
Mapping Trails and Tracks in the Boreal Forest Using LiDAR and Convolutional Neural Networks



Abstract
:1. Introduction
1.1. Mapping Trails and Tracks
1.2. Research Objectives
- To demonstrate the capacity of high-density LiDAR and CNNs to map trails and tracks automatically in a natural environment;
- To compare the accuracy of trail/track maps developed with LiDAR from a drone platform (185 points/m2) and a piloted-aircraft platform (30 points/m2) to evaluate trade-offs between spatial resolution and operational scalability; and
- To measure the abundance and distribution of tracks and trails across different land-cover classes and their co-location with anthropogenic disturbances across our study area in the Canadian boreal forest.
2. Materials and Methods
2.1. Study Area
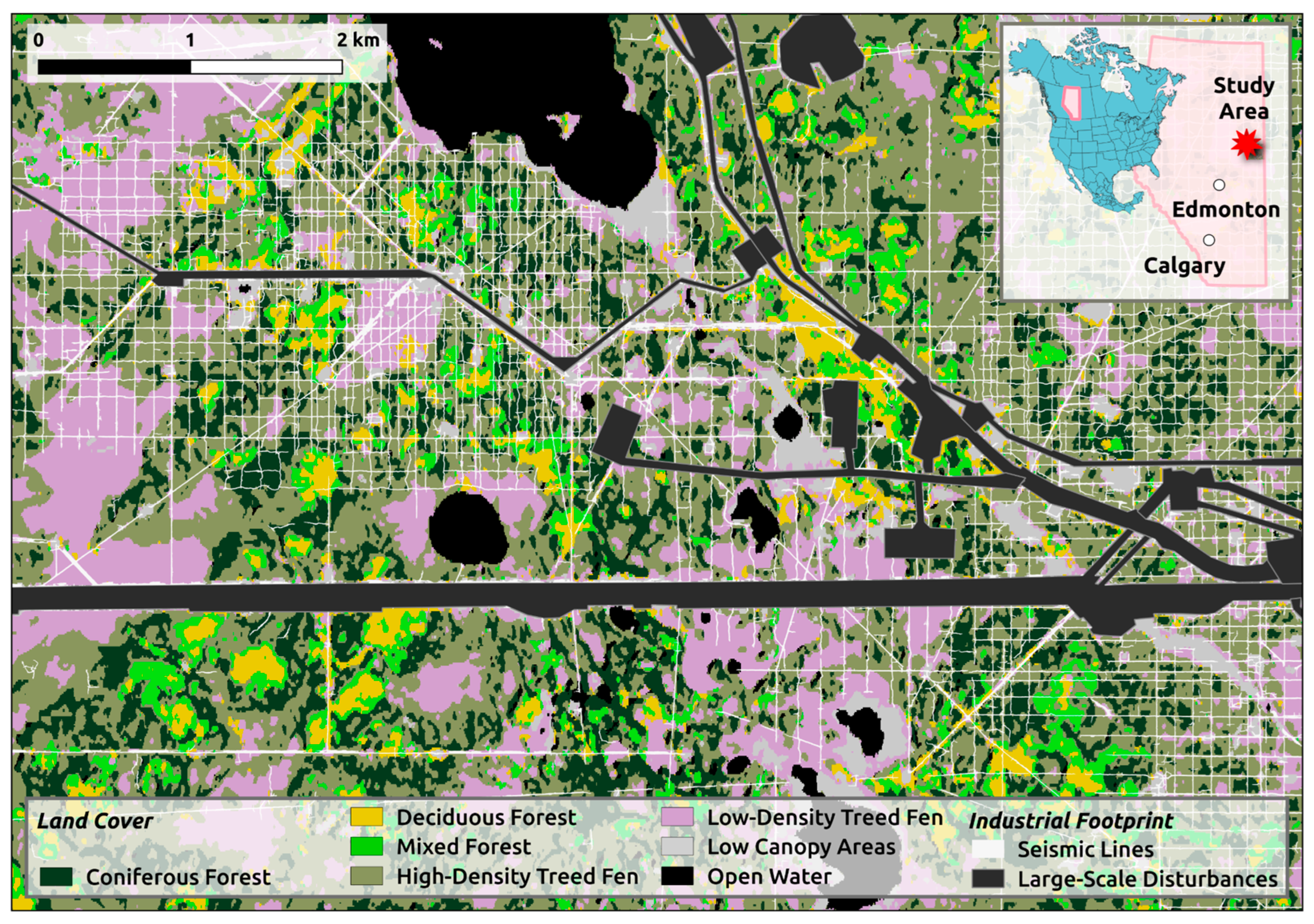

2.2. Data Acquisition and Processing
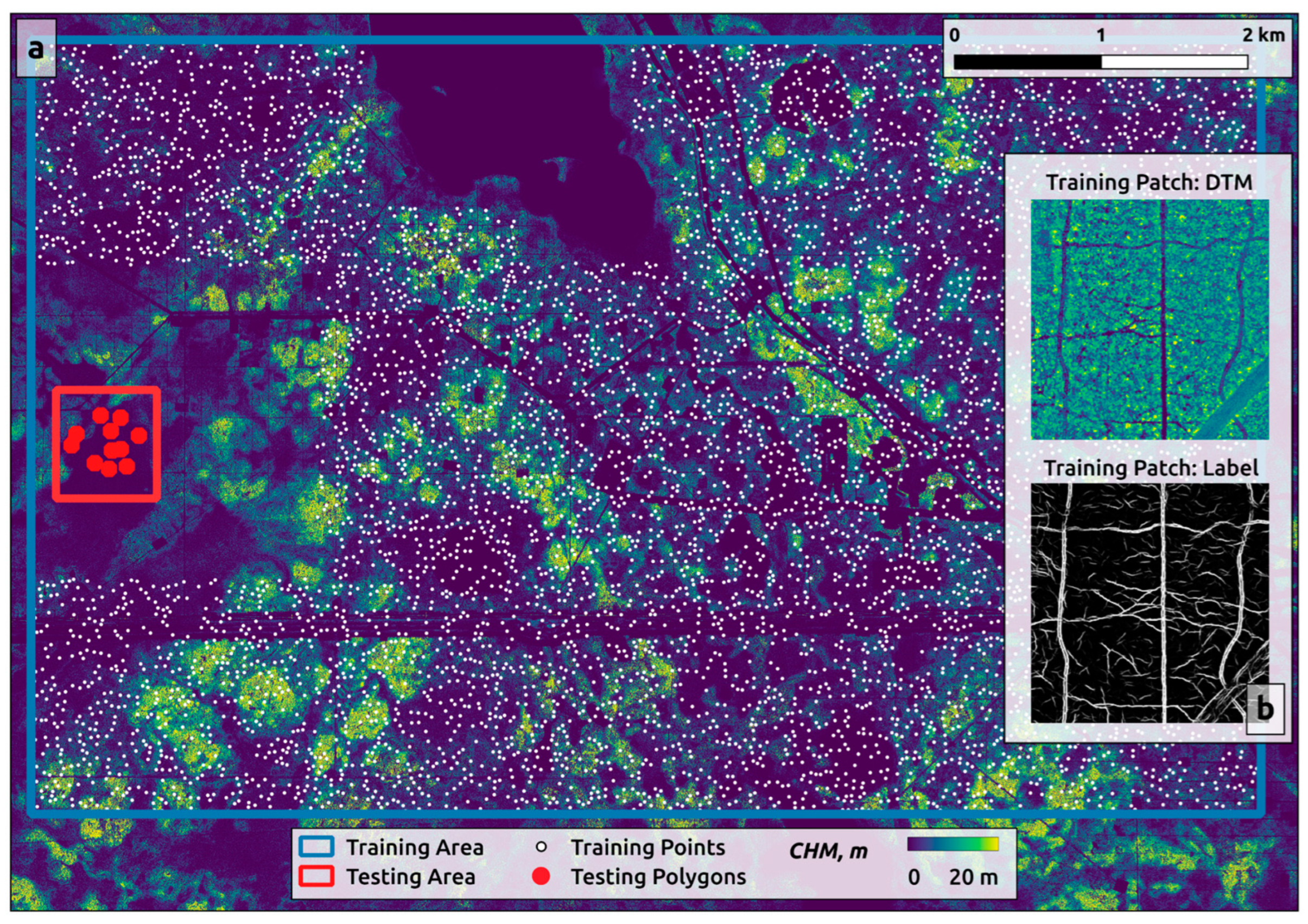
2.3. Mapping Trails and Tracks
2.3.1. Training Data Preparation
2.3.2. U-Net Model Architecture
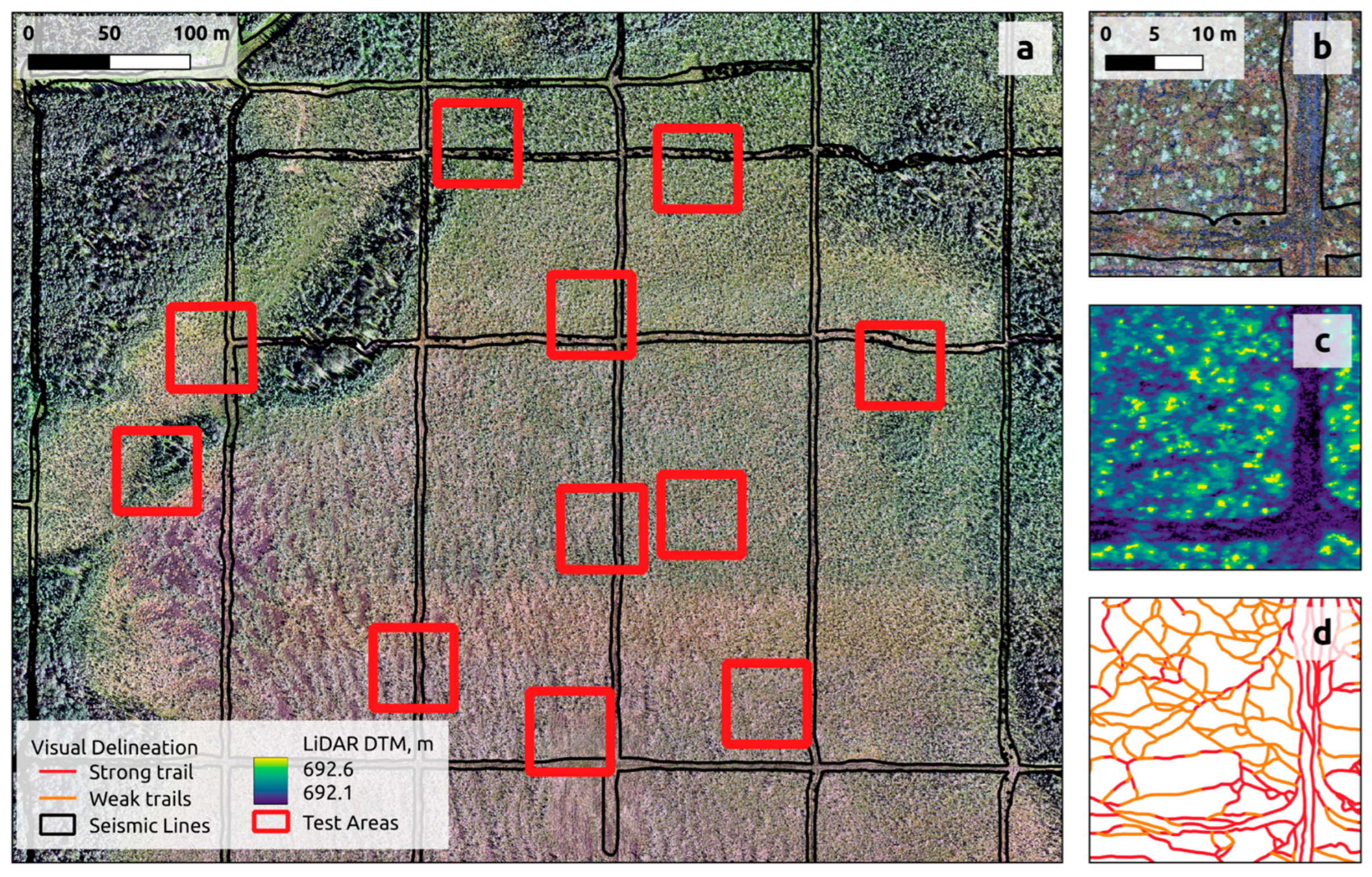
2.3.3. Accuracy Assessment
2.4. Land Cover and Seismic Line Maps
3. Results
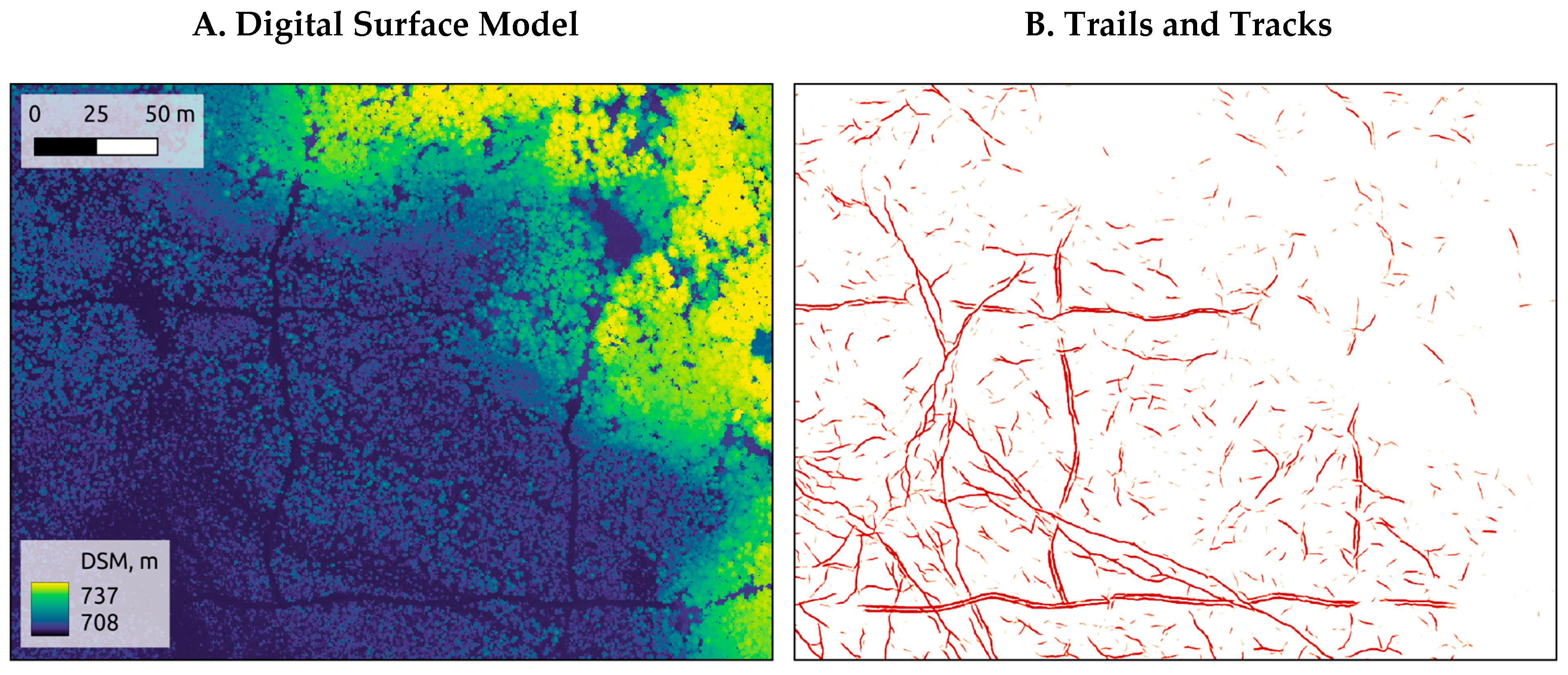
3.1. Mapping Trails and Tracks

3.2. Accuracy of Piloted-Aircraft- and Drone-Based Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Source | Precision (%) | Recall (%) | F1 Score (%) | Average Precision |
|---|---|---|---|---|
| Aerial 50 cm DTM | 77 ± 9 | 78 ± 14 | 77 ± 9 | 0.69 |
| Drone 10 cm DTM | 70 ± 10 | 80 ± 8 | 74 ± 6 | 0.64 |

3.3. Distribution of Trails and Tracks Across Land-Cover Classes
| Trails and Tracks | |||
|---|---|---|---|
| Land-Cover Type | Land-Cover Area km2 (%) | Length km (%) | Density (km/km2) |
| Coniferous forest | 10 (16.9) | 396 (14.0) | 40 |
| Deciduous forest | 3 (5.1) | 62 (2.2) | 21 |
| Mixed forest | 3 (5.1) | 51 (1.8) | 17 |
| High-density treed fen | 24 (40.7) | 1342 (47.4) | 56 |
| Low-density treed fen | 10 (16.9) | 978 (34.6) | 98 |
| Excluded areas (lakes, floodplains, roads, and dense industrial footprint) | 9 (15.2) | N/A | N/A |
| SUM | 59 (100) | 2829 (100) | |
3.4. Seismic Line Influence on Trails and Tracks
| Area, km2 (%) | Trails and Tracks Length km (%) | Trails and Tracks Density km/km2 | |
|---|---|---|---|
| On seismic lines | 4.2 (8) | 765 (27) | 182 |
| Off seismic lines | 45.7 (92) | 2064 (73) | 41 |
| SUM | 49.9 (100) | 2829 (100) |
4. Discussion
4.1. Boreal Trails and Tracks Can Be Mapped with LiDAR and Convolutional Neural Networks
4.2. Canopy Density and Substrate Materials Are Key Factors

4.3. Drone and Piloted-Aircraft Map Accuracies Are Statistically Identical
4.4. Patterns of Trails and Tracks in Our Study Area

4.5. Ecosystem Effects of Trails and Tracks
4.6. Assumptions and Limitations
4.7. Future Research Needs

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LiDAR | Light detection and ranging |
| GNSS | Global navigation satellite system |
| CNN | Convolutional neural network |
| OHV | Off-highway vehicle |
| BERA | Boreal Ecosystem Recovery and Assessment |
| PPP | Precise point positioning |
| RTK | Real-time kinematic |
| DTM | Digital terrain model |
| S1 | Sentinel 1 |
| S2 | Sentinel 2 |
| CHM | Canopy height model |
References
- Perna, A.; Latty, T. Animal transportation networks. J. R. Soc. Interface 2014, 11, 20140334. [Google Scholar] [CrossRef] [PubMed]
- Myslajek, R.W.; Olkowska, E.; Wronka-Tomulewicz, M.; Nowak, S. Mammal use of wildlife crossing structures along a new motorway in an area recently recolonized by wolves. Eur. J. Wildl. Res. 2020, 66, 79. [Google Scholar] [CrossRef]
- Newmark, W.D.; Rickart, E.A. High-use movement pathways and habitat selection by ungulates. Mamm. Biol. 2012, 77, 293–298. [Google Scholar] [CrossRef]
- Squies, J.R.; Olson, L.E.; Roberts, E.K.; Ivan, J.S.; Hebblewhite, M. Winter recreation and Canada lynx: Reducing conflict through niche partitioning. Ecosphere 2019, 10, e02876. [Google Scholar] [CrossRef]
- Hebblewhite, M.; Miguelle, D.G.; Murzin, A.A.; Aramilev, V.V.; Pikunov, D.G. Predicting potential habitat and population size for reintroduction of the Far Eastern leopards in the Russian Far East. Biol. Conserv. 2011, 144, 2403–2413. [Google Scholar] [CrossRef]
- Kondratenkov, I.A.; Oparin, M.L.; Oparina, O.S. Estimation of the Ecological Density of Some Species of Game according to Winter Route Censuses. Biol. Bull. 2023, 50, 2710–2718. [Google Scholar] [CrossRef]
- Whittington, J.; St Clair, C.C.; Mercer, G. Path tortuosity and the permeability of roads and trails to wolf movement. Ecol. Soc. 2004, 9, 4. [Google Scholar] [CrossRef]
- Slovikosky, S.A.; Merrick, M.J.; Morandini, M.; Koprowski, J.L. Movement response of small mammals to burn severity reveals importance of microhabitat features. J. Mammal. 2024, 105, 157–167. [Google Scholar] [CrossRef]
- Kays, R.; Crofoot, M.C.; Jetz, W.; Wikelski, M. Terrestrial animal tracking as an eye on life and planet. Science 2015, 348, aaa2478. [Google Scholar] [CrossRef]
- Muhly, T.B.; Semeniuk, C.; Massolo, A.; Hickman, L.; Musiani, M. Human Activity Helps Prey Win the Predator-Prey Space Race. PLoS ONE 2011, 6, e17050. [Google Scholar] [CrossRef]
- Palm, E.C.; Landguth, E.L.; Holden, Z.A.; Day, C.C.; Lamb, C.T.; Frame, P.F.; Morehouse, A.T.; Mowat, G.; Proctor, M.F.; Sawaya, M.A.; et al. Corridor-based approach with spatial cross-validation reveals scale-dependent effects of geographic distance, human footprint and canopy cover on grizzly bear genetic connectivity. Mol. Ecol. 2023, 32, 5211–5227. [Google Scholar] [CrossRef]
- Harmsen, B.J.; Foster, R.J.; Silver, S.; Ostro, L.; Doncaster, C.P. Differential Use of Trails by Forest Mammals and the Implications for Camera-Trap Studies: A Case Study from Belize. Biotropica 2010, 42, 126–133. [Google Scholar] [CrossRef]
- Rowcliffe, J.M.; Field, J.; Turvey, S.T.; Carbone, C. Estimating animal density using camera traps without the need for individual recognition. J. Appl. Ecol. 2008, 45, 1228–1236. [Google Scholar] [CrossRef]
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. For. Ecol. Manag. 2015, 338, 124–138. [Google Scholar] [CrossRef]
- Kormanek, M.; Dvorák, J.; Tylek, P.; Jankovsky, M.; Nuhlícek, O.; Mateusiak, L. Impact of MHT9002HV Tracked Harvester on Forest Soil after Logging in Steeply Sloping Terrain. Forests 2023, 14, 977. [Google Scholar] [CrossRef]
- Battigelli, J.P.; Spence, J.R.; Langor, D.W.; Berch, S.M. Short-term impact of forest soil compaction and organic matter removal on soil mesofauna density and oribatid mite diversity. Can. J. For. Res. 2004, 34, 1136–1149. [Google Scholar] [CrossRef]
- Croke, J.; Hairsine, P.; Fogarty, P. Soil recovery from track construction and harvesting changes in surface infiltration, erosion and delivery rates with time. For. Ecol. Manag. 2001, 143, 3–12. [Google Scholar] [CrossRef]
- Robroek, B.J.M.; Smart, R.P.; Holden, J. Sensitivity of blanket peat vegetation and hydrochemistry to local disturbances. Sci. Total Environ. 2010, 408, 5028–5034. [Google Scholar] [CrossRef]
- Ares, A.; Terry, T.A.; Miller, R.E.; Anderson, H.W.; Flaming, B.L. Ground-based forest harvesting effects on soil physical properties and Douglas-Fir growth. Soil Sci. Soc. Am. J. 2005, 69, 1822–1832. [Google Scholar] [CrossRef]
- Marra, E.; Cambi, M.; Fernandez-Lacruz, R.; Giannetti, F.; Marchi, E.; Nordfjell, T. Photogrammetric estimation of wheel rut dimensions and soil compaction after increasing numbers of forwarder passes. Scand. J. For. Res. 2018, 33, 613–620. [Google Scholar] [CrossRef]
- Davidson, S.J.; Goud, E.M.; Franklin, C.; Nielsen, S.E.; Strack, M. Seismic Line Disturbance Alters Soil Physical and Chemical Properties Across Boreal Forest and Peatland Soils. Front. Earth Sci. 2020, 8, 281. [Google Scholar] [CrossRef]
- Dabros, A.; Higgins, K.L.; Pinzon, J. Seismic line edge effects on plants, lichens and their environmental conditions in boreal peatlands of Northwest Alberta (Canada). Restor. Ecol. 2022, 30, e13468. [Google Scholar] [CrossRef]
- Lovitt, J.; Rahman, M.M.; McDermid, G.J. Assessing the Value of UAV Photogrammetry for Characterizing Terrain in Complex Peatlands. Remote Sens. 2017, 9, 715. [Google Scholar] [CrossRef]
- Filicetti, A.T.; Nielsen, S.E. Effects of wildfire and soil compaction on recovery of narrow linear disturbances in upland mesic boreal forests. For. Ecol. Manag. 2022, 510, 120073. [Google Scholar] [CrossRef]
- Wimpey, J.F.; Marion, J.L. The influence of use, environmental and managerial factors on the width of recreational trails. J. Environ. Manag. 2010, 91, 2028–2037. [Google Scholar] [CrossRef]
- Costantino, C.; Mantini, N.; Benedetti, A.C.; Bartolomei, C.; Predari, G. Digital and Territorial Trails System for Developing Sustainable Tourism and Enhancing Cultural Heritage in Rural Areas: The Case of San Giovanni Lipioni, Italy. Sustainability 2022, 14, 13982. [Google Scholar] [CrossRef]
- Norman, P.; Pickering, C.M. Using volunteered geographic information to assess park visitation: Comparing three on-line platforms. Appl. Geogr. 2017, 89, 163–172. [Google Scholar] [CrossRef]
- Loosen, A.; Capdevila, T.V.; Pigeon, K.; Wright, P.; Jacob, A.L. Understanding the role of traditional and user-created recreation data in the cumulative footprint of recreation. J. Outdoor Recreat. Tour.-Res. Plan. Manag. 2023, 44, 100615. [Google Scholar] [CrossRef]
- Abdollahi, A.; Pradhan, B.; Shukla, N.; Chakraborty, S.; Alamri, A. Deep Learning Approaches Applied to Remote Sensing Datasets for Road Extraction: A State-Of-The-Art Review. Remote Sens. 2020, 12, 1444. [Google Scholar] [CrossRef]
- Washington-Allen, R.A.; Van Niel, T.G.; Ramsey, D.; West, N.E. Remote Sensing-Based Piosphere Analysis. GIScience Remote Sens. 2004, 41, 136–154. [Google Scholar] [CrossRef]
- Dara, A.; Baumann, M.; Freitag, M.; Hölzel, N.; Hostert, P.; Kamp, J.; Müller, D.; Prishchepov, A.V.; Kuemmerle, T. Annual Landsat time series reveal post-Soviet changes in grazing pressure. Remote Sens. Environ. 2020, 239, 111667. [Google Scholar] [CrossRef]
- Chang, C.C.; Wang, J.; Zhao, Y.B.; Cai, T.Y.; Yang, J.L.; Zhang, G.L.; Wu, X.C.; Otgonbayar, M.; Xiao, X.M.; Xin, X.P.; et al. A 10-m annual grazing intensity dataset in 2015–2021 for the largest temperate meadow steppe in China. Sci. Data 2024, 11, 181. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.; Madden, M.; Doren, R.F. Mapping the Everglades. Photogramm. Eng. Remote Sens. 1999, 65, 163–170. [Google Scholar]
- Kaiser, J.V.; Stow, D.A.; Cao, L. Evaluation of remote sensing techniques for mapping transborder trails. Photogramm. Eng. Remote Sens. 2004, 70, 1441–1447. [Google Scholar] [CrossRef]
- Yamato, C.; Ichikawa, K.; Arai, N.; Tanaka, K.; Nishiyama, T.; Kittiwattanawong, K. Deep neural networks based automated extraction of dugong feeding trails from UAV images in the intertidal seagrass beds. PLoS ONE 2021, 16, e0255586. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, S.; Puliti, S.; Talbot, B.; Heppelmann, J.B.; Breidenbach, J.; Astrup, R. Mapping wheel-ruts from timber harvesting operations using deep learning techniques in drone imagery. Forestry 2022, 95, 698–710. [Google Scholar] [CrossRef]
- Marra, E.; Wictorsson, R.; Bohlin, J.; Marchi, E.; Nordfjell, T. Remote measuring of the depth of wheel ruts in forest terrain using a drone. Int. J. For. Eng. 2021, 32, 224–234. [Google Scholar] [CrossRef]
- Flyckt, J.; Andersson, F.; Lavesson, N.; Nilsson, L.; Gren, A. Detecting ditches using supervised learning on high-resolution digital elevation models. Expert Syst. Appl. 2022, 201, 116961. [Google Scholar] [CrossRef]
- Lidberg, W.; Paul, S.S.; Westphal, F.; Richter, K.F.; Lavesson, N.; Melniks, R.; Ivanovs, J.; Ciesielski, M.; Leinonen, A.; Agren, A.M. Mapping Drainage Ditches in Forested Landscapes Using Deep Learning and Aerial Laser Scanning. J. Irrig. Drain. Eng. 2023, 149, 04022051. [Google Scholar] [CrossRef]
- Du, L.; McCarty, G.W.; Li, X.; Zhang, X.; Rabenhorst, M.C.; Lang, M.W.; Zou, Z.H.; Zhang, X.S.; Hinson, A.L. Drainage ditch network extraction from lidar data using deep convolutional neural networks in a low relief landscape. J. Hydrol. 2024, 628, 130591. [Google Scholar] [CrossRef]
- Natural Regions Committee. Natural Regions and Subregions of Alberta; Compiled by D.J. Downing and W.W. Pettapiece; Government of Alberta: Edmonton, AB, Canada, 2006; Pub. No. T/852.
- Vitt, D.H. An overview of factors that influence the development of Canadian peatlands. Mem. Entomol. Soc. Can. 1994, 126, 7–20. [Google Scholar] [CrossRef]
- Dabros, A.; Pyper, M.; Castilla, G. Seismic lines in the boreal and arctic ecosystems of North America: Environmental impacts, challenges, and opportunities. Environ. Rev. 2018, 26, 214–229. [Google Scholar] [CrossRef]
- Roussel, J.R.; Auty, D.; Coops, N.C.; Tompalski, P.; Goodbody, T.R.H.; Meador, A.S.; Bourdon, J.F.; de Boissieu, F.; Achim, A. lidR: An R package for analysis of Airborne Laser Scanning (ALS) data. Remote Sens. Environ. 2020, 251, 112061. [Google Scholar] [CrossRef]
- Roussel, J.R.; Auty, D. Airborne LiDAR Data Manipulation and Visualization for Forestry Applications. R Package Version 4.1.2. 2024. Available online: https://cran.r-project.org/package=lidR (accessed on 22 April 2025).
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. arXiv 2015, arXiv:1505.04597. [Google Scholar]
- Oktay, O.; Schlemper, J.; Le Folgoc, L.; Lee, M.; Heinrich, M.; Misawa, K.; Mori, K.; McDonagh, S.; Hammerla, N.Y.; Kainz, B.; et al. Attention U-Net: Learning Where to Look for the Pancreas. arXiv 2018, arXiv:1804.03999. [Google Scholar]
- Kingma, D.P.; Ba, J. Adam: A Method for Stochastic Optimization. arXiv 2014, arXiv:1412.6980. [Google Scholar]
- Zhang, T.Y.; Suen, C.Y. A Fast Parallel Algorithm for Thinning Digital Patterns. Commun. ACM 1984, 27, 236–239. [Google Scholar] [CrossRef]
- Queiroz, G.L.; McDermid, G.J.; Rahman, M.M.; Linke, J. The Forest Line Mapper: A Semi-Automated Tool for Mapping Linear Disturbances in Forests. Remote Sens. 2020, 12, 4176. [Google Scholar] [CrossRef]
- Price, J.S.; Cagampan, J.; Kellner, E. Assessment of peat compressibility: Is there an easy way? Hydrol. Process. 2005, 19, 3469–3475. [Google Scholar] [CrossRef]
- Crow, P.; Benham, S.; Devereux, B.J.; Amable, G.S. Woodland vegetation and its implications for archaeological survey using LiDAR. Forestry 2007, 80, 241–252. [Google Scholar] [CrossRef]
- Razak, K.A.; Straatsma, M.W.; van Westen, C.J.; Malet, J.P.; de Jong, S.M. Airborne laser scanning of forested landslides characterization: Terrain model quality and visualization. Geomorphology 2011, 126, 186–200. [Google Scholar] [CrossRef]
- Dietmaier, A.; McDermid, G.J.; Rahman, M.M.; Linke, J.; Ludwig, R. Comparison of LiDAR and Digital Aerial Photogrammetry for Characterizing Canopy Openings in the Boreal Forest of Northern Alberta. Remote Sens. 2019, 11, 1919. [Google Scholar] [CrossRef]
- Finnegan, L.; Hebblewhite, M.; Pigeon, K.E. Whose line is it anyway? Moose (Alces alces) response to linear features. Ecosphere 2023, 14, e4636. [Google Scholar] [CrossRef]
- Finnegan, L.; Pigeon, K.E.; Cranston, J.; Hebblewhite, M.; Musiani, M.; Neufeld, L.; Schmiegelow, F.; Duval, J.; Stenhouse, G.B. Natural regeneration on seismic lines influences movement behaviour of wolves and grizzly bears. PLoS ONE 2018, 13, e0195480. [Google Scholar] [CrossRef]
- Dickie, M.; Serrouya, R.; McNay, R.S.; Boutin, S. Faster and farther: Wolf movement on linear features and implications for hunting behaviour. J. Appl. Ecol. 2017, 54, 253–263. [Google Scholar] [CrossRef]
- Dickie, M.; Sherman, G.G.; Sutherland, G.D.; McNay, R.S.; Cody, M. Evaluating the impact of caribou habitat restoration on predator and prey movement. Conserv. Biol. 2023, 37, e14004. [Google Scholar] [CrossRef]
- Lovitt, J.; McDermid, G.J.; Kariyeva, J.; Mahoney, C.; Hall, R.J.; Franklin, S.E.; Saxena, A.; Kalacska, M. UAV Remote Sensing Can Reveal the Effects of Low-Impact Seismic Lines on Surface Morphology, Hydrology, and Methane (CH4) Release in a Boreal Peatland. Remote Sens. 2018, 10, 1549. [Google Scholar] [CrossRef]
- Evans, M.; Warburton, J. Erosion in peatlands: Recent research progress and future directions. Earth-Sci. Rev. 2010, 103, 30–49. [Google Scholar] [CrossRef]
- Williams-Mounsey, J.; Crowle, A.; Grayson, R.; Lindsay, R.; Holden, J. Surface structure on abandoned upland blanket peatland tracks. J. Environ. Manag. 2023, 325, 116561. [Google Scholar] [CrossRef]
- Pellerin, S.; Huot, J.; Côté, S.D. Long term effects of deer browsing and trampling on the vegetation of peatlands. Biol. Conserv. 2006, 128, 316–326. [Google Scholar] [CrossRef]
- Persson, I.L.; Danell, K.; Bergström, R. Disturbance by large herbivores in boreal forests with special reference to moose. Ann. Zool. Fenn. 2000, 37, 251–263. [Google Scholar]
- Holmgren, M.; Groten, F.; Carracedo, M.R.; Vink, S.; Limpens, J. Rewilding Risks for Peatland Permafrost. Ecosystems 2023, 26, 1806–1818. [Google Scholar] [CrossRef]
- Ramirez, J.I.; Jansen, P.A.; Poorter, L. Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: A semi-quantitative review. For. Ecol. Manag. 2018, 424, 406–419. [Google Scholar] [CrossRef]
- Bengtsson, F.; Rydin, H.; Baltzer, J.L.; Bragazza, L.; Bu, Z.J.; Caporn, S.J.M.; Dorrepaal, E.; Flatberg, K.I.; Galanina, O.; Galka, M.; et al. Environmental drivers of Sphagnum growth in peatlands across the Holarctic region. J. Ecol. 2021, 109, 417–431. [Google Scholar] [CrossRef]
- Adriaensen, F.; Chardon, J.P.; De Blust, G.; Swinnen, E.; Villalba, S.; Gulinck, H.; Matthysen, E. The application of ‘least-cost’ modelling as a functional landscape model. Landsc. Urban Plan. 2003, 64, 233–247. [Google Scholar] [CrossRef]
- Etherington, T.R. Least-cost modelling and landscape ecology: Concepts, applications, and opportunities. Curr. Landsc. Ecol. Rep. 2016, 1, 40–53. [Google Scholar] [CrossRef]
- Nir, N.; Davidovich, U.; Ullman, M.; Schütt, B.; Stahlschmidt, M.C. The environmental footprint of Holocene societies: A multi-temporal study of trails in the Judean Desert, Israel. Front. Earth Sci. 2023, 11, 1148101. [Google Scholar] [CrossRef]
- Godtman Kling, K.; Fredman, P.; Wall-Reinius, S. Trails for tourism and outdoor recreation: A systematic literature review. Tour. Int. Interdiscip. J. 2017, 65, 488–508. [Google Scholar]
- Xie, E.; Wang, W.; Yu, Z.; Anandkumar, A.; Alvarez, J.M.; Luo, P. SegFormer: Simple and efficient design for semantic segmentation with transformers. Adv. Neural Inf. Process. Syst. 2021, 34, 12077–12090. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McDermid, G.J.; Terenteva, I.; Chan, X.Y. Mapping Trails and Tracks in the Boreal Forest Using LiDAR and Convolutional Neural Networks. Remote Sens. 2025, 17, 1539. https://doi.org/10.3390/rs17091539
McDermid GJ, Terenteva I, Chan XY. Mapping Trails and Tracks in the Boreal Forest Using LiDAR and Convolutional Neural Networks. Remote Sensing. 2025; 17(9):1539. https://doi.org/10.3390/rs17091539
Chicago/Turabian StyleMcDermid, Gregory J., Irina Terenteva, and Xue Yan Chan. 2025. "Mapping Trails and Tracks in the Boreal Forest Using LiDAR and Convolutional Neural Networks" Remote Sensing 17, no. 9: 1539. https://doi.org/10.3390/rs17091539
APA StyleMcDermid, G. J., Terenteva, I., & Chan, X. Y. (2025). Mapping Trails and Tracks in the Boreal Forest Using LiDAR and Convolutional Neural Networks. Remote Sensing, 17(9), 1539. https://doi.org/10.3390/rs17091539