Quantifying Carbon Use Efficiency: Unraveling the Impact of Climate Change and Ecological Engineering on Vegetation in the Three Rivers Source Region

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Analysis Methods
2.2.1. Estimation of Carbon Use Efficiency
- CUE represents the carbon use efficiency of vegetation.
- NPP denotes the net primary productivity of vegetation, measured in g C/m2.
- GPP stands for the gross primary productivity of vegetation, quantified in g C/m2.
- Ra signifies the vegetation autotrophic respiration, expressed in g C/m2.
2.2.2. Sen’s Slope Estimator and Mann–Kendall Test
2.2.3. Pearson Correlation Analysis
2.2.4. Residual Analysis
3. Results
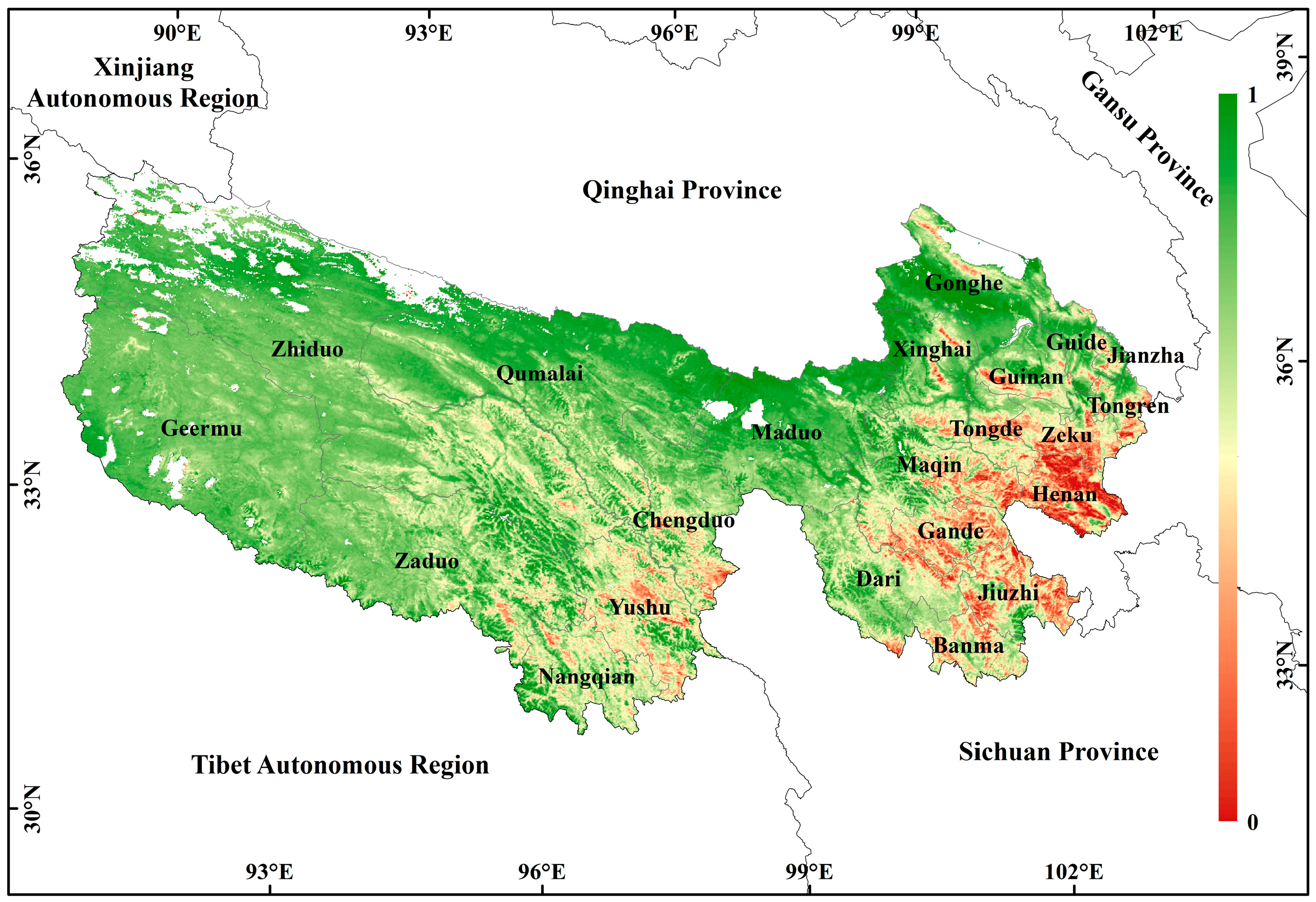
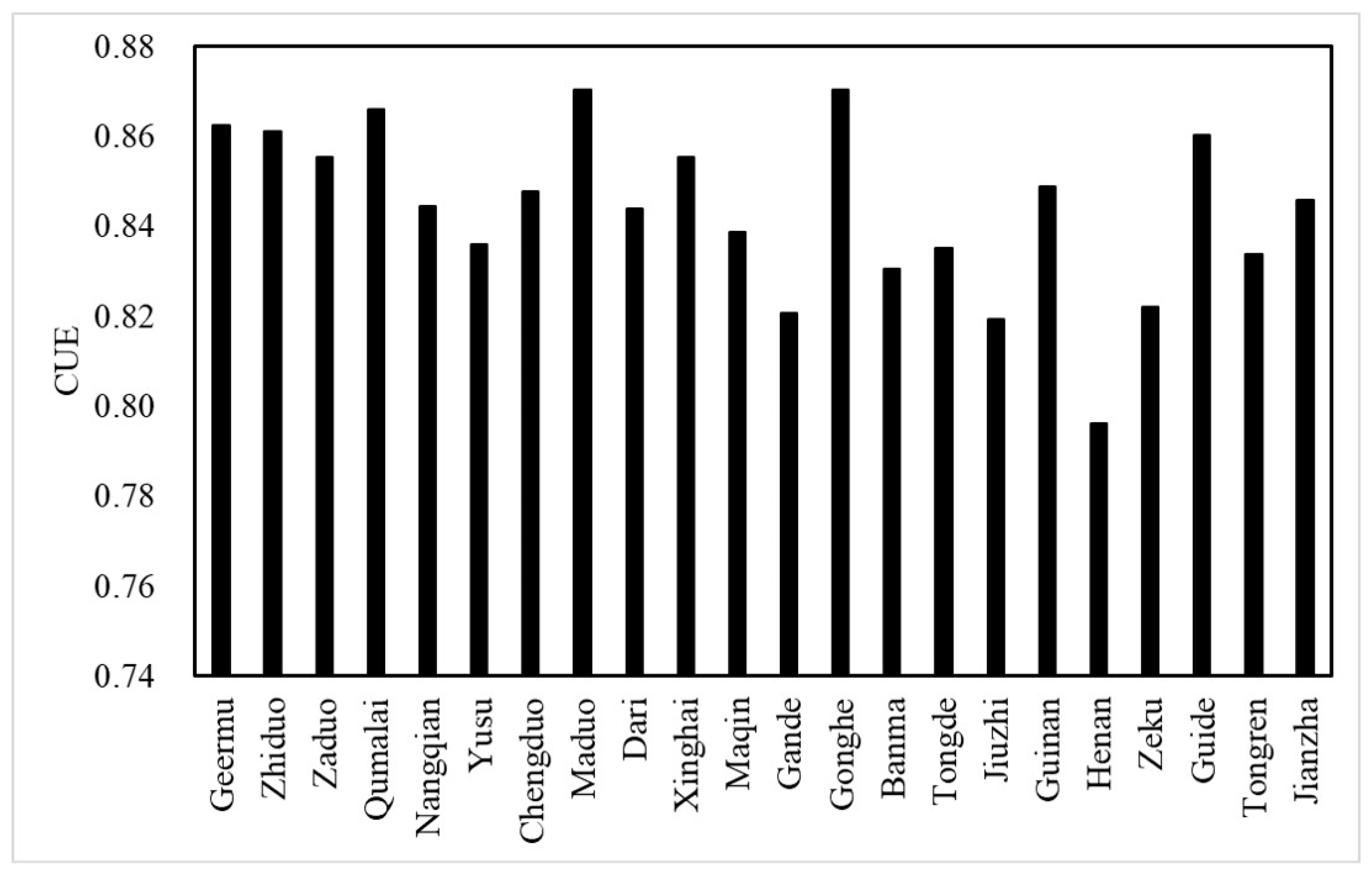
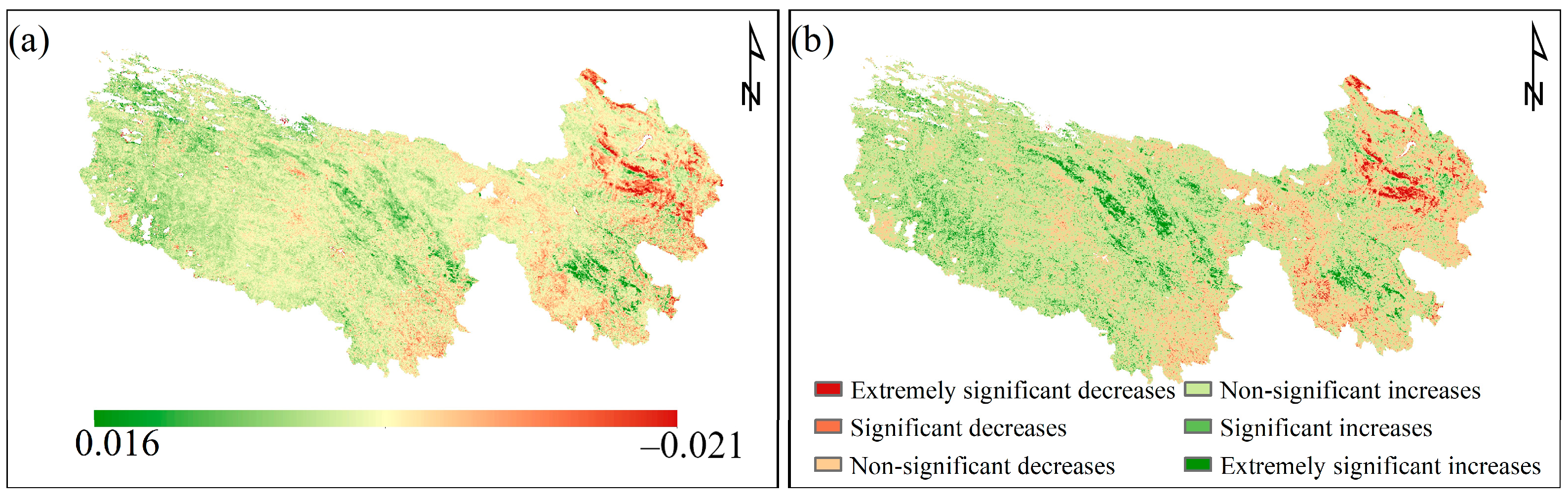
3.1. Spatial Distribution and Change of CUE
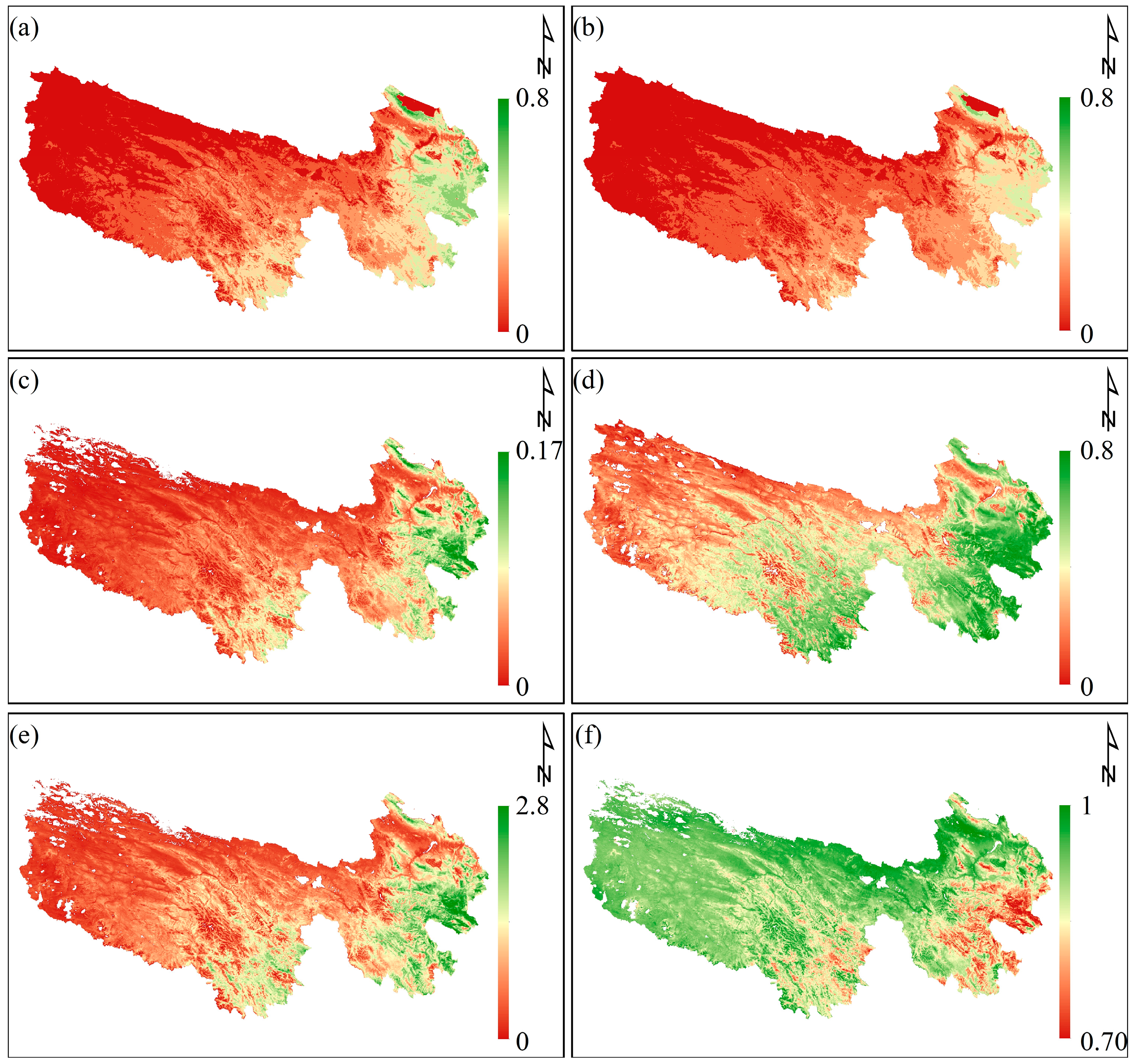
3.2. Carbon Allocation and Interannual Spatial Distribution Characteristics of Vegetation Parameters
3.3. Spatial Correlation Analysis of CUE and Climate Variability
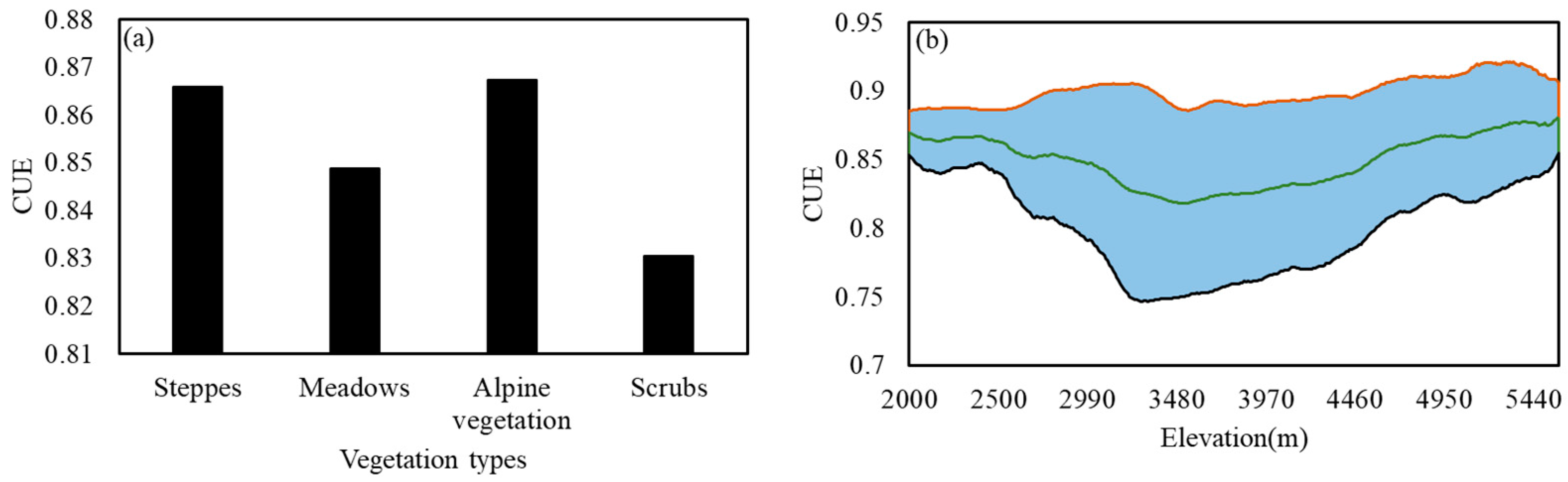
3.4. Impact of Vegetation Types and Elevation Changes on CUE
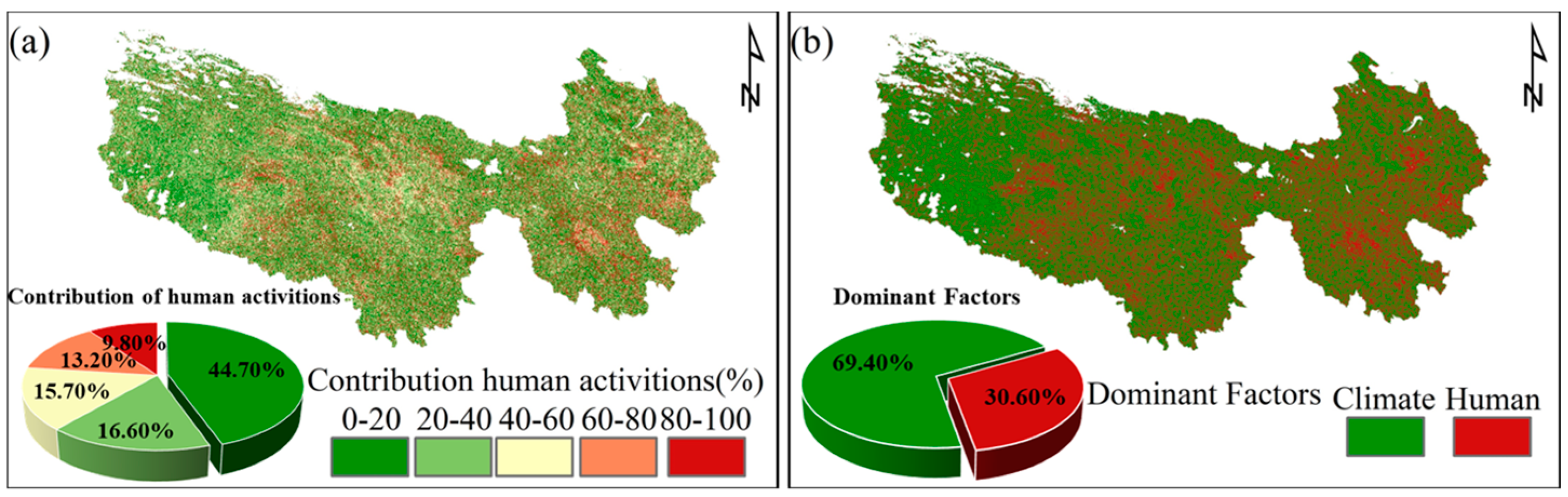
3.5. Contribution of Climate Change and Ecological Engineering to CUE
4. Discussion
4.1. Effects of Vegetation on CUE
4.2. Influence of Climatic Factors on CUE Dynamics
4.3. Ecological Engineering’s Impact on CUE
4.4. Uncertainty
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Valentini, R.; Matteucci, G.; Dolman, A.; Schulze, E.; Rebmann, C.; Moors, E.; Granier, A.; Gross, P.; Jensen, N.; Pilegaard, K.; et al. Respiration as the main determinant of carbon balance in European forests. Nature 2000, 404, 861–865. [Google Scholar] [CrossRef]
- Gifford, R. Plant respiration in productivity models: Conceptualisation, representation and issues for global terrestrial carbon-cycle research. Funct. Plant Biol. 2003, 30, 171–186. [Google Scholar] [CrossRef]
- Manzoni, S.; Taylor, P.; Richter, A.; Porporato, A.; Ågren, G. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol. 2012, 196, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Campioli, M.; Vicca, S.; Luyssaert, S.; Bilcke, J.; Ceschia, E.; Chapin, F.; Ciais, P.; Fernández-Martínez, M.; Malhi, Y.; Obersteiner, M.; et al. Biomass production efficiency controlled by management in temperate and boreal ecosystems. Nat. Geosci. 2015, 8, 843–846. [Google Scholar] [CrossRef]
- Falkowski, P.; Scholes, R.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Högberg, P.; Linder, A.; et al. The global carbon cycle: A test of our knowledge of earth as a system. Science 2000, 290, 291–296. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T., Qin, D., Plattner, G., Tignor, M., Allen, S., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P., et al., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 33–115. [Google Scholar]
- Ise, T.; Litton, C.; Giardina, C.; Ito, A. Comparison of modeling approaches for carbon partitioning: Impact on estimates of global net primary production and equilibrium biomass of woody vegetation from MODIS GPP. J. Geophys. Res. Biogeosciences 2010, 115, G04025. [Google Scholar] [CrossRef]
- Schimel, D.; Pavlick, R.; Fisher, J.; Asner, G.; Saatchi, S.; Townsend, P.; Miller, C.; Frankenberg, C.; Hibbard, K.; Cox, P. Observing terrestrial ecosystems and the carbon cycle from space. Glob. Change Biol. 2015, 21, 1762–1776. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Hu, H.; Sun, W.; Zhu, J.; Liu, G.; Zhou, W.; Zhang, Q.; Shi, P.; Liu, X.; Wu, X.; et al. Effects of national ecological restoration projects on carbon sequestration in China from 2001 to 2010. Proc. Natl. Acad. Sci. USA 2018, 115, 4039–4044. [Google Scholar] [CrossRef]
- Hou, J.; Du, L.; Liu, K.; Hu, Y.; Zhu, Y. Characteristics of vegetation activity and its responses to climate change in desert/grassland biome transition zones in the last 30 years based on GIMMS3g. Theor. Appl. Climatol. 2019, 136, 915–928. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.; Fuch, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Gifford, R. The global carbon cycle: A viewpoint on the missing sink. Funct. Plant Biol. 1994, 21, 1–15. [Google Scholar] [CrossRef]
- Gifford, R. Whole plant respiration and photosynthesis of wheat under increased CO2 concentration and temperature: Long-term vs. short-term distinctions for modelling. Glob. Chang. Biol. 1995, 1, 385–396. [Google Scholar] [CrossRef]
- Dewar, R.; Medlyn, B.; Mcmurtrie, R. A mechanistic analysis of light and carbon use efficiencies. Plant Cell Environ. 1998, 21, 573–588. [Google Scholar] [CrossRef]
- Medlyn, B.; Dewar, R. Comment on the article by R.H. Waring, J.J. Landsberg and M. Williams relating net primary production to gross primary production. Tree Physiol. 1999, 19, 137–138. [Google Scholar] [CrossRef]
- De Lucia, E.; Drake, J.; Thomas, R.; Gonzalez-Meler, M. Forest carbon use efficiency: Is respiration a constant fraction of gross primary production? Glob. Chang. Biol. 2007, 13, 1157–1167. [Google Scholar] [CrossRef]
- Ryan, M.; Lavigne, M.; Gower, S. Annual carbon cost of autotrophic respiration in boreal forest ecosystems in relation to species and climate. J. Geophys. Res. Atmos. 1997, 102, 28871–28883. [Google Scholar] [CrossRef]
- Amthor, J. The McCree–de Wit–Penning de Vries–Thornley respiration paradigms: 30 years later. Ann. Bot. 2000, 86, 1–20. [Google Scholar] [CrossRef]
- Cheng, W.; Sims, D.; Luo, Y.; Coleman, J.; Johnson, D. Photosynthesis, respiration, and net primary production of sunflower stands in ambient and elevated atmospheric CO2 concentrations: An invariant NPP: GPP ratio? Glob. Change Biol. 2000, 6, 931–941. [Google Scholar] [CrossRef]
- Giardina, C.; Ryan, M.; Binkley, D.; Fownes, J. Primary production and carbon allocation in relation to nutrient supply in a tropical experimental forest. Glob. Change Biol. 2003, 9, 1438–1450. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.; Li, J.; Gang, C.; Zhang, Y.; Odeh, I.; Qi, J. Assessing the spatiotemporal dynamic of global grassland carbon use efficiency in response to climate change from 2000 to 2013. Acta Oecologica 2017, 81, 22–31. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, G. Spatial variations and controls of carbon use efficiency in China’s terrestrial ecosystems. Sci. Rep. 2019, 9, 19516. [Google Scholar] [CrossRef] [PubMed]
- Law, B.; Turner, D.; Campbell, J.; Sun, O.; Van Tuyl, S.; Ritts, W.; Cohen, W. Disturbance and climate effects on carbon stocks and fluxes across Western Oregon USA. Glob. Chang. Biol. 2004, 10, 1429–1444. [Google Scholar] [CrossRef]
- Metcalfe, D.; Meir, P.; Aragão, L.; Lobo-do-Vale, R.; Galbraith, D.; Fisher, R.; Chaves, M.; Maroco, J.; Costa, A.; Almeida, S.; et al. Shifts in plant respiration and carbon use efficiency at a large-scale drought experiment in the eastern Amazon. New Phytol. 2010, 187, 608–621. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Piao, S.; Li, X.; Chen, A.; Qin, D. Global patterns of vegetation carbon use efficiency and their climate drivers deduced from MODIS satellite data and process-based models. Agric. For. Meteorol. 2018, 256, 150–158. [Google Scholar] [CrossRef]
- Du, L.; Gong, F.; Zeng, Y.; Ma, L.; Qiao, C.; Wu, H. Carbon use efficiency of terrestrial ecosystems in desert/grassland biome transition zone: A case in Ningxia province, northwest China. Ecol. Indic. 2020, 120, 106971. [Google Scholar] [CrossRef]
- Van Rooyen, N.; Van Rooyen, M.; Theron, G.; Grobbelaar, N. The vegetation of the Roodeplaat Dam Nature Reserve. VI. Photosynthesis and transpiration in woody plants with special reference to an evergreen tree Olea europaea subsp. africana. South Afr. J. Bot. 1986, 52, 301–308. [Google Scholar] [CrossRef]
- Boegh, E.; Soegaard, H.; Broge, N.; Hasager, C.; Jensen, N.; Schelde, K.; Thomsen, A. Airborne multispectral data for quantifying leaf area index, nitrogen concentration, and photosynthetic efficiency in agriculture. Remote Sens. Environ. 2002, 81, 179–193. [Google Scholar] [CrossRef]
- Shao, Q.; Cao, W.; Fan, J.; Huang, L.; Xu, X. Effects of an ecological conservation and restoration project in the Three-River Source Region, China. J. Geogr. Sci. 2017, 27, 183–204. [Google Scholar] [CrossRef]
- Tang, X.; Liu, D.; Song, K.; Munger, J.; Zhang, B.; Wang, Z. A new model of net ecosystem carbon exchange for the deciduous-dominated forest by integrating MODIS and flux data. Ecol. Eng. 2011, 37, 1567–1571. [Google Scholar] [CrossRef]
- Fu, G.; Sun, W.; Yu, C.; Zhang, X.; Shen, Z.; Li, Y.; Yang, P.; Zhou, N. Clip** alters the response of biomass production to experimental warming: A case study in an alpine meadow on the Tibetan Plateau, China. J. Mt. Sci. 2015, 12, 935–942. [Google Scholar] [CrossRef]
- Peng, S.; Ding, Y.; Liu, W.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, D.; Gan, R.; Chiew, F.; McVicar, T.; Zhang, Q.; Yang, Y. Coupled estimation of 500m and 8-day resolution global evapotranspiration and gross primary production in 2002–2017. Remote Sens. Environ. 2019, 222, 165–182. [Google Scholar] [CrossRef]
- Akihiko, I. Constraining size-dependence of vegetation respiration rates. Sci. Rep. 2020, 10, 4304. [Google Scholar]
- Hirsch, R.; Slack, J.; Smith, R. Techniques of trend analysis for monthly water quality data. Water Resour. Res. 1982, 18, 107–121. [Google Scholar] [CrossRef]
- Mann, H. Non-parametric tests against trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M. Rank Correlation Measures. In Charles Griffin, 4th ed.; Charles Griffin: London, UK, 1975. [Google Scholar]
- Zhang, Y.; Kang, S.; Ward, E.; Ding, R.; Zhang, X.; Zheng, R. Evapotranspiration components determined by sap flow and microlysimetry techniques of a vineyard in northwest china: Dynamics and influential factors. Agric. Water Manag. 2011, 98, 1207–1214. [Google Scholar] [CrossRef]
- Nooni, I.; Wang, G.; Hagan, D.; Lu, J.; Ullah, W.; Li, S. Evapotranspiration and its Components in the Nile River Basin Based on Long-Term Satellite Assimilation Product. Water 2019, 11, 1400. [Google Scholar] [CrossRef]
- Tong, S.; Zhang, J.; Bao, Y.; Lai, Q.; Lian, X.; Li, N.; Bao, Y. Analyzing vegetation dynamic trend on the Mongolian Plateau based on the Hurst exponent and influencing factors from 1982–2013. J. Geogr. Sci. 2018, 28, 595–610. [Google Scholar] [CrossRef]
- Partal, T.; Kahya, E. Trend analysis in Turkish precipitation data. Hydrol. Process. Int. J. 2006, 20, 2011–2026. [Google Scholar] [CrossRef]
- Xia, J.; Yi, G.; Zhang, T.; Zhou, X.; Miao, J.; Bie, X. Interannual variation in the start of vegetation growing season and its response to climate change in the Qinghai–Tibet Plateau derived from MODIS data during 2001 to 2016. J. Appl. Remote Sens. 2019, 13, 048506. [Google Scholar] [CrossRef]
- Evans, J.; Geerken, R. Discrimination between climate and human-induced dryland degradation. J. Arid Environ. 2004, 57, 535–554. [Google Scholar] [CrossRef]
- Wessels, K.; Prince, S.; Malherbe, J.; Small, J.; Frost, P.; VanZyl, D. Can human-induced land degradation be distinguished from the effects of rainfall variability? A case study in South Africa. J. Arid Environ. 2007, 68, 271–297. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.; Wang, H.; Deng, F.; Li, X.; Mi, X. Effects of converting natural grasslands into planted grasslands on ecosystem respiration: A case study in Inner Mongolia, China. J. Arid Land 2017, 9, 38–50. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, G.; Liu, T.; Xue, B.; Yinglan, A. Controls of carbon flux in a semi-arid grassland ecosystem experiencing wetland loss: Vegetation patterns and environmental variables. Agric. For. Meteorol. 2018, 259, 196–210. [Google Scholar] [CrossRef]
- Yang, C.; Li, G.; Yan, L.; Wu, J.; Li, J.; Liu, S.; Lu, Y. Effects of different vegetation types on ecosystem respiration in semiarid Loess Hilly Region, Central Gansu Province, China. Ecol. Indic. 2022, 145, 109683. [Google Scholar] [CrossRef]
- Yu, G.; Zhu, X.; Fu, Y.; He, H.; Wang, Q.; Wen, X.; Li, X.; Zhang, M.; Zhang, L.; Su, W.; et al. Spatial patterns and climate drivers of carbon fluxes in terrestrial ecosystems of China. Glob. Chang. Biol. 2013, 19, 798–810. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yu, G.; Ge, J.; Sun, X.; Hirano, T.; Saigusa, N.; Wang, Q.; Zhu, X.; Zhang, Y.; Zhang, J.; et al. Temperature and precipitation control of the spatial variation of terrestrial ecosystem carbon exchange in the Asian region. Agric. For. Meteorol. 2013, 182, 266–276. [Google Scholar] [CrossRef]
- Wei, S.; Yi, C.; Fang, W.; Hendrey, G. A global study of GPP focusing on light-use efficiency in a random forest regression model. Ecosphere 2017, 8, e01724. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, M.; Chen, H.; Adams, J. Global pattern of NPP to GPP ratio derived from MODIS data: Effects of ecosystem type, geographical location and climate. Glob. Ecol. Biogeogr. 2009, 18, 280–290. [Google Scholar] [CrossRef]
- Potter, C.; Randerson, J.; Field, C.; Matson, P.; Vitousek, P.; Mooney, H.; Klooster, S. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Malhi, Y. The productivity, metabolism and carbon cycle of tropical forest vegetation. J. Ecol. 2012, 100, 65–75. [Google Scholar] [CrossRef]
- Atkin, O.; Evans, J.; Ball, M.; Lambers, H.; Pons, T. Leaf respiration of snow gum in the light and dark. Interactions between temperature and irradiance. Plant Physiol. 2000, 122, 915–924. [Google Scholar] [CrossRef]
- Bolstad, P.; Reich, P.; Lee, T. Rapid temperature acclimation of leaf respiration rates in Quercus alba and Quercus rubra. Tree Physiol. 2003, 23, 969–976. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Piao, S.; Luyssaert, S.; Ciais, P.; Janssens, I.; Chen, A.; Cao, C.; Fang, J.; Friedlingstein, P.; Luo, Y.; Wang, S. Forest annual carbon cost: A global-scale analysis of autotrophic respiration. Ecology 2010, 91, 652–661. [Google Scholar] [CrossRef]
- Kwon, Y.; Larsen, C. Effects of forest type and environmental factors on forest carbon use efficiency assessed using MODIS and FIA data across the eastern USA. Int. J. Remote Sens. 2013, 34, 8425–8448. [Google Scholar] [CrossRef]
- Chambers, J.; Tribuzy, E.; Toledo, L.; Crispim, B.; Higuchi, N.; Santos, J.; Araújo, A.; Kruijt, B.; Nobre, A.; Trumbore, S. Respiration from a tropical forest ecosystem: Partitioning of sources and low carbon use efficiency. Ecol. Appl. 2004, 14, 72–88. [Google Scholar] [CrossRef]
- Alfieri, J.; Xiao, X.; Niyogi, D.; Pielke Sr, R.; Chen, F.; LeMone, M. Satellite-based modeling of transpiration from the grasslands in the Southern Great Plains, USA. Glob. Planet. Chang. 2009, 67, 78–86. [Google Scholar] [CrossRef]
- Monteith, J. Does transpiration limit the growth of vegetation or vice versa? J. Hydrol. 1988, 100, 57–68. [Google Scholar] [CrossRef]
- Law, B.; Falge, E.; Gu, L.; Baldocchi, D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.; et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef]
- Berry, S.; Roderick, M. Gross primary productivity and transpiration flux of the Australian vegetation from 1788 to 1988 AD: Effects of CO2 and land use change. Glob. Chang. Biol. 2004, 10, 1884–1898. [Google Scholar] [CrossRef]
- Vesala, T.; Suni, T.; Rannik, Ü.; Keronen, P.; Markkanen, T.; Sevanto, S.; Grönholm, T.; Smolander, S.; Kulmala, H.; Ojansuu, R.; et al. Effect of thinning on surface fluxes in a boreal forest. Glob. Biogeochem. Cycles 2005, 19, GB2001. [Google Scholar] [CrossRef]
- Jardine, K.; Cobello, L.; Teixeira, L.; East, M.; Levine, S.; Gimenez, B.; Robles, E.; Spanner, G.; Koven, C.; Xu, C.; et al. Stem respiration and growth in a central Amazon rainforest. Trees 2022, 36, 991–1004. [Google Scholar] [CrossRef]
- Tong, X.; Brandt, M.; Yue, Y.; Horion, S.; Wang, K.; Keersmaecker, W.; Tian, F.; Schurgers, G.; Xiao, X.; Luo, Y.; et al. Increased vegetation growth and carbon stock in China karst via ecological engineering. Nat. Sustain. 2018, 1, 44–50. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, B.; Wu, C.; Huete, A.; Gonsamo, A.; Lei, L.; Ponce-Campos, G.; Liu, X.; Wu, Y. Country-level net primary production distribution and response to drought and land cover change. Sci. Total Environ. 2017, 574, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, S.; Bai, X.; Tian, Y.; Luo, G.; Wang, J.; Li, Q.; Chen, F.; Deng, Y.; Yang, Y.; et al. Climate change weakens the positive effect of human activities on karst vegetation productivity restoration in southern China. Ecol. Indic. 2020, 115, 106392. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Duan, H.; Pan, J.; Gao, X.; Chen, J.; Wu, S.; Tan, D. Quantifying Carbon Use Efficiency: Unraveling the Impact of Climate Change and Ecological Engineering on Vegetation in the Three Rivers Source Region. Remote Sens. 2024, 16, 2909. https://doi.org/10.3390/rs16162909
Luo Y, Duan H, Pan J, Gao X, Chen J, Wu S, Tan D. Quantifying Carbon Use Efficiency: Unraveling the Impact of Climate Change and Ecological Engineering on Vegetation in the Three Rivers Source Region. Remote Sensing. 2024; 16(16):2909. https://doi.org/10.3390/rs16162909
Chicago/Turabian StyleLuo, Yixia, Hengyi Duan, Jing Pan, Xue Gao, Jilong Chen, Shengjun Wu, and Daming Tan. 2024. "Quantifying Carbon Use Efficiency: Unraveling the Impact of Climate Change and Ecological Engineering on Vegetation in the Three Rivers Source Region" Remote Sensing 16, no. 16: 2909. https://doi.org/10.3390/rs16162909
APA StyleLuo, Y., Duan, H., Pan, J., Gao, X., Chen, J., Wu, S., & Tan, D. (2024). Quantifying Carbon Use Efficiency: Unraveling the Impact of Climate Change and Ecological Engineering on Vegetation in the Three Rivers Source Region. Remote Sensing, 16(16), 2909. https://doi.org/10.3390/rs16162909