
How Many Reindeer? UAV Surveys as an Alternative to Helicopter or Ground Surveys for Estimating Population Abundance in Open Landscapes

 ,
,  , , , ,
, , , ,
Abstract
1. Introduction
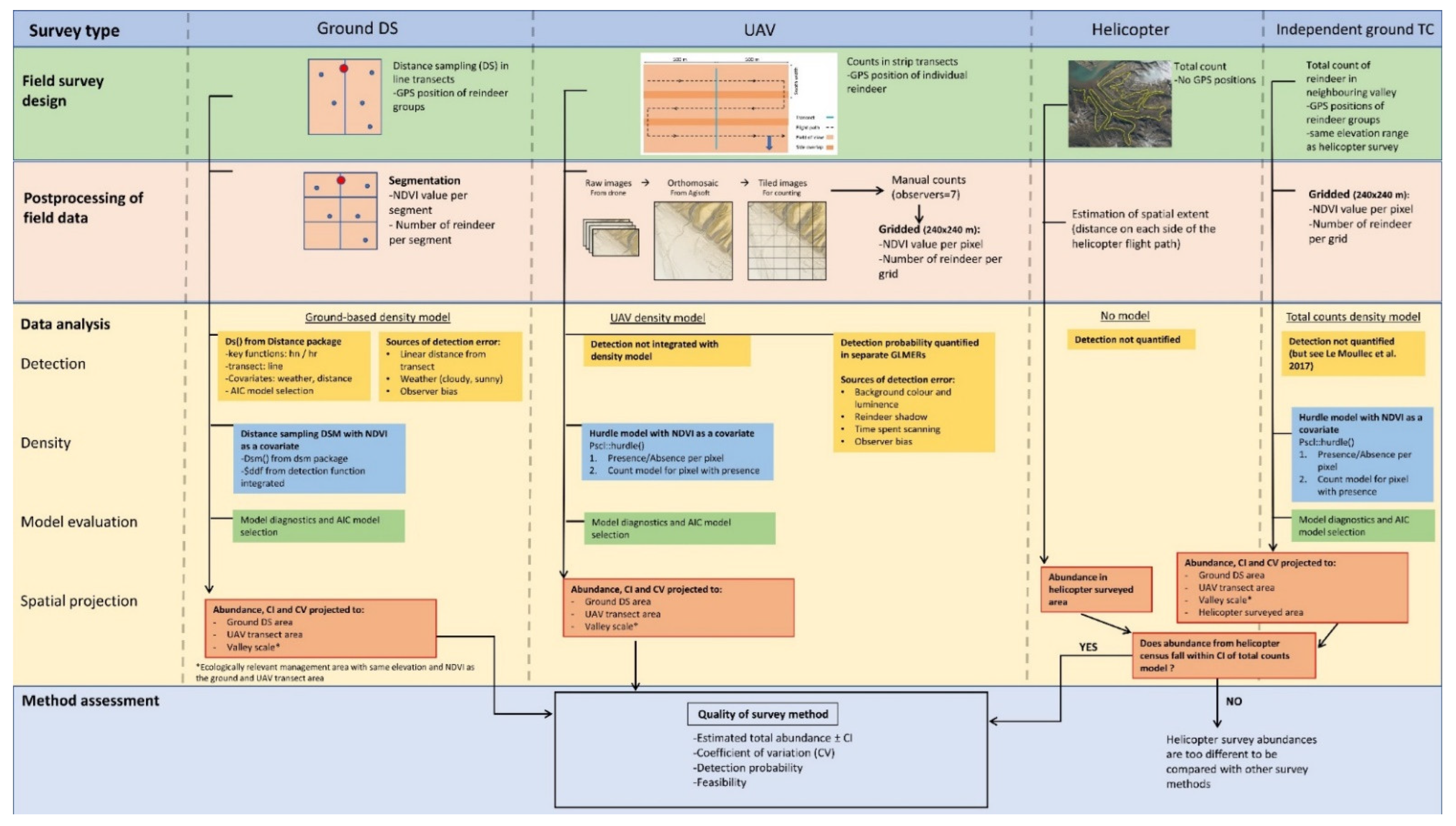
2. Materials and Methods
2.1. Study Area and Species
2.2. Field Data Collection
2.2.1. Ground Distance Sampling Survey
2.2.2. UAV Survey
2.2.3. Helicopter Survey
2.2.4. Independent Total Counts Survey
2.3. Data Analyses
2.3.1. Ground Distance Sampling DSM
2.3.2. UAV DSM
2.3.3. Independent Total Counts DSM
2.4. Comparison of Survey Methods
3. Results
3.1. Field Survey Characteristics
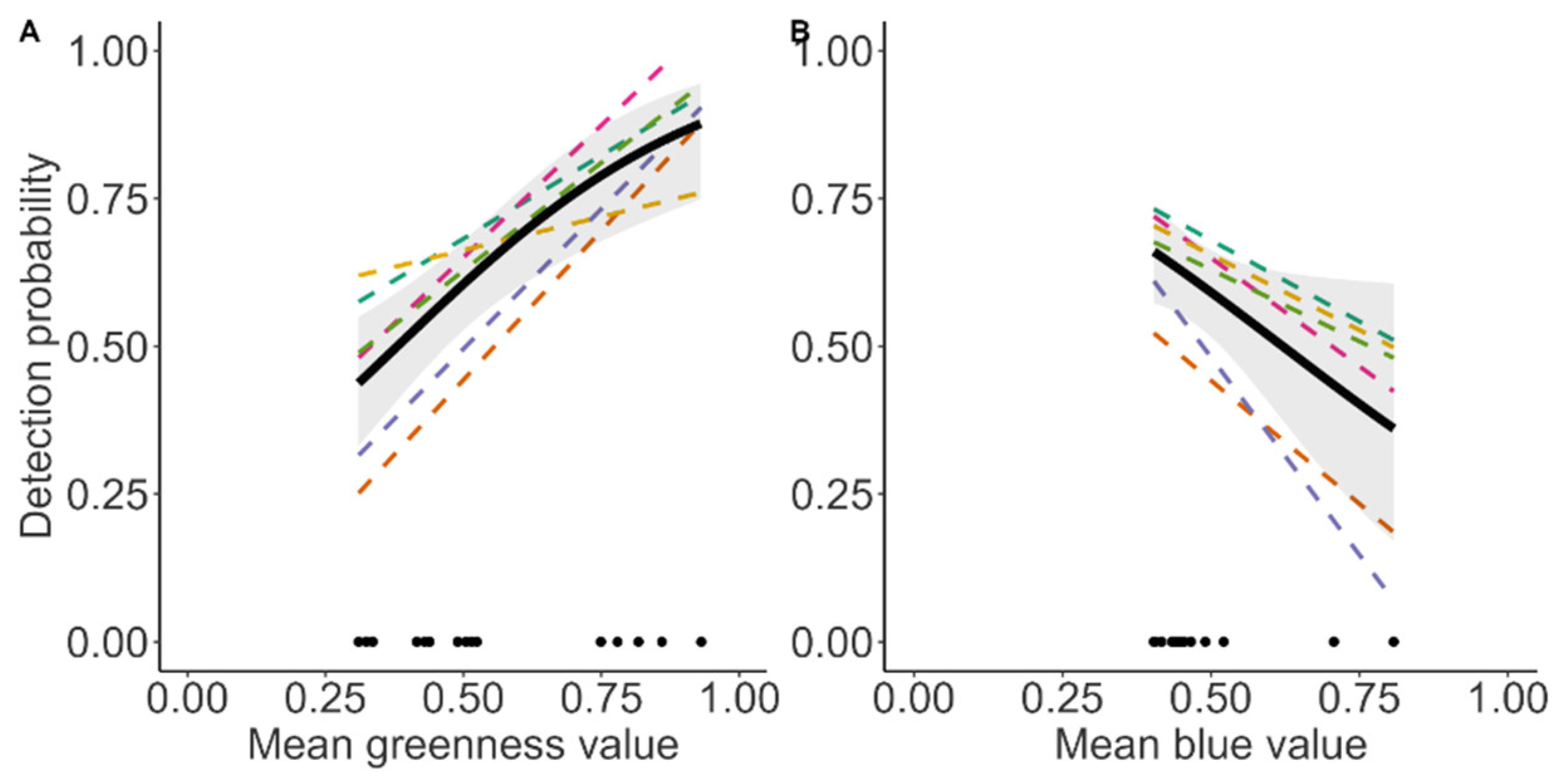
3.2. Detection of Reindeer
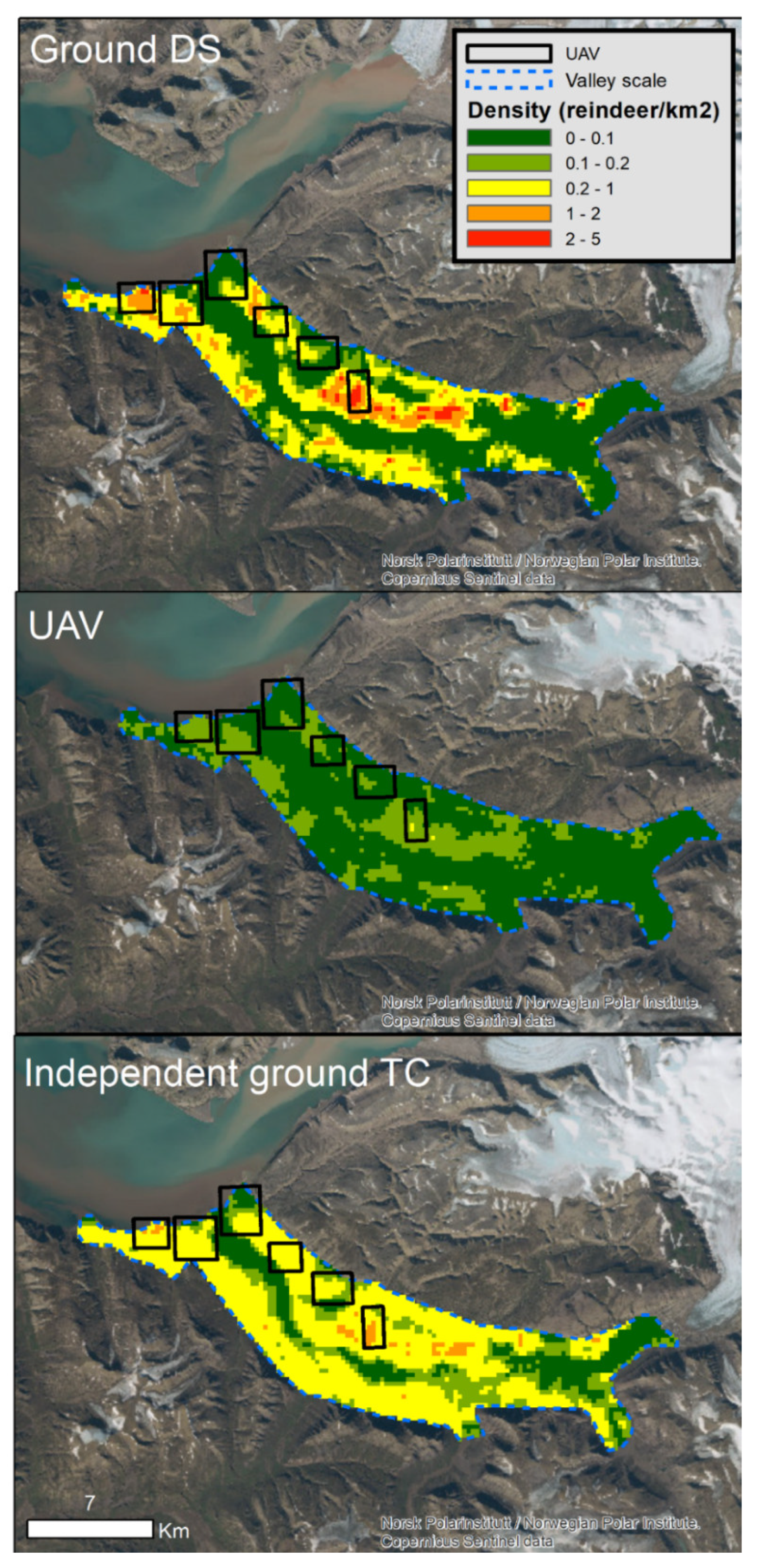
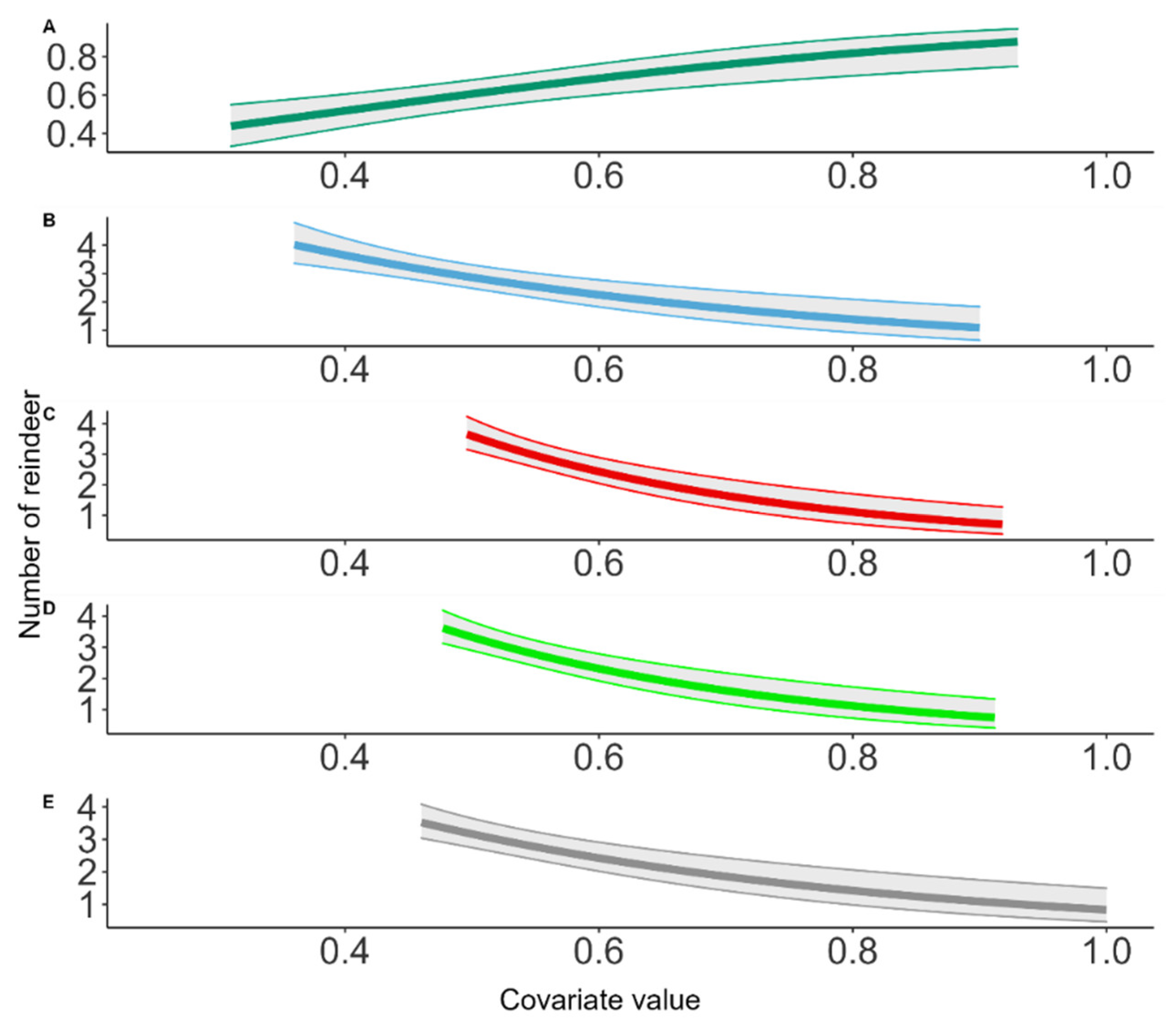
3.3. Reindeer Densities and Spatial Projections
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Independent Total Counts from Adventdalen—Background Data and Results
Statistical Analyses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hurdle Density Model Adventdalen | |||
|---|---|---|---|
| β ± SE | p | ||
| Count model | Intercept | −1.04 ± 0.57 | 0.07 |
| NDVI * | 0.003 ± 0.0009 | 0.002 | |
| P/A model | Intercept | −4.59 ± 0.33 | <0.05 |
| NDVI * | 0.004 ± 0.0005 | 0.02 | |
Appendix B. Model Selection and Detection Curve for Estimating Svalbard Reindeer Abundance by Ground Distance Sampling
| Model | Key | AIC | ΔAIC |
|---|---|---|---|
| ~weather | hr | 657.893 | 0 |
| ~1 | hr | 661.477 | 3.584 |
| ~weather | hn | 663.627 | 5.734 |
| ~1 | hn | 665.857 | 7.964 |

| Ground DS Survey Sassendalen | Model by Le Moullec et al. [22] | |||
|---|---|---|---|---|
| β ± SE | p | β ± SE | p | |
| Intercept | −19.25 ± 2.05 | <0.001 | −13.95 ± 0.38 | <0.001 |
| NDVI * | 0.012 ± 0.003 | <0.001 | 2.65 × 10−3 ± 0.76 × 10−3 | <0.001 |

Appendix C. Protocol for Counting Reindeer from UAV Imagery
Appendix C.1. Counting Svalbard Reindeer from Drone Imagery—Instructions to Observers (Full Version Can Be Available from the Authors upon Request)
Background



Appendix C.2. Download Software and Get Started!
- ▪
- Save and extract the ‘Reindeer_counting_drone_imagery.zip’ to your computer or hard disk. The folder and metadata require about 4 GB of space so make sure you have enough.
Appendix C.3. Set up DotDotGoose Software
- ▪
- Click and open the dotdotgoose.exe file in the ‘Reindeer_counting_drone_imagery’ folder
- ▪
- Click on ‘Load’ in the bottom left corner. Find the imagery folder “drone_imagery_SAS_2021” and select the point file ‘template_reindeer_counting.pnt’
- ▪
- In Survey Id at the top left panel: put your first name and last name with underscore, e.g., ole_olesen. This will create a column in the metadata with your name.
- ▪
- Click the Save button and save a point file with your own name (e.g., ole_olesen.pnt) into the same folder as the drone imagery ‘drone_imagery_SAS_2021’. It is important that it is the same folder as the imagery—if not the save will not work!
- ▪
- If you need to close the programme and finish at another time, you can open your point file in the DotDotGoose software by locating the file and click Import.
Appendix C.4. Reindeer Detection and Assigning Objects to Categories
Appendix C.4.1. Time Tracking
- ▪
- We would like to know how long it takes for each observer to scan through each transect line. The name of each jpeg file starts with the transect number (e.g., Line_1, Line_2).
- ▪
- When you are about to start on the first image of the transect (e.g., Line_1_tile_100.jpeg) write down the time in ‘time_start’ from the Custom Fields (right side panel) from your computer clock (e.g., 09:54).
- ▪
- When you have scanned all images in the transect (e.g., last image is Line_1_tile_99.jpeg) write down the time in time_stop (e.g., 11:00) on this last image of Line_1.
- ▪
- Do this for every transect line (Line_1 to Line_6) so we get the start and end time for each transect. Please try to complete every transect line in one go, but if you need to take breaks write down the end time and start time as well so breaks can be subtracted.
- ▪
- Remember to save frequently and when you take breaks.
Appendix C.4.2. Reindeer Scanning Method
- ▪
- For each image, scan the full-scale image quickly from grid to grid with your eyes (see example below). It might be useful to move your mouse as a guide.
- ▪
- If you cannot find an object of interest, go to the next image by pressing the down arrow key on your keyboard.
- ▪
- If you want to go back to any previous images use the up-arrow key or double-click on a specific photo in the Summary table.

- ▪
- If you do find an object of interest, zoom in on it to check if it is a reindeer or carcass by scrolling your mouse or use the zoom buttons in the right bottom corner (you can also drag the image up, down, and sideways by clicking and holding the mouse).
- ▪
- To mark a reindeer or carcass, you click on the category you want to assign on the left side panel (see left image below). Press the Ctrl key while you click on the object in the image. A dot will be created over the reindeer.
- ▪
- You can double check that the right category was assigned to the object for that image by looking at the Summary table on the left panel (see right image below).
- ▪
- NB! If you accidentally make a point or assign wrong category and need to remove it from the image, press and hold the Shift key on your keyboard, then left click and drag the mouse to draw a box around the points you’d like to delete. A red circle around your point will show up. Press the Delete key to remove the point.
Appendix D. UAV Density Model for Estimating Reindeer Abundance with Hurdle Density Model
| UAV Density Models | ||||||
|---|---|---|---|---|---|---|
| β ± SE | p | AIC | ΔAIC | |||
| Hurdle 1 | Count model | Intercept | −2.61 ± 82.8 | 0.975 | 182.50 | 0 |
| NDVI | −9.59 ± 7.15 | 0.180 | ||||
| P/A model | Intercept | −7.00 ± 1.40 | <0.05 | |||
| NDVI | 6.67 ± 2.15 | 0.002 | ||||
| Hurdle 2 | Count model | Intercept | 114.19 ± −0.08 | 0.93 | 182.54 | 0.04 |
| Log(theta) | −10.16 ± 114.20 | 0.93 | ||||
| P/A model | Intercept | −5.82 ± 1.36 | <0.05 | |||
| NDVI | 5.19 ± 2.080 | 0.012 | ||||
Appendix E. Detection Probability from UAV Imagery
- Binomial linear mixed effect model (GLMER)
- Five separate models with observer id as a random effect and each of the fixed effects median luminance, mean red, green, and blue channels per image. We only show the predicted effect plots for the fixed effects with a statistical significance (p > 0.05) below (intercepts and standard error in results section).
- Response variable: Reindeer seen (1) or reindeer not seen (0) by observers
- Sample size of reindeer n = 234
- Poisson GLMER.
- Five models with observer id as a random effect and each of the fixed effects median luminance, mean red, green, and blue channels per image (intercepts and standard error in results section). Below, we only show the predicted effect plots for the fixed effects with a statistical significance (p > 0.05)
- Response variable: Number of reindeer observed in an image
- Sample size of reindeer n = 179

| Fixed Effect | Random Effect | Coefficient | Fixed Effect (Β ± Se) | Random Effect | AIC | |
|---|---|---|---|---|---|---|
| P/A model | ~Greenness index | observer ID | Intercept Covariate | −1.36 ± 0.48 3.57 ± 0.92 | 0.04, 0.20 | 301.89 |
| ~mean blue channel | observer ID | Intercept Covariate | 1.90 ± 0.73 −3.06 ± 1.48 | 0.03, 0.17 | 315.10 | |
| ~mean green channel | observer ID | Intercept Covariate | 1.63 ± 0.85 −2.25 ± 1.58 | 0.03, 0.17 | 317.43 | |
| ~mean red channel | observer ID | Intercept Covariate | 1.35 ± 0.89 −1.67 ± 1.61 | 0.03, 0.16 | 318.40 | |
| ~median luminance | observer ID | Intercept Covariate | 1.07 ± 0.68 −1.22 ± 1.28 | 0.03, 0.16 | 318.57 | |
| Count model | ~mean red channel | observer ID | Intercept Covariate | −3.91 ± 0.76 −3.91 ± 0.76 | 0.02, 0.13 | 709.63 |
| ~mean green channel | observer ID | Intercept Covariate | 3.01 ± 0.38 −3.62 ± 0.72 | 0.02, 0.13 | 712.19 | |
| ~median luminance | observer ID | Intercept Covariate | −2.66 ± 0.57 −2.66 ± 0.57 | 0.02, 0.14 | 714.17 | |
| ~mean blue channel | observer ID | Intercept Covariate | −2.41 ± 0.58 −2.41 ± 0.58 | 0.02, 0.13 | 725.01 | |
| ~Greenness index | observer ID | Intercept covariate | 1.40 ± 0.14 −0.52 ± 0.22 | 0.03, 0.18 | 740.84 |
References
- Nichols, J.D.; Williams, B.K. Monitoring for conservation. Trends Ecol. Evol. 2006, 21, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.K.; Nichols, J.D.; Conory, M.J. Analysis and Management of Wildlife Populations; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Forsyth, D.M.; Comte, S.; Davis, N.E.; Bengsen, A.J.; Cote, S.D.; Hewitt, D.G.; Morellet, N.; Mysterud, A. Methodology matters when estimating deer abundance: A global systematic review and recommendations for improvements. J. Wildl. Manag. 2022, 86, e22207. [Google Scholar] [CrossRef]
- Thompson, W.L.; White, G.C.; Gowan, C. Chapter 3—Enumeration Methods. In Monitoring Vertebrate Populations; Thompson, W.L., White, G.C., Gowan, C., Eds.; Academic Press: San Diego, CA, USA, 1998; pp. 75–121. [Google Scholar]
- Gamelon, M.; Firth, J.A.; le Moullec, M.; Petry, W.K.; Salguero-Gòmez, R. Longitudinal demographic data collection. In Demographic Methods Across the Tree of Life; Roberto Salguero-Gomez, M.G., Ed.; Oxford Academic: Oxford, UK, 2021. [Google Scholar]
- ENETWILD consortium; Grignolio, S.; Apollonio, M.; Brivio, F.; Vicente, J.; Acevedo, P.; Palencia, P.; Petrovic, K.; Keuling, O. Guidance on estimation of abundance and density data of wild ruminant population: Methods, challenges, possibilities. EFSA Support. Publ. 2020, 17, 1876E. [Google Scholar] [CrossRef]
- Wang, D.L.; Shao, Q.Q.; Yue, H.Y. Surveying Wild Animals from Satellites, Manned Aircraft and Unmanned Aerial Systems (UASs): A Review. Remote Sens. 2019, 11, 1308. [Google Scholar] [CrossRef]
- Pereira, J.A.; Varela, D.; Scarpa, L.J.; Frutos, A.E.; Fracassi, N.G.; Lartigau, B.V.; Pina, C.I. Unmanned aerial vehicle surveys reveal unexpectedly high density of a threatened deer in a plantation forestry landscape. Oryx 2022, First View, 1–9. [Google Scholar] [CrossRef]
- Schofield, G.; Esteban, N.; Katselidis, K.A.; Hays, G.C. Drones for research on sea turtles and other marine vertebrates—A review. Biol. Conserv. 2019, 238, 108214. [Google Scholar] [CrossRef]
- Fettermann, T.; Fiori, L.; Gillman, L.; Stockin, K.A.; Bollard, B. Drone surveys are more accurate than boat-based surveys of bottlenose dolphins (Tursiops truncatus). Drones 2022, 6, 82. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Raja Segaran, R.; Reid, I.; Terauds, A.; Koh, L.P. Drones count wildlife more accurately and precisely than humans. Methods Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef]
- Forsyth, D.M.; MacKenzie, D.I.; Wright, E.F. Monitoring ungulates in steep non-forest habitat: A comparison of faecal pellet and helicopter counts. N. Z. J. Zool. 2014, 41, 248–262. [Google Scholar] [CrossRef]
- Noyes, J.H.; Johnson, B.K.; Riggs, R.A.; Schlegel, M.W.; Coggins, V.L. Assessing aerial survey methods to estimate elk populations: A case study. Wildl. Soc. Bull. 2000, 28, 636–642. [Google Scholar]
- Poole, K.G.; Cuyler, C.; Nymand, J. Evaluation of caribou Rangifer tarandus groenlandicus survey methodology in West Greenland. Wildl. Biol. 2013, 19, 225–239. [Google Scholar] [CrossRef]
- Davis, K.L.; Silverman, E.D.; Sussman, A.L.; Wilson, R.R.; Zipkin, E.F. Errors in aerial survey count data: Identifying pitfalls and solutions. Ecol. Evol. 2022, 12, e8733. [Google Scholar] [CrossRef] [PubMed]
- Reilly, B.K.; van Hensbergen, H.J.; Eiselen, R.J.; Fleming, P.J.S. Statistical power of replicated helicopter surveys in southern African conservation areas. Afr. J. Ecol. 2017, 55, 198–210. [Google Scholar] [CrossRef]
- Dyal, J.; Miller, K.V.; Cherry, M.J.; D’Angelo, G.J. Estimating sightability for helicopter surveys using surrogates of white-tailed deer. J. Wildl. Manag. 2021, 85, 887–896. [Google Scholar] [CrossRef]
- Mansson, J.; Hauser, C.E.; Andren, H.; Possingham, H.P. Survey method choice for wildlife management: The case of moose Alces alces in Sweden. Wildl. Biol. 2011, 17, 176–190. [Google Scholar] [CrossRef]
- Gentle, M.; Finch, N.; Speed, J.; Pople, A. A comparison of unmanned aerial vehicles (drones) and manned helicopters for monitoring macropod populations. J. Wildl. Res. 2018, 45, 586–594. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001; pp. 1–432. [Google Scholar]
- Le Moullec, M.; Pedersen, A.O.; Yoccoz, N.G.; Aanes, R.; Tufto, J.; Hansen, B.B. Ungulate population monitoring in an open tundra landscape: Distance sampling versus total counts. Wildl. Biol. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Le Moullec, M.; Pedersen, Å.Ø.; Stien, A.; Rosvold, J.; Hansen, B.B. A century of conservation: The ongoing recovery of svalbard reindeer. J. Wildl. Manag. 2019, 83, 1676–1686. [Google Scholar] [CrossRef]
- Delplanque, A.; Foucher, S.; Lejeune, P.; Linchant, J.; Théau, J. Multispecies detection and identification of African mammals in aerial imagery using convolutional neural networks. Remote Sens. Ecol. Conserv. 2022, 8, 166–179. [Google Scholar] [CrossRef]
- Yang, F.; Shao, Q.; Jiang, Z. A population census of large herbivores based on UAV and its effects on grazing pressure in the Yellow-River-Source National Park, China. Int. J. Environ. Res. Public Health 2019, 16, 4402. [Google Scholar] [CrossRef]
- Butcher, P.A.; Colefax, A.P.; Gorkin, R.A.; Kajiura, S.M.; López, N.A.; Mourier, J.; Purcell, C.R.; Skomal, G.B.; Tucker, J.P.; Walsh, A.J.; et al. The drone revolution of shark science: A review. Drones 2021, 5, 8. [Google Scholar] [CrossRef]
- Descamps, S.; Aars, J.; Fuglei, E.; Kovacs, K.M.; Lydersen, C.; Pavlova, O.; Pedersen, A.O.; Ravolainen, V.; Strom, H. Climate change impacts on wildlife in a High Arctic archipelago—Svalbard, Norway. Glob. Chang. Biol. 2017, 23, 490–502. [Google Scholar] [CrossRef]
- van der Wal, R.; Bardgett, R.D.; Harrison, K.A.; Stien, A. Vertebrate herbivores and ecosystem control: Cascading effects of faeces on tundra ecosystems. Ecography 2004, 27, 242–252. [Google Scholar] [CrossRef]
- Peeters, B.; Pedersen, Å.Ø.; Veiberg, V.; Hansen, B.B. Hunting quotas, selectivity and stochastic population dynamics challenge the management of wild reindeer. Clim. Res. 2022, 86, 93–111. [Google Scholar] [CrossRef]
- Hansen, B.B.; Gamelon, M.; Albon, S.D.; Lee, A.M.; Stien, A.; Irvine, R.J.; Saether, B.E.; Loe, L.E.; Ropstad, E.; Veiberg, V.; et al. More frequent extreme climate events stabilize reindeer population dynamics. Nat. Commun. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Loe, L.E.; Liston, G.E.; Pigeon, G.; Barker, K.; Horvitz, N.; Stien, A.; Forchhammer, M.; Getz, W.M.; Irvine, R.J.; Lee, A.; et al. The neglected season: Warmer autumns counteract harsher winters and promote population growth in Arctic reindeer. Glob. Chang. Biol. 2021, 27, 993–1002. [Google Scholar] [CrossRef]
- Solberg, E.J.; STrand, O.; Veiberg, V.; Andersen, R.; Heim, M.; Rolandsen, C.M.; Langvatn, R.; Holmstrøm, F.; Solem, M.I.; Eriksen, R.; et al. Hjortevilt 1991–2011. Oppsummeringsrapport fra Overvåkingsprogrammet for hjortevilt. NINA Rapp. 2012, 885, 1–156. [Google Scholar]
- Governor of Svalbard. Plan for forvaltning av svalbardrein, kunnskaps- og forvaltningsstatus. Rapport 2009, 1, 2009. [Google Scholar]
- Hansen, B.B.; Pedersen, A.O.; Peeters, B.; Le Moullec, M.; Albon, S.D.; Herfindal, I.; Saether, B.E.; Grotan, V.; Aanes, R. Spatial heterogeneity in climate change effects decouples the long-term dynamics of wild reindeer populations in the high Arctic. Glob. Chang. Biol. 2019, 25, 3656–3668. [Google Scholar] [CrossRef]
- Albon, S.D.; Irvine, R.J.; Halvorsen, O.; Langvatn, R.; Loe, L.E.; Ropstad, E.; Veiberg, V.; Van Der Wal, R.; Bjørkvoll, E.M.; Duff, E.I.; et al. Contrasting effects of summer and winter warming on body mass explain population dynamics in a food-limited Arctic herbivore. Glob. Chang. Biol. 2017, 23, 1374–1389. [Google Scholar] [CrossRef]
- Pedersen, Å.Ø.; Bårdsen, B.J.; Veiberg, V.; Hansen, B.B. Jegernes egne data. Analyser av jaktstatistikk og kjevemateriale fra svalbardrein. Nor. Polarinst. Kortrapport. 2014, 27, 1504–3215. [Google Scholar]
- Ims, R.A.; Jepsen, J.U.; Stien, A.; Yoccoz, N.G. Science Plan for COAT: Climate-Ecological Observatory for Arctic Tundra; Fram Centre: Tromsø, Norway, 2013. [Google Scholar]
- Hann, R.; Altstädter, B.; Betlem, P.; Deja, K.; Dragańska-Deja, K.; Ewertowski, M.; Hartvich, F.; Jonassen, M.; Lampert, A.; Laska, M.; et al. Scientific Applications of Unmanned Vehicles in Svalbard; SESS report 2020; Svalbard Integrated Arctic Earth Observing System: Longyearbyen, Norway, 2021. [Google Scholar] [CrossRef]
- Johansen, B.E.; Karlsen, S.R.; Tommervik, H. Vegetation mapping of Svalbard utilising Landsat TM/ETM plus data. Polar Rec. 2012, 48, 47–63. [Google Scholar] [CrossRef]
- Elvebakk, A. A vegetation map of Svalbard on the scale 1:3.5 mill. Phytocoenologia 2005, 35, 951–967. [Google Scholar] [CrossRef]
- Elvebakk, A. Bioclimatic delimitation and subdivision of the Arctic. In The Species Concept in the High North—A Panarctic Flora Initiative; Nordal, I., Razzhivin, V.Y., Eds.; Norske Videnskaps-Akademi: Oslo, Norway, 1999; pp. 81–112. [Google Scholar]
- Derocher, A.E.; Wiig, O.; Bangjord, G. Predation of Svalbard reindeer by polar bears. Polar Biol. 2000, 23, 675–678. [Google Scholar] [CrossRef]
- Stempniewicz, L.; Kulaszewicz, I.; Aars, J. Yes, they can: Polar bears Ursus maritimus successfully hunt Svalbard reindeer Rangifer tarandus platyrhynchus. Polar Biol. 2021, 44, 2199–2206. [Google Scholar] [CrossRef]
- Solberg, E.J.; Jordhøy, P.; Strand, O.; Aanes, R.; Loison, A.; Sæther, B.E.; Linnell, J.D.C. Effects of density-dependence and climate on the dynamics of a Svalbard reindeer population. Ecography 2001, 24, 441–451. [Google Scholar] [CrossRef]
- Stien, A.; Ims, R.A.; Albon, S.D.; Fuglei, E.; Irvine, R.J.; Ropstad, E.; Halvorsen, O.; Loe, L.E.; Veiberg, V.; Yoccoz, N.G. Congruent responses to weather variability in high arctic herbivores. Biol. Lett. 2012, 8, 1002–1005. [Google Scholar] [CrossRef]
- Marques, T.A.; Buckland, S.T.; Bispo, R.; Howland, B. Accounting for animal density gradients using independent information in distance sampling surveys. Stat. Methods Appl. 2013, 22, 67–80. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Karlsen, S.R.; Elvebakk, A.; Hogda, K.A.; Grydeland, T. Spatial and Temporal variability in the onset of the growing season on Svalbard, Arctic Norway—Measured by MODIS-NDVI Satellite Data. Remote Sens. 2014, 6, 8088–8106. [Google Scholar] [CrossRef]
- Karlsen, S.R.; Anderson, H.B.; van der Wal, R.; Hansen, B.B. A new NDVI measure that overcomes data sparsity in cloud-covered regions predicts annual variation in ground-based estimates of high arctic plant productivity. Environ. Res. Lett. 2018, 13, 12. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 June 2021).
- Miller, D.L.; Burt, M.L.; Rexstad, E.A.; Thomas, L. Spatial models for distance sampling data: Recent developments and future directions. Methods Ecol. Evol. 2013, 4, 1001–1010. [Google Scholar] [CrossRef]
- Kawashima, S.; Nakatani, M. An algorithm for estimating chlorophyll content in leaves using a Video Camera. Ann. Bot. 1998, 81, 49–54. [Google Scholar] [CrossRef]
- Liu, Y.; Hatou, K.; Aihara, T.; Kurose, S.; Akiyama, T.; Kohno, Y.; Lu, S.; Omasa, K. A Robust Vegetation Index Based on Different UAV RGB Images to Estimate SPAD Values of Naked Barley Leaves. Remote Sens. 2021, 13, 686. [Google Scholar] [CrossRef]
- Skalski, J.R.; Ryding, K.E.; Millspaugh, J.J. 9—Estimating Population Abundance. In Wildlife Demography; Skalski, J.R., Ryding, K.E., Millspaugh, J.J., Eds.; Academic Press: Burlington, ON, Canada, 2005; pp. 435–539. [Google Scholar]
- Witczuk, J.; Pagacz, S.; Zmarz, A.; Cypel, M. Exploring the feasibility of unmanned aerial vehicles and thermal imaging for ungulate surveys in forests—Preliminary results. Int. J. Remote Sens. 2018, 39, 5504–5521. [Google Scholar] [CrossRef]
- Brack, I.V.; Kindel, A.; Oliveira, L.F.B. Detection errors in wildlife abundance estimates from Unmanned Aerial Systems (UAS) surveys: Synthesis, solutions, and challenges. Methods Ecol. Evol. 2018, 9, 1864–1873. [Google Scholar] [CrossRef]
- Acevedo, P.; Ruiz-Fons, F.; Vicente, J.; Reyes-García, A.R.; Alzaga, V.; Gortázar, C. Estimating red deer abundance in a wide range of management situations in Mediterranean habitats. J. Zool. 2008, 276, 37–47. [Google Scholar] [CrossRef]
- Sun, C.; Beirne, C.; Burgar, J.M.; Howey, T.; Fisher, J.T.; Burton, A.C. Simultaneous monitoring of vegetation dynamics and wildlife activity with camera traps to assess habitat change. Remote Sens. Ecol. Conserv. 2021, 7, 666–684. [Google Scholar] [CrossRef]
- Dujon, A.M.; Ierodiaconou, D.; Geeson, J.J.; Arnould, J.P.Y.; Allan, B.M.; Katselidis, K.A.; Schofield, G. Machine learning to detect marine animals in UAV imagery: Effect of morphology, spacing, behaviour and habitat. Remote Sens. Ecol. Conserv. 2021, 7, 341–354. [Google Scholar] [CrossRef]
- Ersts, P.J. DotDotGoose (version 1.5.1). American Museum of Natural History, Center for Biodiversity and Conservation. Available online: https://biodiversityinformatics.amnh.org/open_source/dotdotgoose (accessed on 7 December 2022).





| Estimated Abundance | ||||
|---|---|---|---|---|
| Survey Method | UAV Sampling Area (16.2 km2) | Ground DS Sampling Area (42.7 km2) | Valley Scale (161.7 km2) | Helicopter Surveyed Area (286.2 km2) |
| Ground DS | 164 ± 43 (CV = 0.26) | 351 ± 84 (CV = 0.24) | 920 ± 202 (CV = 0.22) | - |
| UAV | 32 ± 9 (CV = 0.29) | 77 ± 15 (CV = 0.19) | 243 ± 26 (CV = 0.11) | - |
| Helicopter | - | - | - | 1559 * |
| Independent ground TC | 131 ± 32 (CV = 0.24) | 311 ± 48 (CV = 0.15) | 958 ± 82 (CV = 0.09) | 1515 ± 101 (CV = 0.07) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulsen, I.M.G.; Pedersen, Å.Ø.; Hann, R.; Blanchet, M.-A.; Eischeid, I.; van Hazendonk, C.; Ravolainen, V.T.; Stien, A.; Le Moullec, M. How Many Reindeer? UAV Surveys as an Alternative to Helicopter or Ground Surveys for Estimating Population Abundance in Open Landscapes. Remote Sens. 2023, 15, 9. https://doi.org/10.3390/rs15010009
Paulsen IMG, Pedersen ÅØ, Hann R, Blanchet M-A, Eischeid I, van Hazendonk C, Ravolainen VT, Stien A, Le Moullec M. How Many Reindeer? UAV Surveys as an Alternative to Helicopter or Ground Surveys for Estimating Population Abundance in Open Landscapes. Remote Sensing. 2023; 15(1):9. https://doi.org/10.3390/rs15010009
Chicago/Turabian StylePaulsen, Ingrid Marie Garfelt, Åshild Ønvik Pedersen, Richard Hann, Marie-Anne Blanchet, Isabell Eischeid, Charlotte van Hazendonk, Virve Tuulia Ravolainen, Audun Stien, and Mathilde Le Moullec. 2023. "How Many Reindeer? UAV Surveys as an Alternative to Helicopter or Ground Surveys for Estimating Population Abundance in Open Landscapes" Remote Sensing 15, no. 1: 9. https://doi.org/10.3390/rs15010009
APA StylePaulsen, I. M. G., Pedersen, Å. Ø., Hann, R., Blanchet, M.-A., Eischeid, I., van Hazendonk, C., Ravolainen, V. T., Stien, A., & Le Moullec, M. (2023). How Many Reindeer? UAV Surveys as an Alternative to Helicopter or Ground Surveys for Estimating Population Abundance in Open Landscapes. Remote Sensing, 15(1), 9. https://doi.org/10.3390/rs15010009