
Enhancing Accuracy in Historical Forest Vegetation Mapping in Yunnan with Phenological Features, and Climatic and Elevation Variables
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Image Data Pre-Processing
2.3. Ground Reference Data
- (1)
- Random sampling: A total of 445 forest communities, each 20 m × 50 m in size, were sampled using the random sampling method. We used ArcGIS software (version 10.0) and the provincial boundary shapefile data of Yunnan were used to randomly allocate 445 forest sampling sites. During field visits, the nearest forest patches to the allocated forest sites were surveyed. Data collected from 2010 to 2015 included plant community compositions, geographic co-ordinates, elevation, and forest ages.
- (2)
- Stratified sampling: From 2018 to 2021, 348 forest sampling sites were collected using the stratified sampling method based on the floristic regions of Yunnan [49]. First, we calculated the area percentage of each floristic region in Yunnan and multiplied it by 337 to determine the number of sampling sites for each region. We then randomly allocated the forest sampling sites within each floristic region using ArcGIS software (version 10.5). High spatial resolution satellite data (e.g., Google Earth) were used to ensure each forest sampling site within the closest forest patch. At these sites, we investigated plant community compositions, geographic co-ordinates, and elevation.
- (3)
2.4. Separability Analysis of Data Inputs
2.5. Classification and Validation
2.6. Historical Mapping of Forest Vegetation Types
3. Results
3.1. Phenological Features across Nine Forest Vegetation
3.2. The Separability of Forest Vegetation
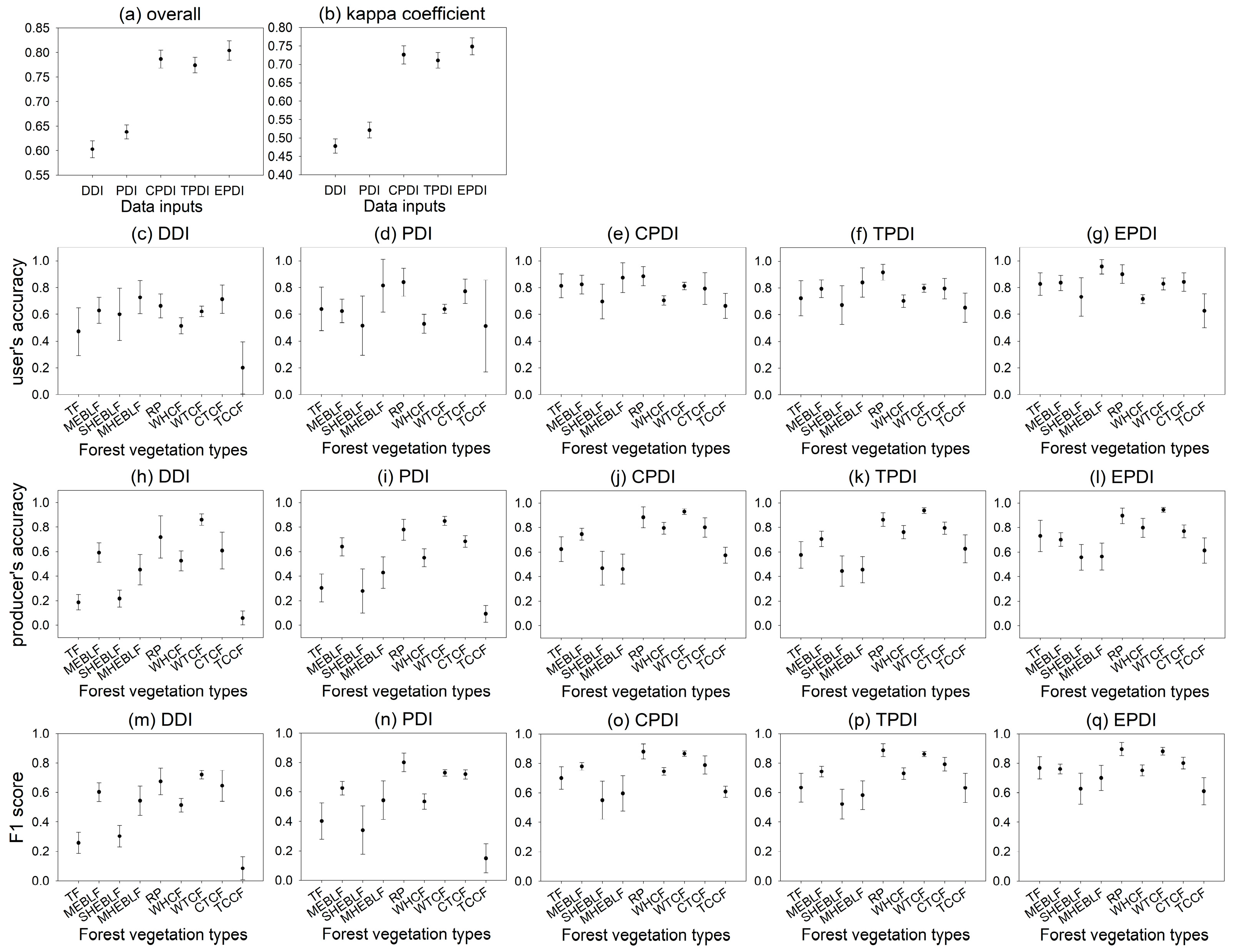
3.3. Mapping Accuracy Comparison
3.4. Characteristic of Historical Forest Vegetation Maps in Yunnan
4. Discussion
4.1. Spectral and Phenological Features in Differentiating Forest Vegetation Types
4.2. The Role of Climatic and Topographic Information in Enhancing Forest Mapping Accuracy
4.3. Changes in Forest Area in Yunnan
4.4. Applications and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- de Groot, R.; Brander, L.; van der Ploeg, S.; Costanza, R.; Bernard, F.; Braat, L.; Christie, M.; Crossman, N.; Ghermandi, A.; Hein, L.; et al. Global estimates of the value of ecosystems and their services in monetary units. Ecosyst. Serv. 2012, 1, 50–61. [Google Scholar] [CrossRef]
- Yang, W.; Wang, Y.; Webb, A.A.; Li, Z.; Tian, X.; Han, Z.; Wang, S.; Yu, P. Influence of climatic and geographic factors on the spatial distribution of Qinghai spruce forests in the dryland Qilian Mountains of Northwest China. Sci. Total Environ. 2018, 612, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Zischg, A.P.; Frehner, M.; Gubelmann, P.; Augustin, S.; Brang, P.; Huber, B. Participatory modelling of upward shifts of altitudinal vegetation belts for assessing site type transformation in Swiss forests due to climate change. Appl. Veg. Sci. 2021, 24, e12621. [Google Scholar] [CrossRef]
- Zhou, Y.; Yi, Y.-J.; Liu, H.-X.; Tang, C.-H.; Zhu, Y.-L.; Zhang, S.-H. Effect of geomorphologic features and climate change on vegetation distribution in the arid hot valleys of Jinsha River, Southwest China. J. Mt. Sci. 2022, 19, 2874–2885. [Google Scholar] [CrossRef]
- Hua, F.; Xu, J.; Wilcove, D.S. A New Opportunity to Recover Native Forests in China. Conserv. Lett. 2018, 11, e12396. [Google Scholar] [CrossRef]
- Hua, F.; Bruijnzeel, L.A.; Meli, P.; Martin, P.A.; Zhang, J.; Nakagawa, S.; Miao, X.; Wang, W.; McEvoy, C.; Peña-Arancibia, J.L.; et al. The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 2022, 376, 839–844. [Google Scholar] [CrossRef]
- Yang, J.; Zhai, D.-L.; Fang, Z.; Alatalo, J.M.; Yao, Z.; Yang, W.; Su, Y.; Bai, Y.; Zhao, G.; Xu, J. Changes in and driving forces of ecosystem services in tropical southwestern China. Ecol. Indic. 2023, 149, 110180. [Google Scholar] [CrossRef]
- Clark, M.L. Comparison of multi-seasonal Landsat 8, Sentinel-2 and hyperspectral images for mapping forest alliances in Northern California. ISPRS-J. Photogramm. Remote Sens. 2020, 159, 26–40. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, L.; Zhao, Y.; Wang, Y.; Wei, D.; Wu, X.; Ma, X. Recent advances in using Chinese Earth observation satellites for remote sensing of vegetation. ISPRS-J. Photogramm. Remote Sens. 2023, 195, 393–407. [Google Scholar] [CrossRef]
- Ghosh, A.; Fassnacht, F.E.; Joshi, P.K.; Koch, B. A framework for mapping tree species combining hyperspectral and LiDAR data: Role of selected classifiers and sensor across three spatial scales. Int. J. Appl. Earth Obs. Geoinf. 2014, 26, 49–63. [Google Scholar] [CrossRef]
- Mäyrä, J.; Keski-Saari, S.; Kivinen, S.; Tanhuanpää, T.; Hurskainen, P.; Kullberg, P.; Poikolainen, L.; Viinikka, A.; Tuominen, S.; Kumpula, T.; et al. Tree species classification from airborne hyperspectral and LiDAR data using 3D convolutional neural networks. Remote Sens. Environ. 2021, 256, 112322. [Google Scholar] [CrossRef]
- Iglseder, A.; Immitzer, M.; Dostálová, A.; Kasper, A.; Pfeifer, N.; Bauerhansl, C.; Schöttl, S.; Hollaus, M. The potential of combining satellite and airborne remote sensing data for habitat classification and monitoring in forest landscapes. Int. J. Appl. Earth Obs. Geoinf. 2023, 117, 103131. [Google Scholar] [CrossRef]
- Hu, T.; Zhang, Y.; Su, Y.; Zheng, Y.; Lin, G.; Guo, Q. Mapping the Global Mangrove Forest Aboveground Biomass Using Multisource Remote Sensing Data. Remote Sens. 2020, 12, 1690. [Google Scholar] [CrossRef]
- Wulder, M.A.; Masek, J.G.; Cohen, W.B.; Loveland, T.R.; Woodcock, C.E. Opening the archive: How free data has enabled the science and monitoring promise of Landsat. Remote Sens. Environ. 2012, 122, 2–10. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A review of vegetation phenological metrics extraction using time-series, multispectral satellite data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Xue, Z.; Du, P.; Feng, L. Phenology-Driven Land Cover Classification and Trend Analysis Based on Long-term Remote Sensing Image Series. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2014, 7, 1142–1156. [Google Scholar] [CrossRef]
- Zhai, D.; Dong, J.; Cadisch, G.; Wang, M.; Kou, W.; Xu, J.; Xiao, X.; Abbas, S. Comparison of Pixel- and Object-Based Approaches in Phenology-Based Rubber Plantation Mapping in Fragmented Landscapes. Remote Sens. 2018, 10, 44. [Google Scholar] [CrossRef]
- Yang, J.; Xu, J.; Zhai, D.-L. Integrating Phenological and Geographical Information with Artificial Intelligence Algorithm to Map Rubber Plantations in Xishuangbanna. Remote Sens. 2021, 13, 2793. [Google Scholar] [CrossRef]
- Senf, C.; Laštovička, J.; Okujeni, A.; Heurich, M.; van der Linden, S. A generalized regression-based unmixing model for mapping forest cover fractions throughout three decades of Landsat data. Remote Sens. Environ. 2020, 240, 111691. [Google Scholar] [CrossRef]
- Guo, J.; Du, S. A Multicenter Soft Supervised Classification Method for Modeling Spectral Diversity in Multispectral Remote Sensing Data. IEEE Trans. Geosci. Remote Sens. 2022, 60, 5605517. [Google Scholar] [CrossRef]
- Woodward, F.I.; Lomas, M.R.; Kelly, C.K. Global Climate and the Distribution of Plant Biomes. Philos. Trans. R. Soc. B-Biol. Sci. 2004, 359, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Barga, S.C.; Dilts, T.E.; Leger, E.A. Contrasting climate niches among co-occurring subdominant forbs of the sagebrush steppe. Divers. Distrib. 2018, 24, 1291–1307. [Google Scholar] [CrossRef]
- Sun, G.-Q.; Li, L.; Li, J.; Liu, C.; Wu, Y.-P.; Gao, S.; Wang, Z.; Feng, G.-L. Impacts of climate change on vegetation pattern: Mathematical modeling and data analysis. Phys. Life Rev. 2022, 43, 239–270. [Google Scholar] [CrossRef]
- Li, F.-F.; Lu, H.-L.; Wang, G.-Q.; Yao, Z.-Y.; Li, Q.; Qiu, J. Zoning of precipitation regimes on the Qinghai–Tibet Plateau and its surrounding areas responded by the vegetation distribution. Sci. Total Environ. 2022, 838, 155844. [Google Scholar] [CrossRef]
- Wu, Z. Vegetation of Yunnan; Science Press: Beijing, China, 1987; pp. 81–417. [Google Scholar]
- Fang, J.; Song, Y.; Liu, H.; Piao, S. Vegetation-climate relationship and its application in the division of vegetation zone in China. Acta Bot. Sin. 2002, 44, 1105–1122. [Google Scholar]
- Fang, J.-y.; Yoda, K. Climate and vegetation in China II. Distribution of main vegetation types and thermal climate. Ecol. Res. 1989, 4, 71–83. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Li, C.-F.; Chen, T.-Y.; Hsieh, C.-F.; Wang, G.; Wang, T.; Hu, J.-M. Climate-based approach for modeling the distribution of montane forest vegetation in Taiwan. Appl. Veg. Sci. 2020, 23, 239–253. [Google Scholar] [CrossRef]
- Shao, J.a.; Li, Y.; Ni, J. The characteristics of temperature variability with terrain, latitude and longitude in Sichuan-Chongqing Region. J. Geogr. Sci. 2012, 22, 223–244. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, D. Accurate mapping of forest types using dense seasonal Landsat time-series. ISPRS-J. Photogramm. Remote Sens. 2014, 96, 1–11. [Google Scholar] [CrossRef]
- Hościło, A.; Lewandowska, A. Mapping Forest Type and Tree Species on a Regional Scale Using Multi-Temporal Sentinel-2 Data. Remote Sens. 2019, 11, 929. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Okolie, C.J.; Smit, J.L. A systematic review and meta-analysis of Digital elevation model (DEM) fusion: Pre-processing, methods and applications. ISPRS-J. Photogramm. Remote Sens. 2022, 188, 1–29. [Google Scholar] [CrossRef]
- Riley, K.L.; Grenfell, I.C.; Finney, M.A. Mapping forest vegetation for the western United States using modified random forests imputation of FIA forest plots. Ecosphere 2016, 7, e01472. [Google Scholar] [CrossRef]
- Zhou, G.; Ren, H.; Liu, T.; Zhou, L.; Ji, Y.; Song, X.; Lv, X. A new regional vegetation mapping method based on terrain-climate-remote sensing and its application on the Qinghai-Xizang Plateau. Sci. China-Earth Sci. 2023, 66, 237–246. [Google Scholar] [CrossRef]
- Wu, F.; Ren, H.; Zhou, G. The 30 m vegetation maps from 1990 to 2020 in the Tibetan Plateau. Sci. Data 2024, 11, 804. [Google Scholar] [CrossRef]
- Reddy, C.S.; Jha, C.S.; Diwakar, P.G.; Dadhwal, V.K. Nationwide classification of forest types of India using remote sensing and GIS. Environ. Monit. Assess. 2015, 187, 777. [Google Scholar] [CrossRef]
- Su, Y.; Guo, Q.; Hu, T.; Guan, H.; Jin, S.; An, S.; Chen, X.; Guo, K.; Hao, Z.; Hu, Y.; et al. An updated Vegetation Map of China (1:1,000,000). Sci. Bull. 2020, 65, 1125–1136. [Google Scholar] [CrossRef]
- Yang, Y.; Tian, K.; Hao, J.; Pei, S.; Yang, Y. Biodiversity and biodiversity conservation in Yunnan, China. Biodivers. Conserv. 2004, 13, 813–826. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Zhang, Y.; Wan, C. Carbon sequestration potential of the stands under the Grain for Green Program in Yunnan Province, China. For. Ecol. Manag. 2009, 258, 199–206. [Google Scholar] [CrossRef]
- Zhou, R.; Li, W.; Zhang, Y.; Peng, M.; Wang, C.; Sha, L.; Liu, Y.; Song, Q.; Fei, X.; Jin, Y.; et al. Responses of the Carbon Storage and Sequestration Potential of Forest Vegetation to Temperature Increases in Yunnan Province, SW China. Forests 2018, 9, 227. [Google Scholar] [CrossRef]
- Peng, J.; Yang, Y.; Liu, Y.; Hu, Y.n.; Du, Y.; Meersmans, J.; Qiu, S. Linking ecosystem services and circuit theory to identify ecological security patterns. Sci. Total Environ. 2018, 644, 781–790. [Google Scholar] [CrossRef]
- Zhu, H.; Tan, Y. Flora and Vegetation of Yunnan, Southwestern China: Diversity, Origin and Evolution. Diversity 2022, 14, 340. [Google Scholar] [CrossRef]
- Hua, Z. Biogeographical Divergence of the Flora of Yunnan, Southwestern China Initiated by the Uplift of Himalaya and Extrusion of Indochina Block. PLoS ONE 2012, 7, e45601. [Google Scholar] [CrossRef]
- Hua, Z. Advances in Biogeography of the Tropical Rain Forest in Southern Yunnan, Southwestern China. Trop. Conserv. Sci. 2008, 1, 34–42. [Google Scholar] [CrossRef]
- Zhang, M.-G.; Zhou, Z.-K.; Chen, W.-Y.; Slik, J.W.F.; Cannon, C.H.; Raes, N. Using species distribution modeling to improve conservation and land use planning of Yunnan, China. Biol. Conserv. 2012, 153, 257–264. [Google Scholar] [CrossRef]
- Wu, Q. geemap: A Python package for interactive mapping with Google Earth Engine. J. Open Source Softw. 2020, 5, 2305. [Google Scholar] [CrossRef]
- Wang, Y.; Hollingsworth, P.M.; Zhai, D.; West, C.D.; Green, J.M.H.; Chen, H.; Hurni, K.; Su, Y.; Warren-Thomas, E.; Xu, J.; et al. High-resolution maps show that rubber causes substantial deforestation. Nature 2023, 623, 340–346. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Defries, R.S.; Townshend, J.R.G. NDVI-derived land cover classifications at a global scale. Int. J. Remote Sens. 1994, 15, 3567–3586. [Google Scholar] [CrossRef]
- Xiao, X.; Boles, S.; Liu, J.; Zhuang, D.; Frolking, S.; Li, C.; Salas, W.; Moore, B. Mapping paddy rice agriculture in southern China using multi-temporal MODIS images. Remote Sens. Environ. 2005, 95, 480–492. [Google Scholar] [CrossRef]
- Yang, R.-Q.; Fu, P.-L.; Fan, Z.-X.; Panthi, S.; Gao, J.; Niu, Y.; Li, Z.-S.; Bräuning, A. Growth-climate sensitivity of two pine species shows species-specific changes along temperature and moisture gradients in southwest China. Agric. For. Meteorol. 2022, 318, 108907. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- Yang, J.; Xu, J.; Zhou, Y.; Zhai, D.; Chen, H.; Li, Q.; Zhao, G. Paddy Rice Phenological Mapping throughout 30-Years Satellite Images in the Honghe Hani Rice Terraces. Remote Sens. 2023, 15, 2398. [Google Scholar] [CrossRef]
- Jung, R.; Ehlers, M. Comparison of two feature selection methods for the separability analysis of intertidal sediments with spectrometric datasets in the German Wadden Sea. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 175–191. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS-J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Ouyang, Z.; Tam, C.; Chen, X. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proc. Natl. Acad. Sci. USA 2008, 105, 9477–9482. [Google Scholar] [CrossRef]
- Zhai, D.-L.; Xu, J.-C. The legacy effects of rubber defoliation period on the refoliation phenology, leaf disease, and latex yield. Plant Divers. 2023, 45, 98–103. [Google Scholar] [CrossRef]
- Chen, B.; Li, X.; Xiao, X.; Zhao, B.; Dong, J.; Kou, W.; Qin, Y.; Yang, C.; Wu, Z.; Sun, R.; et al. Mapping tropical forests and deciduous rubber plantations in Hainan Island, China by integrating PALSAR 25-m and multi-temporal Landsat images. Int. J. Appl. Earth Obs. Geoinf. 2016, 50, 117–130. [Google Scholar] [CrossRef]
- Rautiainen, M.; Lukeš, P.; Homolová, L.; Hovi, A.; Pisek, J.; Mõttus, M. Spectral Properties of Coniferous Forests: A Review of In Situ and Laboratory Measurements. Remote Sens. 2018, 10, 207. [Google Scholar] [CrossRef]
- Lukeš, P.; Stenberg, P.; Rautiainen, M.; Mõttus, M.; Vanhatalo, K.M. Optical properties of leaves and needles for boreal tree species in Europe. Remote Sens. Lett. 2013, 4, 667–676. [Google Scholar] [CrossRef]
- Roberts, D.A.; Ustin, S.L.; Ogunjemiyo, S.; Greenberg, J.; Dobrowski, S.Z.; Chen, J.; Hinckley, T.M. Spectral and Structural Measures of Northwest Forest Vegetation at Leaf to Landscape Scales. Ecosystems 2004, 7, 545–562. [Google Scholar] [CrossRef]
- Robinson, T.M.P.; La Pierre, K.J.; Vadeboncoeur, M.A.; Byrne, K.M.; Thomey, M.L.; Colby, S.E. Seasonal, not annual precipitation drives community productivity across ecosystems. Oikos 2013, 122, 727–738. [Google Scholar] [CrossRef]
- Hiltner, U.; Bräuning, A.; Gebrekirstos, A.; Huth, A.; Fischer, R. Impacts of precipitation variability on the dynamics of a dry tropical montane forest. Ecol. Model. 2016, 320, 92–101. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y. Impacts of climate, phenology, elevation and their interactions on the net primary productivity of vegetation in Yunnan, China under global warming. Ecol. Indic. 2023, 154, 110533. [Google Scholar] [CrossRef]
- Otto, R.; Fernández-Palacios, J.M.; Krüsi, B.O. Variation in species composition and vegetation structure of succulent scrub on Tenerife in relation to environmental variation. J. Veg. Sci. 2001, 12, 237–248. [Google Scholar] [CrossRef]
- Zheng, Y.; Xie, Z.; Jiang, L.; Shimizu, H.; Rimmington, G.M.; Zhou, G. Vegetation responses along environmental gradients on the Ordos plateau, China. Ecol. Res. 2006, 21, 396–404. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Molecular Regulation of Plant Responses to Environmental Temperatures. Mol. Plant. 2020, 13, 544–564. [Google Scholar] [CrossRef]
- Doughty, C.E.; Keany, J.M.; Wiebe, B.C.; Rey-Sanchez, C.; Carter, K.R.; Middleby, K.B.; Cheesman, A.W.; Goulden, M.L.; da Rocha, H.R.; Miller, S.D.; et al. Tropical forests are approaching critical temperature thresholds. Nature 2023, 621, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, X.; Zhou, X.; Zhao, K.; Yang, Q. Holocene Vegetation Succession and Response to Climate Change on the South Bank of the Heilongjiang-Amur River, Mohe County, Northeast China. Adv. Meteorol. 2016, 2016, 2450697. [Google Scholar] [CrossRef]
- Ouyang, Z.; Zheng, H.; Xiao, Y.; Polasky, S.; Liu, J.; Xu, W.; Wang, Q.; Zhang, L.; Xiao, Y.; Rao, E.; et al. Improvements in ecosystem services from investments in natural capital. Science 2016, 352, 1455–1459. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Q.; Corlett, R.T.; Zhai, D. After the rubber boom: Good news and bad news for biodiversity in Xishuangbanna, Yunnan, China. Reg. Envir. Chang. 2019, 19, 1713–1724. [Google Scholar] [CrossRef]
- Kavgacı, A.; Balpınar, N.; Öner, H.H.; Arslan, M.; Bonari, G.; Chytrý, M.; Čarni, A. Classification of forest and shrubland vegetation in Mediterranean Turkey. Appl. Veg. Sci. 2021, 24, e12589. [Google Scholar] [CrossRef]
- Gao, N.; Guan, T.; Zhou, J.; Cai, W.; Zhang, X.; Li, H.; Jiang, L.; Zheng, Y. Vegetation patterns and causal factors in different reaches of an endorheic basin in arid China. Écoscience 2019, 26, 71–83. [Google Scholar] [CrossRef]
- Bueno, M.L.; Rezende, V.L.; Pontara, V.; de Oliveira-Filho, A.T. Floristic distributional patterns in a diverse ecotonal area in South America. Plant Ecol. 2017, 218, 1171–1186. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Vegetation Types | Dominant Tree Species | Elevation Ranges | Numbers of Forest Sites |
|---|---|---|---|
| Tropical forest (TF) | Pometia pinnata, Lasiococca comberi var. pseudoverticillata, and Terminalia myriocarpa | <1100 m | 52 |
| Monsoon evergreen broad-leaved forest (MEBLF) | Castanopsis mekongensis, Schima wallichii, and Castanopsis echinocarpa | 1100–1800 m | 119 |
| Mountainous humid evergreen broad-leaved forest (MHEBLF) | Lithocarpus xylocarpus, Symplocos ramosissima, and Lithocarpus craibianus | 2000–2900 m | 59 |
| Semi-humid evergreen broad-leaved forest (SHEBLF) | Quercus schottkyana, Castanopsis orthacantha, and Manglietia duclouxii | 1700–2500 m | 45 |
| Rubber plantation (RP) | Hevea brasiliensis | <1200 m | 59 |
| Warm-hot coniferous forest (WHCF) | Pinus kesiya | 850–1800 m | 139 |
| Warm-temperate coniferous forest (WTCF) | Pinus yunnanensis, Pinus armandi, and Cunninghamia laceolata | 1500–2800 m | 361 |
| Cold-temperate coniferous forest (CTCF) | Abies georgei, Picea likiangensis, and Larix potaninii var. australis | >2700 m | 70 |
| Temperate-cool coniferous forest (TCCF) | Pinus densata | 1800–3300 m | 44 |
| DDI | PDI | PDI-DDI | CPDI-PDI | TPDI-PDI | EPDI-PDI | ||
|---|---|---|---|---|---|---|---|
| F1 score | TF | 25.750% | 40.163% | 14.413% | 29.862% | 23.202% | 36.723% |
| MEBLF | 60.312% | 62.555% | 2.244% | 15.422% | 11.741% | 13.484% | |
| SHEBLF | 30.327% | 34.026% | 3.699% | 21.035% | 18.159% | 28.575% | |
| MHEBLF | 54.492% | 54.408% | −0.084% | 5.160% | 3.853% | 15.613% | |
| RP | 67.529% | 80.127% | 12.598% | 7.923% | 8.611% | 9.446% | |
| WHCF | 51.339% | 53.502% | 2.163% | 21.041% | 19.424% | 21.645% | |
| WTCF | 72.049% | 72.935% | 0.887% | 13.660% | 13.207% | 15.221% | |
| CTCF | 64.576% | 71.989% | 7.413% | 6.857% | 7.244% | 8.139% | |
| TCCF | 8.491% | 14.893% | 6.402% | 45.899% | 48.287% | 46.050% | |
| Overall accuracy (OA) | 60.295% | 63.825% | 3.530% | 14.845% | 13.613% | 16.583% | |
| Kappa coefficient (KC) | 47.800% | 0.522% | 4.341% | 20.432% | 18.902% | 22.689% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Liu, D.; Li, Q.; Wanasinghe, D.N.; Zhai, D.; Zhao, G.; Xu, J. Enhancing Accuracy in Historical Forest Vegetation Mapping in Yunnan with Phenological Features, and Climatic and Elevation Variables. Remote Sens. 2024, 16, 3687. https://doi.org/10.3390/rs16193687
Yang J, Liu D, Li Q, Wanasinghe DN, Zhai D, Zhao G, Xu J. Enhancing Accuracy in Historical Forest Vegetation Mapping in Yunnan with Phenological Features, and Climatic and Elevation Variables. Remote Sensing. 2024; 16(19):3687. https://doi.org/10.3390/rs16193687
Chicago/Turabian StyleYang, Jianbo, Detuan Liu, Qian Li, Dhanushka N. Wanasinghe, Deli Zhai, Gaojuan Zhao, and Jianchu Xu. 2024. "Enhancing Accuracy in Historical Forest Vegetation Mapping in Yunnan with Phenological Features, and Climatic and Elevation Variables" Remote Sensing 16, no. 19: 3687. https://doi.org/10.3390/rs16193687