholar]
- Zheng, C.A.P.; Fan, X.L.; Wang, C.; Qi, J.Z. Gman: A graph multi-attention network for traffic prediction. In Proceedings of the Thirty-Second Innovative Applications of Artificial Intelligence Conference, New York, NY, USA, 9–11 February 2020. [Google Scholar]
- Li, Y.; Tong, Z.; Tong, S.; Westerdahl, D. A data-driven interval forecasting model for building energy prediction using attention-based LSTM and fuzzy information granulation. Sustain. Cities Soc. 2022, 76, 103481. [Google Scholar] [CrossRef]
- Xue, K.; Yuan, M.M.; Shi, Z.J.; Qin, Y.; Deng, Y.; Cheng, L.; Wu, L.; He, Z.; Van Nostrand, J.D.; Bracho, R.; et al. Tundra soil carbon is vulnerable to rapid microbial decomposition under climate warming. Nat. Clim. Change 2016, 6, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Ni, J. Carbon storage in grasslands of China. J. Arid Environ. 2002, 50, 205–218. [Google Scholar] [CrossRef]
- Su, D. The Atlas of Grassland Resources of China (1:1,000,000); Press of Map: Beijing, China, 1993. (In Chinese) [Google Scholar]
- Yu, G.-R.; Wen, X.-F.; Sun, X.-M.; Tanner, B.D.; Lee, X.; Chen, J.-Y. Overview of ChinaFLUX and evaluation of its eddy covariance measurement. Agric. For. Meteorol. 2006, 137, 125–137. [Google Scholar] [CrossRef]
- Wang, H.; Jia, G.; Fu, C.; Feng, J.; Zhao, T.; Ma, Z. Deriving maximal light use efficiency from coordinated flux measurements and satellite data for regional gross primary production modeling. Remote Sens. Environ. 2010, 114, 2248–2258. [Google Scholar] [CrossRef]
- Li, X.; Cheng, G.D.; Liu, S.M.; Xiao, Q.; Ma, M.G.; Jin, R.; Che, T.; Liu, Q.H.; Wang, W.Z.; Qi, Y.; et al. Heihe watershed allied telemetry experimental research (hiwater): Scientific objectives and experimental design. Bull. Am. Meteorol. Soc. 2013, 94, 1145–1160. [Google Scholar] [CrossRef]
- Schwalm, C.R.; Williams, C.A.; Schaefer, K.; Anderson, R.; Arain, M.A.; Baker, I.; Barr, A.; Black, T.A.; Chen, G.; Chen, J.M.; et al. A model-data intercomparison of CO2 exchange across North America: Results from the North American Carbon Program site synthesis. J. Geophys. Res. Biogeosci. 2010, 115, G3. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; He, H.; Zhang, L.; Yu, G. Global radiation, photosynthetically active radiation, and the diffuse component dataset of China, 1981–2010. Earth Syst. Sci. Data 2018, 10, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.B.; Wang, J.W.; Ye, H.; Liu, Y.; He, H.L. An interpolated temperature and precipitation dataset at 1-km grid resolution in China (2000–2012). China Sci. Data 2017, 2, 73–80. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Carvalhais, N.; Forkel, M.; Khomik, M.; Bellarby, J.; Jung, M.; Migliavacca, M.; Μu, M.; Saatchi, S.; Santoro, M.; Thurner, M.; et al. Global covariation of carbon turnover times with climate in terrestrial ecosystems. Nature 2014, 514, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Kraft, B.; Jung, M.; Körner, M.; Mesa, C.R.; Cortés, J.; Reichstein, M. Identifying Dynamic Memory Effects on Vegetation State Using Recurrent Neural Networks. Front. Big Data 2019, 2, 31. [Google Scholar] [CrossRef] [Green Version]
- Bergstra, J.; Bengio, Y. Random search for hyper-parameter optimization. J. Mach. Learn. Res. 2012, 13, 281–305. [Google Scholar]
- Forkel, M.; Carvalhais, N.; Rödenbeck, C.; Keeling, R.; Heimann, M.; Thonicke, K.; Zaehle, S.; Reichstein, M. Enhanced seasonal CO2 exchange caused by amplified plant productivity in northern ecosystems. Science 2016, 351, 696–699. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.Q.; Randerson, J.T.; Abramowitz, G.; Bacour, C.; Blyth, E.; Carvalhais, N.; Ciais, P.; Dalmonech, D.; Fisher, J.B.; Fisher, R.; et al. A framework for benchmarking land models. Biogeosciences 2012, 9, 3857–3874. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Sitch, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Wang, X.; Ahlström, A.; Anav, A.; Canadell, J.G.; Cong, N.; et al. Evaluation of terrestrial carbon cycle models for their response to climate variability and to CO2 trends. Glob. Change Biol. 2013, 19, 2117–2132. [Google Scholar] [CrossRef] [Green Version]
- Koppa, A.; Rains, D.; Hulsman, P.; Poyatos, R.; Miralles, D.G. A deep learning-based hybrid model of global terrestrial evaporation. Nat. Commun. 2022, 13, 1912. [Google Scholar] [CrossRef] [PubMed]
- Raich, J.W.; Potter, C.S.; Bhagawati, D. Interannual variability in global soil respiration, 1980–1994. Glob. Change Biol. 2002, 8, 800–812. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A.; Luo, Y.Q. On the variability of respiration in terrestrial ecosystems: Moving beyond Q10. Glob. Change Biol. 2006, 12, 154–164. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Change Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Luo, Y.; Xia, J.; Shi, Z.; Jiang, L.; Niu, S.; Zhou, X.; Cao, J. Differential responses of ecosystem respiration components to experimental warming in a meadow grassland on the Tibetan Plateau. Agric. For. Meteorol. 2016, 220, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.-S.; Reynolds, J.F.; Maestre, F.T.; Li, F.-M. Hydrological and ecological responses of ecosystems to extreme precipitation regimes: A test of empirical-based hypotheses with an ecosystem model. Perspect. Plant Ecol. Evol. Syst. 2016, 22, 36–46. [Google Scholar] [CrossRef]
- Sun, G.; Wang, Z.; Zhu-Barker, X.; Zhang, N.; Wu, N.; Liu, L.; Lei, Y. Biotic and abiotic controls in determining exceedingly variable responses of ecosystem functions to extreme seasonal precipitation in a mesophytic alpine grassland. Agric. For. Meteorol. 2016, 228–229, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Curtin, D.; Beare, M.H.; Hernandez-Ramirez, G. Temperature and Moisture Effects on Microbial Biomass and Soil Organic Matter Mineralization. Soil Sci. Soc. Am. J. 2012, 76, 2055–2067. [Google Scholar] [CrossRef]
- Pieper, S.J.; Loewen, V.; Gill, M.; Johnstone, J.F. Plant Responses to Natural and Experimental Variations in Temperature in Alpine Tundra, Southern Yukon, Canada. Arctic Antarct. Alp. Res. 2011, 43, 442–456. [Google Scholar] [CrossRef]
- Ji, S.N.; Classen, A.; Zhang, Z.; He, J. Asymmetric winter warming advanced plant phenology to a greater extent than symmetric warming in an alpine meadow. Funct. Ecol. 2017, 31, 2147–2156. [Google Scholar] [CrossRef] [Green Version]
- Bai, W.; Wan, S.; Niu, S.; Liu, W.; Chen, Q.; Wang, Q.; Zhang, W.; Han, X.; Li, L. Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling. Glob. Change Biol. 2010, 16, 1306–1316. [Google Scholar] [CrossRef]
- Sjöström, M.; Ardö, J.; Arneth, A.; Boulain, N.; Cappelaere, B.; Eklundh, L.; de Grandcourt, A.; Kutsch, W.; Merbold, L.; Nouvellon, Y.; et al. Exploring the potential of MODIS EVI for modeling gross primary production across African ecosystems. Remote Sens. Environ. 2011, 115, 1081–1089. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, Q.; Braswell, B.; Urbanski, S.; Boles, S.; Wofsy, S.; Moore, B.; Ojima, D. Modeling gross primary production of temperate deciduous broadleaf forest using satellite images and climate data. Remote Sens. Environ. 2004, 91, 256–270. [Google Scholar] [CrossRef]
- He, H.L.; Wang, S.Q.; Zhang, L.; Wang, J.B.; Ren, X.L.; Zhou, L.; Piao, S.L.; Yan, H.; Ju, W.M.; Gu, F.X.; et al. Altered trends in carbon uptake in china’s terrestrial ecosystems under the enhanced summer monsoon and warming hiatus. Natl. Sci. Rev. 2019, 6, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Papale, D.; Valentini, R.; Aubinet, M.; Bernhofer, C.; Knohl, A.; Laurila, T.; Lindroth, A.; Moors, E.; Pilegaard, K.; et al. Determinants of terrestrial ecosystem carbon balance inferred from European eddy covariance flux sites. Geophys. Res. Lett. 2007, 34, 1. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Wang, Y.; Yang, K.; Wang, S.; Zeng, H.; Baumann, F.; Kuehn, P.; Scholten, T.; He, J.-S. Soil Respiration in Tibetan Alpine Grasslands: Belowground Biomass and Soil Moisture, but Not Soil Temperature, Best Explain the Large-Scale Patterns. PLoS ONE 2012, 7, e34968. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Wallenstein, M.D.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Gómez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef]
- Dacal, M.; Delgado-Baquerizo, M.; Barquero, J.; Berhe, A.A.; Gallardo, A.; Maestre, F.T.; García-Palacios, P. Temperature increases soil respiration across ecosystem types and soil development, but soil properties determine the magnitude of this effect. Ecosystems 2022, 25, 184–198. [Google Scholar] [CrossRef]
- Knohl, A.; Soe, A.R.B.; Kutsch, W.L.; Gockede, M.; Buchmann, N. Representative estimates of soil and ecosystem respiration in an old beech forest. Plant Soil. 2008, 302, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yu, G.; Li, S.; Yan, H.; Zhu, X.; Wang, Q.; Shi, P.; Zhao, L.; Li, Y.; Zhang, F.; et al. A remote sensing model to estimate ecosystem respiration in Northern China and the Tibetan Plateau. Ecol. Model. 2015, 304, 34–43. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Tang, Y.; Ji, C.; Zheng, C.; He, J.; Zhu, B. Storage, patterns and controls of soil organic carbon in the Tibetan grasslands. Glob. Change Biol. 2008, 14, 1592–1599. [Google Scholar] [CrossRef]
- Chu, H.; Baldocchi, D.D.; John, R.; Wolf, S.; Reichstein, M. Fluxes all of the time? A primer on the temporal representativeness of FLUXNET. J. Geophys. Res. Biogeosci. 2017, 122, 289–307. [Google Scholar] [CrossRef]
- Kumar, J.; Hoffman, F.M.; Hargrove, W.W.; Collier, N. Understanding the representativeness of fluxnet for upscaling carbon flux from eddy covariance measurements. Earth Syst. Sci. Data Discuss. 2016, 36, 1–25. [Google Scholar]
- Pastorello, G.; Trotta, C.; Canfora, E.; Chu, H.S.; Christianson, D.; Cheah, Y.W.; Poindexter, C.; Chen, J.Q.; Elbashandy, A.; Humphrey, M.; et al. The fluxnet2015 dataset and the oneflux processing pipeline for eddy covariance data. Sci. Data. 2020, 7, 225. [Google Scholar] [CrossRef]
- Hargrove, W.W.; Hoffman, F.M.; Law, B.E. New analysis reveals representativeness of the ameriflux network. Eos Trans. Am. Geophys. Union 2003, 84, 529–535. [Google Scholar] [CrossRef]
- Sulkava, M.; Luyssaert, S.; Zaehle, S.; Papale, D. Assessing and improving the representativeness of monitoring networks: The European flux tower network example. J. Geophys. Res. Biogeosci. 2011, 116, G3. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Zhang, L.; Gao, Y.; Ren, X.; Zhang, L.; Yu, G.; Wang, S. Regional representativeness assessment and improvement of eddy flux observations in China. Sci. Total Environ. 2015, 502, 688–698. [Google Scholar] [CrossRef]
- Xiao, J.; Zhuang, Q.; Law, B.; Chen, J.; Baldocchi, D.; Cook, D.R.; Oren, R.; Richardson, A.D.; Wharton, S.; Martin, T. A continuous measure of gross primary production for the conterminous United States derived from MODIS and AmeriFlux data. Remote Sens. Environ. 2010, 114, 576–591. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Ollinger, S.V.; Frolking, S.; Hurtt, G.C.; Hollinger, D.Y.; Davis, K.J.; Pan, Y.; Zhang, X.; Deng, F.; Chen, J.; et al. Data-driven diagnostics of terrestrial carbon dynamics over North America. Agric. For. Meteorol. 2014, 197, 142–157. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zone | Grassland Type | Site | Location | Elevation (m) | Operation Period |
|---|---|---|---|---|---|
| TP 1 | KO 3 | AR | 38.04°N, 100.46°E | 3033 | 2014 |
| GL | 34.35°N, 100.56°E | 3980 | 2007, 2010–2011, 2013 | ||
| HBKO | 37.61°N, 101.31°E | 3148 | 2003–2004 | ||
| SH 4 | HBSH | 37.67°N, 101.33°E | 3293 | 2003–2012 | |
| SW 5 | DXSW | 30.47°N, 91.06°E | 4286 | 2009–2010 | |
| HBSW | 37.61°N, 101.33°E | 3160 | 2004–2008, 2010–2012 | ||
| AS 6 | DXST | 30.5°N, 91.06°E | 4333 | 2004–2005, 2007, 2009–2010 | |
| NMC | 30.77°N, 90.96°E | 4730 | 2009 | ||
| ZF | 28.36°N, 86.95°E | 4293 | 2009 | ||
| IM 2 | MS 7 | HLBE | 49.06°N, 119.4°E | 628 | 2012 |
| TY | 44.57°N, 122.92°E | 151 | 2008–2009 | ||
| TS 8 | DL | 42.05°N, 116.28°E | 1324 | 2010–2011 | |
| NMG | 43.53°N, 116.28°E | 1200 | 2004, 2007–2008, 2010–2011 | ||
| XLHT | 44.13°N, 116.32°E | 1187 | 2010–2011 | ||
| YZ | 35.95°N, 104.13°E | 1968 | 2008–2009 | ||
| DS 9 | DS | 44.09°N, 113.57°E | 990 | 2008–2009 | |
| SZWQ | 41.8°N, 111.9°E | 1438 | 2012 | ||
| XLS | 35.77°N, 104.05°E | 2481 | 2008 |
| Grassland Type | KO | SH | SW | AS | MS | TS | DS | Total |
|---|---|---|---|---|---|---|---|---|
| Grid number | 471,226 | 79,952 | 104,752 | 65,294 | 147,111 | 368,673 | 239,440 | 1,476,448 |
| Percentage | 31.92% | 5.42% | 7.09% | 4.42% | 9.96% | 24.97% | 16.22% | 100% |
| The Growing Season | ||||||||
|---|---|---|---|---|---|---|---|---|
| Grassland Subtypes | SOCD | EVI | PAR | PRE | Ta | SoilTex | VT | |
| TP | KO | 0.080 | 0.095 | 0.110 | 0.111 | 0.156 | 0.188 | 0.261 |
| SH | 0.093 | 0.102 | 0.102 | 0.115 | 0.157 | 0.181 | 0.250 | |
| SW | 0.065 | 0.094 | 0.121 | 0.117 | 0.147 | 0.194 | 0.263 | |
| AS | 0.040 | 0.051 | 0.146 | 0.117 | 0.165 | 0.205 | 0.276 | |
| IM | MS | 0.050 | 0.128 | 0.096 | 0.169 | 0.117 | 0.182 | 0.258 |
| TS | 0.038 | 0.083 | 0.125 | 0.184 | 0.120 | 0.188 | 0.263 | |
| DS | 0.034 | 0.042 | 0.152 | 0.155 | 0.110 | 0.200 | 0.306 | |
| The Non-Growing Season | ||||||||
| Grassland subtypes | SOCD | EVI | PAR | PRE | Ta | SoilTex | VT | |
| TP | KO | 0.122 | 0.075 | 0.110 | 0.097 | 0.084 | 0.216 | 0.296 |
| SH | 0.139 | 0.079 | 0.105 | 0.101 | 0.088 | 0.207 | 0.281 | |
| SW | 0.106 | 0.074 | 0.103 | 0.092 | 0.085 | 0.231 | 0.308 | |
| AS | 0.070 | 0.069 | 0.083 | 0.088 | 0.135 | 0.238 | 0.317 | |
| IM | MS | 0.116 | 0.085 | 0.080 | 0.086 | 0.085 | 0.226 | 0.322 |
| TS | 0.121 | 0.077 | 0.092 | 0.083 | 0.069 | 0.233 | 0.324 | |
| DS | 0.113 | 0.066 | 0.061 | 0.077 | 0.100 | 0.228 | 0.354 | |
| Grassland Subtypes | EVI | PAR | PRE | Ta | |
|---|---|---|---|---|---|
| TP | KO | 0.927 | 0.528 | 0.801 | 0.931 |
| SH | 0.938 | 0.515 | 0.807 | 0.929 | |
| SW | 0.925 | 0.674 | 0.826 | 0.951 | |
| AS | 0.624 | 0.697 | 0.564 | 0.834 | |
| IM | MS | 0.892 | 0.773 | 0.736 | 0.956 |
| TS | 0.853 | 0.822 | 0.771 | 0.928 | |
| DS | 0.748 | 0.689 | 0.672 | 0.727 |
| Mean Annual Value | R2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Grassland Subtypes | EVI | PAR (mol m−2 yr−1) | PRE (mm) | Ta (℃) | EVI | PAR | PRE | Ta | |
| TP | KO | 0.15 | 2723.14 | 628.23 | 0.10 | 0.44 | 0.00 | 0.07 | 0.91 |
| SH | 0.17 | 2649.46 | 692.66 | 1.35 | 0.63 | 0.01 | 0.11 | 0.88 | |
| SW | 0.15 | 2673.11 | 508.06 | −0.21 | 0.48 | 0.00 | 0.00 | 0.80 | |
| AS | 0.10 | 2975.61 | 433.30 | −0.62 | 0.01 | 0.44 | 0.51 | 0.73 | |
| IM | MS | 0.18 | 2182.35 | 431.45 | 3.48 | 0.28 | 0.08 | 0.20 | 0.17 |
| TS | 0.14 | 2412.42 | 340.07 | 4.69 | 0.63 | 0.02 | 0.62 | 0.05 | |
| DS | 0.08 | 2728.92 | 275.18 | 2.13 | 0.62 | 0.03 | 0.64 | 0.12 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; He, H.; Wu, X.; Ren, X.; Zhang, L.; Zhu, X.; Feng, L.; Lv, Y.; Chang, Q.; Xu, Q.; et al. Spatiotemporal Changes and Driver Analysis of Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands. Remote Sens. 2022, 14, 3563. https://doi.org/10.3390/rs14153563
Liu W, He H, Wu X, Ren X, Zhang L, Zhu X, Feng L, Lv Y, Chang Q, Xu Q, et al. Spatiotemporal Changes and Driver Analysis of Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands. Remote Sensing. 2022; 14(15):3563. https://doi.org/10.3390/rs14153563
Chicago/Turabian StyleLiu, Weihua, Honglin He, Xiaojing Wu, Xiaoli Ren, Li Zhang, Xiaobo Zhu, Lili Feng, Yan Lv, Qingqing Chang, Qian Xu, and et al. 2022. "Spatiotemporal Changes and Driver Analysis of Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands" Remote Sensing 14, no. 15: 3563. https://doi.org/10.3390/rs14153563
APA StyleLiu, W., He, H., Wu, X., Ren, X., Zhang, L., Zhu, X., Feng, L., Lv, Y., Chang, Q., Xu, Q., Zhang, M., Zhang, Y., & Wang, T. (2022). Spatiotemporal Changes and Driver Analysis of Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands. Remote Sensing, 14(15), 3563. https://doi.org/10.3390/rs14153563