
Driving Climatic Factors at Critical Plant Developmental Stages for Qinghai–Tibet Plateau Alpine Grassland Productivity

 ,
,
Abstract
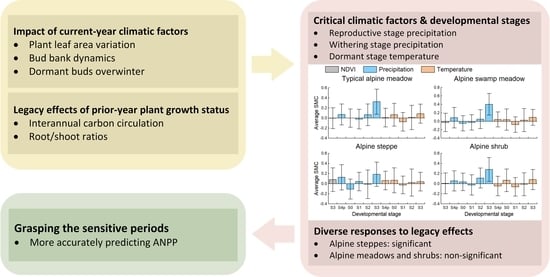
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Preprocessing
2.3. Methods
2.3.1. Developmental Stages of Grasslands on QTP
2.3.2. Indices of Input Variables
2.3.3. Impact of Driving Factors on ANPP
3. Results
3.1. Impact of Current-Year Climatic Factors
3.2. Impact of Prior-Year Plant Growth Status
4. Discussion
4.1. Comparison with Previous Studies
4.1.1. Comparison with Results of Previous Studies on QTP
4.1.2. Possible Reasons for Discrepancies from Other Studies
4.2. Low-Temperature Acclimation in the Alpine Ecosystem May Introduce a Pronounced Impact of Precipitation on ANPP Rather than Temperature
4.3. Possible Mechanisms for ANPP Responses to Climatic Factors at the Critical Developmental Stage
4.3.1. More Water Consumption from the Largest Plant Leaf Area May Lead to the Most Pronounced Impacts of S3 Stage Precipitation
4.3.2. Better Water Availability Promotes Bud Bank Size and Status at the S4 Stage Leading to a Higher ANPP
4.3.3. Higher Temperature at the S0 Stage Benefits Overwintering Buds Facilitating Plant Growth Next Year
4.4. Interannual Carbon Circulation May Decrease the ANPP Responses to Antecedent Vegetation Growth Status
4.5. Research Implications and Uncertainties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, Q.; Hu, Z.M.; Li, S.G.; Li, X.R.; Sun, X.M.; Yu, G.R. Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: Effects of mean annual precipitation and its seasonal distribution. Glob. Chang. Biol. 2012, 18, 3624–3631. [Google Scholar] [CrossRef]
- Craine, J.M.; Nippert, J.B.; Elmore, A.J.; Skibbe, A.M.; Hutchinson, S.L.; Brunsell, N.A. Timing of climate variability and grassland productivity. Proc. Natl. Acad. Sci. USA 2012, 109, 3401–3405. [Google Scholar] [CrossRef] [PubMed]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, F.; Li, Y.; Zhao, X.; Cao, G. Thirty-year variations of above-ground net primary production and precipitation-use efficiency of an alpine meadow in the north-eastern Qinghai-Tibetan Plateau. Grass Forage Sci. 2016, 71, 208–218. [Google Scholar] [CrossRef]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Bjork, R.G.; Bjorkman, A.D.; Callaghan, T.V.; Collier, L.S.; Cooper, E.J.; Cornelissen, J.H.; Day, T.A. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecol. Lett. 2012, 15, 164–175. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid. Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Wang, C.Z. A remote sensing perspective of alpine grasslands on the Tibetan Plateau: Better or worse under “Tibet Warming”? Remote Sens. Appl. Soc. Environ. 2016, 3, 36–44. [Google Scholar] [CrossRef]
- Dong, S.K.; Wang, X.X.; Liu, S.L.; Li, Y.Y.; Su, X.K.; Wen, L.; Zhu, L. Reproductive responses of alpine plants to grassland degradation and artificial restoration in the Qinghai-Tibetan Plateau. Grass Forage Sci. 2015, 70, 229–238. [Google Scholar] [CrossRef]
- Wilcox, K.R.; Blair, J.M.; Smith, M.D.; Knapp, A.K. Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions? Ecology 2016, 97, 561–568. [Google Scholar] [CrossRef]
- Adler, P.B.; White, E.P.; Cortez, M.H. Matching the forecast horizon with the relevant spatial and temporal processes and data sources. Ecography 2020, 43, 1729–1739. [Google Scholar] [CrossRef]
- Knapp, A.K.; Ciais, P.; Smith, M.D. Reconciling inconsistencies in precipitation-productivity relationships: Implications for climate change. New Phytol. 2017, 214, 41–47. [Google Scholar] [CrossRef]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Zak, J.C.; Weltzin, J.F.; et al. Convergence across biomes to a common rain-use efficiency. Nature 2004, 429, 651–654. [Google Scholar] [CrossRef]
- Munson, S.M.; Bunting, E.L.; Bradford, J.B.; Butterfield, B.J.; Gremer, J.R. Plant Production Responses to Precipitation Differ Along an Elevation Gradient and Are Enhanced Under Extremes. Ecosystems 2018, 22, 699–708. [Google Scholar] [CrossRef]
- Zhang, G.L.; Zhang, Y.J.; Dong, J.W.; Xiao, X.M. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc. Natl. Acad. Sci. USA 2013, 110, 4309–4314. [Google Scholar] [CrossRef]
- Zheng, Z.T.; Zhu, W.Q.; Zhang, Y.J. Seasonally and spatially varied controls of climatic factors on net primary productivity in alpine grasslands on the Tibetan Plateau. Glob. Ecol. Conserv. 2020, 21, e00814. [Google Scholar] [CrossRef]
- Gao, Q.Z.; Guo, Y.Q.; Xu, H.M.; Ganjurjav, H.; Li, Y.; Wan, Y.F.; Qin, X.B.; Ma, X.; Liu, S. Climate change and its impacts on vegetation distribution and net primary productivity of the alpine ecosystem in the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 554–555, 34–41. [Google Scholar] [CrossRef]
- Gao, Y.H.; Zhou, X.; Wang, Q.; Wang, C.Z.; Zhan, Z.M.; Chen, L.F.; Yan, J.X.; Qu, R. Vegetation net primary productivity and its response to climate change during 2001–2008 in the Tibetan Plateau. Sci. Total Environ. 2013, 444, 356–362. [Google Scholar] [CrossRef]
- Chen, B.X.; Zhang, X.Z.; Tao, J.; Wu, J.S.; Wang, J.S.; Shi, P.L.; Zhang, Y.J.; Yu, C.Q. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agric. For. Meteorol. 2014, 189–190, 11–18. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.G.; Yang, W.; Yang, Z.Y.; Zhang, G.X.; Piao, S.L. Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland. Int. J. Biometeorol. 2017, 61, 1433–1444. [Google Scholar] [CrossRef]
- Li, L.H.; Zhang, Y.L.; Wu, J.S.; Li, S.C.; Zhang, B.H.; Zu, J.X.; Zhang, H.M.; Ding, M.J.; Paudel, B. Increasing sensitivity of alpine grasslands to climate variability along an elevational gradient on the Qinghai-Tibet Plateau. Sci. Total Environ. 2019, 678, 21–29. [Google Scholar] [CrossRef]
- Zhe, M.; Zhang, X.Q. Time-lag effects of NDVI responses to climate change in the Yamzhog Yumco Basin, South Tibet. Ecol. Indic. 2021, 124, 107431. [Google Scholar] [CrossRef]
- Pang, G.J.; Wang, X.J.; Yang, M.X. Using the NDVI to identify variations in, and responses of, vegetation to climate change on the Tibetan Plateau from 1982 to 2012. Quat. Int. 2017, 444, 87–96. [Google Scholar] [CrossRef]
- Xu, W.X.; Gu, S.; Zhao, X.Q.; Xiao, J.S.; Tang, Y.H.; Fang, J.Y.; Zhang, J.; Jiang, S. High positive correlation between soil temperature and NDVI from 1982 to 2006 in alpine meadow of the Three-River Source Region on the Qinghai-Tibetan Plateau. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 528–535. [Google Scholar] [CrossRef]
- Van der Weide, B.L.; Hartnett, D.C.; Carter, D.L. Belowground bud banks of tallgrass prairie are insensitive to multi-year, growing-season drought. Ecosphere 2014, 5, 103. [Google Scholar] [CrossRef]
- Ott, J.P.; Klimesova, J.; Hartnett, D.C. The ecology and significance of below-ground bud banks in plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef]
- Chapin III, F.S.; Bret-Harte, M.S.; Hobbie, S.E.; Zhong, H. Plant functional types as predictors of transient responses of arctic vegetation to global change. J. Veg. Sci. 1996, 7, 347–358. [Google Scholar] [CrossRef]
- Robinson, T.M.P.; La Pierre, K.J.; Vadeboncoeur, M.A.; Byrne, K.M.; Thomey, M.L.; Colby, S.E. Seasonal, not annual precipitation drives community productivity across ecosystems. Oikos 2013, 122, 727–738. [Google Scholar] [CrossRef]
- La Pierre, K.J.; Yuan, S.; Chang, C.C.; Avolio, M.L.; Hallett, L.M.; Schreck, T.; Smith, M.D. Explaining temporal variation in above-ground productivity in a mesic grassland: The role of climate and flowering. J. Ecol. 2011, 99, 1250–1262. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.D.; Wang, C.Z.; Ji, L.; Li, J.; Wang, K.; Dai, L. The long-term trends (1982–2006) in vegetation greenness of the alpine ecosystem in the Qinghai-Tibetan Plateau. Environ. Earth Sci. 2014, 72, 1827–1841. [Google Scholar] [CrossRef]
- Jiang, W.G.; Yuan, L.H.; Wang, W.J.; Cao, R.; Zhang, Y.F.; Shen, W.M. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecol. Indic. 2015, 51, 117–126. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP estimates from NDVI for the central grassland region of the United States. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Dudney, J.; Hallett, L.M.; Larios, L.; Farrer, E.C.; Spotswood, E.N.; Stein, C.; Suding, K.N. Lagging behind: Have we overlooked previous-year rainfall effects in annual grasslands? J. Ecol. 2017, 105, 484–495. [Google Scholar] [CrossRef]
- Sherry, R.A.; Arnone, J.A.; Johnson, D.W.; Schimel, D.S.; Verburg, P.S.; Luo, Y. Carry over from previous year environmental conditions alters dominance hierarchy in a prairie plant community. J. Plant Ecol. 2012, 5, 134–146. [Google Scholar] [CrossRef]
- Tenhumberg, B.; Crone, E.E.; Ramula, S.; Tyre, A.J. Time-lagged effects of weather on plant demography: Drought and Astragalus scaphoides. Ecology 2018, 99, 915–925. [Google Scholar] [CrossRef]
- Zhong, L.; Ma, Y.M.; Xue, Y.K.; Piao, S.L. Climate Change Trends and Impacts on Vegetation Greening Over the Tibetan Plateau. J. Geophys. Res. Atmos. 2019, 124, 7540–7552. [Google Scholar] [CrossRef]
- Hoover, D.L.; Lauenroth, W.K.; Milchunas, D.G.; Porensky, L.M.; Augustine, D.J.; Derner, J.D. Sensitivity of productivity to precipitation amount and pattern varies by topographic position in a semiarid grassland. Ecosphere 2021, 12, e03376. [Google Scholar] [CrossRef]
- Sherry, R.A.; Weng, E.; Arnone Iii, J.A.; Johnson, D.W.; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y.Q. Lagged effects of experimental warming and doubled precipitation on annual and seasonal aboveground biomass production in a tallgrass prairie. Glob. Chang. Biol. 2008, 14, 2923–2936. [Google Scholar] [CrossRef]
- Sun, Q.Q.; Meyer, W.S.; Marschner, P. Direct and carry-over effects of summer rainfall on ecosystem carbon uptake and water use efficiency in a semi-arid woodland. Agric. For. Meteorol. 2018, 263, 15–24. [Google Scholar] [CrossRef]
- Chen, T.; de Jeu, R.A.M.; Liu, Y.Y.; van der Werf, G.R.; Dolman, A.J. Using satellite based soil moisture to quantify the water driven variability in NDVI: A case study over mainland Australia. Remote Sens. Environ. 2014, 140, 330–338. [Google Scholar] [CrossRef]
- Saatchi, S.; Asefi-Najafabady, S.; Malhi, Y.; Aragao, L.E.; Anderson, L.O.; Myneni, R.B.; Nemani, R. Persistent effects of a severe drought on Amazonian forest canopy. Proc. Natl. Acad. Sci. USA. 2013, 110, 565–570. [Google Scholar] [CrossRef]
- Evans, S.E.; Burke, I.C. Carbon and nitrogen decoupling under an 11-year drought in the shortgrass steppe. Ecosystems 2013, 16, 20–33. [Google Scholar] [CrossRef]
- Giese, M.; Gao, Y.Z.; Lin, S.; Brueck, H. Nitrogen availability in a grazed semi-arid grassland is dominated by seasonal rainfall. Plant Soil 2010, 340, 157–167. [Google Scholar] [CrossRef]
- Shen, W.; Jenerette, G.D.; Hui, D.; Scott, R.L. Precipitation legacy effects on dryland ecosystem carbon fluxes: Direction, magnitude and biogeochemical carryovers. Biogeosciences 2016, 13, 425–439. [Google Scholar] [CrossRef]
- Wiegand, T.; Snyman, H.A.; Kellner, K.; Paruelo, J.M. Do grasslands have a memory: Modeling phytomass production of a semiarid South African grassland. Ecosystems 2004, 7, 243–258. [Google Scholar] [CrossRef]
- Gong, Y.H.; Zhao, D.M.; Ke, W.B.; Fang, C.; Pei, J.Y.; Sun, G.J.; Ye, J.S. Legacy effects of precipitation amount and frequency on the aboveground plant biomass of a semi-arid grassland. Sci. Total Environ. 2020, 705, 135899. [Google Scholar] [CrossRef]
- Han, J.J.; Chen, J.Q.; Shi, W.Y.; Song, J.; Hui, D.F.; Ru, J.Y.; Wan, S.Q. Asymmetric responses of resource use efficiency to previous-year precipitation in a semi-arid grassland. Funct. Ecol. 2021, 35, 807–814. [Google Scholar] [CrossRef]
- Shi, Z.; Lin, Y.; Wilcox, K.R.; Souza, L.; Jiang, L.F.; Jiang, J.; Jung, C.G.; Xu, X.; Yuan, M.T.; Guo, X. Successional change in species composition alters climate sensitivity of grassland productivity. Glob. Chang. Biol. 2018, 24, 4993–5003. [Google Scholar] [CrossRef]
- Reichmann, L.G.; Sala, O.E.; Peters, D.P.C. Precipitation legacies in desert grassland primary production occur through previous-year tiller density. Ecology 2013, 94, 435–443. [Google Scholar] [CrossRef]
- Reichmann, L.G.; Sala, O.E.; Whitehead, D. Differential sensitivities of grassland structural components to changes in precipitation mediate productivity response in a desert ecosystem. Funct. Ecol. 2014, 28, 1292–1298. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Gao, Z.Y.; Lin, Z.J.; Niu, F.J.; Luo, J. Soil water dynamics in the active layers under different land-cover types in the permafrost regions of the Qinghai–Tibet Plateau, China. Geoderma 2020, 364, 114176. [Google Scholar] [CrossRef]
- Yahdjian, L.; Sala, O.E. Size of Precipitation Pulses Controls Nitrogen Transformation and Losses in an Arid Patagonian Ecosystem. Ecosystems 2010, 13, 575–585. [Google Scholar] [CrossRef]
- Diaz-Toribio, M.H.; Putz, F.E. Underground carbohydrate stores and storage organs in fire-maintained longleaf pine savannas in Florida, USA. Am. J. Bot. 2021, 108, 432–442. [Google Scholar] [CrossRef]
- De Moraes, M.G.; de Carvalho, M.A.M.; Franco, A.C.; Pollock, C.J.; Figueiredo-Ribeiro, R.; de Cássia, L. Fire and Drought: Soluble Carbohydrate Storage and Survival Mechanisms in Herbaceous Plants from the Cerrado. BioScience 2016, 66, 107–117. [Google Scholar] [CrossRef]
- Wyka, T. Carbohydrate storage and use in an alpine population of the perennial herb, Oxytropis sericea. Oecologia 1999, 120, 198–208. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2003; ISBN 978-3642189692. [Google Scholar]
- Mou, X.M.; Li, X.G.; Zhao, N.N.; Yu, Y.W.; Kuzyakov, Y. Tibetan sedges sequester more carbon belowground than grasses: A 13C labeling study. Plant Soil 2018, 426, 287–298. [Google Scholar] [CrossRef]
- Song, M.H.; Hu, Q.W.; Tian, Y.Q.; Ouyang, H. Seasonal patterns of root and shoot interactions in an alpine meadow on the Tibetan Plateau. J. Plant Ecol. 2010, 5, 182–190. [Google Scholar] [CrossRef]
- You, Q.G.; Xue, X.; Peng, F.; Xu, M.H.; Duan, H.C.; Dong, S.Y. Comparison of ecosystem characteristics between degraded and intact alpine meadow in the Qinghai-Tibetan Plateau, China. Ecol. Eng. 2014, 71, 133–143. [Google Scholar] [CrossRef]
- Li, J.; Zhang, F.W.; Lin, L.; Li, H.Q.; Du, Y.G.; Li, Y.K.; Cao, G.M. Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the Qinghai–Tibetan Plateau, China. Ecol. Res. 2015, 30, 589–596. [Google Scholar] [CrossRef]
- Zeng, C.X.; Wu, J.S.; Zhang, X.Z. Effects of Grazing on Above-vs. Below-Ground Biomass Allocation of Alpine Grasslands on the Northern Tibetan Plateau. PLoS ONE 2015, 10, e0135173. [Google Scholar] [CrossRef]
- Zhou, X.M. Chinese Kobresia Meadow; Science Press: Beijing, China, 2001; ISBN 9787030085702. (In Chinese) [Google Scholar]
- Zhang, T.; Li, B.L.; Yuan, Y.C.; Gao, X.Z.; Sun, Q.L.; Xu, L.L.; Jiang, Y.H. Spatial downscaling of TRMM precipitation data considering the impacts of macro-geographical factors and local elevation in the Three-River Headwaters Region. Remote Sens. Environ. 2018, 215, 109–127. [Google Scholar] [CrossRef]
- Qin, D.H. Ecological Protection and Sustainable Development in the Three-River Headwater; Regin Science Press: Beijing, China, 2014; ISBN 9787030387189. (In Chinese) [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Stow, D.; Petersen, A.; Hope, A.; Engstrom, R.; Coulter, L. Greenness trends of Arctic tundra vegetation in the 1990s: Comparison of two NDVI data sets from NOAA AVHRR systems. Int. J. Remote Sens. 2010, 28, 4807–4822. [Google Scholar] [CrossRef]
- Montandon, L.M.; Small, E.E. The impact of soil reflectance on the quantification of the green vegetation fraction from NDVI. Remote Sens. Environ. 2008, 112, 1835–1845. [Google Scholar] [CrossRef]
- Piao, S.L.; Mohammat, A.; Fang, J.Y.; Cai, Q.; Feng, J.M. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; He, J.S. Variations in Vegetation Net Primary Production in the Qinghai-Xizang Plateau, China, from 1982 to 1999. Clim. Chang. 2006, 74, 253–267. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Wen, Z.M.; Chen, Y.M.; Cao, Y.; Ren, J.Y. Spatiotemporal change and trend analysis of potential evapotranspiration over the Loess Plateau of China during 2011–2100. Agric. For. Meteorol. 2017, 233, 183–194. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Liu, W.Z.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Zhang, X.S. 1:1 Million Vegetation Map of CHINA; Geological Publishing House: Beijing, China, 2008; ISBN 978-7-116-05146-1. [Google Scholar]
- Xu, W.; Zhu, M.Y.; Zhang, Z.H.; Ma, Z.Y.; Liu, H.Y.; Chen, L.T.; Cao, G.M.; Zhao, X.Q.; Schmid, B.; He, J.S. Experimentally simulating warmer and wetter climate additively improves rangeland quality on the Tibetan Plateau. J. Appl. Ecol. 2018, 55, 1486–1497. [Google Scholar] [CrossRef]
- Klein, J.A.; Harte, J.; Zhao, X.Q. Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecol. Appl. 2007, 17, 541–557. [Google Scholar] [CrossRef]
- Hu, Z.M.; Yu, G.R.; Fan, J.W.; Zhong, H.P.; Wang, S.Q.; Li, S.G. Precipitation-use efficiency along a 4500-km grassland transect. Glob. Ecol. Biogeogr. 2010, 19, 842–851. [Google Scholar] [CrossRef]
- Wang, S.P.; Duan, J.C.; Xu, G.P.; Wang, Y.F.; Zhang, Z.H.; Rui, Y.C.; Luo, C.Y.; Xu, B.; Zhu, X.X.; Chang, X.F. Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 2012, 93, 2365–2376. [Google Scholar] [CrossRef]
- Qiu, B.; Li, W.K.; Wang, X.Q.; Shang, L.Y.; Song, C.Q.; Guo, W.D.; Zhang, Y.G. Satellite-observed solar-induced chlorophyll fluorescence reveals higher sensitivity of alpine ecosystems to snow cover on the Tibetan Plateau. Agric. For. Meteorol. 2019, 271, 126–134. [Google Scholar] [CrossRef]
- Bell, K.L.; Bliss, L.C. Autecology of Kobresia bellardii: Why winter snow accumulation limits local distribution. Ecol. Monogr. 1979, 49, 377–402. [Google Scholar] [CrossRef]
- Yang, Y.W.; Li, X.L. Pilot study of clonal growth and reproduction of Kobresia humilis. J. Qinghai Univ. 2008, 26, 31–34. (In Chinese) [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Hunt, H.W.; Swift, D.M.; Singh, J.S. Estimating aboveground net primary production in grasslands: A simulation approach. Ecol. Model. 1986, 33, 297–314. [Google Scholar] [CrossRef]
- Singh, J.S.; Lauenroth, W.K.; Steinhorst, R.K. Review and assessment of various techniques for estimating net aerial primary production in grasslands from harvest data. Bot. Rev. 1975, 41, 181–232. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Sala, O.E.; Paruelo, J.M. Patterns and controls of primary production in the Patagonian steppe: A remote sensing approach. Ecology 2002, 83, 307–319. [Google Scholar] [CrossRef]
- Piao, S.L.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Oesterheld, M.; Loreti, J.; Semmartin, M.; Sala, O.E. Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. J. Veg. Sci. 2001, 12, 137–142. [Google Scholar] [CrossRef]
- Griffin-Nolan, R.J.; Carroll, C.J.W.; Denton, E.M.; Johnston, M.K.; Collins, S.L.; Smith, M.D.; Knapp, A.K. Legacy effects of a regional drought on aboveground net primary production in six central US grasslands. Plant Ecol. 2018, 219, 505–515. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.D.; Shi, P.J.; Singh, V.P.; Sun, P. Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Guo, L.; Cheng, J.M.; Luedeling, E.; Koerner, S.E.; He, J.S.; Xu, J.C.; Gang, C.C.; Li, W.; Luo, R.M.; Peng, C.H. Critical climate periods for grassland productivity on China’s Loess Plateau. Agric. For. Meteorol. 2017, 233, 101–109. [Google Scholar] [CrossRef]
- Wang, L.; Yu, H.Y.; Zhang, Q.; Xu, Y.J.; Tao, Z.X.; Alatalo, J.H.; Dai, J.H. Responses of aboveground biomass of alpine grasslands to climate changes on the Qinghai-Tibet Plateau. J. Geogr. Sci. 2018, 28, 1953–1964. [Google Scholar] [CrossRef]
- Luedeling, E.; Gassner, A. Partial Least Squares Regression for analyzing walnut phenology in California. Agric. For. Meteorol. 2012, 158–159, 43–52. [Google Scholar] [CrossRef]
- Wold, S. PLS for multivariate linear modeling. In Chemometric Methods in Molecular Design: Methods and Principles in Medicinal Chemistry; VCH Verlagsgesellschaft mbH: Weinheim, Germany, 1995; pp. 195–218. [Google Scholar]
- Bai, Y.; Guo, C.; Degen, A.A.; Ahmad, A.A.; Wang, W.; Zhang, T.; Li, W.; Ma, L.; Huang, M.; Zeng, H.; et al. Climate warming benefits alpine vegetation growth in Three-River Headwater Region, China. Sci. Total Environ. 2020, 742, 140574. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Zhang, X.X. Alpine grasslands response to climatic factors and anthropogenic activities on the Tibetan Plateau from 2000 to 2012. Ecol. Eng. 2016, 92, 251–259. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.B.; Wang, Z.Q.; Chen, Y.Z.; Gang, C.C.; An, R.; Li, J.L. Vegetation dynamics and its driving forces from climate change and human activities in the Three-River Source Region, China from 1982 to 2012. Sci. Total Environ. 2016, 563–564, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.; Ma, Y.; Ma, W.; Liang, C.; Flynn, D.F.B.; Schmid, B.; Fang, J.; He, J.S. Field-based observations of regional-scale, temporal variation in net primary production in Tibetan alpine grasslands. Biogeosciences 2014, 11, 2003–2016. [Google Scholar] [CrossRef]
- Li, P.L.; Hu, Z.M.; Liu, Y.W. Shift in the trend of browning in Southwestern Tibetan Plateau in the past two decades. Agric. For. Meteorol. 2020, 287, 107950. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X.; Zhang, X.Z. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau. Agric. For. Meteorol. 2018, 249, 11–21. [Google Scholar] [CrossRef]
- Yuan, X.; Chen, Y.; Qin, W.K.; Xu, T.L.; Mao, Y.H.; Wang, Q.; Chen, K.L.; Zhu, B. Plant and microbial regulations of soil carbon dynamics under warming in two alpine swamp meadow ecosystems on the Tibetan Plateau. Sci. Total Environ. 2021, 790, 148072. [Google Scholar] [CrossRef]
- Yang, Y.; Hopping, K.A.; Wang, G.; Chen, J.; Peng, A.; Klein, J.A. Permafrost and drought regulate vulnerability of Tibetan Plateau grasslands to warming. Ecosphere 2018, 9, e02233. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.J.; Xu, M.J.; Zhu, J.T.; Chen, N.; Jiang, Y.B.; Huang, K.; Zu, J.X.; Liu, Y.J.; Yu, G.R. Water availability is more important than temperature in driving the carbon fluxes of an alpine meadow on the Tibetan Plateau. Agric. For. Meteorol. 2018, 256–257, 22–31. [Google Scholar] [CrossRef]
- Wu, Z.T.; Dijkstra, P.; Koch, G.W.; PeÑUelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Li, P.L.; Zhu, D.; Wang, Y.L.; Liu, D. Elevation dependence of drought legacy effects on vegetation greenness over the Tibetan Plateau. Agric. For. Meteorol. 2020, 295, 108190. [Google Scholar] [CrossRef]
- Fensholt, R.; Proud, S.R. Evaluation of earth observation based global long term vegetation trends—Comparing GIMMS and MODIS global NDVI time series. Remote Sens. Environ. 2012, 119, 131–147. [Google Scholar] [CrossRef]
- Liu, Y.C.; Li, Z.; Chen, Y.N.; Li, Y.P.; Li, H.W.; Xia, Q.Q.; Kayumba, P.M. Evaluation of consistency among three NDVI products applied to High Mountain Asia in 2000–2015. Remote Sens. Environ. 2022, 269, 112821. [Google Scholar] [CrossRef]
- Tian, F.; Fensholt, R.; Verbesselt, J.; Grogan, K.; Horion, S.; Wang, Y.J. Evaluating temporal consistency of long-term global NDVI datasets for trend analysis. Remote Sens. Environ. 2015, 163, 326–340. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.Y.; Huang, N.; Bi, J.; Ma, X.L.; Ma, Z.Y.; Shangguan, Z.J.; Zhao, H.F.; Feng, Q.S.; Liang, T.G. Satellite-derived NDVI underestimates the advancement of alpine vegetation growth over the past three decades. Ecology 2021, 102, e03518. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Yamori, W. Thermal acclimation of photosynthesis: On the importance of adjusting our definitions and accounting for thermal acclimation of respiration. Photosynth. Res. 2014, 119, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Björk, R.G.; Boulanger-Lapointe, N.; Cooper, E.J.; Cornelissen, J.H.C.; Day, T.A.; Dorrepaal, E.; Elumeeva, T.G. Plot-scale evidence of tundra vegetation change and links to recent summer warming. Nat. Clim. Chang. 2012, 2, 453–457. [Google Scholar] [CrossRef]
- Yamasaki, T.; Yamakawa, T.; Yamane, Y.; Koike, H.; Satoh, K.; Katoh, S. Temperature acclimation of photosynthesis and related changes in photosystem II electron transport in winter wheat. Plant Physiol. 2002, 128, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Mawson, B.T.; Cummins, W.R. Thermal acclimation of photosynthetic electron transport activity by thylakoids of Saxifraga cernua. Plant Physiol. 1989, 89, 325–332. [Google Scholar] [CrossRef]
- Erschbamer, B.; Kiebacher, T.; Mallaun, M.; Unterluggauer, P. Short-term signals of climate change along an altitudinal gradient in the South Alps. Plant Ecol. 2009, 202, 79–89. [Google Scholar] [CrossRef]
- Theurillat, J.P.; Guisan, A. Potential impact of climate change on vegetation in the European Alps: A review. Clim. Chang. 2001, 50, 77–109. [Google Scholar] [CrossRef]
- Billings, W.D.; Mooney, H.A. The ecology of arctic and alpine plants. Biol. Rev. 1968, 43, 481–529. [Google Scholar] [CrossRef]
- Pisek, A.; Larcher, W.; Vegis, A.; Napp-Zinn, K. The normal temperature range. In Temperature and Life; Springer: Berlin/Heidelberg, Germany, 1973; pp. 102–194. [Google Scholar]
- Kammer, P.M.; Schöb, C.; Eberhard, G.; Gallina, R.; Meyer, R.; Tschanz, C. The relationship between soil water storage capacity and plant species diversity in high alpine vegetation. Plant Ecol. Divers. 2013, 6, 457–466. [Google Scholar] [CrossRef]
- Liu, Y.W.; Geng, X.D.; Tenzintarchen; Wei, D.; Dai, D.X.; Xu, R. Divergence in ecosystem carbon fluxes and soil nitrogen characteristics across alpine steppe, alpine meadow and alpine swamp ecosystems in a biome transition zone. Sci. Total Environ. 2020, 748, 142453. [Google Scholar] [CrossRef]
- Hu, Z.M.; Yu, G.R.; Zhou, Y.L.; Sun, X.M.; Li, Y.N.; Shi, P.L.; Wang, Y.F.; Song, X.; Zheng, Z.M.; Zhang, L.; et al. Partitioning of evapotranspiration and its controls in four grassland ecosystems: Application of a two-source model. Agric. For. Meteorol. 2009, 149, 1410–1420. [Google Scholar] [CrossRef]
- Li, H.Q.; Wang, C.Y.; Zhang, F.W.; He, Y.T.; Shi, P.L.; Guo, X.W.; Wang, J.B.; Zhang, L.M.; Li, Y.N.; Cao, G.M.; et al. Atmospheric water vapor and soil moisture jointly determine the spatiotemporal variations of CO2 fluxes and evapotranspiration across the Qinghai-Tibetan Plateau grasslands. Sci. Total Environ. 2021, 791, 148379. [Google Scholar] [CrossRef]
- Qin, Y.; Yi, S.; Ren, S.; Li, N.; Chen, J. Responses of typical grasslands in a semi-arid basin on the Qinghai-Tibetan Plateau to climate change and disturbances. Environ. Earth Sci. 2013, 71, 1421–1431. [Google Scholar] [CrossRef]
- Wang, J.F.; Shi, Y.J.; Ao, Y.N.; Yu, D.F.; Wang, J.; Gao, S.; Knops, J.M.H.; Mu, C.S.; Li, Z.J. Summer drought decreases Leymus chinensis productivity through constraining the bud, tiller and shoot production. J. Agron. Crop Sci. 2019, 205, 554–561. [Google Scholar] [CrossRef]
- Benson, E.J.; Hartnett, D.C. The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie. Plant Ecol. 2006, 187, 163–178. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Hartnett, D.C. Below-ground bud banks increase along a precipitation gradient of the North American Great Plains: A test of the meristem limitation hypothesis. New Phytol. 2006, 171, 81–89. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef]
- Busso, C.A.; Mueller, R.J.; Richards, J.H. Effects of drought and defoliation on bud viability in two caespitose grasses. Ann. Bot. 1989, 63, 477–485. [Google Scholar] [CrossRef]
- Deng, Z.F.; Xie, X.L.; Zhou, X.M. Primary Study on Reproductive Strategies of Kobresia humilis Population in Alpine Meadow. Chin. J. Ecol. 2001, 20, 68–70. (In Chinese) [Google Scholar] [CrossRef]
- Ott, J.P.; Hartnett, D.C. Contrasting bud bank dynamics of two co-occurring grasses in tallgrass prairie: Implications for grassland dynamics. Plant Ecol. 2012, 213, 1437–1448. [Google Scholar] [CrossRef]
- Malyshev, A.V.; Henry, H.A.L. Frost damage and winter nitrogen uptake by the grass Poa pratensis L.: Consequences for vegetative versus reproductive growth. Plant Ecol. 2012, 213, 1739–1747. [Google Scholar] [CrossRef]
- Rixen, C.; Dawes, M.A.; Wipf, S.; Hagedorn, F. Evidence of enhanced freezing damage in treeline plants during six years of CO2 enrichment and soil warming. Oikos 2012, 121, 1532–1543. [Google Scholar] [CrossRef]
- Wheeler, J.A.; Hoch, G.; Cortes, A.J.; Sedlacek, J.; Wipf, S.; Rixen, C. Increased spring freezing vulnerability for alpine shrubs under early snowmelt. Oecologia 2014, 175, 219–229. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Li, B.L.; Yuan, Y.C.; Sun, Q.L.; Zhang, T.; Liu, Y.; Li, Y.; Li, R. Divergent shifts in flowering phenology of herbaceous plants on the warming Qinghai–Tibetan plateau. Agric. For. Meteorol. 2021, 307, 108502. [Google Scholar] [CrossRef]
- Bjorkman, A.D.; Elmendorf, S.C.; Beamish, A.L.; Vellend, M.; Henry, G.H. Contrasting effects of warming and increased snowfall on Arctic tundra plant phenology over the past two decades. Glob. Chang. Biol. 2015, 21, 4651–4661. [Google Scholar] [CrossRef]
- Esteban, P.; Jones, P.D.; Martín-Vide, J.; Mases, M. Atmospheric circulation patterns related to heavy snowfall days in Andorra, Pyrenees. Int. J. Climatol. 2005, 25, 319–329. [Google Scholar] [CrossRef]
- Ma, N.; Yu, K.L.; Zhang, Y.S.; Zhai, J.Q.; Zhang, Y.Q.; Zhang, H.B. Ground observed climatology and trend in snow cover phenology across China with consideration of snow-free breaks. Clim. Dyn. 2020, 55, 2867–2887. [Google Scholar] [CrossRef]
- Li, G.D. The study of underground phytomass and its seasonal changes in alpine grass—Kobresia meadow of Tianzhu area, Gansu. Cao Ye Xue Bao 1990, 1, 42–46. (In Chinese) [Google Scholar]
- Pu, J.Y.; Li, Y.N.; Zhao, L.; Yang, S.H. The relationship between seasonal changes of Kobresia humilis meadow biomass and the meteorological factors. Acta Agrestia Sin. 2005, 13, 238–241. (In Chinese) [Google Scholar] [CrossRef]
- Zhu, B.W.; Zhou, H.K.; Xu, Y.X.; Li, Y.N.; Tang, K. Study on seasonal dynamics of biomass in meadow grassland of north shore of Qinghai lake. Pratacult. Sci. 2008, 25, 62–66. (In Chinese) [Google Scholar] [CrossRef]
- Wu, Y.B.; Tan, H.C.; Deng, Y.C.; Wu, J.; Xu, X.L.; Wang, Y.F.; Tang, Y.H.; Higashi, T.R.; Cui, X.Y. Partitioning pattern of carbon flux in a Kobresia grassland on the Qinghai-Tibetan Plateau revealed by field 13C pulse-labeling. Glob. Chang. Biol. 2009, 16, 2322–2333. [Google Scholar] [CrossRef]
- Lenney, M.P.; Woodcock, C.E.; Collins, J.B.; Hamdi, H. The status of agricultural lands in Egypt: The use of multitemporal NDVI features derived from Landsat TM. Remote Sens. Environ. 1996, 56, 8–20. [Google Scholar] [CrossRef]
- Li, L.H.; Zhang, Y.L.; Liu, L.S.; Wu, J.S.; Li, S.C.; Zhang, H.Y.; Zhang, B.H.; Ding, M.J.; Wang, Z.F.; Paudel, B. Current challenges in distinguishing climatic and anthropogenic contributions to alpine grassland variation on the Tibetan Plateau. Ecol. Evol. 2018, 8, 5949–5963. [Google Scholar] [CrossRef]
- Guo, B.; Zang, W.Q.; Yang, F.; Han, B.M.; Chen, S.T.; Liu, Y.; Yang, X.; He, T.L.; Chen, X.; Liu, C.T.; et al. Spatial and temporal change patterns of net primary productivity and its response to climate change in the Qinghai-Tibet Plateau of China from 2000 to 2015. J. Arid. Land 2019, 12, 1–17. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.D.; Ji, L.; Lei, L.P.; Wang, C.Z.; Yan, D.M.; Li, B.; Li, J. Vegetation greenness trend (2000 to 2009) and the climate controls in the Qinghai-Tibetan Plateau. J. Appl. Remote Sens. 2013, 7, 73572. [Google Scholar] [CrossRef]
- Yang, Y.H.; Piao, S.L. Variations in grassland vegetation cover in relation to climatic factors on the Tibetan Plateau. J. Plant Ecol. 2006, 30, 1–8. (In Chinese) [Google Scholar] [CrossRef]
- Du, J.Q.; Zhao, C.X.; Shu, J.M.; Jiaerheng, A.; Yuan, X.J.; Yin, J.Q.; Fang, S.F.; He, P. Spatiotemporal changes of vegetation on the Tibetan Plateau and relationship to climatic variables during multiyear periods from 1982–2012. Environ. Earth Sci. 2015, 75. [Google Scholar] [CrossRef]
- Xu, W.X.; Liu, X.D. Response of vegetation in the Qinghai-Tibet Plateau to global warming. Chin. Geogr. Sci. 2007, 17, 151–159. [Google Scholar] [CrossRef]
- Ding, M.J.; Zhang, Y.L.; Liu, L.S.; Zhang, W.; Wang, Z.F.; Bai, W.Q. The relationship between NDVI and precipitation on the Tibetan Plateau. J. Geogr. Sci. 2007, 17, 259–268. [Google Scholar] [CrossRef]
- Gao, Q.Z.; Li, Y.; Wan, Y.F.; Qin, X.B.; Jiangcun, W.Z.; Liu, Y.H. Dynamics of alpine grassland NPP and its response to climate change in Northern Tibet. Clim. Change 2009, 97, 515–528. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, D.; Gao, X.; Li, B.; Yuan, Y.; Jiang, Y.; Liu, Y.; Li, Y.; Li, R.; Liu, W.; Xu, J. Driving Climatic Factors at Critical Plant Developmental Stages for Qinghai–Tibet Plateau Alpine Grassland Productivity. Remote Sens. 2022, 14, 1564. https://doi.org/10.3390/rs14071564
Zhai D, Gao X, Li B, Yuan Y, Jiang Y, Liu Y, Li Y, Li R, Liu W, Xu J. Driving Climatic Factors at Critical Plant Developmental Stages for Qinghai–Tibet Plateau Alpine Grassland Productivity. Remote Sensing. 2022; 14(7):1564. https://doi.org/10.3390/rs14071564
Chicago/Turabian StyleZhai, Dechao, Xizhang Gao, Baolin Li, Yecheng Yuan, Yuhao Jiang, Yan Liu, Ying Li, Rui Li, Wei Liu, and Jie Xu. 2022. "Driving Climatic Factors at Critical Plant Developmental Stages for Qinghai–Tibet Plateau Alpine Grassland Productivity" Remote Sensing 14, no. 7: 1564. https://doi.org/10.3390/rs14071564
APA StyleZhai, D., Gao, X., Li, B., Yuan, Y., Jiang, Y., Liu, Y., Li, Y., Li, R., Liu, W., & Xu, J. (2022). Driving Climatic Factors at Critical Plant Developmental Stages for Qinghai–Tibet Plateau Alpine Grassland Productivity. Remote Sensing, 14(7), 1564. https://doi.org/10.3390/rs14071564