1. Introduction
Stem volume is one of the most relevant resource attributes of forest inventories. In most European countries, volume is usually estimated or modeled based on field measurements [
1]. In the Nordic countries, conventional forest inventories at various geographical scales have over the past few decades been enhanced by using remotely sensed data such as airborne laser scanning (ALS) and stereo aerial photography [
2,
3]. It has been shown that for local management planning, data from ALS may reduce the overall costs by reducing the economic losses caused by incorrect decisions due to erroneous data [
4]. One of the biggest challenges in remote sensing-assisted inventories is the discrimination among tree species [
5]. Moreover, combining ALS data with hyperspectral images may improve the accuracy of species identification [
6]. The tree species information is needed for many forest applications, especially to retrieve species-specific forest biophysical properties, such as volume and diameter at breast height (DBH), to derive biodiversity indicators and to plan silvicultural activities and cutting regimes.
Due to accurate three-dimensional measurements obtained by ALS [
7,
8], the ALS technology has become one of the most valuable remote sensing methods for providing forest information. By using ALS data, biophysical attributes such as volume, height, DBH, crown area, and stem density can be derived or modeled with high accuracies [
2,
9,
10,
11,
12]. However, the estimation of biophysical attributes can be challenging in dense, multispecies, heterogeneous forest stands [
13,
14]. In the past, ALS data were examined to classify the tree species at the individual tree crown (ITC) level using shape, structure, and intensity features of the tree crowns [
5,
15]. In addition, when species-specific biophysical attributes are required, ALS data are often fused with optical images, i.e., color-infrared [
16,
17,
18], multispectral [
19,
20], or hyperspectral [
21,
22] images, in order to improve the tree species classification [
6,
23,
24]. Recently, multi- or hyper-spectral ALS sensors were developed with a high potential to be used as a future single-sensor solution for forest mapping [
25,
26,
27]. Currently, remotely sensed hyperspectral data are the most promising data source for classifying tree species due to their ability to detect subtle variations in the chemical and structural properties of the tree canopy. Likewise, other studies demonstrated the success of identifying tree species in other ecosystems [
28,
29]. Thus, the remote sensing technologies that currently are considered to have the greatest potential to improve forest attribute characterization are ALS, describing the 3D forest structure, and hyperspectral imagery, outperforming other techniques for the identification of tree species [
30]. The fusion of both has the potential to improve the forest attribute prediction accuracy [
31].
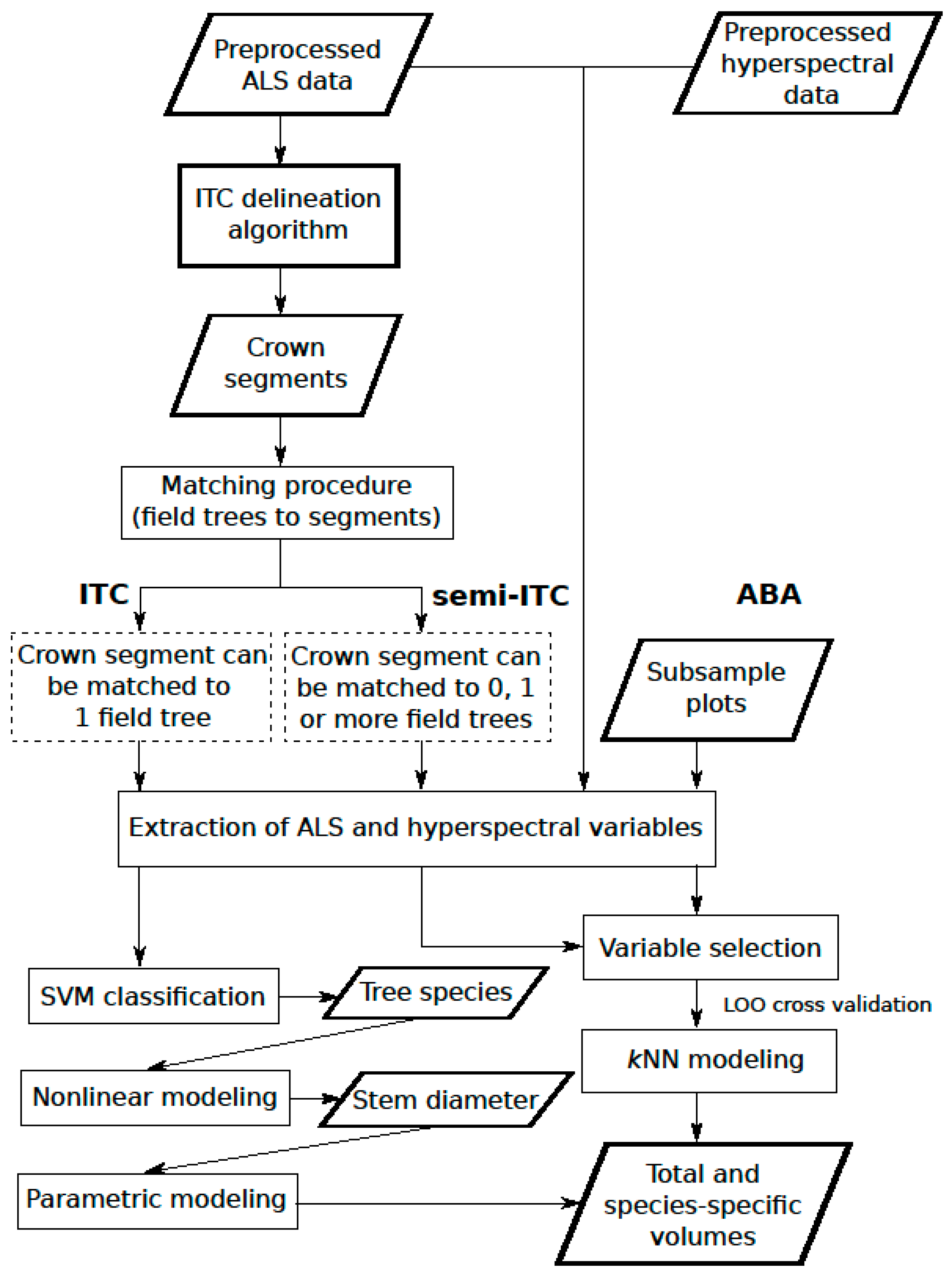
Most local forest management inventories assisted by ALS data follow the area-based approach (ABA). For field sample plots distributed across the area of interest, several variables are extracted from the ALS echoes describing structural properties, for which the statistical relationships with field-measured biophysical attributes are constructed. The relationships, typically in the form of regression models, are then used to predict biophysical attributes for larger areas [
9,
32]. As an alternative, individual trees can be identified and the forest resource variables can be estimated as an aggregate of individual tree properties. To obtain tree attributes, the ITC approach was introduced [
33]. With the ITC approach, crown segments are detected and delineated by applying segmentation algorithms [
34,
35]. The crown segments, often referred to as ITCs, can contain none, one, or several trees. In particular, this approach presumes that one crown segment contains exclusively one tree. For each crown segment, various ALS-derived structural variables are extracted, such as height and crown area. Based on these variables, the biophysical attributes, such as volume and DBH, are predicted for each segment and can be aggregated to other scales, e.g., a forest stand. Detection errors, i.e., omission error (failure to detect a tree or segmenting multiple trees into a single crown segment) and commission error (detecting an object that is not a tree or splitting a single tree into multiple crown segments), usually lead to underprediction of the forest biophysical attributes [
36]. To overcome this problem, the semi-ITC approach has been proposed [
37], which, in contrast to the ITC approach, allows a crown segment to contain multiple trees.
Several studies in Finland aimed to estimate species-specific forest volume using the combination of ALS data and aerial imagery based on ABA for three species groups: Norway spruce, Scots pine, and deciduous [
16,
17,
18,
38]. All these studies used a non-parametric
k-most similar neighbor (
kMSN) approach [
39] based on Packalén and Maltamo [
16]. Packalén and Maltamo [
16] compared two approaches for determining species-specific volumes at the plot level. The first approach predicted the total volume using ALS data, upon which the species-specific volume was obtained by multiplying the total volume by the proportion of each tree species obtained from aerial photographs and fuzzy classification. The second approach used the
kMSN method for simultaneous prediction of volumes by tree species. The
kMSN method resulted in more accurate species-specific volume estimates, except for the total volume where the fuzzy classification performed slightly better. Packalén and Maltamo [
17] extended the
kMSN approach from the previous study [
16] to a stand level and tested the simultaneous prediction of volume, stem number, basal area, basal area median diameter, and tree height for the same tree species groups. The attributes of coniferous tree species were predicted accurately and those of deciduous trees less accurately, since they were minority species. Vauhkonen et al. [
38] tested the performance of alpha shape metrics calculated from ALS data combined with image variables for species-specific volume predictions at the plot level. They demonstrated that using only ALS variables resulted in less accurate estimates compared to using combined ALS and images variables. Niska et al. [
18] compared the
kMSN method with three artificial neural network modeling methods: the multilayer perceptron (MLP), support vector regression (SVR), and self-organizing map (SOM) at the plot and stand level. The results revealed that the SVR and MLP models reached the greatest prediction accuracy, the
kMSN a lower accuracy, and the SOM the smallest prediction accuracy. It should be noted that the SVR and MLP yielded negative estimates to some extent, whereas the
kMSN and SOM estimates were always within the range of the modeling data.
In Norway, Breidenbach et al. [
37] determined species-specific volumes for four dominant tree species (spruce, pine, birch, and aspen) using the ABA and semi-ITC approaches, combining the ALS and multispectral data. They applied the
kNN approach using MSN and random forest (RF) methods for calculating statistical distances between the neighbors. The volumes predicted with the semi-ITC approach resulted in smaller RMSE compared with the ABA results. Ørka et al. [
23] evaluated tree species composition in a Norwegian forest using (1) ALS data, (2) multispectral imagery, (3) hyperspectral imagery, (4) fused ALS and multispectral data, and (5) fused ALS and hyperspectral data. Subsequently, they predicted the basal area for spruce, pine, and deciduous trees for three inventory approaches: ABA, semi-ITC, and ITC. For the ABA and semi-ITC approach, the
kNN algorithm with an Euclidean distance matrix was applied. The results suggested that the greatest species accuracy was achieved by fusing the ALS and hyperspectral data. The ITC approach resulted in the highest accuracy for deciduous species, while the ABA performed the best for the other tree species.
As mentioned above, different remote sensing-assisted forest inventory approaches have been suggested to assess species-specific forest attributes. Each of them has their own pros and cons. The advantage of the ABA is that it provides predictions with small systematic errors, i.e., mean differences, but at the same time only the attributes of the main tree species can be predicted with high accuracy, while for the minority species the errors were large and consequently resulted in inaccurate species-specific attributes [
23]. The ITC-based approaches are suitable for developing species-specific models which could lead to more accurate stand-level estimations, particularly for mixed stands. The main drawback of the ITC procedure are the under-predictions due to problems in detecting suppressed and understory trees, especially in stands with a complex forest structure. We expect that a solution for alleviating systematic errors lies in the semi-ITC approach, which has not been sufficiently explored and tested in various forest conditions and ecosystems. In fact, only a few studies have evaluated and compared the results of species-specific volumes and basal areas obtained from different remote sensing-assisted forest inventory approaches [
23,
37]. In particular, most of these studies were carried out in boreal forest conditions, accounting for spruce, pine, and deciduous tree species groups. Moreover, most of the remote sensing-assisted inventory methods combine ALS data with aerial photogrammetry or multispectral imagery to obtain species-specific volumes. The fusion of complementary data sources, in particular ALS and hyperspectral data, typically result in greater accuracies in contrast to using the respective data separately [
6,
23,
40]. Therefore, the fusion of ALS and hyperspectral data can offer a powerful basis for discriminating tree species and enables an accurate prediction of species-specific attributes. The
kNN approach has grown in popularity due to its ability to successfully and simultaneously relate multiple forest attributes derived from field observations to remotely sensed data [
41]. However, to the very best of our knowledge, there are no studies to date that have tested the ITC, semi-ITC, and ABA approaches on a comparative basis with emphasis on species-specific volumes.
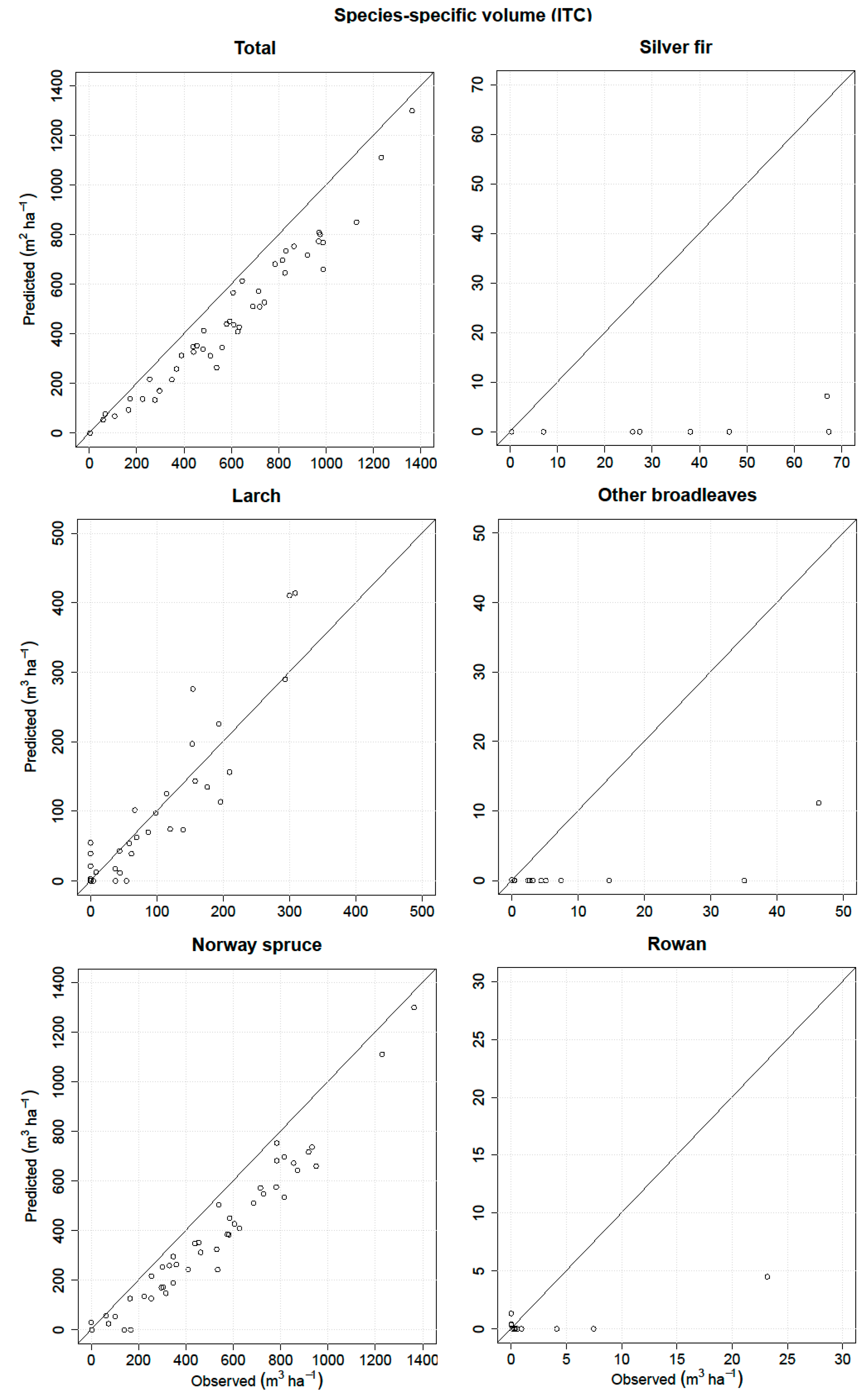
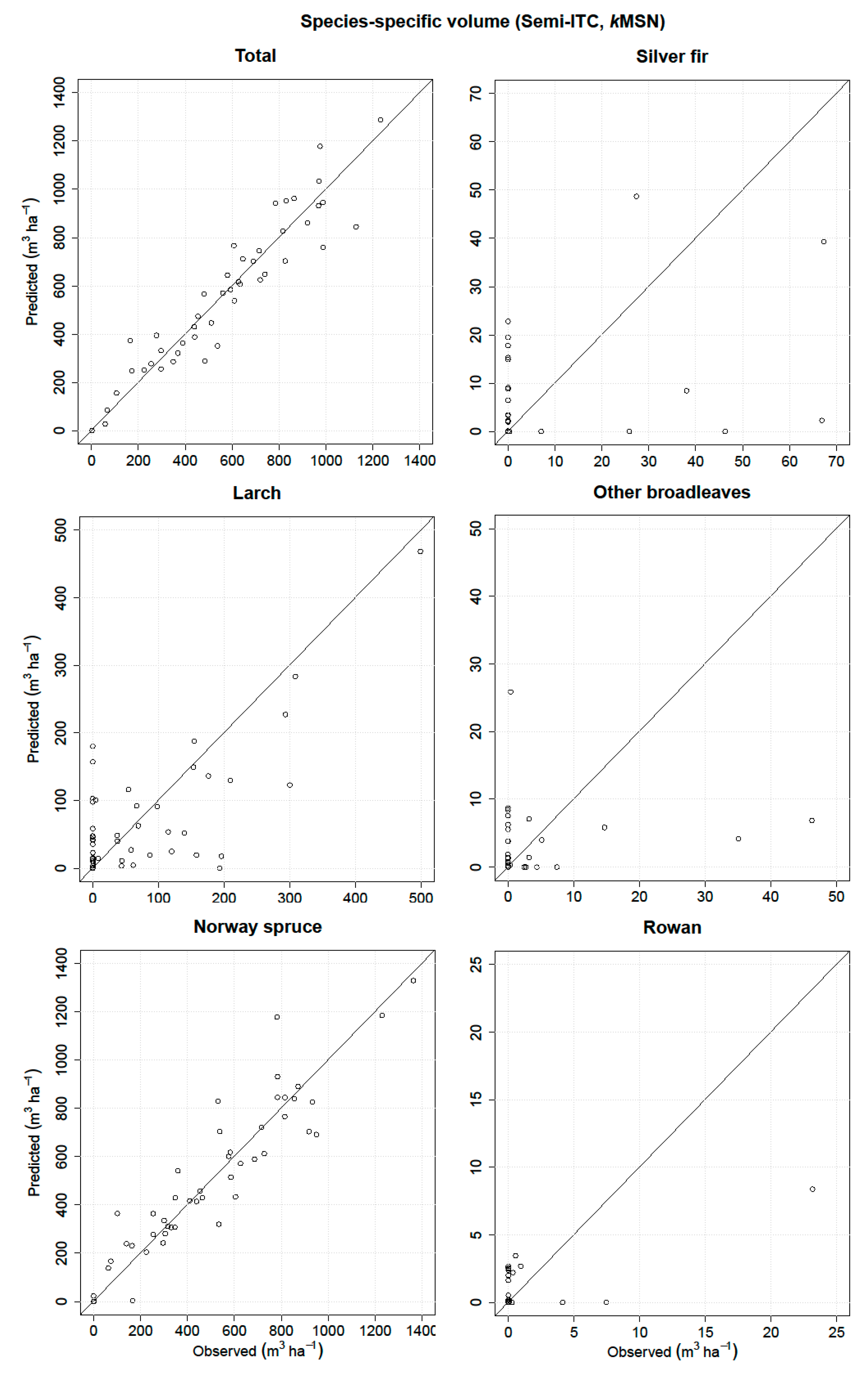
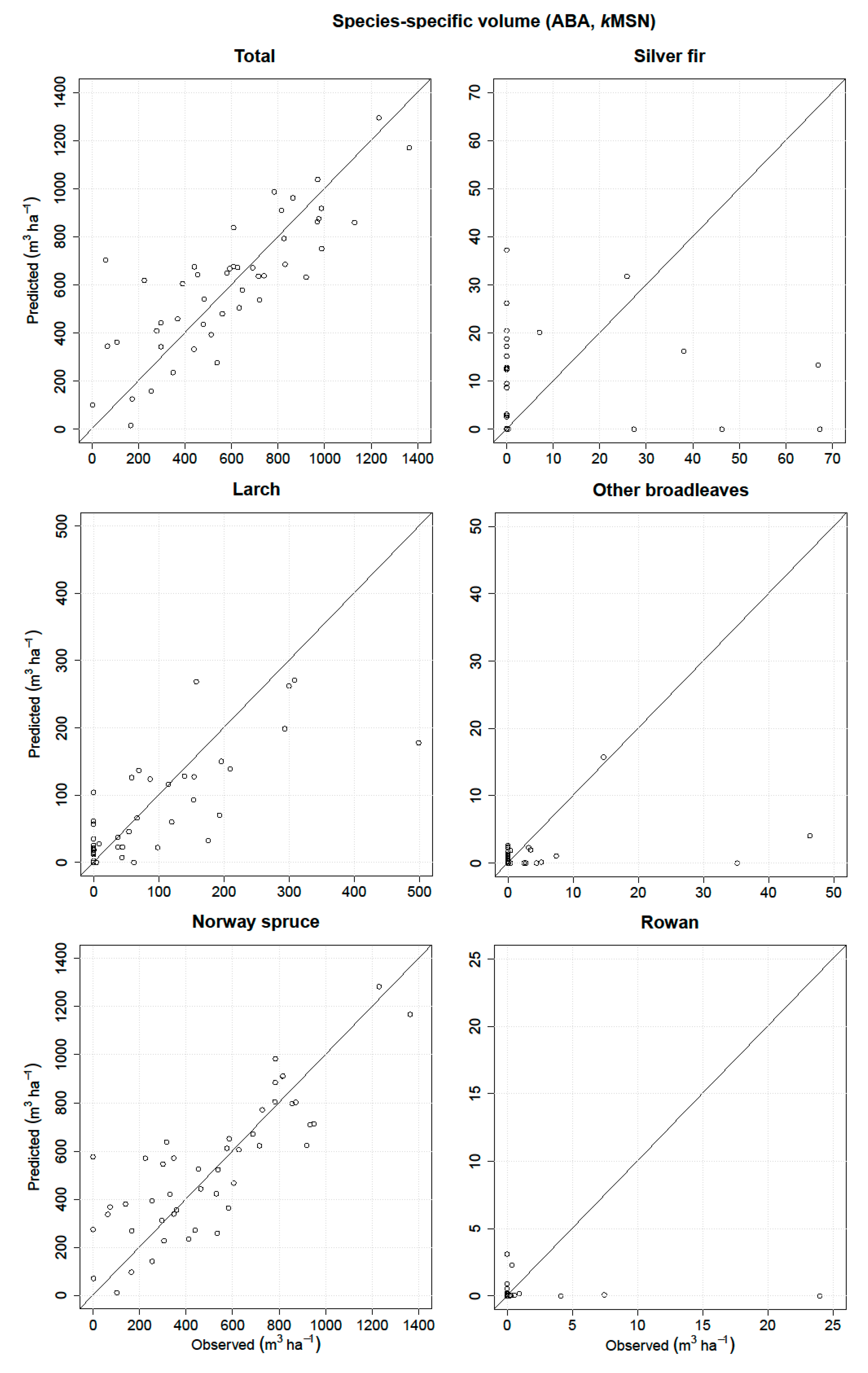
Thus, the main goal of the present study was to predict and evaluate the species-specific volume in a complex Alpine forest based on fused ALS and airborne hyperspectral data using three remote sensing-assisted forest inventory approaches, i.e., the ITC, semi-ITC, and ABA. The performances of all three inventory approaches were analyzed and compared for the total volume and volume of five species classes.
4. Discussion
Many methods have been proposed for fusing ALS data with stereo or airborne multispectral or hyperspectral images in order to achieve a more accurate species recognition [
23,
56,
57]. In this study, we showed that the fusion of ALS and hyperspectral data enabled the prediction of volumes at good accuracy levels. Moreover, hyperspectral variables were not only used for species classification, but they were also incorporated in the common ALS framework as they were selected as important variables for volume modeling. At the moment, hyperspectral data are the most powerful tool for species identification [
6] and consequently they can improve the accuracy of the predicted biophysical attributes [
31]. Forest inventories can be improved by the use of these combined data as they can increase the spatial detail, coverage, and accuracy of forest biophysical attributes. Thus, the combined data also helps forest managers to develop a broad and detailed database of forestry information to be coupled with a decision support system [
58].
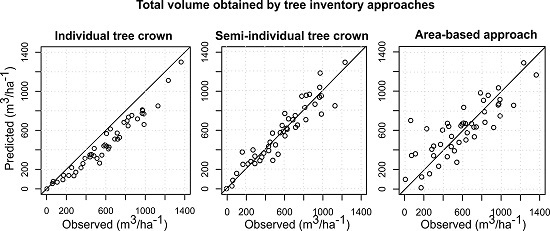
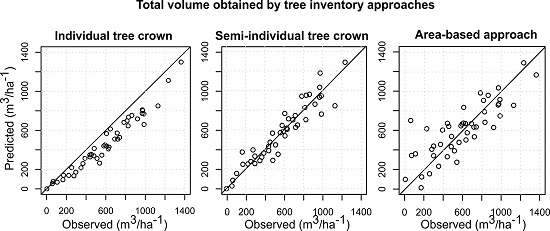
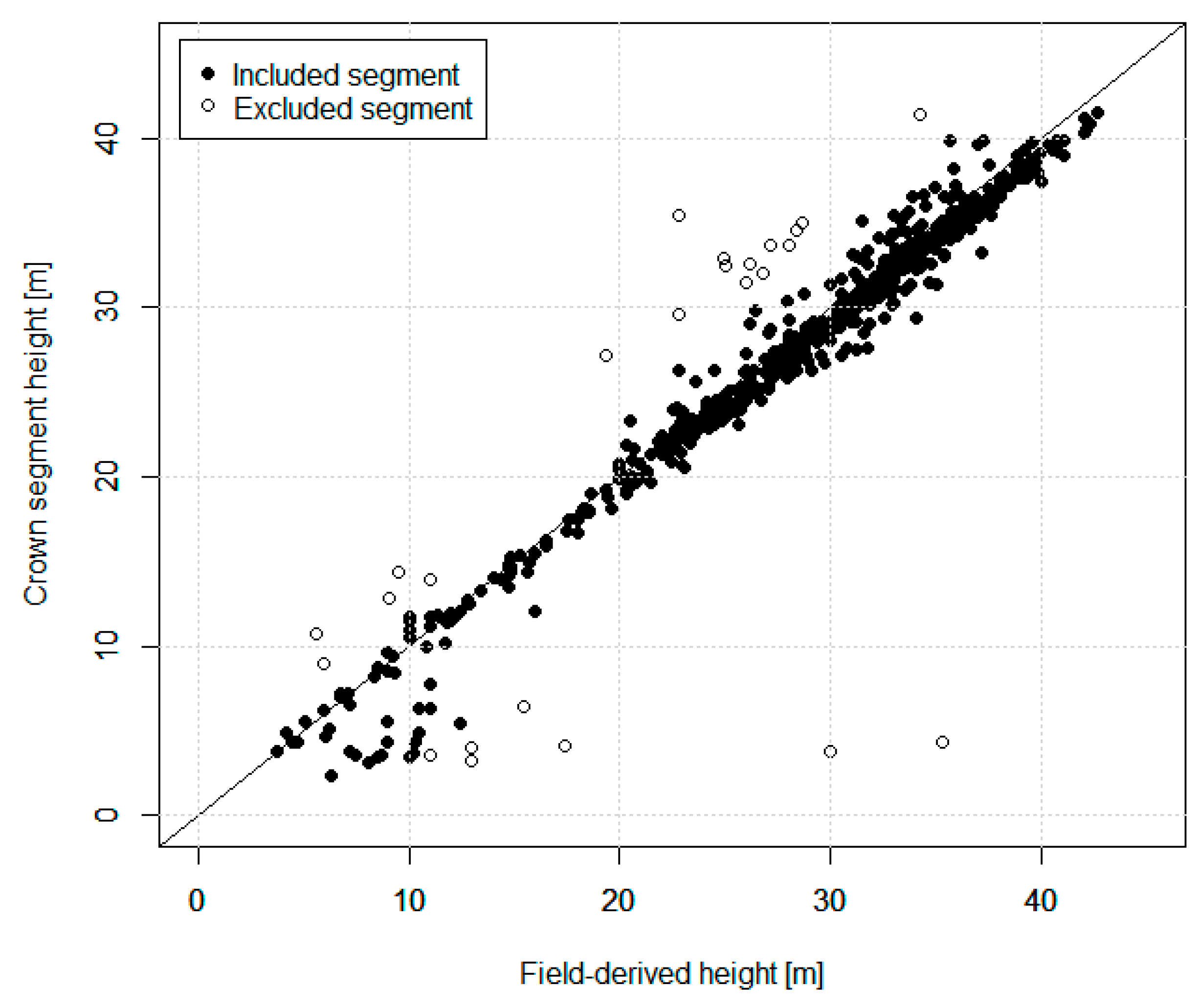
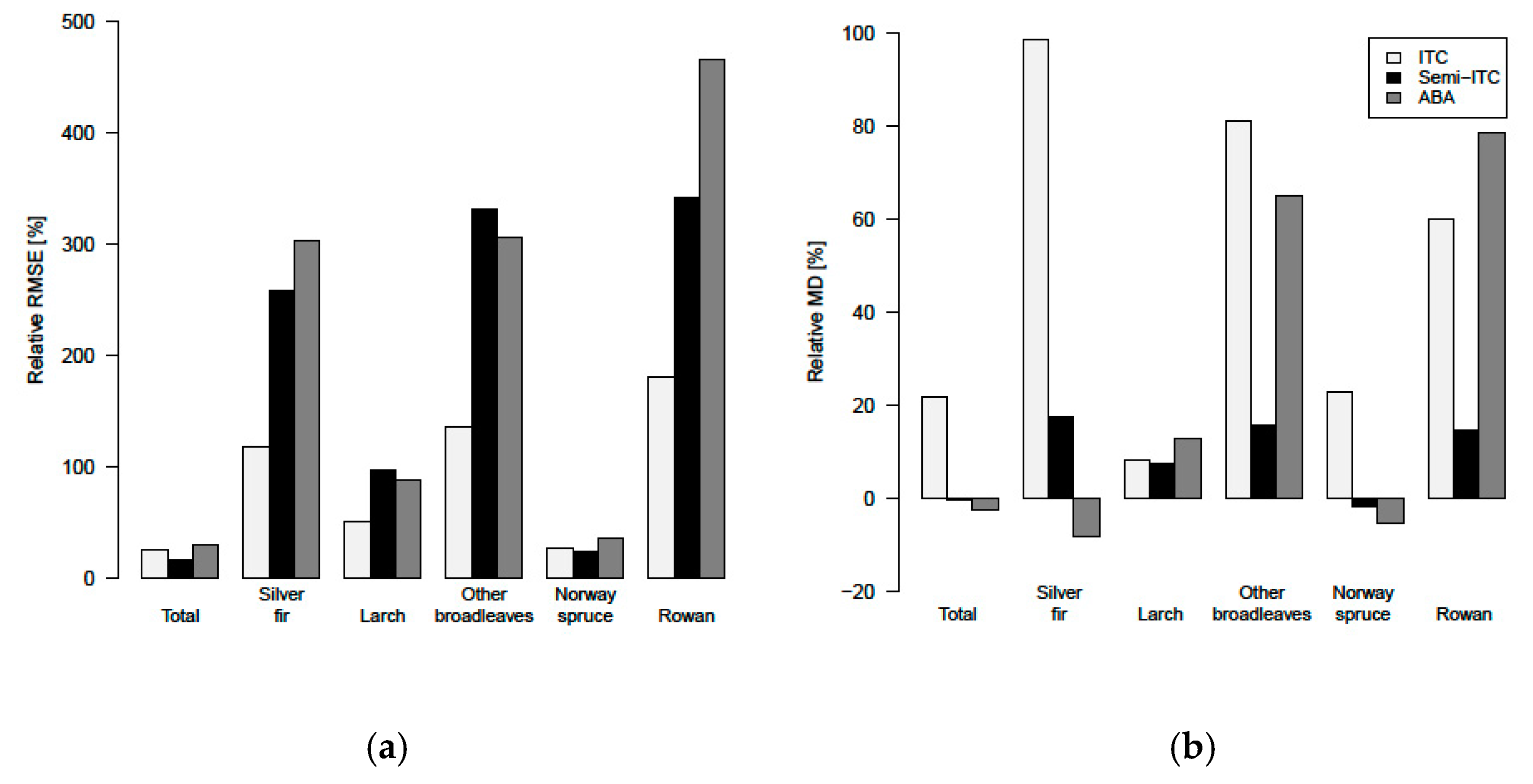
The accuracies of the species-specific volumes of the minority species obtained by the ITC approach were relatively higher compared to the semi-ITC and ABA approaches, but the accuracies of the more frequent species, i.e., the Norway spruce, were very much alike. The semi-ITC approach had slightly smaller RMSEs compared to the ABA, except for the larch and other broadleaves where the RMSEs were similar for both approaches. The largest systematic errors occurred in the ITC approach, caused by non-detected trees. Although all delineated crown segments were used for the volume modeling, a large underprediction due to omission errors of the ITC delineation still appeared in the ITC approach, mainly due to the forest structure [
14]. The semi-ITC approach underpredicted all volumes, except for the Norway spruce volume, which was negligibly overpredicted. Even though, the semi-ITC is compensating the problem related to the omission error, it is not able to eliminate it completely. On average, the species-specific volumes were achieved with the smallest systematic errors with the semi-ITC approach. The ABA approach also underpredicted all volumes, but overpredicted the Norway spruce and silver fir volumes. The overprediction in ABA can be the result of species presence of the nearest sample plots. As the volumes were predicted based on selected nearest neighbors, the
kMSN method can predict small volumes for tree species classes that do not really occur in a sample plot. Since the
kNN method was used, the predicted values will have a smaller range than the observed because of an averaging effect, which can caused the underprediction for the ABA and semi-ITC approach. We have to consider that the systematic error of the semi-ITC approach is the result of the combination of the delineation and
kNN method’s characteristics. In most cases, the systematic errors obtained with the semi-ITC approach were smaller compared to the ABA. A similar observation was also found in the study of Breidenbach et al. [
37].
In the literature, there are no similar studies comparing the species-specific attributes obtained by the ITC and semi-ITC approaches, except for Ørka et al. [
23]. In general, the main problem of the ITC methods is the quite large omission error, caused by undetected understory and suppressed trees [
59], mostly resulting in underprediction [
12] of biophysical attributes when aggregated at the plot level [
60]. This systematic error can be considerably reduced with the semi-ITC approach as demonstrated in our study. Comparing the semi-ITC and ABA inventory approaches, Breidenbach et al. and Rahlf et al. [
37,
61] showed that the semi-ITC approach provided prediction accuracies that were higher or similar to the ABA, while in the study of Ørka et al. [
23], the ABA performed better than the semi-ITC approach. Based on a trade-off between the goodness of fit and the systematic error, the compared total and species-specific results suggest that the semi-ITC approach in total outperformed the other approaches.
For the
kNN methods, a sufficient number of sample plots is required for the ABA in order to have a sufficient number of neighbors available for the calculation of the distance matrix. For example, 300–500 sample plots are usually applied in operational forest inventories. On the other hand, many studies showed that nonparametric models were in line with parametric models also with a lower number of observations (e.g., 200 sample plots) [
44,
62]. However, we assumed that 94 subsample plots using three nearest neighbors were sufficient to achieve good results. The reason to not apply a parametric method was that the field-observed species-specific volumes contained many values close to zero or zeros due to the absence of minority tree species in the sample plots. Moreover, in complex stands with a wide variety of tree sizes and species, the nonparametric methods might be preferred to avoid implausible (e.g., negative) predictions and to obtain a reasonable extrapolation beyond the range of calibration data. In fact, among the nonparametric methods, the
kNN, in particular the
kMSN method, have been shown to be efficient in providing simultaneous multivariate predictions at a satisfactory accuracy level [
17,
37,
54,
63,
64].
The volumes of the minor species were always predicted with smaller accuracies than the volumes of the majority species. The reason could be the small share of some tree species, especially broadleaves, and thus it was difficult to obtain accurate results. Moreover, a large share of only one species is the typical situation of the European forests. This phenomenon of small accuracy of the minor species is typical also for other studies [
23,
37], but despite the small RMSEs, the information on the existence of the minority species in stands can be valuable information, for example in tree-oriented silvicultural practices. Looking at the problem from an economical perspective, minority species are of secondary interest as the majority of the harvested volume is coming from Norway spruce and larch. Other species are mainly used for energy wood, as is the case of the broadleaves volume, or for very specific uses (e.g., high quality furniture). Minority species are important from the ecological perspective and thus, the use of ITC or semi-ITCs approaches is essential in this case as it is difficult to monitor such species with the ABA approach [
23]. The ITC and semi-ITC approaches can also be used to detect and report the presence and spread of non-native invasive species that can irreversibly alter the productivity of the systems they invade. The semi-ITC approach is overall quite good for predicting volumes of the minority and majority species. However, in studies where the volume of majority species is of high interest, the ABA is recommended, as the field data collection (e.g., tree positions) is less demanding and less costly compared to what is required for the semi-ITC approach.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}