
Effects of Increasing C4-Crop Cover and Stomatal Conductance on Evapotranspiration: Simulations for a Lake Erie Watershed

 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
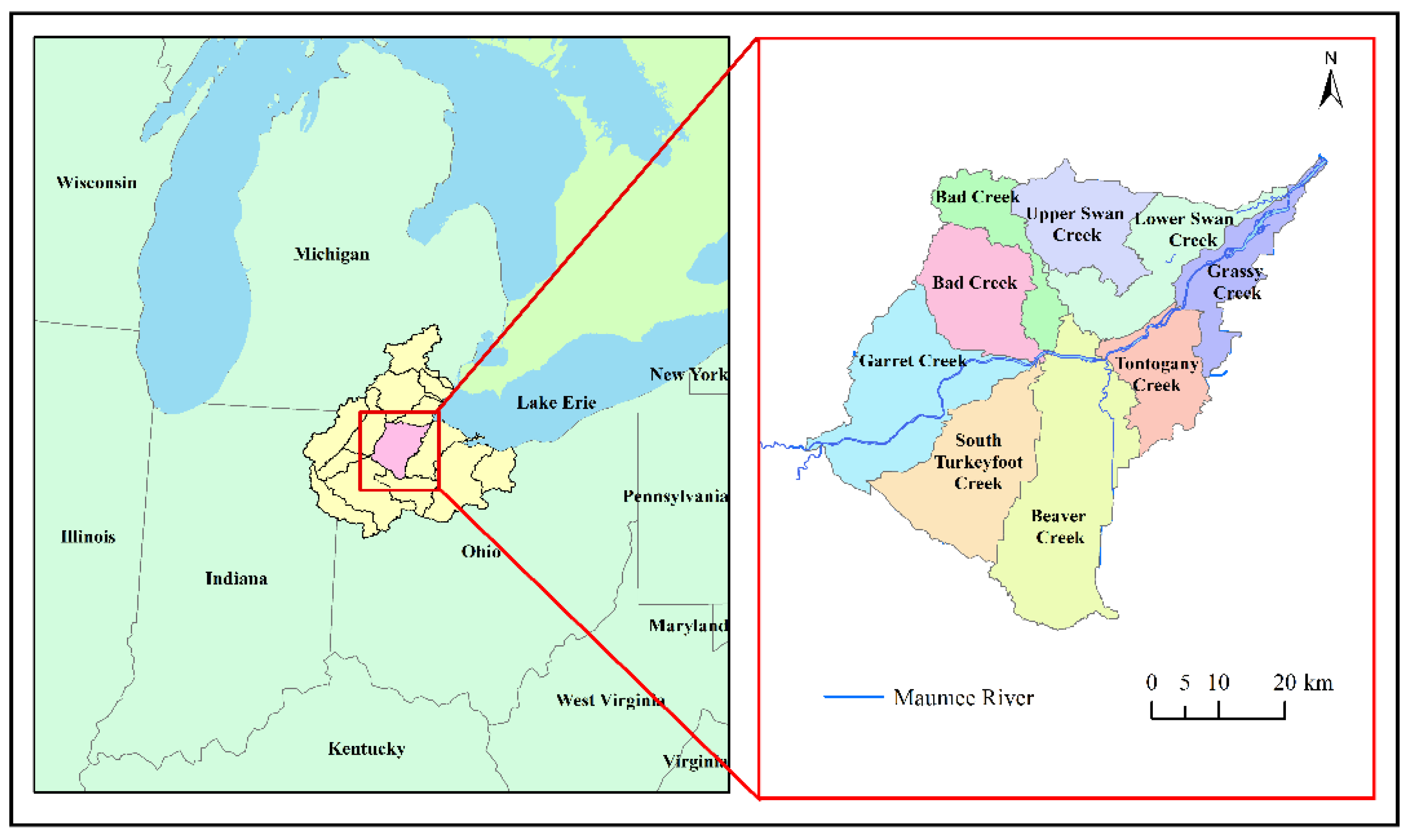
2.1. Description of Study Area
2.2. Data
2.2.1. Meteorological Data
2.2.2. Satellite Imagery Data and Products
2.2.3. Land Use/Land Cover (LULC) Data
2.2.4. Leaf Area Index
2.2.5. ECOSTRESS Data
2.2.6. Available Water Holding Capacity (AWHC) Data
2.3. Evapotranspiration Modeling
2.3.1. BEPS Model
2.3.2. Hypothetical Impact Analyses—Alterations in Stomatal Conductance and Corn Cover Extent
3. Results
3.1. Estimated LAI
3.2. Estimated ET Rates under Different Stomatal Conductance Values
3.3. Estimated ET Rates under Different Percentages of Corn Cover
3.4. Comparison of Daily BEPS-Generated ET and ECOSTRESS ET Product
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kanae, S.; Oki, T. Global hydrological cycles and world water resources. Science 2006, 313, 1068–1072. [Google Scholar] [CrossRef] [Green Version]
- U.S. Geological Survey. Science for a Changing World. 2020. Available online: https://www.usgs.gov (accessed on 12 December 2020).
- Irmak, S. Evapotranspiration Basics and Estimating Actual Crop Evapotranspiration from Reference Evapotranspiration and Crop-Specific Coefficients; G1994 Index: Crops, Irrigation Engineering—UNL Extension 2017. Available online: https://extensionpublications.unl.edu/assets/pdf/g1994.pdf (accessed on 2 January 2022).
- Liu, J.; Chen, J.M.; Cihlar, J.; Chen, W. Net primary productivity mapped for Canada at 1-km resolution. Glob. Ecol. Biogeogr. 2002, 11, 115–129. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, J.M.; Chillar, J.; Park, W.M. Mapping evapotranspiration based on remote sensing: An application to Canada Landmass. Water Resour. 2003, 39, 1189. [Google Scholar] [CrossRef]
- Myneni, R.B.; Yang, W.; Nemani, R.R.; Huete, A.R.; Dickinson, R.E.; Knyazikhin, Y.; Didan, K.; Fu, R.; Negrón Juárez, R.I.; Saatchi, S.S.; et al. Large seasonal swings in leaf area of Amazon rainforests. Proc. Natl. Acad. Sci. USA 2007, 104, 4820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, Y.; Kar, S.K. Evapotranspiration estimation with remote sensing and various surface energy balance algorithms—A review. Energies 2014, 7, 2821–2849. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Black, T.A. Measuring leaf area index of plant canopies with branch architecture. Agric. For. Meteorol. 1991, 57, 1–12. [Google Scholar] [CrossRef]
- Chen, J.M.; Menges, C.H.; Leblanc, S.G. Global mapping of foliage clumping index using multi-angular satellite data. Remote Sens. Environ. 2005, 97, 447–457. [Google Scholar] [CrossRef]
- Chen, J.M.; Liu, J. Evolution of evapotranspiration models using thermal and shortwave remote sensing data. Remote Sens. Environ. 2020, 237, 111594. [Google Scholar] [CrossRef]
- Govind, A.; Chen, J.M.; Margolis, H.; Ju, W.; Sonnentag, O.; Giasson, M. A spatially explicit hydro-ecological modeling framework (BEPS-Terrainlab V2.0): Model description and test in a Boreal ecosystem in Eastern North America. J. Hydrol. 2009, 367, 200–216. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 response of stomata and its dependence on environmental factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper; FAO: Rome, Italy, 1998; Volume 56, pp. 1–155. [Google Scholar]
- Urban, j.; Miles Ingwers, M.; McGuire, M.A.; Teskey, R.O. Stomatal conductance increases with rising temperature. Plant Signal. Behavior. 2017, 12, e1356534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damour, G.; Simonneau, T.; Cochard, H.; Urban, L. An overview of models of stomatal conductance at the leaf level. Plant Cell Environ. 2010, 33, 1419–1438. [Google Scholar] [CrossRef]
- Hoorman, J.J.; Islam, R.; Sundermeier, A. Sustainable Crop Rotations with Cover Crops. Agriculture and Natural Resources. Available online: https://ohioline.osu.edu/factsheet/SAG-9 (accessed on 20 October 2021).
- Busari, M.A.; Kukal, S.S.; Kaur, A.; Bhatt, R.; Ally Dulazib, A.A. Conservation tillage impacts on soil, crop and the environment. Int. Soil Water Conserv. Res. 2015, 3, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Qin, J.; Li, J.; Zhao, Y.; Nie, L.; Zhang, Z. Interactions between soil water content and fertilizer on growth characteristics and biomass yield of Chinese White Poplar (Populus tomentosa Carr.) Seedlings. Soil Sci. Plant Nutr. 2011, 57, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Irmak, S.; Rudnick, D. Corn Soil Water Extraction and Effective Rooting Depth in a Silty-Loam Soil; Nebraska Extension, Agriculture and Natural Resources. G2245. 2014. Available online: http://extensionpublications.unl.edu/assets/pdf/g2245.pdf (accessed on 3 January 2022).
- Zou, M.; Niu, J.; Kang, S. The contribution of human agricultural activities to increasing evapotranspiration is significantly greater than climate change effect over Heihe agricultural region. Sci. Rep. 2017, 7, 8805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahajpal, R.; Zhang, X.; Izaurralde, R.C.; Gelfand, I.; Hurtt, G.C. Identifying representative crop rotation patterns and grassland loss in the US western corn belt. Comput. Electron. Agric. 2014, 108, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Mladenoff, D.J.; Sahajpal, R.; Johnson, C.P.; Rothstein, D.E. Recent land use change to agriculture in the U.S. Lake states: Impacts on cellulosic biomass potential and natural lands. PLoS ONE 2016, 11, e0148566. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Sheshukov, A.Y.; Yen, H.; Kastens, J.H.; Peterson, D.L. Impacts of incorporating dominant crop rotation patterns as primary land use change on hydrologic model performance. Agric. Ecosyst. Environ. 2017, 247, 33–42. [Google Scholar] [CrossRef]
- USDA-FAS. United States Department of Agriculture—Foreign Agricultural Service. Available online: https://www.fas.usda.gov (accessed on 12 June 2020).
- USEIA. U.S. Energy Information Administration. Available online: https://www.eia.gov/todayinenergy/detail.php?id=32152 (accessed on 18 February 2021).
- Ueno, O. Environmental regulation of C3 and C4 differentiation in the amphibious Sedge Eleocharis vivipara1. Plant Physiol. 2001, 127, 1524–1532. [Google Scholar] [CrossRef] [PubMed]
- Garner, D.M.; Mure, C.M.; Yerramsetty, P.; Berry, J.O. Kranz Anatomy and the C4 Pathway. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 1–10. [Google Scholar] [CrossRef]
- Eisenhut, M.; Brautigam, A.; Timm, S.; Florian, A.; Tohge, T.; Fernie, A.R.; Weber, A.P.M. Photorespiration is crucial for dynamic response of photosynthetic metabolism and stomatal movement to altered CO2 availability. Mol. Plant. 2017, 10, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, M.R.; Christin, P.A. Despite phylogenetic effects, C3–C4 lineages bridge the ecological gap to c4 photosynthesis. J. Exp. Bot. 2017, 68, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, O.; Kawano, Y.; Wakayama, M.; Takeda, T. Leaf vascular systems in C3 and C4 Grasses: A two-dimensional analysis. Ann. Bot. 2006, 97, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Cernusak, L.A.; Aranda, J.; Marshall, J.D.; Winter, K. Large variation in whole plant water-use efficiency among tropical tree species. New Physiol. 2007, 173, 294–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKown, A.D.; Dengler, N.G. Shifts in leaf vein density through accelerated vein formation in C4 Flaveria (Asteraceae). Ann. Bot. 2009, 104, 1085–1098. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, J.M.; Chillar, J.; Park, W.M. A process-based Boreal Ecosystem Productivity Simulator using remote sensing inputs. Remote Sens. Environ. 1997, 62, 158–175. [Google Scholar] [CrossRef]
- Mu, Q.; Zhao, M.; Running, S.W. Improvements to A MODIS global terrestrial evapotranspiration algorithm. Remote Sens. Environ. 2011, 115, 1781–1800. [Google Scholar] [CrossRef]
- Wei, Z.; Yoshimura, K.; Wang, L.; Miralles, D.G.; Jasechko, S.; Lee, X. Revisiting the contribution of transpiration to global terrestrial evapotranspiration. Geophys. Res. Lett. 2017, 44, 2792–2801. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Kimball, J.S.; Running, S.W. A review of remote sensing based actual evapotranspiration estimation. WIREs Water 2016, 3, 834–853. [Google Scholar] [CrossRef]
- NASA Earth Data. ECOSTRESS. Available online: https://search.earthdata.nasa.gov/search?q=ecostress&ac=true (accessed on 15 November 2020).
- American Rivers. Manual for the Lower Maumee and Ottawa River Watersheds; Tetra Tech. Low Impact Development Center: Washington, DC, USA, 2010. Available online: http://mvparkdistrict.org/pdf/nwo_lid_manual.pdf (accessed on 5 February 2022).
- U.S. Environmental Protection Agency. Action Plan for Lake Erie 2018. Available online: https://www.epa.gov/sites/default/files/2018-03/documents/us_dap_final_march_1.pdf (accessed on 20 January 2021).
- Natural Resources Conservation Service-NRCS. Lower Maumee Watershed. 2009. Available online: https://www.nrcs.usda.gov (accessed on 12 December 2020).
- National Centers for Environmental Information—NOAA. U.S. Climate Normals. Available online: https://www.ncei.noaa.gov/access/us-climate-normals (accessed on 19 February 2022).
- Beck, H.E.; Zimmerman, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- USDA. Crop Production. National Agriculture Statistics Service. Available online: https://www.nass.usda.gov/ (accessed on 18 December 2020).
- U.S. Geological Survey Earth Explorer. Available online: https://earthexplorer.usgs.gov (accessed on 20 December 2020).
- Abatzoglou, J.T. Development of gridded surface meteorological data for ecological applications and modelling. Int. J. Climatol. 2013, 33, 121–131. [Google Scholar] [CrossRef]
- Allen, R.G.; Irmak, A.; Trezza, R.; Hendrickx, J.M.H.; Bastiaanssen, W.; Kjaersgaard, J. Satellite-based ET estimation in agriculture using SEBAL and METRIC. Hydrol. Process. 2011, 25, 4011–4027. [Google Scholar] [CrossRef]
- PRISM Climate Group. Oregon State University. 2018. Available online: http://prism.oregonstate.edu (accessed on 2 February 2020).
- ESA Copernicus Open Access Hub. Available online: https://scihub.copernicus.eu (accessed on 15 May 2020).
- Schmitt, M.; Zhu, X.X. Data fusion and remote sensing: An ever-growing relationship. IEEE Geosci. Remote Sens. Mag. 2016, 4, 6–23. [Google Scholar] [CrossRef]
- Mandanici, E.; Bitelli, G. Preliminary comparison of sentinel-2 and Landsat 8 imagery for combined use. Remote Sens. 2016, 8, 1014. [Google Scholar] [CrossRef] [Green Version]
- Useya, J.; Chen, S. Comparative performance evaluation of pixel-level and decision level data fusion of Landsat 8 OLI, Landsat 7 ETM+, and sentinel-2 MSI for crop ensemble classification. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 11, 4441–4451. [Google Scholar] [CrossRef]
- USDA. CropScape—Cropland Data Layer. Available online: https://nassgeodata.gmu.edu/CropScape (accessed on 11 November 2019).
- Marambe, Y.; Simic Milas, A. Modeling Evapotranspiration for C4 and C3 Crops in the Western Lake Erie Basin Using Remote Sensing Data. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2020, 42, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Huete, A. A Soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Hong, S.Y.; Sudduth, K.A.; Kitchen, N.R.; Fraisse, C.W.; Palm, H.L.; Wiebold, W.J. Comparison of remote sensing and crop growth models for estimating within-field LAI variability. Korean J. Remote Sens. 2004, 20, 175–188. [Google Scholar]
- Kang, Y.; Özdoğan, M.; Zipper, S.C.; Román, M.O.; Walker, J.; Hong, S.Y.; Marshall, M.; Magliulo, V.; Moreno, J.; Alonso, L.; et al. How universal is the relationship between remotely sensed vegetation indices and crop leaf area index? A global assessment. Remote Sens. 2016, 8, 597. [Google Scholar] [CrossRef] [Green Version]
- Boegh, E.; Soegaard, H.; Broge, N.; Hasager, C.B.; Jensen, N.O.; Schelde, K.; Thomsen, A. Airborne multispectral data for quantifying leaf area index, nitrogen concentration, and photosynthetic efficiency in agriculture. Remote Sens. Environ. 2002, 81, 179–193. [Google Scholar] [CrossRef]
- Blinn, C.; House, M.; Wynne, R.; Thomas, V.; Fox, T.; Sumnall, M. Landsat 8 based leaf area index estimation in loblolly pine plantations. Forests 2019, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.J.; Cawse-Nicholson, K.; Barsi, J.; Radocinski, R.; Hulley, G.; Johnson, W.; Rivera, G.; Markham, B. In-flight validation of ECOSTRESS, Landsat 7 and 8 thermal infrared spectral channels using the lake Tahoe CA/NV and Salton sea CA automated validation sites. IEEE Trans. Geosci. Remote Sens. 2019, 58, 1294–1302. [Google Scholar] [CrossRef]
- Fisher, J.B.; Lee, B.; Purdy, A.J.; Halverson, G.H.; Dohlen, M.B.; Cawse-Nicholson, K.; Wang, A.; Anderson, R.G.; Aragon, B.; Arain, M.A.; et al. ECOSTRESS: NASA’s next generation mission to measure evapotranspiration from the international space station. Water Resour. Res. 2020, 56, e2019WR026058. [Google Scholar] [CrossRef]
- USDA. Description of SSURGO Database. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/survey/?cid=nrcs142p2_053627 (accessed on 18 December 2020).
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Corn Growth and Development; PMR (1009); Iowa State University Extension and Outreach: Ames, IA, USA, 2011; 50p. [Google Scholar]
- Simic, A.; Fernandes, R.; Wang, S. Assessing the impact of leaf area index on evapotranspiration and groundwater recharge across a shallow water region for diverse land cover and soil properties. J. Water Resour. Hydraul. Eng. 2004, 3, 60–73. [Google Scholar]
- Cui, T.; Sun, R.; Xiao, Z.; Liang, Z.; Wang, J. Simulating spatially distributed solar-induced chlorophyll fluorescence using a BEPS-SCOPE coupling framework. Agric. For. Meteorol. 2020, 295, 108169. [Google Scholar] [CrossRef]
- Norman, J.M. Simulation of Microclimates. In Biometeorology in Integrated Pest Management; Hatfield, J.L., Thomason, I.J., Eds.; Academic Press: New York, NY, USA, 1982; pp. 65–99. [Google Scholar]
- Rochette, P.; Pattey, E.; Desjardins, R.L.; Dwyer, L.M.; Stewart, D.W.; Dubé, P.A. Estimation of maize (Zea mays L.) canopy conductance by scaling up leaf stomatal conductance. Agric. For. Meteorol. 1991, 54, 241–261. [Google Scholar] [CrossRef]
- SWAT. SWAT Input Output File Information Version 2012. Available online: https://swat.tamu.edu/documentation/2012-io (accessed on 25 December 2020).
- Ocheltree, T.W.; Nippert, J.B.; Prasad, P.V.V. Changes in stomatal conductance along grass blades reflect changes in leaf structure. Plant Cell Environ. 2012, 35, 1040–1049. [Google Scholar] [CrossRef]
- Blonquist, J.M.; Norman, J.M.; Bugbee, B. Automated measurement of canopy stomatal conductance based on infrared temperature. Agric. For. Meteorol. 2009, 149, 1931–1945. [Google Scholar] [CrossRef]
- Renninger, H.; Carlo, N.; Clark, K.; Schafer, K. Resource use and efficiency, and stomatal responses to environmental drivers of oak and pine species in an Atlantic coastal plain forest. Front. Plant Sci. 2015, 6, 297. [Google Scholar] [CrossRef] [Green Version]
- Matzner, S.L.; Rice, K.J.; Richards, J.H. Patterns of stomatal conductance among blue oak (Quercus douglasii) size classes and populations: Implications for seedling establishment. Tree Physiol. 2003, 23, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Taconet, O.; Olioso, A.; Mehrez, M.B.; Brisson, N. Seasonal estimation of evaporation and stomatal conductance over a soybean field using surface IR temperatures. Agric. For. Meteorol. 1994, 73, 321–337. [Google Scholar] [CrossRef]
- NDSU. Corn Growth Management. 2020. Available online: https://www.ag.ndsu.edu/pubs/plantsci/crops/a1173.pdf (accessed on 15 August 2021).
- Berglund, D.R.; Endres, G.J.; McWilliams, D.A. Corn Growth and Management Quick Guide; A1173; NDSU Extension Service: Bismarck ND, USA, 1999; pp. 1–8. [Google Scholar]
- Wolf, D.D.; Blaser, R.E. Flexible alfalfa management: Early spring utilization. Crop Sci. 1991, 21, 90–93. [Google Scholar] [CrossRef]
- Iio, A.; Ito, A. A Global Database of Field-Observed Leaf Area Index in Woody Plant Species, 1932–2011; Oak Ridge National Laboratory Distributed Active Archive Center: Oak Ridge, TN, USA, 2014. Available online: https://daac.ornl.gov/cgibin/theme_dataset_lister.pl?theme_id=1 (accessed on 15 December 2021).
- Viña, A.; Gitelson, A.A.; Nguy-Robertson, A.L.; Peng, Y. Comparison of different vegetation indices for the remote assessment of green leaf area index of crops. Remote Sens. Environ. 2011, 115, 3468–3478. [Google Scholar] [CrossRef]
- Beard, J.B.; Green, R.L. The role of turf grasses in environmental protection and their benefits to humans. J. Environ. Qual. 1994, 23, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Hill, H.R. Using Evapotranspiration Data to Schedule Irrigation of Forages. Proceedings, Western Alfalfa and Forage Conference 2002. Available online: http://alfalfa.ucdavis.edu (accessed on 18 November 2021).
- Anda, A.; Simon, B.; Soos, G.; Teixeira da Silva, J.A.; Farkas, Z.; Menyhart, L. Assessment of soybean evapotranspiration and controlled water stress using traditional and converted evapotranspirometers. Atmosphere 2020, 11, 830. [Google Scholar] [CrossRef]
- Kimball, J.S.; Thornton, P.E.; White, M.A.; Running, S.W. Simulating Forest productivity and surface-atmosphere carbon exchange in the BOREAS study region. Tree Physiol. 1997, 17, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gai, Z.; Zhang, J.; Li, C. Effects of starter nitrogen fertilizer on soybean root activity, leaf photosynthesis and grain yield. PLoS ONE 2017, 12, e0174841. [Google Scholar] [CrossRef] [Green Version]
- Scagel, C.F.; Bi, G.; Fuchigami, L.H.; Regan, R.P. Effects of irrigation frequency and nitrogen fertilizer rate on water stress, nitrogen uptake, and plant growth of container-grown rhododendron. HortScience 2011, 46, 1598–1603. [Google Scholar] [CrossRef] [Green Version]
- Healy, R.W.; Winter, T.C.; la Baugh, J.W.; Franke, O.L. Water Budgets: Foundations for Effective Water Resources and Environmental Management; US Geological Survey: Reston, VI, USA, 2007; Volume 1308, p. 90. [Google Scholar]
- Jackson, R.B.; Esteban, G.J.; Avissar, R.; Somnath, B.R.; Barrett, D.J.; Cook, C.W.; Farley, K.A.; Maitre, D.C.L.; McCarl, B.A.; Murray, B.C. Trading water for carbon with biological carbon sequestration. Science 2005, 310, 1944–1947. [Google Scholar] [CrossRef] [Green Version]
- Levia, D.F.; Creed, I.F.; Hannah, D.M.; Nanko, K.; Boyer, E.W.; Carlyle-Moses, D.; Bruen, M. Homogenization of the terrestrial water cycle. Nat. Geosci. 2020, 13, 656–658. [Google Scholar] [CrossRef]
- Zeiger, E. The biology of stomatal guard cells. Annu. Rev. Plant Physiol. 1983, 34, 441–474. [Google Scholar] [CrossRef]
- Araújo, W.L.; Fernie, A.R.; Nunes-Nesi, A. Control of stomatal aperture: A renaissance of the old guard. Plant Signal. Behav. 2011, 6, 1305–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faralli, M.; Matthews, J.; Lawson, T. Exploiting natural variation and genetic manipulation of stomatal conductance for crop improvement. Curr. Opin. Plant Biol. 2019, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U.F.; McMillan, A.M.S. Warming and elevated CO2 have opposing influences on transpiration. Which is more important? Curr. For. Rep. 2018, 4, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Li, C.; Kong, D.; Guo, F.; Wei, H. Light-mediated signaling and metabolic changes coordinate stomatal opening and closure. Front. Plant Sci. 2020, 11, 601478. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, P.J.; Chang, J. Concerning the relationship between evapotranspiration and soil moisture. J. Appl. Meteorol. Climatol. 1987, 26, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Shaxson, F.; Barber, R. Optimizing Soil Moisture for Plant Production: The Significance of Soil Porosity; FAO Soils Bulletin 79; Food and Agriculture Organization of the United Nations: Rome, Italy, 1993. [Google Scholar]
- Casson, S.A.; Hetherington, A.M. Environmental regulation of stomatal development. Curr. Opin. Plant Biol. 2010, 13, 90–95. [Google Scholar] [CrossRef]
- He, L.; Chen, J.M.; Liu, J.; Bélair, S.; Luo, X. Assessment of SMAP soil moisture for global simulation of gross primary production. J. Geophys. Res. Biogeosci. 2017, 122, 1549–1563. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, J.M.; Qin, Q.; Huang, M.; Wang, L.; Li, J.; Cao, B. Assimilating Remote Sensing based Soil Moisture in an Ecosystem Model (BEPS) for Agricultural Drought Assessment. In Proceedings of the IGARSS 2008—2008 IEEE International Geoscience and Remote Sensing Symposium, Boston, MA, USA, 7–11 July 2008; Volume 5, p. 440. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Day | Sensor Type | Cloud Cover |
|---|---|---|---|
| May | 8 | Landsat 8 OLI | 0 |
| 24 | Landsat 8 OLI | 0 | |
| July | 11 | Landsat 8 OLI | 0 |
| 12 | Landsat 8 OLI | ~15% | |
| August | 11 | Sentinel-2A MSI | 0 |
| 23 | Sentinel-2B MSI | 0 | |
| 28 | Landsat 8 OLI | ~15% | |
| September | 27 | Landsat 8 OLI | 0 |
| Vegetation Indices | LAI Algorithm |
|---|---|
| Simple Ratio | |
| Enhanced Vegetation Index | |
| Enhanced Vegetation Index 2 | |
| Soil-Adjusted Vegetation Index |
| LULC | LAI Algorithm | References | Months |
|---|---|---|---|
| Grass | LAI = (a × (EVI)2 + b)4/3; a = 2.84, b = 0.88 | [56] | May–September |
| Winter wheat/cover crops/Alfalfa | LAI = 3.618 × EVI − 0.118 | [57] | May–September |
| Soybean | LAI = 6.59 SAVI − 2.34 | [55] | May/September |
| LAI = (a × EVI + b)2; a = 2.53, b = 0.69 | [56] | June/July/August | |
| Corn | LAI = 2.62 × SAVI − 0.1314 | [55] | May/September |
| LAI = (a × EVI + b)2; a = 2.42, b = 0.34 | [56] | June/July/August | |
| Conifer | LAI = 0.332915 × SR − 0.00212 | [58] | May–September |
| Crop Type | Scenario 1 (High SC (ms−1)) | Scenario 2 (Medium SC (ms−1)) | Scenario 3 (Low SC (ms−1)) |
|---|---|---|---|
| Grass | 0.0550 [67] | 0.0054 [68] | 0.0012 [68] |
| Alfalfa | 0.0100 [67] | 0.0054 [33] | 0.0012 [69] |
| Deciduous | 0.0064 [70] | 0.0050 [33] | 0.0043 [71] |
| Soybean | 0.0076 [72] | 0.0071 [67] | 0.0025 [72] |
| Corn | 0.0090 [66] | 0.0071 [67] | 0.0040 [66] |
| Month | May | June | July | August | September | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Crop | LAI | sd | LAI | sd | LAI | sd | LAI | sd | LAI | sd | |
| Grass | 1.36 | 0.32 | 1.83 | 0.37 | 1.94 | 0.42 | 1.80 | 0.37 | 1.59 | 0.27 | |
| Alfalfa | 1.21 | 0.62 | 2.23 | 0.60 | 2.24 | 0.69 | 2.16 | 0.60 | 2.03 | 0.58 | |
| Deciduous | 2.21 | 0.58 | 4.82 | 0.67 | 5.41 | 0.68 | 5.34 | 0.71 | 4.60 | 0.58 | |
| Soybean | 0.02 | 0.33 | 1.39 | 0.87 | 3.17 | 0.44 | 4.25 | 0.64 | 1.79 | 0.67 | |
| Corn | 0.30 | 0.22 | 3.16 | 0.79 | 5.04 | 0.29 | 4.63 | 0.78 | 1.99 | 0.60 | |
| Winter wheat/cover crops | 2.36 | 0.49 | 2.12 | 0.28 | 1.54 | 0.42 | 1.75 | 0.73 | 1.55 | 0.77 | |
| Crop | Estimated Mean LAI | LAI Values from Literature | References |
|---|---|---|---|
| Grass | 1.70 | 1.60–2.00 | [75] |
| Alfalfa | 1.99 | 1.50–2.10 | [75] |
| Deciduous | 4.48 | 5.60–6.70 | [76] |
| Soybean | 4.25 | 0.20–5.60 | [77] |
| Corn | 5.04 | 0.00–6.40 | [77] |
| LULC | BEPS-High SC | BEPS-Middle SC | BEPS-Low SC | ECOSTRESS | ET Rates from Literature | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ET (mm day−1) | sd | ET (mm day−1) | sd | ET (mm day−1) | sd | ET (mm day−1) | sd | ET (mm day−1) | Reference | |
| Grass | 4.09 | 1.61 | 4.03 | 1.61 | 2.50 | 1.27 | 4.85 | 1.13 | 3.00–8.00 | [78] |
| Alfalfa | 4.83 | 1.34 | 4.30 | 1.26 | 2.76 | 1.11 | 5.43 | 1.20 | 3.80–7.50 | [79] |
| Deciduous | 4.68 | 1.38 | 4.26 | 1.40 | 3.67 | 1.42 | 5.09 | 1.15 | ||
| Soybean | 5.20 | 1.16 | 5.10 | 1.14 | 3.22 | 0.76 | 5.13 | 1.14 | 2.54–5.30 | [80] |
| Corn | 5.20 | 1.10 | 4.43 | 0.98 | 3.19 | 0.73 | 5.13 | 1.12 | 5.08 | [81] |
| Winter wheat/cover crops | 3.47 | 1.18 | 3.89 | 1.08 | 3.46 | 0.99 | 4.83 | 1.29 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senevirathne, C.K.; Simic Milas, A.; Liu, G.; Yacobucci, M.M.; Marambe, Y.A. Effects of Increasing C4-Crop Cover and Stomatal Conductance on Evapotranspiration: Simulations for a Lake Erie Watershed. Remote Sens. 2022, 14, 1914. https://doi.org/10.3390/rs14081914
Senevirathne CK, Simic Milas A, Liu G, Yacobucci MM, Marambe YA. Effects of Increasing C4-Crop Cover and Stomatal Conductance on Evapotranspiration: Simulations for a Lake Erie Watershed. Remote Sensing. 2022; 14(8):1914. https://doi.org/10.3390/rs14081914
Chicago/Turabian StyleSenevirathne, Chathuranga Kumara, Anita Simic Milas, Ganming Liu, Margaret Mary Yacobucci, and Yahampath Anuruddha Marambe. 2022. "Effects of Increasing C4-Crop Cover and Stomatal Conductance on Evapotranspiration: Simulations for a Lake Erie Watershed" Remote Sensing 14, no. 8: 1914. https://doi.org/10.3390/rs14081914