
Spatial–Temporal Variations in the Climate, Net Ecosystem Productivity, and Efficiency of Water and Carbon Use in the Middle Reaches of the Yellow River

Abstract
:
1. Introduction
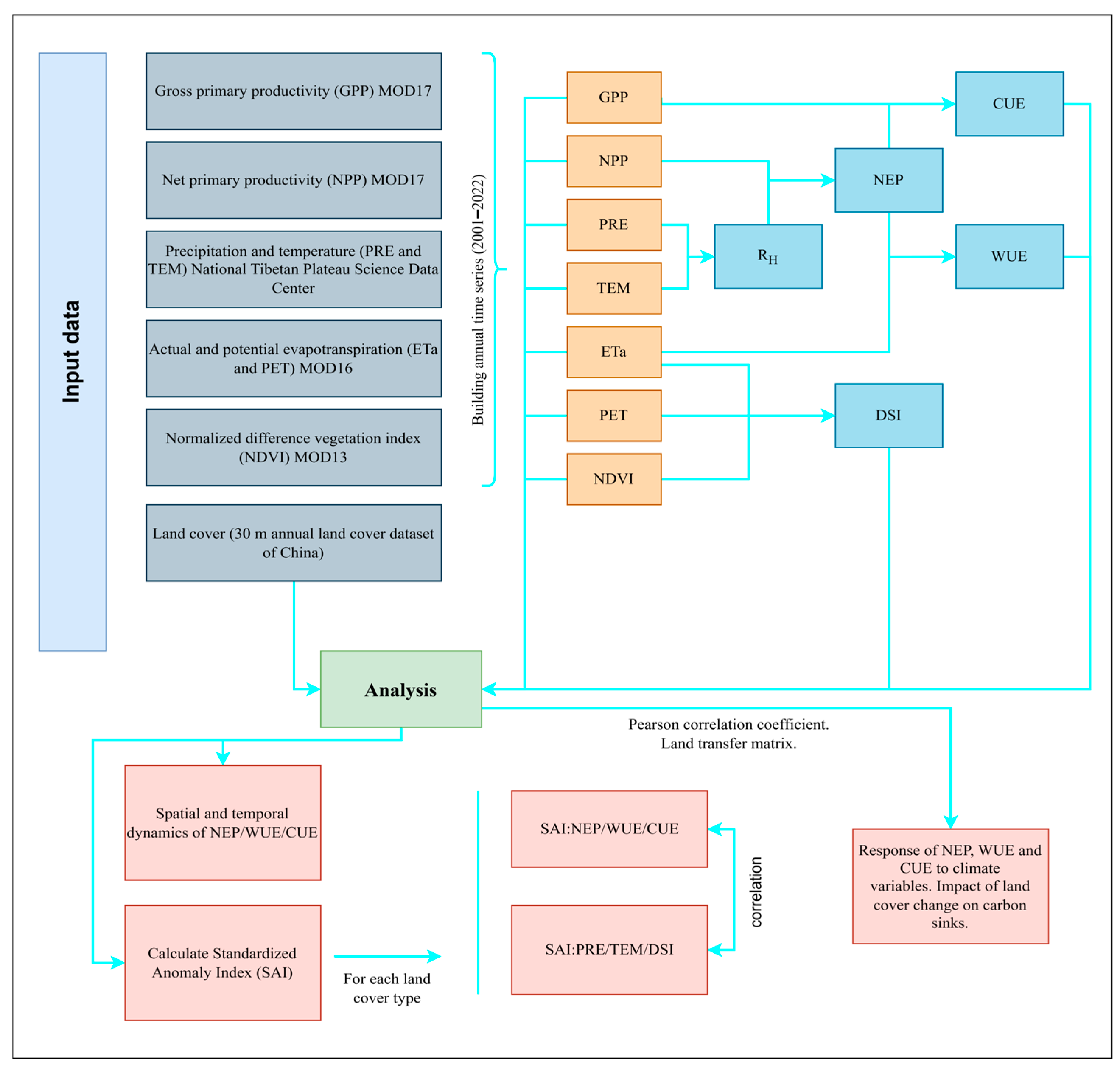
2. Materials and Methods
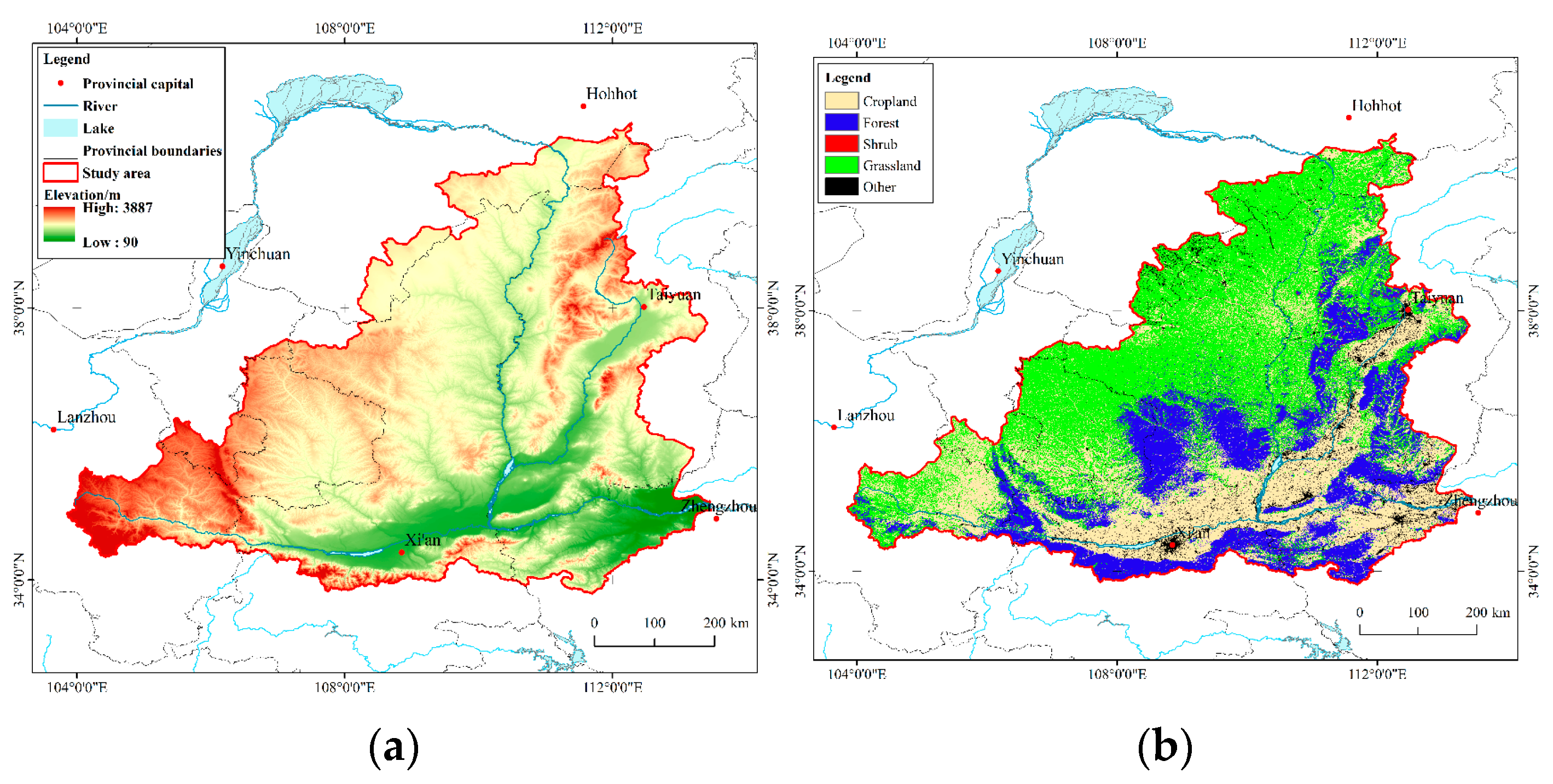
2.1. Study Area
2.2. Data and Pre-Processing
2.3. Methodology
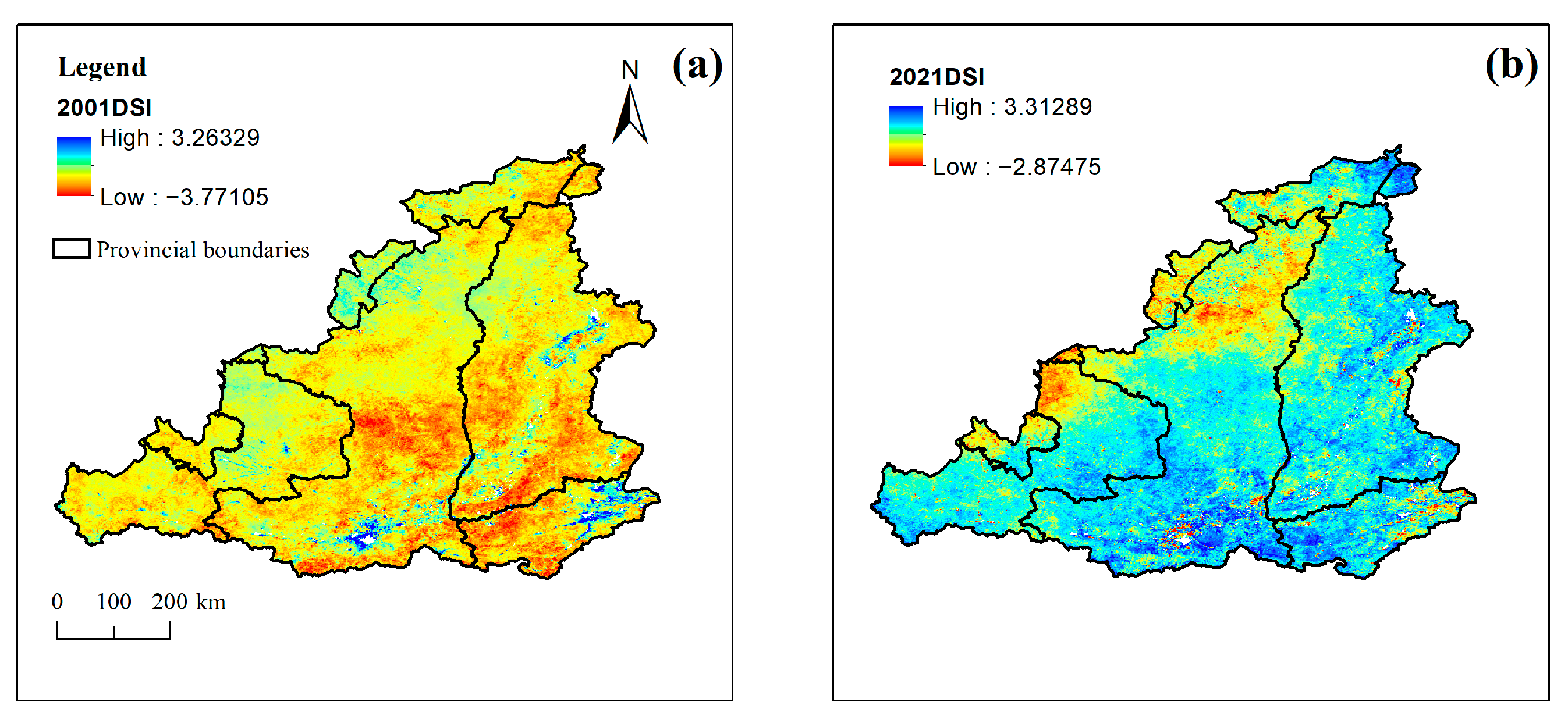
2.3.1. Drought Severity Index (DSI)
2.3.2. NEP, WUE, and CUE
2.3.3. Trend Analysis
2.3.4. Standardized Anomaly Indices
2.3.5. Pearson Correlation Coefficient
3. Results
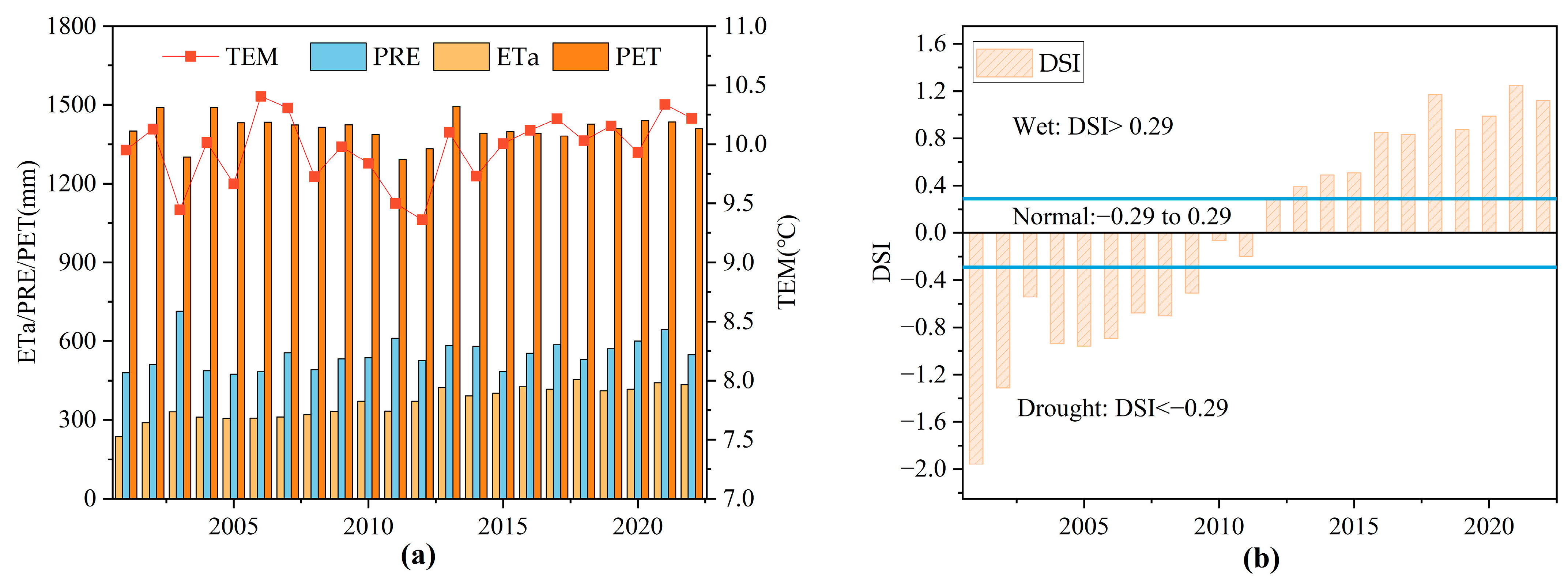
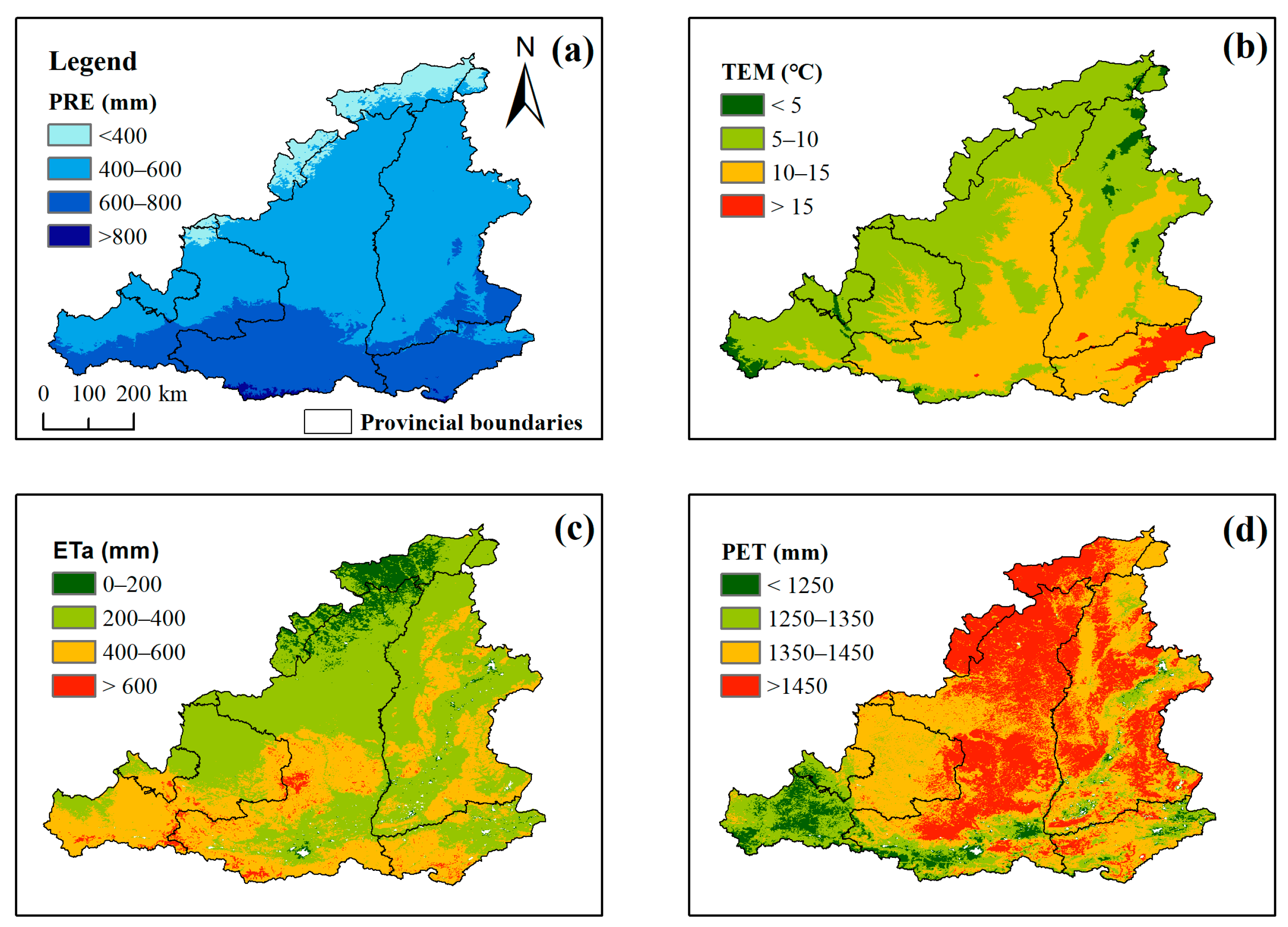
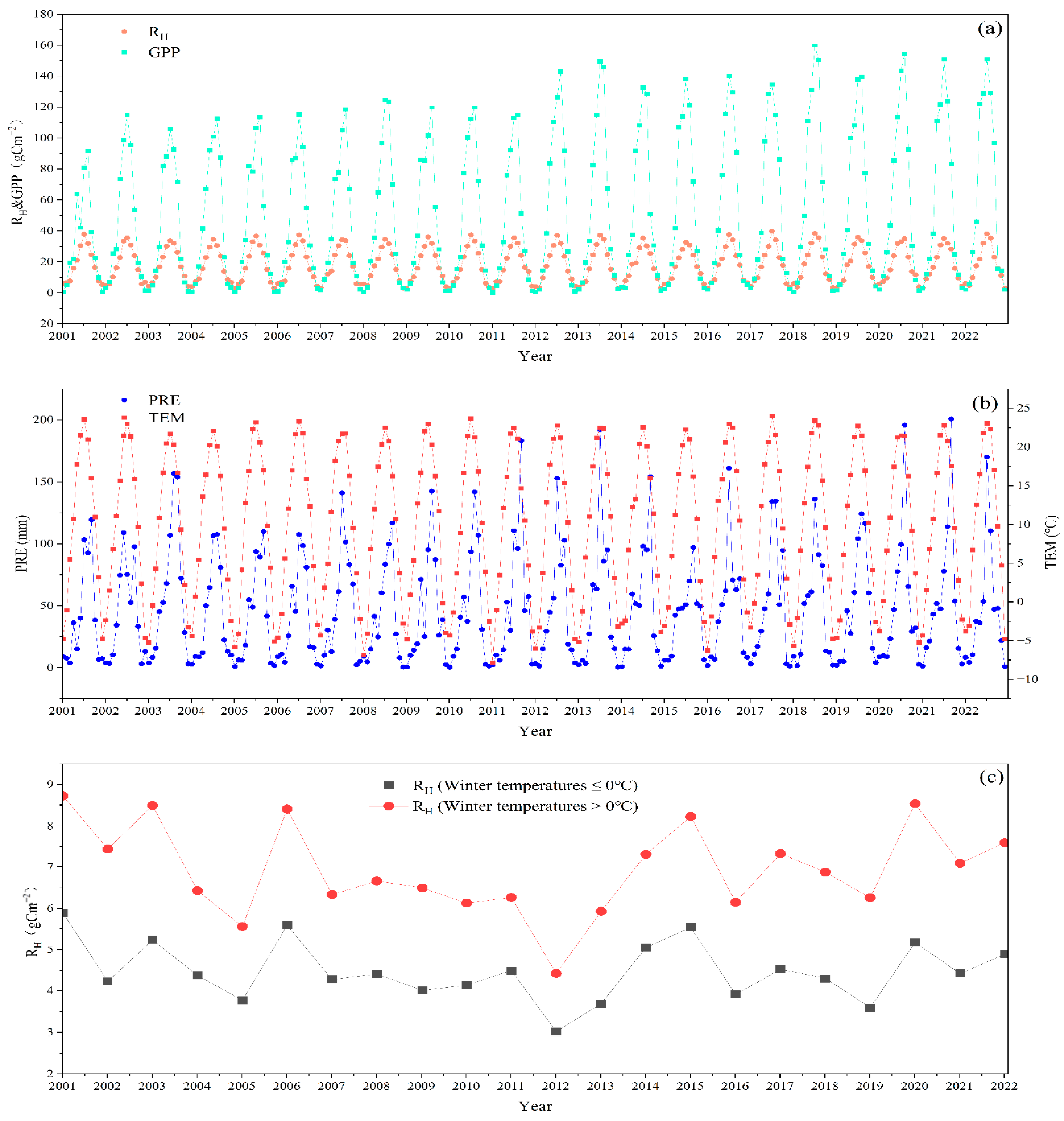
3.1. Climate Conditions from 2001 to 2022
3.2. Temporal and Spatial Dynamics of NEP/WUE/CUE
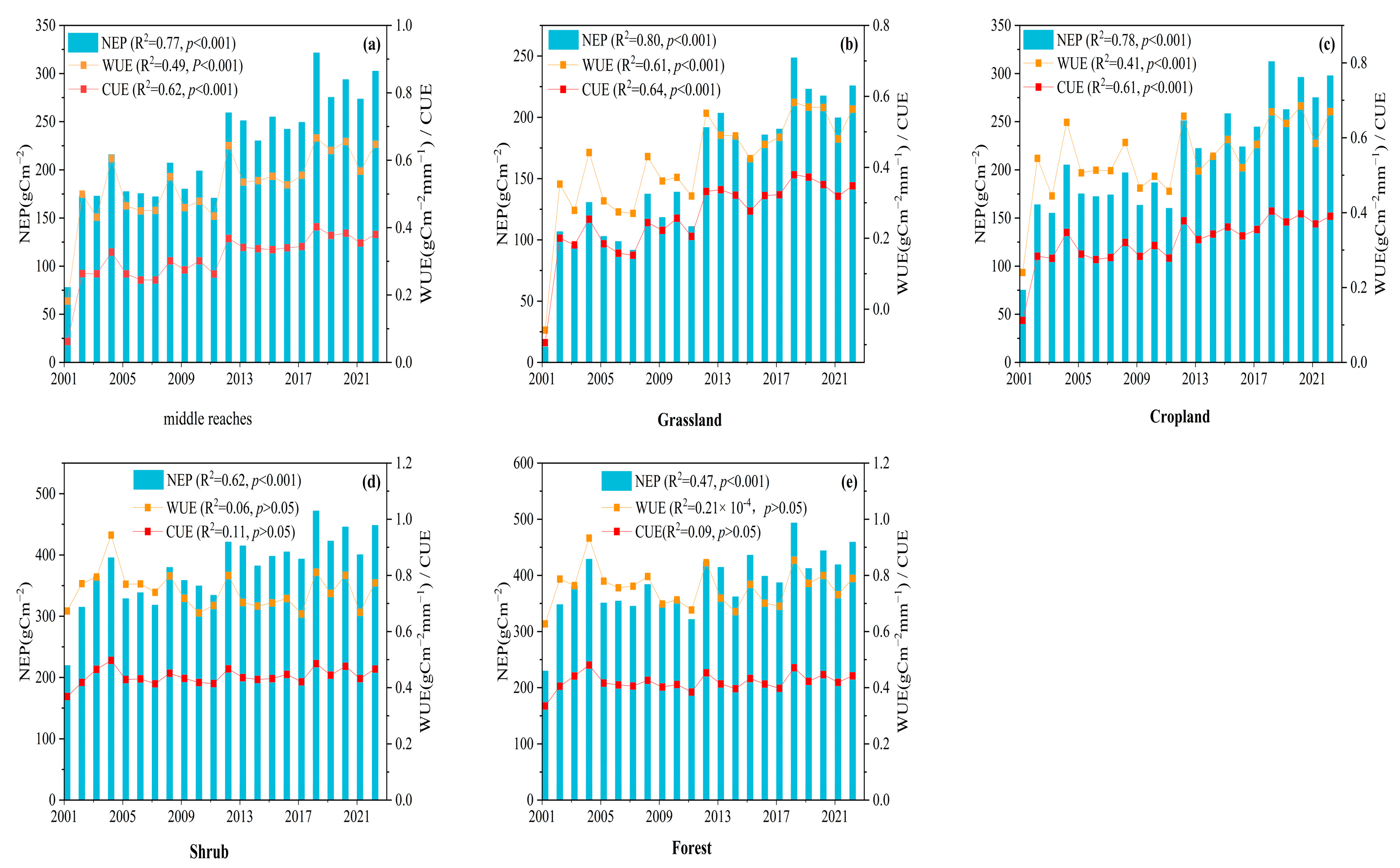
3.2.1. Temporal Variability in NEP/WUE/CUE
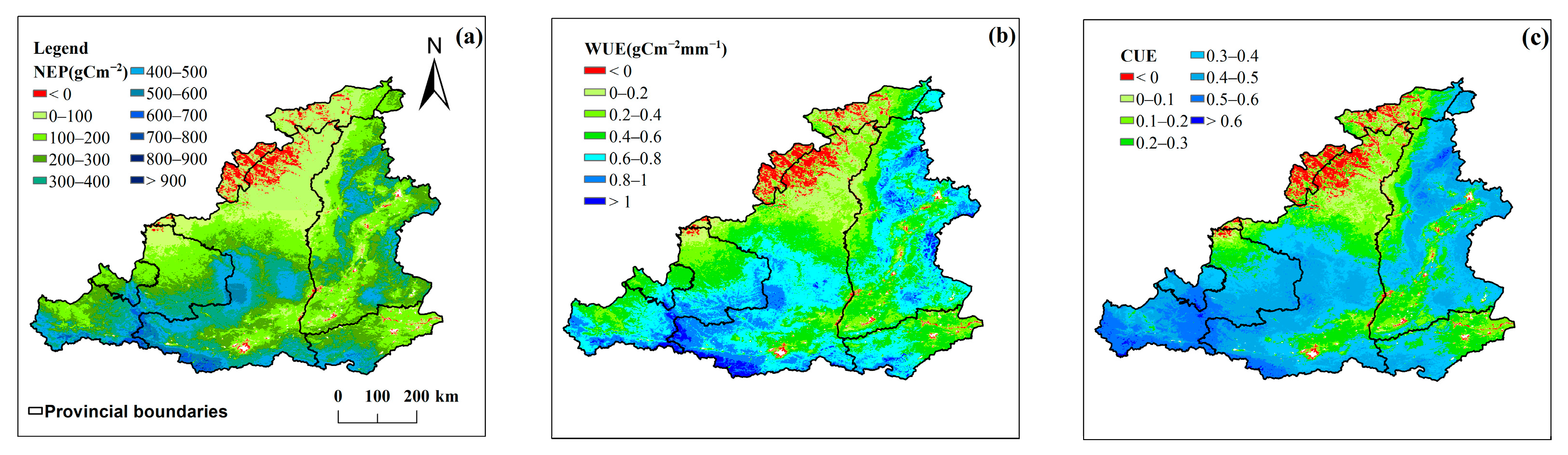
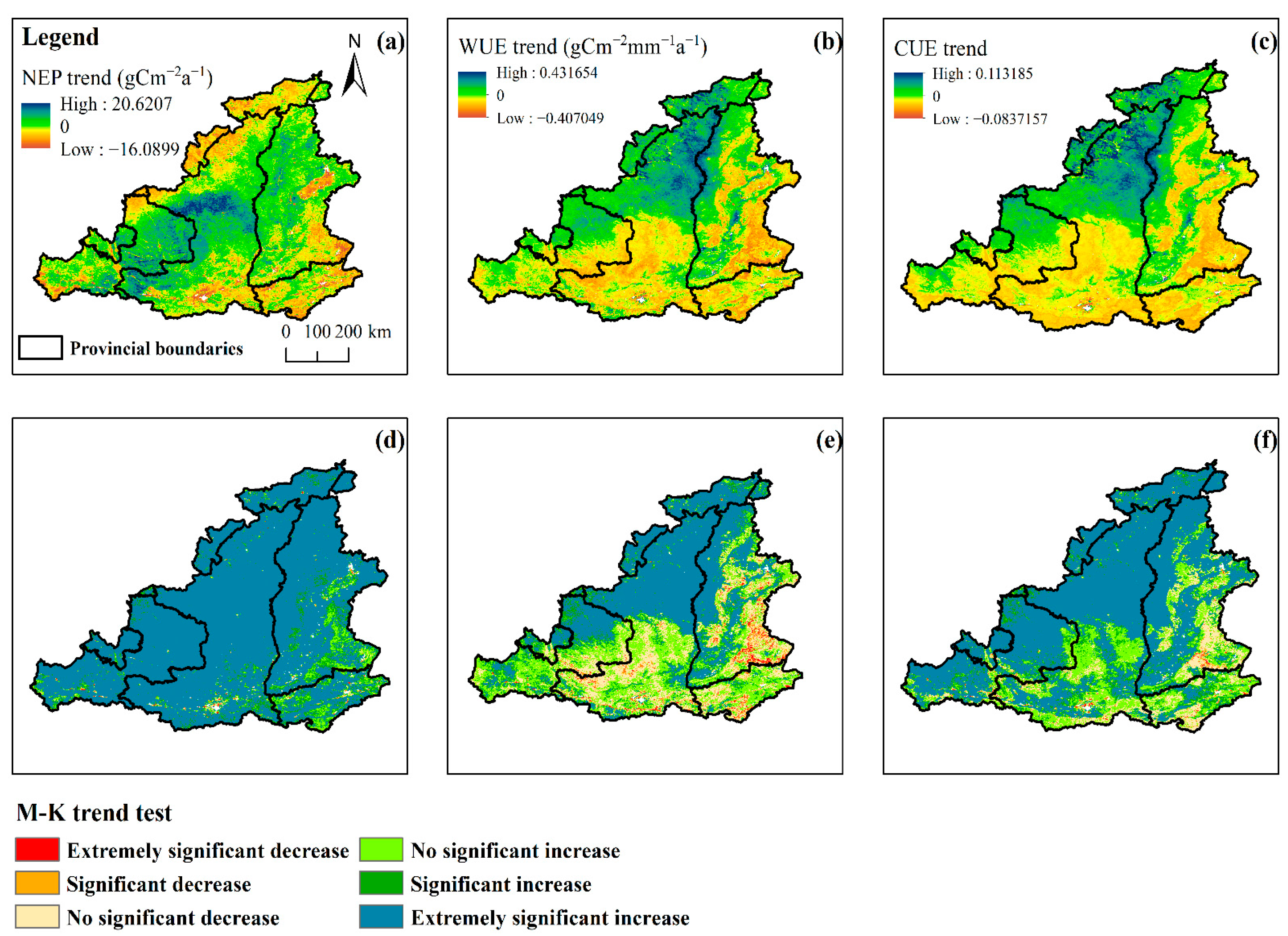
3.2.2. NEP/WUE/CUE Spatial Patterns
3.3. Interannual Anomalies in NEP, WUE, CUE, and Climate Variables
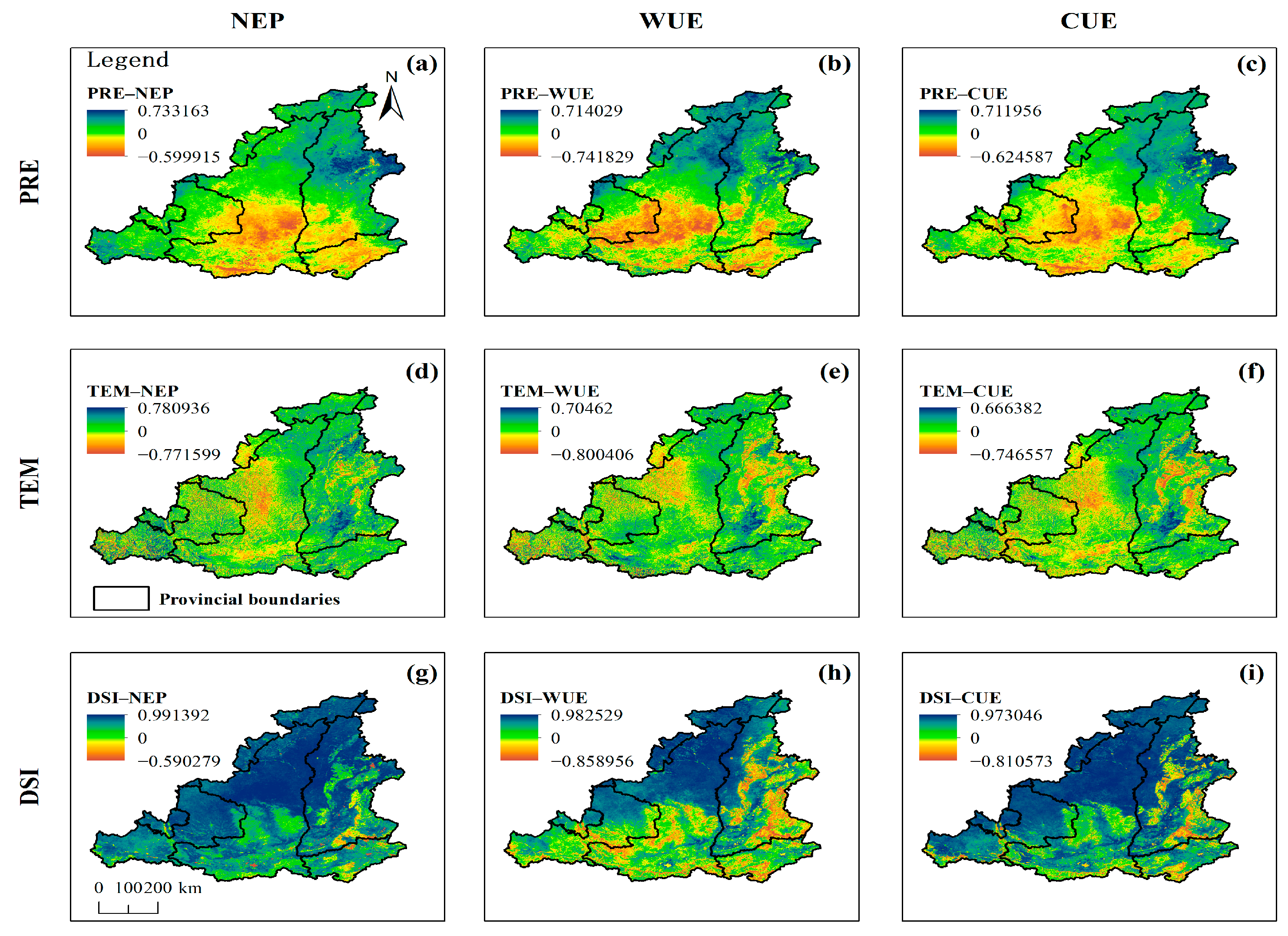
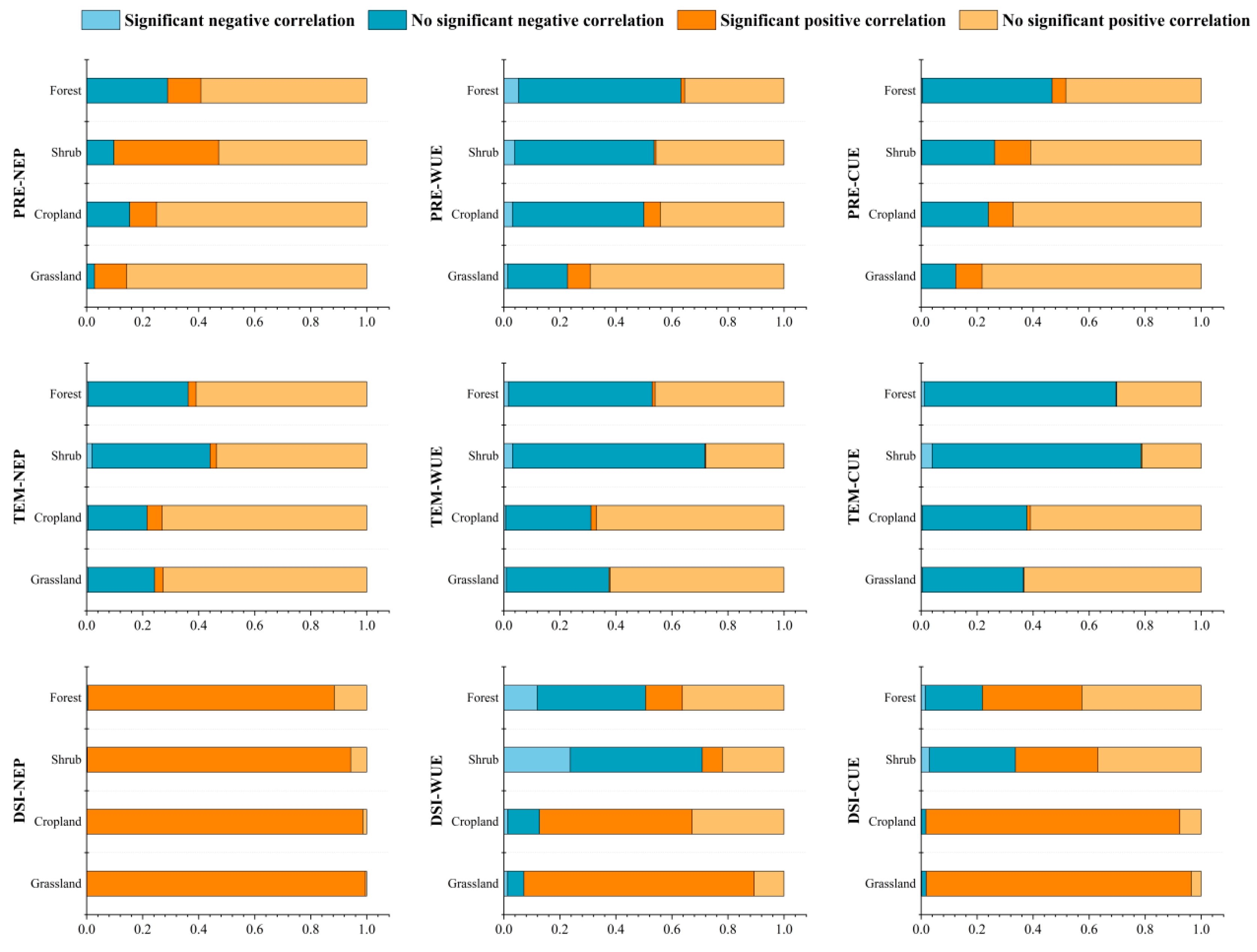
3.4. Correlation of NEP, WUE, and CUE with Climate Variables
3.4.1. Correlation with Precipitation
3.4.2. Correlation with Temperature
3.4.3. Correlation with DSI
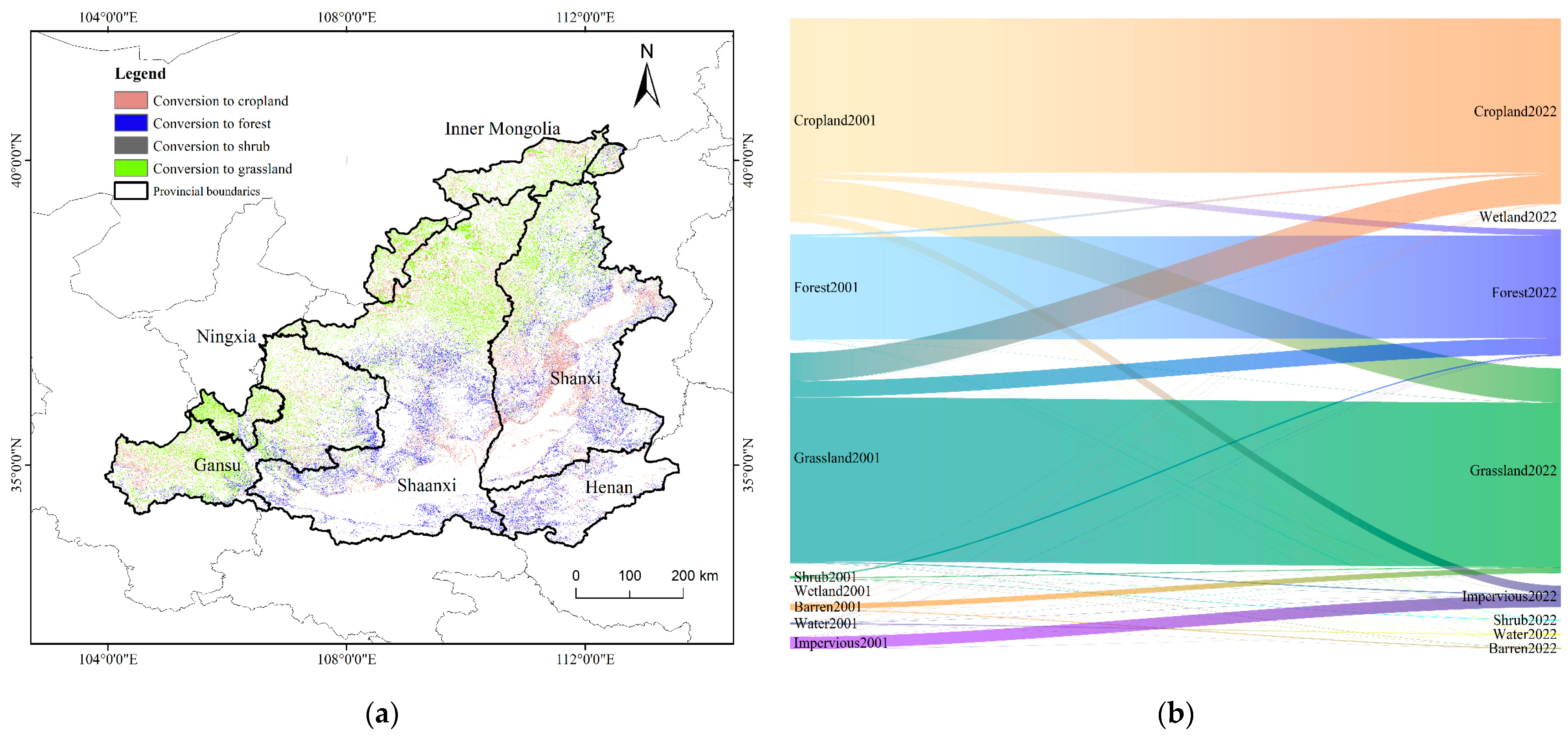
3.5. Impact of Land Cover on Carbon Sinks
4. Discussion
4.1. Climate Change and Its Impacts
4.2. Characterization of Changes in NEP, WUE, and CUE
4.3. Analysis of Impact Mechanisms
4.4. Uncertainty Analysis and Directions for Further Research
5. Conclusions
- (1)
- Based on the changes in precipitation, temperature, ETa, PET, and DSI from 2001 to 2022, the drought trend in the MRYR is weakening, and climate change has been moving toward warming and humidification.
- (2)
- The NEP, WUE, and CUE generally showed an upward trend from 2001 to 2022, with annual mean values of 221.49 gCm−2, 0.52 gC m−2mm−1, and 0.31, respectively, and average growth rates of 7.75 gC m−2a−1, 0.012 gC m−2 mm−1a−1, and 0.009 a−1, respectively. Spatially, the fastest growth rate was observed in the more arid northwestern part of the MRYR. In contrast, the growth rate was lower in the southeastern region, where water resources are relatively abundant. Of the four vegetation types, the grassland NEP, WUE, and CUE had the highest growth rates, followed by cropland; however, in the forests and shrubs, the WUE and CUE growth rates tended to be close to 0 or less than 0.
- (3)
- An analysis of the standardized anomaly indices showed that precipitation accumulation contributed to the accumulation of carbon sinks. Among the three selected meteorological factors, the DSI had the highest correlation with the NEP, WUE, and CUE, and the correlation of precipitation with the NEP, WUE, and CUE showed latitudinal zonality. This suggests that precipitation is the main control factor affecting the water–carbon cycle in the region, not temperature.
- (4)
- From 2001 to 2022, a total of 67,671.27 km2 of land cover types changed in the MRYR, and the regional NEP increased by 15.07 Tg. The interconversion between grassland, forest, and cropland accounted for 81.60% of the changed area.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vörösmarty, C.J.; Green, P.; Salisbury, J.; Lammers, R.B. Global Water Resources: Vulnerability from Climate Change and Population Growth. Science 2000, 289, 284–288. [Google Scholar] [CrossRef]
- Rosegrant, M.; Cai, X.; Cline, S. Will The World Run Dry? Global Water and Food Security. Environ. Sci. Policy Sustain. Dev. 2003, 45, 24–36. [Google Scholar] [CrossRef]
- Gang, C.; Wang, Z.; Chen, Y.; Yang, Y.; Li, J.; Cheng, J.; Qi, J.; Odeh, I. Drought-Induced Dynamics of Carbon and Water Use Efficiency of Global Grasslands from 2000 to 2011. Ecol. Indic. 2016, 67, 788–797. [Google Scholar] [CrossRef]
- Yu, P.; Krysanova, V.; Wang, Y.; Xiong, W.; Mo, F.; Shi, Z.; Liu, H.; Vetter, T.; Huang, S. Quantitative Estimate of Water Yield Reduction Caused by Forestation in a Water-Limited Area in Northwest China. Geophys. Res. Lett. 2009, 36, L02406. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India Lead in Greening of the World through Land-Use Management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, B.; Wu, C.; Huete, A.R.; Gonsamo, A.; Lei, L.; Ponce-Campos, G.E.; Liu, X.; Wu, Y. Country-Level Net Primary Production Distribution and Response to Drought and Land Cover Change. Sci. Total Environ. 2017, 574, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.A.; Exbrayat, J.-F.; van der Velde, I.R.; Feng, L.; Williams, M. The Decadal State of the Terrestrial Carbon Cycle: Global Retrievals of Terrestrial Carbon Allocation, Pools, and Residence Times. Proc. Natl. Acad. Sci. USA 2016, 113, 1285–1290. [Google Scholar] [CrossRef]
- Zhang, J.; Hao, X.; Hao, H.; Fan, X.; Li, Y. Climate Change Decreased Net Ecosystem Productivity in the Arid Region of Central Asia. Remote Sens. 2021, 13, 4449. [Google Scholar] [CrossRef]
- Chapin, F.S.; Woodwell, G.M.; Randerson, J.T.; Rastetter, E.B.; Lovett, G.M.; Baldocchi, D.D.; Clark, D.A.; Harmon, M.E.; Schimel, D.S.; Valentini, R.; et al. Reconciling Carbon-Cycle Concepts, Terminology, and Methods. Ecosystems 2006, 9, 1041–1050. [Google Scholar] [CrossRef]
- Liang, L.; Geng, D.; Yan, J.; Qiu, S.; Shi, Y.; Wang, S.; Wang, L.; Zhang, L.; Kang, J. Remote Sensing Estimation and Spatiotemporal Pattern Analysis of Terrestrial Net Ecosystem Productivity in China. Remote Sens. 2022, 14, 1902. [Google Scholar] [CrossRef]
- Ruimy, A.; Saugier, B.; Dedieu, G. Methodology for the Estimation of Terrestrial Net Primary Production from Remotely Sensed Data. J. Geophys. Res. Atmos. 1994, 99, 5263–5283. [Google Scholar] [CrossRef]
- Feng, X.; Fan, Q.; Qu, J.; Ding, X.; Niu, Z. Characteristics of Carbon Sources and Sinks and Their Relationships with Climate Factors during the Desertification Reversal Process in Yulin, China. Front. For. Glob. Chang. 2023, 6, 1288449. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Wang, Y.; Fang, G. Dynamic Changes in Terrestrial Net Primary Production and Their Effects on Evapotranspiration. Hydrol. Earth Syst. Sci. 2016, 20, 2169–2178. [Google Scholar] [CrossRef]
- Liu, Y.; Xiao, J.; Ju, W.; Zhou, Y.; Wang, S.; Wu, X. Water Use Efficiency of China’s Terrestrial Ecosystems and Responses to Drought. Sci. Rep. 2015, 5, 13799. [Google Scholar] [CrossRef]
- Qiu, R.; Han, G.; Ma, X.; Xu, H.; Shi, T.; Zhang, M. A Comparison of OCO-2 SIF, MODIS GPP, and GOSIF Data from Gross Primary Production (GPP) Estimation and Seasonal Cycles in North America. Remote Sens. 2020, 12, 258. [Google Scholar] [CrossRef]
- Liu, W.L.; Jiang, L.L.; Liu, B.; Xiao, Z.L.; Zhang, Y.W. Spatio-Temporal Evolution Characteristics and Driving Factors Analysis of Vegetation Carbon Sources/Sinks in China. Acta Ecol. Sin. 2024, 44, 1456–1467. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation Among Biomes in Temporal Dynamics of Aboveground Primary Production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef]
- van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.E.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M.; et al. Drought and Ecosystem Carbon Cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- Khalifa, M.; Elagib, N.A.; Ribbe, L.; Schneider, K. Spatio-Temporal Variations in Climate, Primary Productivity and Efficiency of Water and Carbon Use of the Land Cover Types in Sudan and Ethiopia. Sci. Total Environ. 2018, 624, 790–806. [Google Scholar] [CrossRef]
- Tucker, C.L.; Bell, J.; Pendall, E.; Ogle, K. Does Declining Carbon-use Efficiency Explain Thermal Acclimation of Soil Respiration with Warming? Glob. Chang. Biol. 2013, 19, 252–263. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D.L.; Xu, X.; Litvak, M.E. Plant, Microbial and Ecosystem Carbon Use Efficiencies Interact to Stabilize Microbial Growth as a Fraction of Gross Primary Production. New Phytol. 2017, 214, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- DI, Y.; Zeng, H.; Zhang, Y.; Chen, N.; Cong, N. Research Advances in Carbon Use Efficiency at Multiple Scales. Chin. J. Ecol. 2021, 40, 1849–1860. [Google Scholar] [CrossRef]
- Xia, J.; McGuire, A.D.; Lawrence, D.; Burke, E.; Chen, G.; Chen, X.; Delire, C.; Koven, C.; MacDougall, A.; Peng, S.; et al. Terrestrial Ecosystem Model Performance in Simulating Productivity and Its Vulnerability to Climate Change in the Northern Permafrost Region. J. Geophys. Res. Biogeosci. 2017, 122, 430–446. [Google Scholar] [CrossRef]
- Manzoni, S.; Čapek, P.; Porada, P.; Thurner, M.; Winterdahl, M.; Beer, C.; Brüchert, V.; Frouz, J.; Herrmann, A.M.; Lindahl, B.D.; et al. Reviews and Syntheses: Carbon Use Efficiency from Organisms to Ecosystems—Definitions, Theories, and Empirical Evidence. Biogeosciences 2018, 15, 5929–5949. [Google Scholar] [CrossRef]
- Tang, X.; Li, H.; Desai, A.R.; Nagy, Z.; Luo, J.; Kolb, T.E.; Olioso, A.; Xu, X.; Yao, L.; Kutsch, W.; et al. How Is Water-Use Efficiency of Terrestrial Ecosystems Distributed and Changing on Earth? Sci. Rep. 2014, 4, 7483. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Inatomi, M. Water-Use Efficiency of the Terrestrial Biosphere: A Model Analysis Focusing on Interactions between the Global Carbon and Water Cycles. J. Hydrometeorol. 2012, 13, 681–694. [Google Scholar] [CrossRef]
- DeLUCIA, E.H.; Drake, J.E.; Thomas, R.B.; Gonzalez-Meler, M. Forest Carbon Use Efficiency: Is Respiration a Constant Fraction of Gross Primary Production? Glob. Chang. Biol. 2007, 13, 1157–1167. [Google Scholar] [CrossRef]
- Zhang, T.; Peng, J.; Liang, W.; Yang, Y.; Liu, Y. Spatial–Temporal Patterns of Water Use Efficiency and Climate Controls in China’s Loess Plateau during 2000–2010. Sci. Total Environ. 2016, 565, 105–122. [Google Scholar] [CrossRef]
- Niu, S.; Xing, X.; Zhang, Z.; Xia, J.; Zhou, X.; Song, B.; Li, L.; Wan, S. Water-Use Efficiency in Response to Climate Change: From Leaf to Ecosystem in a Temperate Steppe. Glob. Chang. Biol. 2011, 17, 1073–1082. [Google Scholar] [CrossRef]
- Fu, B.; Wu, X.; Wang, Z.; Wu, X.; Wang, S. Coupling Human and Natural Systems for Sustainability: Experience from China’s Loess Plateau. Earth Syst. Dyn. 2022, 13, 795–808. [Google Scholar] [CrossRef]
- Yu, Y.; Zhao, W.; Martinez-Murillo, J.F.; Pereira, P. Loess Plateau: From Degradation to Restoration. Sci. Total Environ. 2020, 738, 140206. [Google Scholar] [CrossRef]
- Naeem, S.; Zhang, Y.; Zhang, X.; Tian, J.; Abbas, S.; Luo, L.; Meresa, H.K. Both Climate and Socioeconomic Drivers Contribute to Vegetation Greening of the Loess Plateau. Sci. Bull. 2021, 66, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, S.; Fu, B.; Feng, X.; Chen, Y. Socio-Ecological Changes on the Loess Plateau of China after Grain to Green Program. Sci. Total Environ. 2019, 678, 565–573. [Google Scholar] [CrossRef]
- Ogutu, B.O.; D’Adamo, F.; Dash, J. Impact of Vegetation Greening on Carbon and Water Cycle in the African Sahel-Sudano-Guinean Region. Glob. Planet. Chang. 2021, 202, 103524. [Google Scholar] [CrossRef]
- Li, Y.; Liu, C.; Liu, X.; Liang, K.; Bai, P.; Feng, Y. Impact of the Grain for Green Project on the Land Use/Cover Change in the Middle Yellow River. J. Nat. Resour. 2016, 31, 2005–2020. [Google Scholar] [CrossRef]
- Yang, J.; Huang, X. The 30 m Annual Land Cover Dataset and Its Dynamics in China from 1990 to 2019. Earth Syst. Sci. Data 2021, 13, 3907–3925. [Google Scholar] [CrossRef]
- Peng, S.; Ding, Y.; Liu, W.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Mu, Q.; Zhao, M.; Kimball, J.S.; McDowell, N.G.; Running, S.W. A Remotely Sensed Global Terrestrial Drought Severity Index. Bull. Am. Meteorol. Soc. 2013, 94, 83–98. [Google Scholar] [CrossRef]
- Khan, R.; Gilani, H. Global Drought Monitoring with Drought Severity Index (DSI) Using Google Earth Engine. Theor. Appl. Climatol. 2021, 146, 411–427. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef]
- Zhang, X.; Yamaguchi, Y. Characterization and Evaluation of MODIS Derived Drought Severity Index (DSI) for Monitoring the 2009/2010 Drought over Southwestern China. Nat. Hazards 2014, 74, 2129–2145. [Google Scholar] [CrossRef]
- Woodwell, G.M.; Whittaker, R.H.; Reiners, W.A.; Likens, G.E.; Delwiche, C.C.; Botkin, D.B. The Biota and the World Carbon Budget. Science 1978, 199, 141–146. [Google Scholar] [CrossRef]
- Pei, Z.-Y.; Ouyang, H.; Zhou, C.-P.; Xu, X.-L. Carbon Balance in an Alpine Steppe in the Qinghai-Tibet Plateau. J. Integr. Plant Biol. 2009, 51, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, W.; Zhang, Y. The Change in Net Ecosystem Productivity and Its Driving Mechanism in a Mountain Ecosystem of Arid Regions, Northwest China. Remote Sens. 2022, 14, 4046. [Google Scholar] [CrossRef]
- Li, X.; Lin, G.; Jiang, D.; Fu, J.; Wang, Y. Spatiotemporal Evolution Characteristics and the Climatic Response of Carbon Sources and Sinks in the Chinese Grassland Ecosystem from 2010 to 2020. Sustainability 2022, 14, 8461. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, J.; Yu, P.; Li, J.; Kang, Y.; Zhang, B. Response of Ecosystem Gross Primary Productivity to Drought in Northern China Based on Multi-Source Remote Sensing Data. J. Hydrol. 2023, 616, 128808. [Google Scholar] [CrossRef]
- Zhang, J.; Ren, Z. Spatiotemporal Characteristics of Evapotranspiration Based on MOD16 in the Hanjiang River Basin. Sci. Geogr. Sin. 2017, 37, 274–282. [Google Scholar] [CrossRef]
- Zhang, K.; Zhu, C.; Ma, X.; Zhang, X.; Yang, D.; Shao, Y. Spatiotemporal Variation Characteristics and Dynamic Persistence Analysis of Carbon Sources/Sinks in the Yellow River Basin. Remote Sens. 2023, 15, 323. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Zhang, S.; Wang, J.; Bai, Y.; Yao, F.; Guo, H. The Potential of Remote Sensing-Based Models on Global Water-Use Efficiency Estimation: An Evaluation and Intercomparison of an Ecosystem Model (BESS) and Algorithm (MODIS) Using Site Level and Upscaled Eddy Covariance Data. Agric. For. Meteorol. 2020, 287, 107959. [Google Scholar] [CrossRef]
- Tang, X.; Ma, M.; Ding, Z.; Xu, X.; Yao, L.; Huang, X.; Gu, Q.; Song, L. Remotely Monitoring Ecosystem Water Use Efficiency of Grassland and Cropland in China’s Arid and Semi-Arid Regions with MODIS Data. REMOTE Sens. 2017, 9, 616. [Google Scholar] [CrossRef]
- Gang, C.; Zhang, Y.; Guo, L.; Gao, X.; Peng, S.; Chen, M.; Wen, Z. Drought-Induced Carbon and Water Use Efficiency Responses in Dryland Vegetation of Northern China. Front. Plant Sci. 2019, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Gong, F.; Zeng, Y.; Ma, L.; Qiao, C.; Wu, H. Carbon Use Efficiency of Terrestrial Ecosystems in Desert/Grassland Biome Transition Zone: A Case in Ningxia Province, Northwest China. Ecol. Indic. 2021, 120, 106971. [Google Scholar] [CrossRef]
- He, Y.; Piao, S.; Li, X.; Chen, A.; Qin, D. Global Patterns of Vegetation Carbon Use Efficiency and Their Climate Drivers Deduced from MODIS Satellite Data and Process-Based Models. Agric. For. Meteorol. 2018, 256–257, 150–158. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, B.; Chen, J.; Zhou, J.; He, Q.-Q.; Yu, H. Response of Vegetation Productivity to Greening and Drought in the Loess Plateau Based on VIs and SIF. Forests 2024, 15, 339. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Zheng, Z.; Shen, W.; Li, Y.; Rong, P.; Qin, Y. Trends in Drought and Effects on Carbon Sequestration over the Chinese Mainland. Sci. Total Environ. 2023, 856, 159075. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.O.; Wang, Y. Recent Trends in Satellite Vegetation Index Observations Indicate Decreasing Vegetation Biomass in the Southeastern Saline Everglades Wetlands. Wetlands 2014, 34, 67–77. [Google Scholar] [CrossRef]
- Li, Y.; Qin, Y.; Rong, P. Evolution of Potential Evapotranspiration and Its Sensitivity to Climate Change Based on the Thornthwaite, Hargreaves, and Penman–Monteith Equation in Environmental Sensitive Areas of China. Atmos. Res. 2022, 273, 106178. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, J.; Duan, X.; Ma, P.; Lu, G.; Zhu, B.; Liu, X.; Yue, P.; Wang, Y.; Liu, W. The Eastward Expansion of the Climate Humidification Trend in Northwest China and the Synergistic Influences on the Circulation Mechanism. Clim. Dyn. 2022, 59, 2481–2497. [Google Scholar] [CrossRef]
- Zhang, X.; Susan Moran, M.; Zhao, X.; Liu, S.; Zhou, T.; Ponce-Campos, G.E.; Liu, F. Impact of Prolonged Drought on Rainfall Use Efficiency Using MODIS Data across China in the Early 21st Century. Remote Sens. Environ. 2014, 150, 188–197. [Google Scholar] [CrossRef]
- SONG, L.; ZHANG, C. Changing Features of Precipitation over Northwest China During the 20th Century. J. Glaciol. Geocryol. 2003, 25, 143–148. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, J.H.; Wang, P.L.; Yu, H.P.; Yue, P.; Liu, X.Y.; Lin, J.J.; Duan, X.Y.; Zhu, B.; Yan, X.Y. Progress and prospect on climate warming and humidification in Northwest China. Chin. Sci. Bull. 2023, 68, 1814–1828. (In Chinese) [Google Scholar] [CrossRef]
- Yang, L.; Feng, Q.; Adamowski, J.F.; Yin, Z.; Wen, X.; Wu, M.; Jia, B.; Hao, Q. Spatio-Temporal Variation of Reference Evapotranspiration in Northwest China Based on CORDEX-EA. Atmos. Res. 2020, 238, 104868. [Google Scholar] [CrossRef]
- Huo, Z.; Dai, X.; Feng, S.; Kang, S.; Huang, G. Effect of Climate Change on Reference Evapotranspiration and Aridity Index in Arid Region of China. J. Hydrol. 2013, 492, 24–34. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Shen, Y.; Liu, Y.; Zhang, S. Analysis of Changing Pan Evaporation in the Arid Region of Northwest China. Water Resour. Res. 2013, 49, 2205–2212. [Google Scholar] [CrossRef]
- Xing, W.; Wang, W.; Shao, Q.; Yu, Z.; Yang, T.; Fu, J. Periodic Fluctuation of Reference Evapotranspiration during the Past Five Decades: Does Evaporation Paradox Really Exist in China? Sci. Rep. 2016, 6, 39503. [Google Scholar] [CrossRef]
- Yang, Y.; Roderick, M.L.; Guo, H.; Miralles, D.G.; Zhang, L.; Fatichi, S.; Luo, X.; Zhang, Y.; McVicar, T.R.; Tu, Z.; et al. Evapotranspiration on a Greening Earth. Nat. Rev. Earth Environ. 2023, 4, 626–641. [Google Scholar] [CrossRef]
- Kurganova, I.; Teepe, R.; Loftfield, N. Influence of Freeze-Thaw Events on Carbon Dioxide Emission from Soils at Different Moisture and Land Use. Carbon Balance Manag. 2007, 2, 2. [Google Scholar] [CrossRef]
- Wang, X.; Li, F.; Zhao, S. Freeze-Thaw Regime Effects on Soil CO2 Emission: A Review. Chin. J. Soil Sci. 2022, 53, 728–737. [Google Scholar]
- Goulden, M.L.; Wofsy, S.C.; Harden, J.W.; Trumbore, S.E.; Crill, P.M.; Gower, S.T.; Fries, T.; Daube, B.C.; Fan, S.-M.; Sutton, D.J.; et al. Sensitivity of Boreal Forest Carbon Balance to Soil Thaw. Science 1998, 279, 214–217. [Google Scholar] [CrossRef]
- Mikan, C.J.; Schimel, J.P.; Doyle, A.P. Temperature Controls of Microbial Respiration in Arctic Tundra Soils above and below Freezing. Soil Biol. Biochem. 2002, 34, 1785–1795. [Google Scholar] [CrossRef]
- Edwards, A.C.; Creasey, J.; Cresser, M.S. Soil Freezing Effects on Upland Stream Solute Chemistry. Water Res. 1986, 20, 831–834. [Google Scholar] [CrossRef]
- Bo, L.; Li, Z.; Li, P.; Xu, G.; Xiao, L.; Ma, B. Soil Freeze-Thaw and Water Transport Characteristics Under Different Vegetation Types in Seasonal Freeze-Thaw Areas of the Loess Plateau. Front. Earth Sci. 2021, 9, 704901. [Google Scholar] [CrossRef]
- Xiao, L.; Yao, K.; Li, P.; Liu, Y.; Zhang, Y. Effects of Freeze-Thaw Cycles and Initial Soil Moisture Content on Soil Aggregate Stability in Natural Grassland and Chinese Pine Forest on the Loess Plateau of China. J. Soils Sediments 2020, 20, 1222–1230. [Google Scholar] [CrossRef]
- Liu, S.; Wu, J.; Li, G.; Yang, C.; Yuan, J.; Xie, M. Seasonal Freeze-Thaw Characteristics of Soil Carbon Pools under Different Vegetation Restoration Types on the Longzhong Loess Plateau. Front. Ecol. Evol. 2022, 10, 1019627. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, B.; Yang, J.H.; Wang, D.W. New Characteristics about the Climate Humidification Trend in Northwest China. Chin. Sci. Bull. 2021, 66, 3757–3771. (In Chinese) [Google Scholar] [CrossRef]
- Han, T.; Guo, X.; Zhou, B.; Hao, X. Recent Changes in Heavy Precipitation Events in Northern Central China and Associated Atmospheric Circulation. Asia-Pac. J. Atmos. Sci. 2021, 57, 301–310. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Li, K.; Zhang, F.; Yang, K. Change Characteristics of Precipitation in Northwest China from 1961 to 2018. Chin. J. Atmos. Sci. 2021, 45, 713–724. [Google Scholar] [CrossRef]
- Li, X.; Zhang, K.; Bao, H.; Zhang, H. Climatology and Changes in Hourly Precipitation Extremes over China during 1970–2018. Sci. Total Environ. 2022, 839, 156297. [Google Scholar] [CrossRef]
- Huang, H.; Cui, H.; Singh, V.P. Clustering Daily Extreme Precipitation Patterns in China. Water 2023, 15, 3651. [Google Scholar] [CrossRef]
- Yang, M.; Zhao, Y.; Wang, T. Extreme Precipitation Changes and Its Effects on Fractional Vegetation Cover in the Loess Plateau. Sci. Technol. Eng. 2024, 24, 3086–3095. [Google Scholar] [CrossRef]
- PAN, J.; LI, Z. Temporal-Spatial Change of Vegetation Net Primary Productivity in the Arid Region of Northwest China during 2001 and 2012. Chin. J. Ecol. 2015, 34, 3333. [Google Scholar] [CrossRef]
- Wei, X.; Yang, J.; Luo, P.; Lin, L.; Lin, K.; Guan, J. Assessment of the Variation and Influencing Factors of Vegetation NPP and Carbon Sink Capacity under Different Natural Conditions. Ecol. Indic. 2022, 138, 108834. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, Z.; Meng, Y.; Pan, Y.; Guo, Z.; Zhang, Y. Spatio-Temporal Variation in Net Primary Productivity of Different Vegetation Types and Its Influencing Factors Exploration in Southwest China. Environ. Sci. 2024, 45, 262–274. [Google Scholar] [CrossRef]
- Tian, H.; Lu, C.; Chen, G.; Xu, X.; Liu, M.; Ren, W.; Tao, B.; Sun, G.; Pan, S.; Liu, J. Climate and Land Use Controls over Terrestrial Water Use Efficiency in Monsoon Asia. Ecohydrology 2011, 4, 322–340. [Google Scholar] [CrossRef]
- Liu, Y.; Xiao, J.; Ju, W.; Xu, K.; Zhou, Y.; Zhao, Y. Recent Trends in Vegetation Greenness in China Significantly Altered Annual Evapotranspiration and Water Yield. Environ. Res. Lett. 2016, 11, 094010. [Google Scholar] [CrossRef]
- Shao, M.; Jia, X.; Wang, Y.; Zhu, Y. A Review of Studies on Dried Soil Layers in the Loess Plateau. Adv. Earth Sci. 2016, 31, 14–22. [Google Scholar]
- Keenan, T.F.; Luo, X.; Stocker, B.D.; De Kauwe, M.G.; Medlyn, B.E.; Prentice, I.C.; Smith, N.G.; Terrer, C.; Wang, H.; Zhang, Y.; et al. A Constraint on Historic Growth in Global Photosynthesis Due to Rising CO2. Nat. Clim. Chang. 2023, 13, 1376–1381. [Google Scholar] [CrossRef]
- Ahlström, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The Dominant Role of Semi-Arid Ecosystems in the Trend and Variability of the Land CO2 Sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhong, X.; Deng, S.; Xu, H. Assessment of the Impact of LUCC on NPP and Its Influencing Factors in the Yangtze River Basin, China. Catena 2021, 206, 105542. [Google Scholar] [CrossRef]
- Oishi, A.C.; Miniat, C.F.; Novick, K.A.; Brantley, S.T.; Vose, J.M.; Walker, J.T. Warmer Temperatures Reduce Net Carbon Uptake, but Do Not Affect Water Use, in a Mature Southern Appalachian Forest. Agric. For. Meteorol. 2018, 252, 269–282. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, J.; Wang, S.; Chen, B.; Li, Z. Detection and Attribution of Positive Net Ecosystem Productivity Extremes in China’s Terrestrial Ecosystems during 2000–2016. Ecol. Indic. 2021, 132, 108323. [Google Scholar] [CrossRef]
- Fang, X.; Zhang, C.; Wang, Q.; Chen, X.; Ding, J.; Karamage, F. Isolating and Quantifying the Effects of Climate and CO2 Changes (1980–2014) on the Net Primary Productivity in Arid and Semiarid China. Forests 2017, 8, 60. [Google Scholar] [CrossRef]
- Liang, W.; Yang, Y.; Fan, D.; Guan, H.; Zhang, T.; Long, D.; Zhou, Y.; Bai, D. Analysis of Spatial and Temporal Patterns of Net Primary Production and Their Climate Controls in China from 1982 to 2010. Agric. For. Meteorol. 2015, 204, 22–36. [Google Scholar] [CrossRef]
- Feng, X.; Fu, B.; Lu, N.; Zeng, Y.; Wu, B. How Ecological Restoration Alters Ecosystem Services: An Analysis of Carbon Sequestration in China’s Loess Plateau. Sci. Rep. 2013, 3, 2846. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Wu, Y.; Yao, Y.; Sun, K.; Zhang, X.; Winowiecki, L.; Vågen, T.-G.; Xu, J.; Qiu, L.; Sun, P.; et al. Predicting the Climate Change Impacts on Water-Carbon Coupling Cycles for a Loess Hilly-Gully Watershed. J. Hydrol. 2020, 581, 124388. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, X.; Yu, G.; He, N.; Wang, Q.; Tian, J. Water Use Efficiency Threshold for Terrestrial Ecosystem Carbon Sequestration in China under Afforestation. Agric. For. Meteorol. 2014, 195–196, 32–37. [Google Scholar] [CrossRef]
- Sun, B.; Zhao, H.; Wang, X. Effects of Drought on Net Primary Productivity: Roles of Temperature, Drought Intensity, and Duration. Chin. Geogr. Sci. 2016, 26, 270–282. [Google Scholar] [CrossRef]
- Pei, F.; Li, X.; Liu, X.; Lao, C. Assessing the Impacts of Droughts on Net Primary Productivity in China. J. Environ. Manag. 2013, 114, 362–371. [Google Scholar] [CrossRef]
- Turner, D.P.; Ritts, W.D.; Cohen, W.B.; Gower, S.T.; Running, S.W.; Zhao, M.; Costa, M.H.; Kirschbaum, A.A.; Ham, J.M.; Saleska, S.R.; et al. Evaluation of MODIS NPP and GPP Products across Multiple Biomes. Remote Sens. Environ. 2006, 102, 282–292. [Google Scholar] [CrossRef]
- Wang, Y.; Li, R.; Hu, J.; Wang, X.; Kabeja, C.; Min, Q.; Wang, Y. Evaluations of MODIS and Microwave Based Satellite Evapotranspiration Products under Varied Cloud Conditions over East Asia Forests. Remote Sens. Environ. 2021, 264, 112606. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Source | Time Resolution | Spatial Resolution | Download Site |
|---|---|---|---|---|
| Land cover | National Cryosphere Desert Data Centre | Annual | 30 m | http://www.ncdc.ac.cn/portal/ (accessed on 1 April 2024) |
| PRE and TEM | National Tibetan Plateau Science Data Center | Monthly | 1 km | http://data.tpdc.ac.cn/ (accessed on 1 April 2024) |
| ETa and PET | MOD16A2 | 8 days | 500 m | https://earthengine.google.com/ (accessed on 1 April 2024) |
| NDVI | MOD13A1 | 16 days | 500 m | |
| GPP | MOD17A2H | 8 days | 500 m | |
| NPP | MOD17A2H | Annual | 500 m |
| Form | Degree (Level or Extent) | DSI |
|---|---|---|
| D5 | Extreme drought | <−1.50 |
| D4 | Severe drought | −1.50–−1.20 |
| D3 | Moderate drought | −1.19–−0.90 |
| D2 | Mild drought | −0.89–−0.60 |
| D1 | Incipient drought | −0.59–−0.30 |
| WD | Near normal | −0.29–0.29 |
| W1 | Incipient wet | 0.30–0.59 |
| W2 | Slightly wet | 0.60–0.89 |
| W3 | Moderately wet | 0.90–1.19 |
| W4 | Very wet | 1.20–1.50 |
| W5 | Extremely wet | >1.50 |
| Study | Model Used | Location | Study Period | Key Results |
|---|---|---|---|---|
| Zhang et al. [48] | NEP = NPP − RH | Yellow River Basin | 2000–2020 |
|
| Wang et al. [44] | NEP = NPP − RH | Qilian Mountains (QLM) | 2000–2020 |
|
| Li et al. [45] | Vegetation production model; NEP = NPP − RH | China Grassland | 2010–2020 |
|
| Zhang et al. [8] | CASA model; NEP = NPP − RH | Central Asia | 2001–2019 |
|
| Yang et al. [49] | WUE = GPP/ETa; BESS model | Global | 2000–2014 |
|
| Tang et al. [50] | WUE = GPP/ETa | Heihe River Basin | 2013–2015 |
|
| Gang et al. [51] | WUE = NPP/ETa; CUE = NPP/GPP | Northern China | 2000–2011 |
|
| Du et al. [52] | CUE = NPP/ETa | Ningxia | 2001–2017 |
|
| He et al. [53] | CUE = NPP/GPP; Process-based carbon cycle models | Global | 2000–2012 |
|
| LUCC (2001–2022) | Area/km2 | Percentage of Change | Cumulative Percentage | Change in NEP/Tg (1 Tg = 1012 g) |
|---|---|---|---|---|
| Cropland–Grassland | 21,592.86 | 31.91% | 31.91% | 3.25 |
| Grassland–Cropland | 18,064.26 | 26.69% | 58.60% | 5.15 |
| Grassland–Forest | 10,166.11 | 15.02% | 73.63% | 4.54 |
| Cropland–Impervious | 5306.00 | 7.84% | 81.47% | −0.40 |
| Cropland–Forest | 3824.11 | 5.65% | 87.12% | 1.49 |
| Barren–Grassland | 3340.92 | 4.94% | 92.05% | 0.754 |
| Forest–Cropland | 1407.19 | 2.08% | 94.13% | 0.096 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, X.; Zhang, B.; He, Q.-Q.; Shao, Z.-L.; Yu, H.; Zhang, X.-Y. Spatial–Temporal Variations in the Climate, Net Ecosystem Productivity, and Efficiency of Water and Carbon Use in the Middle Reaches of the Yellow River. Remote Sens. 2024, 16, 3312. https://doi.org/10.3390/rs16173312
Hou X, Zhang B, He Q-Q, Shao Z-L, Yu H, Zhang X-Y. Spatial–Temporal Variations in the Climate, Net Ecosystem Productivity, and Efficiency of Water and Carbon Use in the Middle Reaches of the Yellow River. Remote Sensing. 2024; 16(17):3312. https://doi.org/10.3390/rs16173312
Chicago/Turabian StyleHou, Xiao, Bo Zhang, Qian-Qian He, Zhuan-Ling Shao, Hui Yu, and Xue-Ying Zhang. 2024. "Spatial–Temporal Variations in the Climate, Net Ecosystem Productivity, and Efficiency of Water and Carbon Use in the Middle Reaches of the Yellow River" Remote Sensing 16, no. 17: 3312. https://doi.org/10.3390/rs16173312
APA StyleHou, X., Zhang, B., He, Q.-Q., Shao, Z.-L., Yu, H., & Zhang, X.-Y. (2024). Spatial–Temporal Variations in the Climate, Net Ecosystem Productivity, and Efficiency of Water and Carbon Use in the Middle Reaches of the Yellow River. Remote Sensing, 16(17), 3312. https://doi.org/10.3390/rs16173312