
Use of Remote Sensing and Biogeochemical Modeling to Simulate the Impact of Climatic and Anthropogenic Factors on Forest Carbon Fluxes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Data
2.3. GPP and NEP Modeling Strategy
2.4. Data Processing
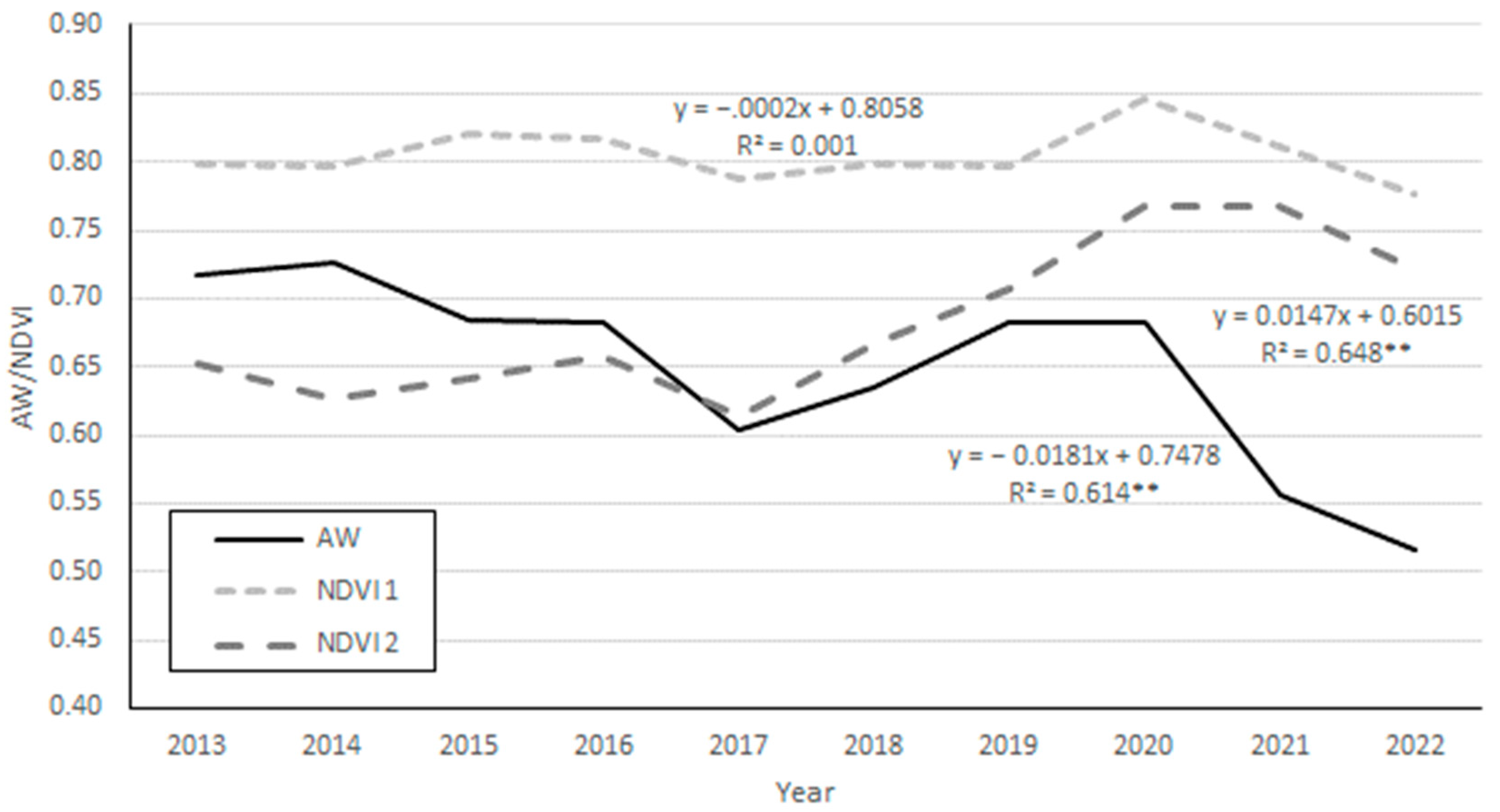
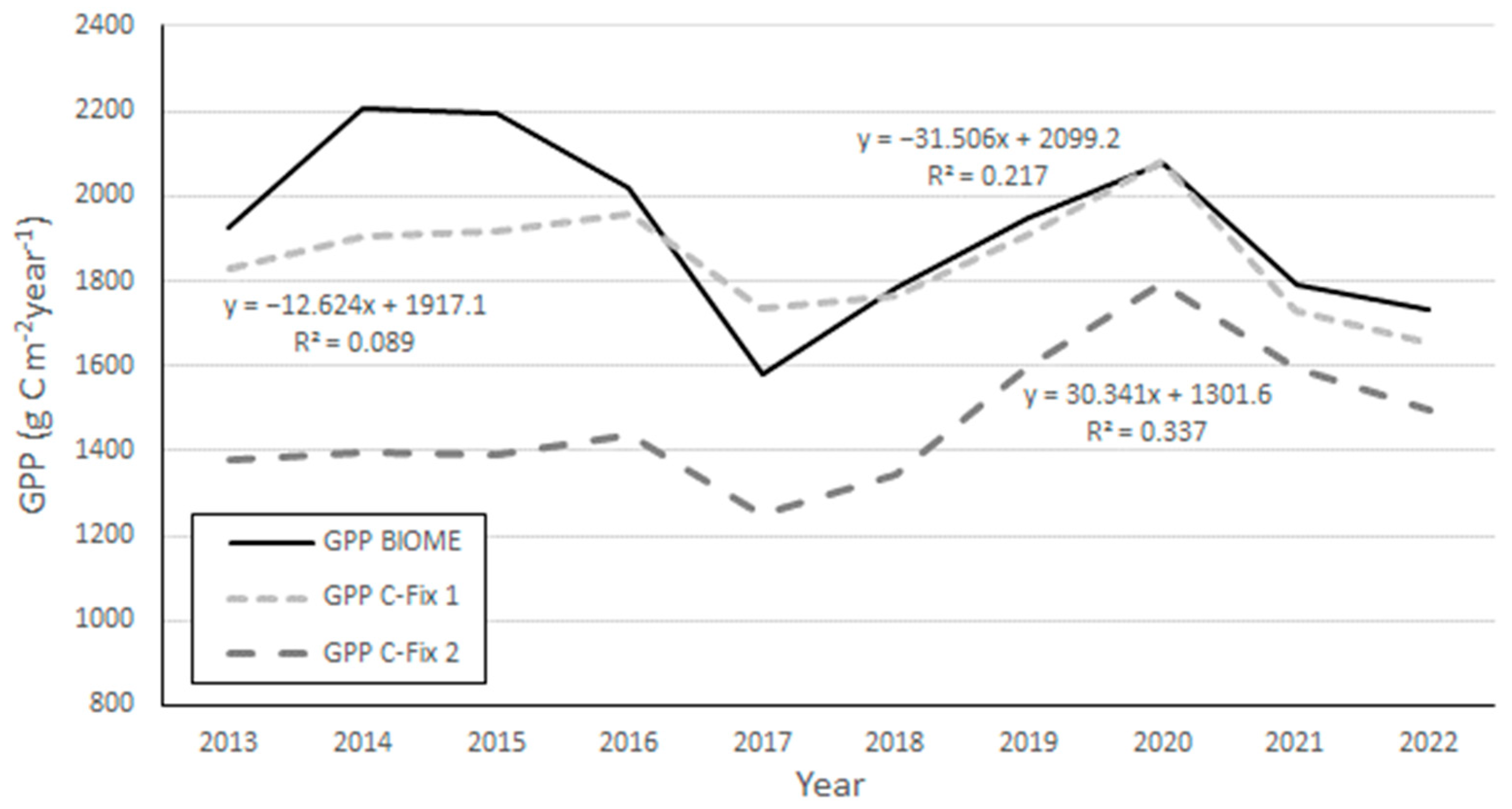
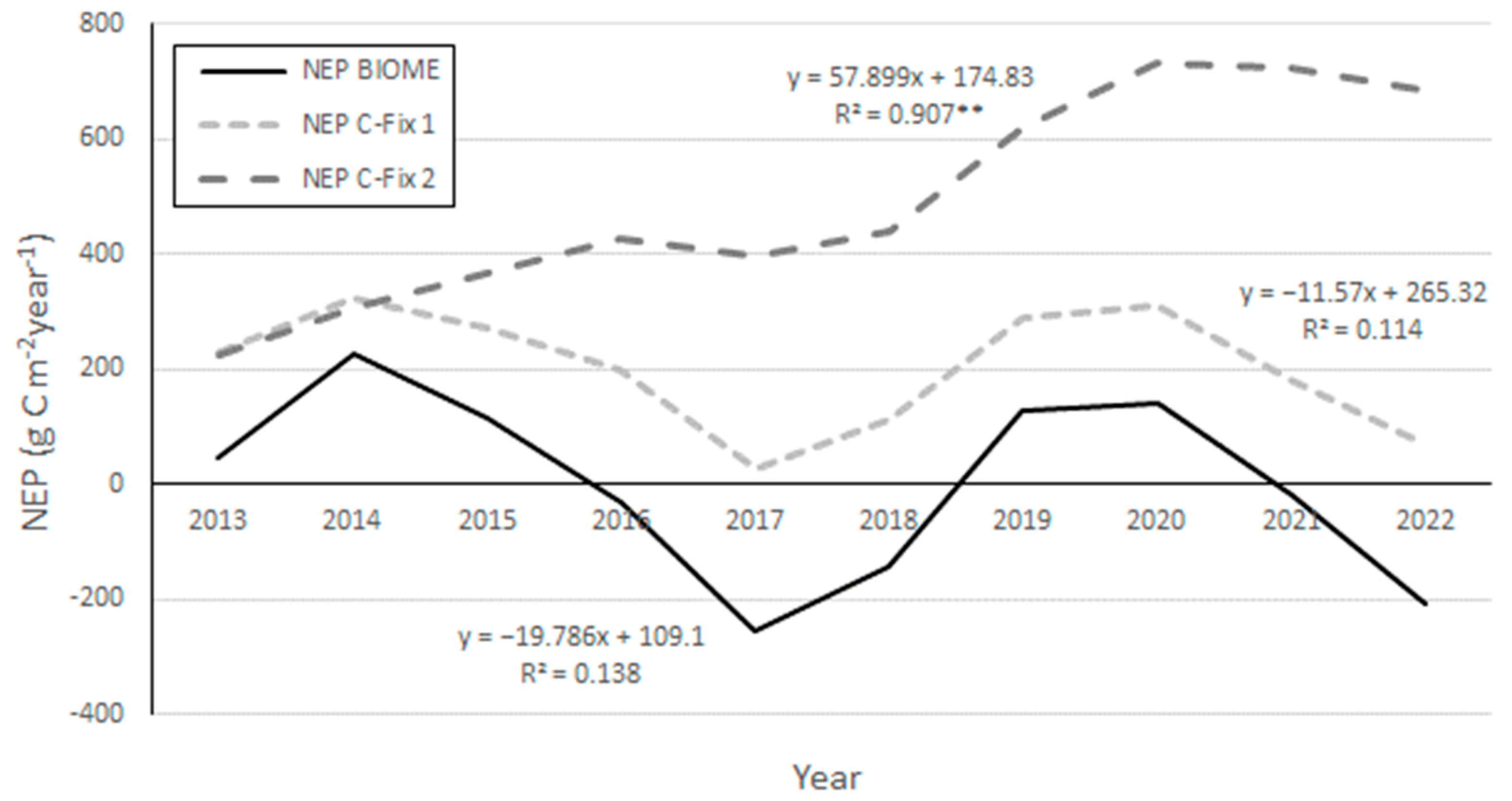
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waring, H.R.; Running, S.W. Forest Ecosystems. In Analysis at Multiples Scales, 3rd ed.; Academic Press: San Diego, CA, USA, 2007. [Google Scholar]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, W.; Liu, B.; Tian, X.; Chen, S.; Wang, H.; Mao, Y. Forest carbon flux simulation using multi-source data and incorporation of remotely sensed model with process-based model. Remote Sens. 2022, 14, 4766. [Google Scholar] [CrossRef]
- Yan, M.; Tian, X.; Li, Z.; Chen, E.; Wang, X.; Han, Z.; Sun, H. Simulation of forest carbon fluxes using model incorporation and data assimilation. Remote Sens. 2016, 8, 567. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, D.; Cao, Y.; Zhang, L.; Peng, H.; Wang, K.; Xi, H.; Wang, C. An integrated remote sensing and model approach for assessing forest carbon fluxes in China. Sci. Total Environ. 2022, 811, 152480. [Google Scholar] [CrossRef]
- Srinet, R.; Nandy, S.; Patel, N.R.; Padalia, H.; Watham, T.; Singh, S.K.; Chauhan, P. Simulation of forest carbon fluxes by integrating remote sensing data into biome-BGC model. Ecol. Model. 2023, 475, 110185. [Google Scholar] [CrossRef]
- Maselli, F.; Chiesi, M.; Moriondo, M.; Fibbi, L.; Bindi, M.; Running, S.W. Modelling the forest carbon budget of a Mediterranean region through the integration of ground and satellite data. Ecol. Model. 2009, 220, 330–342. [Google Scholar] [CrossRef]
- Fibbi, L.; Moriondo, M.; Chiesi, M.; Bindi, M.; Maselli, F. Impacts of climate changes on the gross primary production of Italian forests. Ann. For. Sci. 2019, 76, 59. [Google Scholar] [CrossRef]
- Sciarretta, A.; Marziali, L.; Squarcini, M.; Marianelli, L.; Benassai, D.; Logli, F.; Roversi, P.F. Adaptive management of invasive pests in natural protected areas: The case of Matsucoccus feytaudi in Central Italy. Bull. Entomol. Res. 2015, 106, 9–18. [Google Scholar] [CrossRef]
- Thornton, P.E.; Running, S.W.; White, M.A. Generating surfaces of daily meteorological variables over large regions of complex terrain. J. Hydrol. 1997, 190, 214–251. [Google Scholar] [CrossRef]
- Thornton, P.E.; Hasenauer, H.; White, M.A. Simultaneous estimation of daily solar radiation and humidity from observed temperature and precipitation: An application over complex terrain in Austria. Agric. For. Meteorol. 2000, 104, 255–271. [Google Scholar] [CrossRef]
- Gardin, L.; Chiesi, M.; Fibbi, L.; Maselli, F. Mapping soil organic carbon in Tuscany through the statistical combination of ground observations with ancillary and remote sensing data. Geoderma 2021, 404, 115386. [Google Scholar] [CrossRef]
- Chirici, G.; Chiesi, M.; Corona, P.; Puletti, N.; Mura, M.; Maselli, F. Prediction of forest NPP in Italy by the combination of ground and remote sensing data. Eur. J. For. Res. 2015, 134, 453–467. [Google Scholar] [CrossRef]
- Veroustraete, F.; Sabbe, H.; Eerens, H. Estimation of carbon mass fluxes over Europe using the C-Fix model and Euroflux data. Remote Sens. Environ. 2002, 83, 376–399. [Google Scholar] [CrossRef]
- Maselli, F.; Papale, D.; Puletti, N.; Chirici, G.; Corona, P. Combining remote sensing and ancillary data to monitor the gross productivity of water-limited forest ecosystems. Remote Sens. Environ. 2009, 113, 657–667. [Google Scholar] [CrossRef]
- Golinkoff, J. Biome BGC Version 4.2: Theoretical Framework of Biome-BGC. January 2010. Available online: http://www.ntsg.umt.edu/project/biome-bgc (accessed on 14 March 2023).
- White, M.A.; Thornton, P.E.; Running, S.W.; Nemani, R.R. Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model: Net primary production controls. Earth Interact. 2000, 4, 1–85. [Google Scholar] [CrossRef]
- Liu, S.; Bond-Lamberty, B.; Hicke, J.A.; Vargas, R.; Zhao, S.; Chen, J.; Edburg, S.L.; Hu, Y.; Liu, J.; McGuire, A.D.; et al. Simulating the impacts of disturbances on forest carbon cycling in North America: Processes, data, models, and challenges. J. Geophys. Res. Biogeoscience 2011, 116, G00K08. [Google Scholar] [CrossRef]
- Chirici, G.; Chiesi, M.; Fibbi, L.; Giannetti, F.; Corona, P.; Maselli, F. High spatial resolution modelling of net forest carbon fluxes based on ground and remote sensing data. Agric. For. Meteorol. 2022, 316, 108866. [Google Scholar] [CrossRef]
- Maselli, F.; Mari, R.; Chiesi, M. Use of LiDAR data to simulate forest net primary production. Int. J. Remote Sens. 2013, 34, 2487–2501. [Google Scholar] [CrossRef]
- Heute, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Ju, J.; Roy, D.P. The availability of cloud-free Landsat ETM+ data over the conterminous United States and globally. Remote Sens. Environ. 2008, 112, 1196–1211. [Google Scholar] [CrossRef]
- Wu, J.; Lin, L.; Li, T.; Cheng, Q.; Zhang, C.; Shen, H. Fusing Landsat 8 and Sentinel-2 data for 10-m dense time-series imagery using a degradation-term constrained deep network. Int. J. Appl. Earth Obs. Geoinf. 2022, 108, 102738. [Google Scholar] [CrossRef]
- Pouladi, N.; Gholizadeh, A.; Khosrawi, V.; Boruvka, L. Digital mapping of soil organic carbon using remote sensing data: A systematic review. Catena 2023, 232, 107409. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NDVI 1 | NDVI 2 | GPP BIOME | GPP C-Fix 1 | GPP C-Fix 2 | NEP BIOME | NEP C-Fix 1 | NEP C-Fix 2 | |
|---|---|---|---|---|---|---|---|---|
| AW | 0.410 * | −0.427 * | 0.744 ** | 0.769 ** | −0.032 | 0.744 ** | 0.755 ** | −0.600 ** |
| NDVI 1 | 0.364 | 0.562 * | 0.799 ** | 0.585 * | 0.519 * | 0.584 ** | 0.210 | |
| NDVI 2 | 0.332 | 0.063 | 0.887 ** | 0.088 | 0.137 | 0.906 ** | ||
| GPP BIOME | 0.778 ** | 0.282 | 0.900 ** | 0.904 ** | −0.236 | |||
| GPP C-Fix 1 | 0.484 * | 0.750 ** | 0.808 ** | −0.035 | ||||
| GPP C-Fix 2 | 0.444 * | 0.503 * | 0.787 ** | |||||
| NEP BIOME | 0.993 ** | −0.109 | ||||||
| NEP C-Fix 1 | −0.064 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiesi, M.; Fibbi, L.; Vanucci, S.; Maselli, F. Use of Remote Sensing and Biogeochemical Modeling to Simulate the Impact of Climatic and Anthropogenic Factors on Forest Carbon Fluxes. Remote Sens. 2024, 16, 232. https://doi.org/10.3390/rs16020232
Chiesi M, Fibbi L, Vanucci S, Maselli F. Use of Remote Sensing and Biogeochemical Modeling to Simulate the Impact of Climatic and Anthropogenic Factors on Forest Carbon Fluxes. Remote Sensing. 2024; 16(2):232. https://doi.org/10.3390/rs16020232
Chicago/Turabian StyleChiesi, Marta, Luca Fibbi, Silvana Vanucci, and Fabio Maselli. 2024. "Use of Remote Sensing and Biogeochemical Modeling to Simulate the Impact of Climatic and Anthropogenic Factors on Forest Carbon Fluxes" Remote Sensing 16, no. 2: 232. https://doi.org/10.3390/rs16020232
APA StyleChiesi, M., Fibbi, L., Vanucci, S., & Maselli, F. (2024). Use of Remote Sensing and Biogeochemical Modeling to Simulate the Impact of Climatic and Anthropogenic Factors on Forest Carbon Fluxes. Remote Sensing, 16(2), 232. https://doi.org/10.3390/rs16020232