Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies



Abstract
1. Introduction
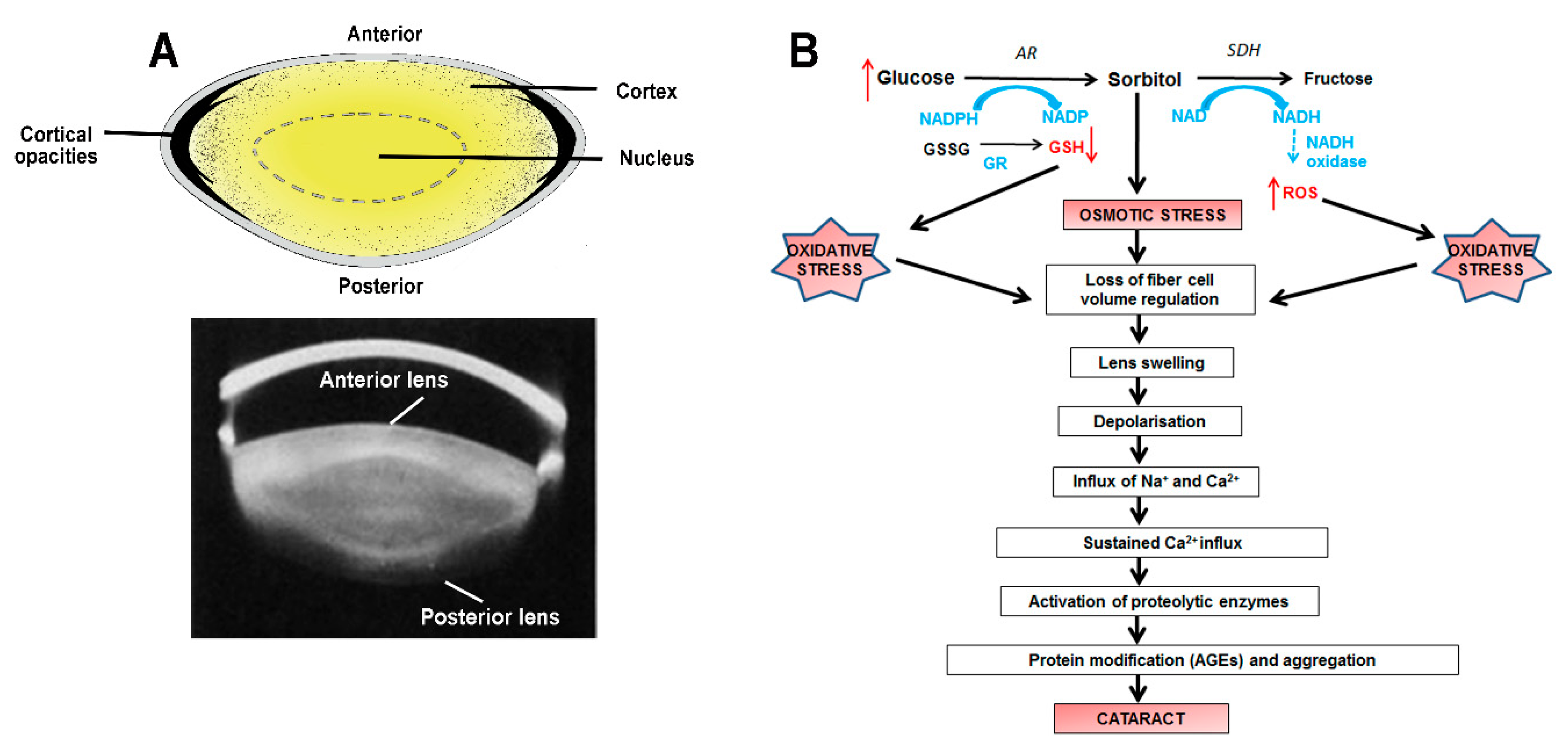
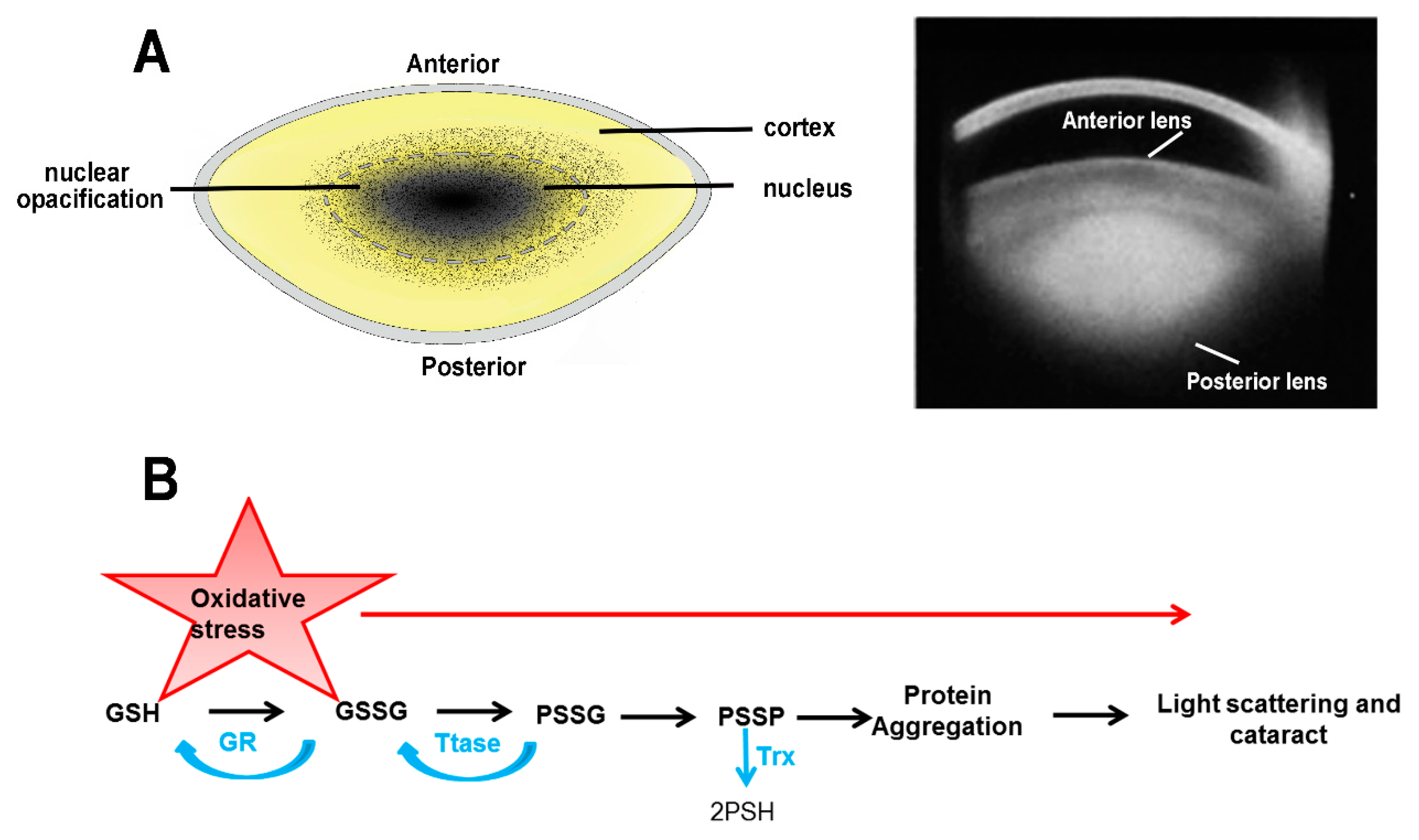
2. The Etiology of Lens Cataract
2.1. Cortical Cataract
2.2. Age-Related Nuclear (ARN) Cataract
3. Evidence for and Against Nutritional Strategies to Prevent Lens Cataract
Effectiveness of Antioxidant Therapies on Cataract Progression—A Survey of the Literature
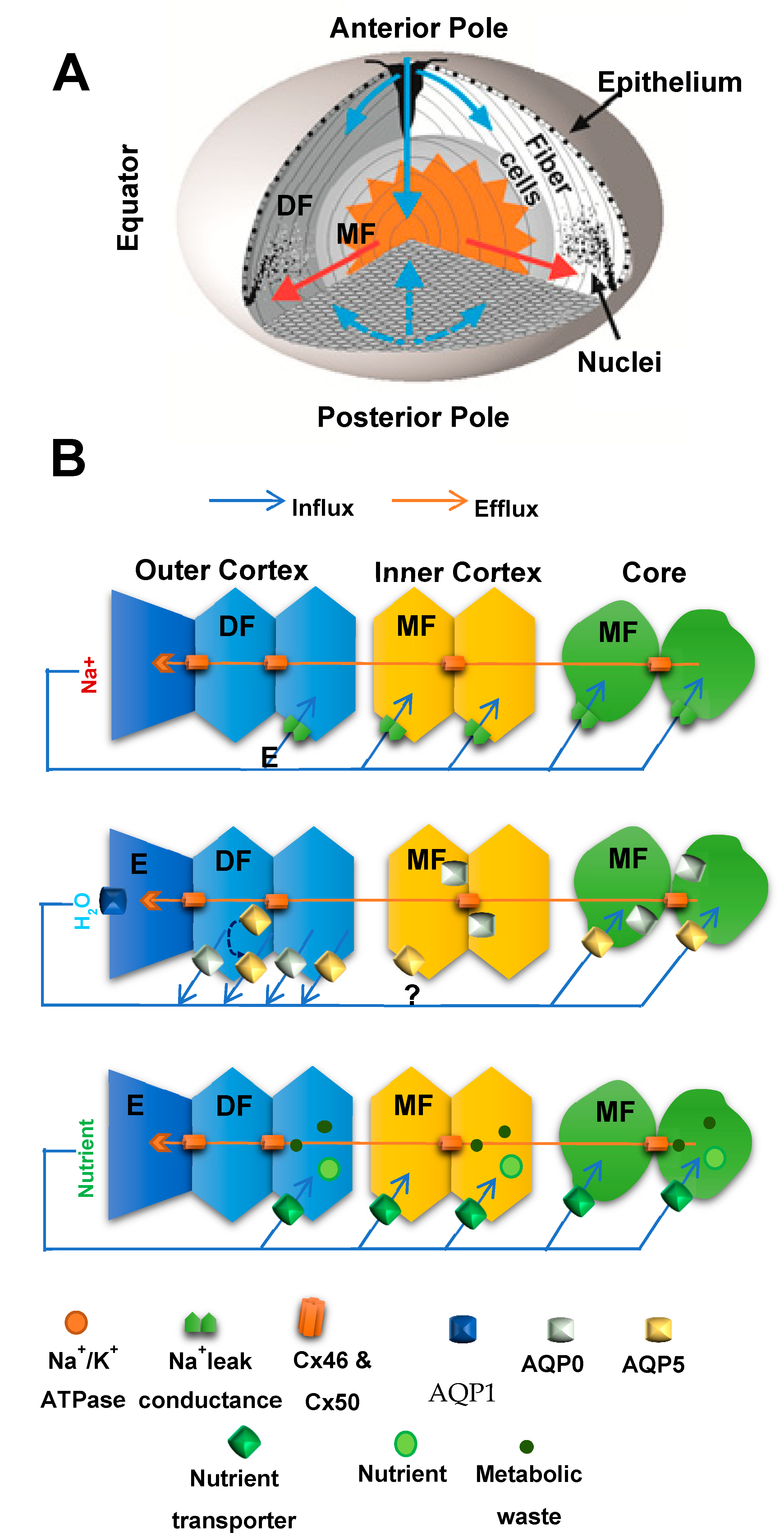
4. Lens Structure and Function
5. Conclusions and Future Strategies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hobbs, R.P.; Bernstein, P.S. Nutrient Supplementation for Age-related Macular Degeneration, Cataract, and Dry Eye. J. Ophthalmic Vis. Res. 2014, 9, 487–493. [Google Scholar] [CrossRef]
- Pascolini, D.; Mariotti, S.P. Global estimates of visual impairment: 2010. Br. J. Ophthalmol 2012, 96, 614–618. [Google Scholar] [CrossRef]
- Weikel, K.A.; Garber, C.; Baburins, A.; Taylor, A. Nutritional modulation of cataract. Nutr. Rev. 2014, 72, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.E.; Klein, R.; Wang, Q.; Moss, S.E. Older-onset diabetes and lens opacities. The Beaver Dam Eye Study. Ophthalm. Epid. 1995, 2, 49–55. [Google Scholar] [CrossRef]
- United Nations Department of Economic and Social Affairs, Population Division. World Population Ageing 2017-Highlights (ST/ESA/SER.A/397); United Nations: New York, NY, USA, 2017. [Google Scholar]
- McCarty, C.A.; Taylor, H.R. The Genetics of Cataract. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1677–1678. [Google Scholar]
- Access Economics. Clear focus—The economic impact of vision loss in New Zealand in 2009. A report prepared for RANZCO and the RNZFB, 2010. [Google Scholar]
- Brian, G.; Taylor, H. Cataract blindness--challenges for the 21st century. Bull. World Health Organ. 2001, 79, 249–256. [Google Scholar]
- Cumming, R.G.; Mitchell, P.; Smith, W. Diet and cataract: The blue mountains eye study. Ophthalmology 2000, 107, 450–456. [Google Scholar] [CrossRef]
- Kuzniarz, M.; Mitchell, P.; Cumming, R.G.; Flood, V.M. Use of vitamin supplements and cataract: The blue mountains eye study. Am. J. Ophthalmol. 2001, 132, 19–26. [Google Scholar] [CrossRef]
- Valero, M.P.; Fletcher, A.E.; de Stavola, B.L.; Vioque, J.S.; Alepuz, V.C.S. Vitamin C Is Associated with Reduced Risk of Cataract in a Mediterranean Population. J. Nutr. 2002, 132, 1299–1306. [Google Scholar] [CrossRef]
- McNeil, J.J.; Robman, L.; Tikellis, G.; Sinclair, M.I.; McCarty, C.A.; Taylor, H.R. Vitamin E supplementation and cataract: Randomized controlled trial. Ophthalmology 2004, 111, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E and beta carotene for age-related cataract and vision loss: AREDS report no. 9. Arch Ophthalmol 2001, 119, 1439–1452. [Google Scholar] [CrossRef]
- Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. Vitamin C supplements and the risk of age-related cataract: A population-based prospective cohort study in women. Am. J. Clin. Nutr. 2009, 91, 487–493. [Google Scholar] [CrossRef]
- Cheng, R.; Feng, Q.; Ortwerth, B.J. LC-MS display of the total modified amino acids in cataract lens proteins and in lens proteins glycated by ascorbic acid in vitro. Biochim. Biophys. Acta (Bba) Mol. Basis Dis. 2006, 1762, 533–543. [Google Scholar] [CrossRef][Green Version]
- Linetsky, M.; Shipova, E.; Cheng, R.; Ortwerth, B.J. Glycation by ascorbic acid oxidation products leads to the aggregation of lens proteins. Biochim. Et Biophys. Acta (Bba) Mol. Basis Dis. 2008, 1782, 22–34. [Google Scholar] [CrossRef]
- Christen, W.G.; Glynn, R.J.; Chew, E.Y.; Albert, C.M.; Manson, J.E. Folic Acid, Vitamin B6, and Vitamin B12 in Combination and Age-Related Cataract in a Randomized Trial of Women. Ophthalmic Epidemiol. 2016, 23, 32–39. [Google Scholar] [CrossRef]
- Camacho-Barcia, M.L.; Bulló, M.; Garcia-Gavilán, J.F.; Ruiz-Canela, M.; Corella, D.; Estruch, R.; Fitó, M.; García-Layana, A.; Arós, F.; Fiol, M.; et al. Association of Dietary Vitamin K1 Intake with the Incidence of Cataract Surgery in an Adult Mediterranean Population: A Secondary Analysis of a Randomized Clinical Trial. JAMA Ophthalmol. 2017, 135, 657–661. [Google Scholar] [CrossRef]
- Bourne, R.R.; Stevens, G.A.; White, R.A.; Smith, J.L.; Flaxman, S.R.; Price, H.; Jonas, J.B.; Keeffe, J.; Leasher, J.; Naidoo, K.; et al. Causes of vision loss worldwide, 1990–2010: A systematic analysis. Lancet Glob Health 2013, 1, e339–e349. [Google Scholar]
- Michael, R.; Bron, A.J. The ageing lens and cataract: A model of normal and pathological ageing. Philos. Trans. R. Soc. Lond. Ser. Biol. Sci. 2011, 366, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Michael, R. Cortical cataract. In Encyclopedia of the Eye; Darlene, A.D., Ed.; Academic Press: Oxford, UK, 2010; Volume 1, pp. 532–536. [Google Scholar]
- Pesudovs, K.; Elliott, D.B. Refractive error changes in cortical, nuclear, and posterior subcapsular cataracts. Br. J. Ophthalmol. 2003, 87, 964–967. [Google Scholar] [CrossRef]
- Planten, J.T. Changes of refraction in the adult eye due to changing refractive indices of the layers of the lens. Ophthalmol. J. Int. D’ophtalmologie Int. J. Ophthalmol. Z. Fur Augenheilkd. 1981, 183, 86–90. [Google Scholar] [CrossRef]
- Al-Ghoul, K.J.; Costello, M.J. Morphological changes in human nuclear cataracts of late-onset diabetics. Exp. Eye Res. 1993, 57, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, J.H. Pathways of glucose metabolism in the lens. Investig. Ophthalmol. 1965, 4, 619–628. [Google Scholar]
- Kinoshita, J.H. Cataracts in galactosemia: The Jonas, S. Friedenwald Memorial Lecture. Investig. Ophthalmol. 1965, 4, 786–799. [Google Scholar]
- Kinoshita, J.H. Mechanisms initiating cataract formation. Proctor Lecture. Investig. Ophthalmol. 1974, 13, 713–724. [Google Scholar]
- Kador, P.F.; Wyman, M.; Oates, P.J. Aldose reductase, ocular diabetic complications and the development of topical Kinostat®. Prog. Retin. Eye Res. 2016. [Google Scholar] [CrossRef]
- Chung, S.S.; Ho, E.C.; Lam, K.S.; Chung, S.K. Contribution of polyol pathway to diabetes-induced oxidative stress. J. Am. Soc. Nephrol. JASN 2003, 14, S233–S236. [Google Scholar] [CrossRef]
- Chan, A.W.; Ho, Y.S.; Chung, S.K.; Chung, S.S. Synergistic effect of osmotic and oxidative stress in slow-developing cataract formation. Exp. Eye Res. 2008, 87, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Pirie, A. Color and solubility of the proteins of human cataracts. Investig. Ophthalmol. 1968, 7, 634–650. [Google Scholar]
- Al-Ghoul, K.J.; Costello, M.J. Fiber cell morphology and cytoplasmic texture in cataractous and normal human lens nuclei. Curr. Eye Res. 1996, 15, 533–542. [Google Scholar] [CrossRef]
- Costello, M.J.; Oliver, T.N.; Cobo, L.M. Cellular architecture in age-related human nuclear cataracts. Invest Ophthalmol. Vis. Sci. 1992, 33, 3209–3227. [Google Scholar] [PubMed]
- Garner, M.H.; Spector, A. Sulfur oxidation in selected human cortical cataracts and nuclear cataracts. Exp. Eye Res. 1980, 31, 361–369. [Google Scholar] [CrossRef]
- Garner, M.H.; Spector, A. Selective oxidation of cysteine and methionine in normal and senile cataractous lenses. Proc. Natl. Acad. Sci. USA 1980, 77, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.; Roy, D. Disulfide-linked high molecular weight protein associated with human cataract. Proc. Natl. Acad. Sci. USA 1978, 75, 3244–3248. [Google Scholar] [CrossRef] [PubMed]
- Truscott, R.J.; Augusteyn, R.C. The state of sulphydryl groups in normal and cataractous human lenses. Exp. Eye Res. 1977, 25, 139–148. [Google Scholar] [CrossRef]
- Lou, M.F.; Dickerson, J.E., Jr.; Garadi, R. The role of protein-thiol mixed disulfides in cataractogenesis. Exp. Eye Res. 1990, 50, 819–826. [Google Scholar] [CrossRef]
- Lou, M.F.; Dickerson, J.E., Jr.; Tung, W.H.; Wolfe, J.K.; Chylack, L.T., Jr. Correlation of nuclear color and opalescence with protein S-thiolation in human lenses. Exp. Eye Res. 1999, 68, 547–552. [Google Scholar] [CrossRef]
- Lim, J.C.; Umapathy, A.; Donaldson, P.J. Tools to fight the cataract epidemic: A review of experimental animal models that mimic age related nuclear cataract. Exp. Eye Res. 2016, 145, 432–443. [Google Scholar] [CrossRef]
- Lou, M.F. Redox regulation in the lens. Prog. Retin. Eye Res. 2003, 22, 657–682. [Google Scholar] [CrossRef]
- Truscott, R.J. Age-related nuclear cataract-oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar] [CrossRef]
- Reddy, V.N. Glutathione and its function in the lens--an overview. Exp. Eye Res. 1990, 50, 771–778. [Google Scholar] [CrossRef]
- Rathbun, W.B. Glutathione Biosynthesis in the Lens and Erythrocyte; Elsevier/North Holland: Amsterdam, The Netherlands, 1980; pp. 169–173. [Google Scholar]
- Kannan, R.; Yi, J.R.; Zlokovic, B.V.; Kaplowitz, N. Molecular characterization of a reduced glutathione transporter in the lens. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1785–1792. [Google Scholar]
- Mackic, J.B.; Jinagouda, S.; McComb, G.J.; Weiss, M.H.; Kannan, R.A.M.; Kaplowitz, N.; Zlokovic, B.V. Transport of Circulating Reduced Glutathione at the Basolateral Side of the Anterior Lens Epithelium: Physiologic Importance and Manipulations. Exp. Eye Res. 1996, 62, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Giblin, F.J. Glutathione: A Vital Lens Antioxidant. J. Ocul. Pharmacol. Ther. 2000, 16, 121. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Monnier, V.M.; Whitson, J. Lens glutathione homeostasis: Discrepancies and gaps in knowledge standing in the way of novel therapeutic approaches. Exp. Eye Res. 2017, 156, 103–111. [Google Scholar] [CrossRef]
- Donaldson, P.J.; Musil, L.S.; Mathias, R.T. Point: A Critical Appraisal of the Lens Circulation Model—An Experimental Paradigm for Understanding the Maintenance of Lens Transparency? Investig. Ophthalmol. Vis. Sci. 2010, 51, 2303. [Google Scholar] [CrossRef]
- Rathbun, W.B.; Murray, D.L. Age-related cysteine uptake as rate-limiting in glutathione synthesis and glutathione half-life in the cultured human lens. Exp. Eye Res. 1991, 53, 205–212. [Google Scholar] [CrossRef]
- Sweeney, M.H.J.; Truscott, R.J.W. An impediment to glutathione diffusion in older normal human lenses: A possible precondition for nuclear cataract. Exp. Eye Res. 1998, 67, 587–595. [Google Scholar] [CrossRef]
- Truscott, R.J. Age-related nuclear cataract: A lens transport problem. Ophthalmic Res. 2000, 32, 185–194. [Google Scholar] [CrossRef]
- Zhang, H.; Jay Forman, H.; Choi, J. Υ-Glutamyl Transpeptidase in Glutathione Biosynthesis. In Methods in Enzymology; Sies, H., Packer, L., Eds.; Academic Press: Waltham, MA, USA, 2005; Volume 401, pp. 468–483. [Google Scholar]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double-edged swords in cellular redox state Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid Med. Cell Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef]
- Agte, V.; Tarwadi, K. The Importance of Nutrition in the Prevention of Ocular Disease with Special Reference to Cataract. Ophthalmic Res. 2010, 44, 166–172. [Google Scholar] [CrossRef]
- Cui, Y.-H.; Jing, C.-X.; Pan, H.-W. Association of blood antioxidants and vitamins with risk of age-related cataract: A meta-analysis of observational studies. Am. J. Clin. Nutr. 2013, 98, 778–786. [Google Scholar] [CrossRef]
- Fernandez, M.M.; Afshari, N.A. Nutrition and the prevention of cataracts. Curr. Opin. Ophthalmol. 2008, 19, 66–70. [Google Scholar] [CrossRef]
- Fletcher, A.E. Free Radicals, Antioxidants and Eye Diseases: Evidence from Epidemiological Studies on Cataract and Age-Related Macular Degeneration. Ophthalmic Res. 2010, 44, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Mathew, M.C.; Ervin, A.M.; Tao, J.; Davis, R.M. Antioxidant vitamin supplementation for preventing and slowing the progression of age-related cataract. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef] [PubMed]
- McCusker, M.M.; Durrani, K.; Payette, M.J.; Suchecki, J. An eye on nutrition: The role of vitamins, essential fatty acids, and antioxidants in age-related macular degeneration, dry eye syndrome, and cataract. Clin. Dermatol. 2016, 34, 276–285. [Google Scholar] [CrossRef]
- Moeller, S.M.; Jacques, P.F.; Blumberg, J.B. The Potential Role of Dietary Xanthophylls in Cataract and Age-Related Macular Degeneration. J. Am. Coll. Nutr. 2000, 19, 522S–527S. [Google Scholar] [CrossRef]
- Raman, R.; Vaghefi, E.; Braakhuis, A.J. Food components and ocular pathophysiology: A critical appraisal of the role of oxidative mechanisms. Asia Pac. J. Clin. Nutr. 2017, 26, 572–585. [Google Scholar] [CrossRef]
- Seddon, J.M. Multivitamin-multimineral supplements and eye disease: Age-related macular degeneration and cataract. Am. J. Clin. Nutr. 2007, 85, 304S–307S. [Google Scholar] [CrossRef] [PubMed]
- Siegal, M.; Chiu, C.-J.; Taylor, A. Antioxidant Status and Risk for Cataract. In Preventive Nutrition: The Comprehensive Guide for Health Professionals; Bendich, A., Deckelbaum, R.J., Eds.; Humana Press: Totowa, NJ, USA, 2005; pp. 463–503. [Google Scholar]
- Trumbo, P.R.; Ellwood, K.C. Lutein and zeaxanthin intakes and risk of age-related macular degeneration and cataracts: An evaluation using the Food and Drug Administration’s evidence-based review system for health claims. Am. J. Clin. Nutr. 2006, 84, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.-Q.; Li, L.-M.; Zhu, H. The Effect of Multivitamin/Mineral Supplements on Age-Related Cataracts: A Systematic Review and Meta-Analysis. Nutrients 2014, 6, 931. [Google Scholar] [CrossRef] [PubMed]
- Christen, W.G.; Liu, S.; Glynn, R.J.; Gaziano, J.M.; Buring, J.E. Dietary carotenoids, vitamins C and E, and risk of cataract in women: A prospective study. Arch. Ophthalmol. 2008, 126, 102–109. [Google Scholar] [CrossRef]
- Dherani, M.; Murthy, G.V.S.; Gupta, S.K.; Young, I.S.; Maraini, G.; Camparini, M.; Price, G.M.; John, N.; Chakravarthy, U.; Fletcher, A.E. Blood Levels of Vitamin C, Carotenoids and Retinol Are Inversely Associated with Cataract in a North Indian Population. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3328–3335. [Google Scholar] [CrossRef] [PubMed]
- Jacques, P.F.; Chylack, L.T., Jr.; Hankinson, S.E.; Khu, P.M.; Rogers, G.; Friend, J.; Tung, W.; Wolfe, J.K.; Padhye, N.; Willett, W.C.; et al. Long-term Nutrient Intake and Early Age-Related Nuclear Lens Opacities. Arch. Ophthalmol. 2001, 119, 1009–1019. [Google Scholar] [CrossRef]
- Pastor-Valero, M. Fruit and vegetable intake and vitamins C and E are associated with a reduced prevalence of cataract in a Spanish Mediterranean population. BMC Ophthalmol. 2013, 13, 52. [Google Scholar] [CrossRef]
- Ravindran, R.D.; Vashist, P.; Gupta, S.K.; Young, I.S.; Maraini, G.; Camparini, M.; Jayanthi, R.; John, N.; Fitzpatrick, K.E.; Chakravarthy, U.; et al. Inverse Association of Vitamin C with Cataract in Older People in India. Ophthalmology 2011, 118, 1958–1965.e1952. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.G.; Rochtchina, E.; Burlutsky, G.; Mitchell, P.; Cumming, R.G.; Flood, V.M.; Wang, J.J. Antioxidant nutrient intake and the long-term incidence of age-related cataract: The Blue Mountains Eye Study. Am. J. Clin. Nutr. 2008, 87, 1899–1905. [Google Scholar] [CrossRef]
- Theodoropoulou, S.; Samoli, E.; Theodossiadis, P.G.; Papathanassiou, M.; Lagiou, A.; Lagiou, P.; Tzonou, A. Diet and cataract: A case–control study. Int. Ophthalmol. 2014, 34, 59–68. [Google Scholar] [CrossRef]
- Yoshida, M.; Takashima, Y.; Inoue, M.; Iwasaki, M.; Otani, T.; Sasaki, S.; Tsugane, S.; JPHC Study Group. Prospective study showing that dietary vitamin C reduced the risk of age-related cataracts in a middle-aged Japanese population. Eur. J. Nutr. 2007, 46, 118–124. [Google Scholar] [CrossRef]
- Delcourt, C.C.; Carriére, I.; Delage, M.; Barberger-Gateau, P.; Schalch, W.; POLA Study Group. Plasma Lutein and Zeaxanthin and Other Carotenoids as Modifiable Risk Factors for Age-Related Maculopathy and Cataract: The POLA Study. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2329–2335. [Google Scholar] [CrossRef]
- Moeller, S.M.; Voland, R.; Tinker, L.; Blodi, B.A.; Klein, M.L.; Gehrs, K.M.; Johnson, E.J.; Snodderly, D.M.; Wallace, R.B.; Chappell, R.J.; et al. Associations Between Age-Related Nuclear Cataract and Lutein and Zeaxanthin in the Diet and Serum in the Carotenoids in the Age-Related Eye Disease Study (CAREDS), an Ancillary Study of the Women’s Health Initiative. Arch. Ophthalmol. 2008, 126, 354–364. [Google Scholar] [CrossRef]
- Mares, J.A.; Voland, R.; Adler, R.; Tinker, L.; Millen, A.E.; Moeller, S.M.; Blodi, B.; Gehrs, K.M.; Wallace, R.B.; Chappell, R.J.; et al. Healthy Diets and the Subsequent Prevalence of Nuclear Cataract in WomenHealthy Diets and Nuclear Cataract in Women. Arch. Ophthalmol. 2010, 128, 738–749. [Google Scholar] [CrossRef]
- Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. Total Antioxidant Capacity of the Diet and Risk of Age-Related Cataract: A Population-Based Prospective Cohort of WomenAntioxidant Capacity of Diet and Risk of CataractsAntioxidant Capacity of Diet and Risk of Cataracts. JAMA Ophthalmol. 2014, 132, 247–252. [Google Scholar] [CrossRef]
- Zheng Selin, J.; Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. High-Dose Supplements of Vitamins C and E, Low-Dose Multivitamins, and the Risk of Age-related Cataract: A Population-based Prospective Cohort Study of Men. Am. J. Epidemiol. 2013, 177, 548–555. [Google Scholar] [CrossRef]
- Group, T.R.; Chylack, L.T.; Brown, N.P.; Bron, A.; Hurst, M.; Köpcke, W.; Thien, U.; Schalch, W. The Roche European American Cataract Trial (REACT): A randomized clinical trial to investigate the efficacy of an oral antioxidant micronutrient mixture to slow progression of age-related cataract. Ophthalmic Epidemiol 2002, 9, 49–80. [Google Scholar] [CrossRef]
- Mares-Perlman, J.A.; Lyle, B.J.; Klein, R.; Fisher, A.I.; Brady, W.E.; VandenLangenberg, G.M.; Trabulsi, J.N.; Palta, M. Vitamin Supplement Use and Incident Cataracts in a Population-Based Study. Arch. Ophthalmol. 2000, 118, 1556–1563. [Google Scholar] [CrossRef]
- Group, A.-R.E.D.S.R. Centrum Use and Progression of Age-Related Cataract in the Age-Related Eye Disease Study: A Propensity Score Approach. AREDS Report No. 21. Ophthalmology 2006, 113, 1264–1270. [Google Scholar] [CrossRef]
- Christen, W.; Glynn, R.; Sperduto, R.; Chew, E.; Buring, J. Age-related cataract in a randomized trial of beta-carotene in women. Ophthalmic Epidemiol. 2004, 11, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Christen, W.G.; Glynn, R.J.; Chew, E.Y.; Buring, J.E. Vitamin E and Age-Related Cataract in a Randomized Trial of Women. Ophthalmology 2008, 115, 822–829.e821. [Google Scholar] [CrossRef]
- Christen, W.G.; Glynn, R.J.; Gaziano, J.M.; Darke, A.K.; Crowley, J.J.; Goodman, P.J.; Lippman, S.M.; Lad, T.E.; Bearden, J.D.; Goodman, G.E.; et al. Age-Related Cataract in Men in the Selenium and Vitamin E Cancer Prevention Trial Eye Endpoints Study: A Randomized Clinical TrialAge-Related Cataract and Selenium and Vitamin E in MenAge-Related Cataract and Selenium and Vitamin E in Men. JAMA Ophthalmol. 2015, 133, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Christen, W.G.; Glynn, R.J.; Sesso, H.D.; Kurth, T.; MacFadyen, J.; Bubes, V.; Buring, J.E.; Manson, J.E.; Gaziano, J.M. Age-Related Cataract in a Randomized Trial of Vitamins E and C in Men Vitamins E and C and Cataract. Arch. Ophthalmol. 2010, 128, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Christen, W.G.; Manson, J.E.; Glynn, R.J.; Gaziano, J.M.; Sperduto, R.D.; Buring, J.E.; Hennekens, C.H. A Randomized Trial of Beta Carotene and Age-Related Cataract in US Physicians. Arch. Ophthalmol. 2002, 121, 372–378. [Google Scholar] [CrossRef]
- Ferrigno, L.; Aldigeri, R.; Rosmini, F.; Sperduto, R.D.; Maraini, G. Associations Between Plasma Levels of Vitamins and Cataract in the Italian-American Clinical Trial of Nutritional Supplements and Age-Related Cataract (CTNS): CTNS Report #2. Ophthalmic Epidemiol. 2005, 12, 71–80. [Google Scholar] [CrossRef]
- Bassnett, S. Lens organelle degradation. Exp. Eye Res. 2002, 74, 1–6. [Google Scholar] [CrossRef]
- Menko, A.S. Lens epithelial cell differentiation. Exp. Eye Res. 2002, 75, 485–490. [Google Scholar] [CrossRef]
- Kuszak, J.R.; Zoltoski, R.K.; Tiedemann, C.E. Development of lens sutures. Int. J. Dev. Biol. 2004, 48, 889–902. [Google Scholar] [CrossRef]
- Donaldson, P.; Kistler, J.; Mathias, R.T. Molecular solutions to mammalian lens transparency. News Physiol. Sci. 2001, 16, 118–123. [Google Scholar] [CrossRef]
- Mathias, R.T.; Kistler, J.; Donaldson, P. The lens circulation. J. Membr Biol 2007, 216, 1–16. [Google Scholar] [CrossRef]
- Mathias, R.T.; Rae, J.L.; Baldo, G.J. Physiological properties of the normal lens. Physiol. Rev. 1997, 77, 21–50. [Google Scholar] [CrossRef]
- Gao, J.; Sun, X.; Moore, L.C.; White, T.W.; Brink, P.R.; Mathias, R.T. Lens intracellular hydrostatic pressure is generated by the circulation of sodium and modulated by gap junction coupling. J. Gen. Physiol. 2011, 137, 507–520. [Google Scholar] [CrossRef]
- Beebe, D.C.; Truscott, R.J.W. Counterpoint: The Lens Fluid Circulation Model—A Critical Appraisal. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2306–2310. [Google Scholar] [CrossRef]
- Candia, O.A.; Mathias, R.; Gerometta, R. Fluid circulation determined in the isolated bovine lens. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7087–7096. [Google Scholar] [CrossRef]
- Vaghefi, E.; Pontre, B.P.; Jacobs, M.D.; Donaldson, P.J. Visualizing ocular lens fluid dynamics using MRI: Manipulation of steady state water content and water fluxes. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 301, R335–R342. [Google Scholar] [CrossRef]
- Vaghefi, E.; Walker, K.; Pontre, B.P.; Jacobs, M.D.; Donaldson, P.J. Magnetic resonance and confocal imaging of solute penetration into the lens reveals a zone of restricted extracellular space diffusion. Am. J. Physiol.-Regul. Integr. Compar. Physiol. 2012, 302, R1250–R1259. [Google Scholar] [CrossRef]
- Vaghefi, E.; Donaldson, P.J. The lens internal microcirculation system delivers solutes to the lens core faster than would be predicted by passive diffusion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R994–R1002. [Google Scholar] [CrossRef]
- Li, L.; Lim, J.; Jacobs, M.D.; Kistler, J.; Donaldson, P.J. Regional differences in cystine accumulation point to a sutural delivery pathway to the lens core. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1253–1260. [Google Scholar] [CrossRef]
- Lim, J.; Lam, Y.C.; Kistler, J.; Donaldson, P.J. Molecular characterization of the cystine/glutamate exchanger and the excitatory amino acid transporters in the rat lens. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2869–2877. [Google Scholar] [CrossRef]
- Lim, J.; Li, L.; Jacobs, M.D.; Kistler, J.; Donaldson, P.J. Mapping of glutathione and its precursor amino acids reveals a role for GLYT2 in glycine uptake in the lens core. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5142. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Lorentzen, K.A.; Kistler, J.; Donaldson, P.J. Molecular identification and characterisation of the glycine transporter (GLYT1) and the glutamine/glutamate transporter (ASCT2) in the rat lens. Exp. Eye Res. 2006, 83, 447–455. [Google Scholar] [CrossRef]
- Merriman-Smith, B.R.; Krushinsky, A.; Kistler, J.; Donaldson, P.J. Expression patterns for glucose transporters glut1 and glut3 in the normal rat lens and in models of diabetic cataract. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3458–3466. [Google Scholar] [CrossRef]
- Merriman-Smith, R.; Donaldson, P.; Kistler, J. Differential expression of facilitative glucose transporters GLUT1 and GLUT3 in the lens. Investig. Ophthalmol. Vis. Sci. 1999, 40, 3224. [Google Scholar]
- Nye-Wood, M.G.; Spraggins, J.M.; Caprioli, R.M.; Schey, K.L.; Donaldson, P.J.; Grey, A.C. Spatial distributions of glutathione and its endogenous conjugates in normal bovine lens and a model of lens aging. Exp. Eye Res. 2017, 154, 70–78. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Vitamin C | ||||
| Author | Any CAT | NUC | CX | PSC |
| Christen, 2008 [71] | RR 1.00; 95% CI 0.86–1.16; P = 0.61 | - | - | - |
| Dherani, 2008 [72] | OR 0.64; 95% CI 0.48–0.85; P < 0.01 | OR 0.62; 95% CI 0.4–0.96; P = 0.06 | OR 0.62; 95% CI 0.40–0.97; P = 0.10 | OR 0.59; 95% CI 0.35–0.99; P = 0.10 |
| Jaques, 2001 [73] | - | OR 0.31; 95% CI 0.16–0.58; P = 0.003 | - | - |
| Pastor-Valero, 2013 [74] | OR 0.46; 95% CI 0.24–0.88; P = 0.047 | - | - | - |
| Ravindran, 2011 [75] | Plasma levels: OR 0.61; 95% CI 0.57–0.82; P < 0.0001Dietary OR 0.78; 95% CI 0.62–0.98; P = 0.006 | OR 0.66; 95% CI 0.54–0.80; P < 0.0001 | OR 0.70; 95% CI 0.54–0.90; P < 0.002 | OR 0.58; 95% CI 0.45–0.74; P < 0.00003 |
| Tan, 2008 [76] | - | OR 0.55; 95% CI 0.36–0.86; P = 0.045 | OR 0.94; 95% CI 0.63–1.40; P = 0.698 | OR 1.15; 95% CI 0.06–2.23; P = 0.954 |
| Theodoropoulou, 2014 [77] | OR 0.50; 95% CI 0.39–0.64; P < 0.001 | OR 0.55; 95% CI 0.41–0.72; P < 0.001 | OR 0.62; 95% CI 0.37–1.04; P = 0.071 | OR 0.30; 95% CI 0.19–0.49; P<0.001 |
| Valero, 2002 [12] | OR 0.70; 95% CI 0.44–1.13; P = 0.04 | OR 0.56; 95% CI 0.38–0.82 | OR 0.92; 95% CI 0.60–1.40 | OR 0.75; 95% CI 0.51–1.05 |
| Yoshida, 2007 [78] | Men: OR 0.65; 95% CI 0.42–0.97; P = 0.094 Women: OR 0.59; 95% CI 0.43–0.89; P = 0.047 | - | - | - |
| Vitamin E | ||||
| Author | Any CAT | NUC | CX | PSC |
| Christen, 2008 [71] | RR 0.92; 95% CI 0.80–1.06; P = 0.39 | - | - | - |
| Jaques, 2001 [73] | - | OR 0.45; 95% CI 0.23–0.86; P = 0.06 | - | - |
| Pastor-Valero, 2013 [74] | OR 0.46; 95% CI 0.24–0.88; P = 0.944 | - | - | - |
| Tan, 2008 [76] | - | OR 0.73; 95% CI 0.47–1.13; P = 0.155 | OR 0.91; 95% CI 0.62–1.33; P = 0.944 | OR 0.95; 95% CI 0.50-1.83; P=0.597 |
| Theodoropoulou, 2014 [77] | OR 0.50; 95% CI 0.38–0.66; P < 0.001 | OR 0.50; 95% CI 0.36–0.69; P < 0.001 | OR 0.71; 95% CI 0.41–1.25; P = 0.238 | OR 0.42; 95% CI 0.26–0.68; P < 0.001 |
| Valero, 2002 [12] | OR 0.77; 95% CI 0.48–1.24; P = 0.60 | OR 0.81; 95% CI 0.50–1.28 | OR 1.00; 95% CI 0.59–1.72 | OR 1.16; 95% CI 0.71–1.90 |
| Vitamin A (Retinol) | ||||
| Author | Any CAT | NUC | CX | PSC |
| Dherani, 2008 [72] | OR 0.58; 95% CI 0.37–0.91; P < 0.02 | OR 0.56; 95% CI 0.33–0.46; P = 0.04 | OR 0.69; 95% CI 0.38–1.26; P = 0.20 | OR 0.69; 95% CI 0.39–1.23; P = 0.20 |
| Tan, 2008 [76] | - | OR 066; 95% CI 0.42–10.3; P = 0.056 | OR 0.84; 95% CI 0.56–1.25; P = 0.305 | OR 1.04; 95% CI 0.54–2.02; P = 0.604 |
| Theodoropoulou, 2014 [77] | OR 1.47; 95% CI 1.150–1.88; P = 0.002 | OR 1.46; 95% CI 1.11–1.92; P = 0.007 | OR 1.02; 95% CI 0.51–2.02; P = 0.962 | OR 1.88; 95% CI 1.35–2.63; P < 0.001 |
| Valero, 2002 [12] | OR 0.82; 95% CI 0.50–1.03; P = 0.21 | Plasma levels: OR 1.67; 95% CI 1.02–2.72 | Plasma levels: OR 1.82; 95% CI 1.09–3.08 | Plasma levels: OR 1.22; 95% CI 0.73–2.03 |
| Selenium | ||||
| Author | Any CAT | NUC | CX | PSC |
| Valero, 2002 [12] | OR 0.97; 95% CI 0.60–1.58; P = 0.34 | OR 0.71; 95% CI 0.48–1.04 | OR 0.88; 95% CI 0.58–2.46 | OR 1.03; 95% CI 0.70–1.51 |
| Carotenoids | ||||
| Author | Any CAT | NUC | CX | PSC |
| Christen, 2008 [71] | Lutein/zeaxanthin: RR 0.82; 95% CI 0.71–0.95; P = 0.045 Alpha-carotene: RR 0.96; 95% CI 0.84–1.11; P = 0.77 Beta-carotene: RR 0.89; 95% CI 0.77–1.02; P = 0.27 Beta-cryptoxanthin: RR 0.92; 95% CI 0.80–1.06; P = 0.19 Lycopene: RR 0.96; 95% CI 0.52–30.7; P = 0.6 | - | - | - |
| Delcourt, 2006 [79] | Plasma levels:Lutein: OR 0.82; 95% CI 0.48–1.41; P = 0.48 Zeaxanthin: OR 0.57; 95% CI 0.34–0.95; P = 0.03 Beta-carotene: OR 0.69; 95% CI 0.40–1.19; P = 0.17 Alpha-carotene: OR 0.69; 95% CI 0.4–1.19; P = 0.17 Beta-cryptoxanthin: OR 0.71; 95% CI 0.42–1.20; P = 0.20 Lycopene: OR 1.17; 95% CI 0.68–2.01; P = 0.58 | Plasma levels:Lutein: OR 0.60; 95% CI 0.24–1.47; P = 0.26 Zeaxanthin: OR 0.25; 95% CI 0.08–0.71; P = 0.004 Beta-carotene: OR 0.42; 95% CI 0.16–1.12; P = 0.07 Alpha-carotene: OR 0.76; 95% CI 0.29–2.03; P = 0.52 Beat-cryptoxanthin: OR 0.70; 95% CI 0.29–1.68; P = 0.40 Lycopene: OR 1.01; 95% CI 0.41–2.51; P = 0.99 | Plasma levels:Lutein: OR 0.75; 95% CI 0.23–2.47; P = 0.63 Zeaxanthin: OR 1.09; 95% CI 0.37–3.26; P = 0.83 Beta-carotene: OR 1.10; 95% CI 0.28–4.26; P = 0.93 Alpha-carotene: OR 0.97; 95% CI 0.32–2.91; P = 0.96 Beta-cryptoxanthin: OR 1.49; 95% CI 0.48–4.68; P = 0.49 Lycopene: OR 0.92; 95% CI 0.27–3.13; P = 0.59 | Plasma levels:Lutein: OR 1.26; 95% CI 0.52–3.07; P = 0.60 Zeaxanthin: OR 0.84; 95% CI 0.34–2.07; P = 0.68 Beta-carotene: OR 0.51; 95% CI 0.19–1.36; P = 0.16 Alpha-carotene: OR 0.72; 95% CI 0.30–1.73; P = 0.46 Beta-cryptoxanthin: OR 0.42; 95% CI 0.15–1.18; P = 0.09 Lycopene: OR 1.19; 95% CI 0.49–2.88; P = 0.70 |
| Dherani, 2008 [72] | Lutein: OR 0.66; 95% CI 0.43–1.02; P = 0.06 Zeaxanthin: OR 0.66; 95% CI 0.45–0.96; P < 0.03 Beta-cryptoxanthin: OR 0.88; 95% CI 0.63–1.23; P = 0.50 Alpha-carotene: OR 0.69; 95% CI 0.50–0.95; P < 0.05 Beta-carotene: OR 0.77; 95% CI 0.45–1.32; P = 0.30 Lycopene: OR 0.78; 95% CI 0.49–1.23; P = 0.20 Alpha-tocopherol: OR 0.58; 95% CI 0.36–0.94; P = 0.04 Gamma-tocopherol: OR 0.75; 95% CI 0.57–0.98; P = 0.06 | Lutein: OR 0.75; 95% CI 0.44–1.31; P = 0.30 Zeaxanthin: OR 0.71; 95% CI 0.43–1.17; P = 0.20 Beta-cryptoxanthin: OR 0.83; 95% CI 0.56–1.22; P = 0.30 Alpha-carotene: OR 0.74; 95% CI 0.48–1.14; P = 0.20 Beta-carotene: OR 0.75; 95% CI 0.44–1.26; P = 0.20 Lycopene: OR 0.83; 95% CI 0.50–1.37; P = 0.40 Alpha-tocopherol: OR 0.60; 95% CI 0.30–1.20; P = 0.10 Gamma-tocopherol: OR 0.99; 95% CI 0.62–1.58; P = 0.90 | Lutein: OR 0.53; 95% CI0.28–1.02; P = 0.10 Zeaxanthin: OR 0.58; 95% CI 0.30–1.12; P = 0.10 Beta-cryptoxanthin: OR 0.93; 95% CI 0.54–1.56; P = 0.90 Alpha-carotene: OR 0.86; 95% CI 0.60–1.22; P = 0.70 Beta-carotene: OR 1.02; 95% CI 0.52–1.99; P = 0.90 Lycopene: OR 1.17; 95% CI 0.54–2.53; P = 0.70 Alpha-tocopherol: OR 0.58; 95% CI 0.25–1.35; P = 0.20 Gamma-tocopherol: OR 0.84; 95% CI 0.44–1.00; P = 0.50 | Lutein: OR 0.72; 95% CI 0.30–1.71; P = 0.40 Zeaxanthin: OR 0.84; 95% CI 0.43–1.67; P = 0.60 Beta-cryptoxanthin: OR 1.06; 95% CI 0.54–2.05; P = 0.80 Alpha-carotene: OR 0.80; 95% CI 0.39–1.65; P = 0.60 Beta-carotene: OR 0.71; 95% CI 0.30–1.68; P = 0.40 Lycopene: OR 0.68; 95% CI 0.29–1.61; P = 0.30 Alpha-tocopherol: OR 1.11; 95% CI 0.42–2.91; P = 0.90 Gamma-tocopherol: OR 1.16; 95% CI 0.69–1.93; P = 0.60 |
| Jaques, 2001 [73] | - | Alpha-carotene: OR 0.71; 95% CI 0.37–1.35; P = 0.39 Beta-carotene: OR 0.52; 95% CI 0.28–0.97; P = 0.08 Beta-cryptoxanthin: OR 0.68; 95% CI 0.34–1.35; P = 0.06 Lutein/zeaxanthin: OR 0.52; 95% CI 0.29–0.91; P = 0.08 Lycopene: OR 1.16; 95% CI 0.63–2.16; P = 0.79 | - | - |
| Moeller, 2008 [80] | - | Lutein: OR 0.68; 95% CI 0.48–0.97; P = 0.04 Zeaxanthin: OR 0.68; 95% CI 0.47–0.98; P = 0.01 | - | - |
| Tan, 2008 [76] | - | Beta-carotene: OR 1.09; 95% CI 0.69–1.72; P = 0.715 | Beta-carotene: OR 1.06; 95% CI 0.7–1.6; P = 0.854 | Beta-carotene: OR 0.76; 95% CI 0.37–1.59; P = 0.317 |
| Theodoropoulou, 2014 [77] | Carotene: OR 0.56; 95% CI 0.45–0.69; P < 0.001 | Carotene: OR 0.50; 95% CI 0.39–0.65; P < 0.001 | Carotene: OR 0.68; 95% CI 0.43–1.05; P = 0.084 | Carotene: OR 0.58; 95% CI 0.40–0.86; P = 0.007 |
| Valero, 2002 [12] | Beta-carotene: OR 0.82; 95% CI 0.51–1.33; P = 0.34 Alpha-carotene: OR 0.64; 95% CI 0.39–1.04; P = 0.07 Beta-cryptoxanthin: OR 0.97; 95% CI 0.61–1.56; P = 0.41 Lutein: OR 1.00; 95% CI 0.64–1.64; P = 0.78 Zeaxanthin: OR 0.99; 95% CI 0.61–1.60; P = 0.83 Lycopene: OR 1.11; 95% CI 0.69–1.78; P = 0.81 | Blood lycopene: OR 1.55; 95% CI 1.00–2.38 | Blood lycopene: OR 1.20; 95% CI 0.76–1.90 | Blood lycopene: OR 1.34; 95% CI 0.87–2.07 |
| Other | ||||
| Author | Any CAT | NUC | CX | PSC |
| Mares, 2010 [81] | - | High vs low HEI score: OR 0.63; 95% CI 0.43–0.91 | - | - |
| Rautiainen, 2014 [82] | Highest vs lowest TAC quintile: OR 0.87 95% CI 0.79–0.96; P = 0.03 | - | - | - |
| Author | Sample Size (n), Age (years) | Nutrients Examined | Key Findings |
|---|---|---|---|
| Multi-Vitamins | |||
| Mares-Perlman, 2000 [85] | n = 3089 | Supplementary Multivitamin, Vitamin C, Vitamin E | The 5-year risk for any CAT was 60% lower for multivitamins or any supplement use containing Vitamin C or E for more than 10 years. 10-year multivitamin use lowered the risk for NUC and CX but not for PSC (OR 0.6, 95% CI 0.4–0.9; OR 0.4, 95% CI 0.2–0.8; and OR, 0.9 95% CI 0.5–1.9; respectively). |
| Kuzniarz, 2001 [11] | n = 2873, 49–97 years | Supplementary Vitamin A, Thiamine, Riboflavin, Niacin, Pyridoxine, Folate, Vitamin B12 | Use of multivitamin supplements was associated with reduced prevalence of NUC, OR 0.6, 95% CI 0.4–1.0, P =0.05. For both NUC and CX, longer duration of multivitamin use was associated with reduced prevalence (NUC, trend P = 0.02; CX, trend P = 0.03). Use of thiamin supplements was associated with reduced prevalence of NUC (OR 0.6, 95% CI 0.4–1.0, P = 0.03, dose trend P = 0.03) and CX (OR 0.7, 95% CI 0.5–0.9, P = 0.01, dose trend P = 0.02). Riboflavin (OR 0.8, 95% CI 0.6–1.0, P = 0.05) and niacin (OR 0.7, 95% CI 0.6–1.0, P = 0.04) supplements exerted a weaker protective influence on CX. Vitamin A supplements were protective against NUC (OR 0.4, 95% CI 0.2–0.8, P = 0.01, dose trend P = 0.01). Folate (OR 0.4, 95% CI 0.2–0.9, P = 0.03) appeared protective for NUC, whereas both folate (NUC 0.6, 95% CI 0.3–0.9, P = 0.01, dose trend P = 0.04) and Vitamin B12 supplements (OR 0.7, 95% CI 0.5–1.0, P = 0.03, dose trend P = 0.02) were strongly protective against CX. |
| Age-Related Eye Disease Study Research Group, 2006 [86] | n = 4590, - | Supplementary CentrumTM Multivitamin (Vitamin A, E, C, B1, B2, B12, B6, D, Folic acid, Niacinamide, Biotin, Pantothenic acid, Calcium, Phosphorus, Iodine, Iron, Magnesium, Copper Zinc) | CentrumTM use is associated with a reduction in any lens opacity progression (OR 0.84, 95% CI 0.72–0.98, P = 0.025). Also protective for nuclear opacity events (OR 0.75, 95% CI 0.61–0.91, P = 0.004). |
| Zheng Selin, 2013 [83] | n = 31120, 45–79 years |
Supplemental Vitamin C, Vitamin E, Low dose multivitamins | The multivariable- adjusted HR for Vitamin C supplements only was 1.21 (95% CI 1.04–1.41) in compared to non-users. The HR for long-term Vitamin C users (≥10 years before baseline) was 1.36 (95% CI 1.02–1.81). The HR for Vitamin E use only was 1.59 (95% CI 1.12–2.26). Use of multivitamins only or multiple supplements in addition to Vitamin C or E was not associated with cataract risk. |
| Single Vitamins | |||
| Christen, 2004 [87] | n = 39876, ≥ 45 years | Supplementary Beta-carotene (50mg.d-1, alternate days) | 129 CAT in the beta-carotene group and 133 in the placebo group (RR 0.95, 95% CI 0.75–1.21). For cataract extraction, there were 94 cases in the beta-carotene group and 89 cases in the placebo group (RR 1.04, 95% CI 0.78–1.39). |
| Christen, 2008 [88] | n = 39876, ≥ 45 years | Supplementary Vitamin E (600 IU.d-1, alternate days) | No significant difference between the Vitamin E and placebo groups in the incidence of CAT (RR 0.96; 95% CI 0.88–1.04). No significant effects of Vitamin E on the incidence of NUC (RR 0.94; 95% CI 0.87–1.02), CX (RR 0.93; 95% CI 0.81–1.06), or PSC (RR 1.00; 95% CI 0.86–1.16). |
| Christen, 2015 [89] | n = 35533, ≥50 years | Supplementary Selenium (200 μg.d-1 from L-selenomethionine), Vitamin E (400 IU.d-1 of all rac-α-tocopheryl acetate) | 185 CAT in the selenium group and 204 in placebo (HR 0.91; 95 % CI 0.75–1.11; P = 0.37). For Vitamin E, there were 197 cases in the Vitamin E group and 192 in placebo (HR 1.02; 95 % CI 0.84–1.25; P = 0.81) |
| Christen,2010 [90] | n = 11545, ≥50 years | Supplementary Vitamin E (400 IU.d-1, alternate days) Vitamin C (500 mg.d-1 alternate days) | 579 CAT in the Vitamin E treated group and 595 in the Vitamin E placebo group (HR 0.99; 95% CI 0.88–1.11). For Vitamin C, there were 593 cataracts in the treated group and 581 in the placebo group (HR 1.02; 95% CI 0.91–1.14). |
| Christen,2002 [91] | n = 22071 | Supplementary Beta-carotene (50 mg.d-1, alternate days) | No difference between the beta-carotene and placebo groups in the overall incidence of CAT (998 cases vs 1017 cases; RR 1.00; 95% CI 0.91–1.09) or CAT extraction (584 vs 593; RR 1.00; 95% CI 0.89–1.12). |
| Ferringo, 2005 [92] | n = 1020 | Supplementary Vitamin A, Vitamin C, Vitamin E, Beta-carotene | High Vitamin C levels were associated with a protective effect on NUC (OR 0.54; 95% CI 0.30, 0.97) and PSC (OR: 0.37; 95% CI: 0.15–0.93). High Vitamin E levels were associated with increased prevalence of CX (OR 1.99; 95% CI 1.02–3.90), PSC (OR 3.27; 95% CI 1.34–7.96) and of any CAT (OR 1.86; 95% CI 1.08–3.18). |
| Rautiainen, 2009 [15] | n = 24593, 49–83 years | Supplementary Vitamin C | HR of Vitamin C supplement users compared with that for nonusers was 1.25 (95% CI 1.05, 1.50). The HR for the duration of 10 y of use before baseline was 1.46 (95% CI 0.93, 2.31). The HR for the use of multivitamins containing Vitamin C was 1.09 (95% CI 0.94, 1.25). Among women aged 65 y, Vitamin C supplement use increased the risk of CAT by 38% (95% CI 12%, 69%). |
| The REACT Group,2002 [84] | n = 297 | Supplementary Vitamin E (200 mg all-rac alpha-tocopherol acetate), Vitamin C (250 mg ascorbic acid), and b-carotene (6 mg) | After two years of treatment, there was a small positive treatment effect in U.S. patients (p = 0.0001); after three years a positive effect (p = 0.048) in both the U.S. and the U.K. groups. The positive effect in the U.S. group was even greater after three years: (IPO = 0.389 (Vitamin) vs. IPO = 2.517 (placebo); p = 0.0001). |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braakhuis, A.J.; Donaldson, C.I.; Lim, J.C.; Donaldson, P.J. Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies. Nutrients 2019, 11, 1186. https://doi.org/10.3390/nu11051186
Braakhuis AJ, Donaldson CI, Lim JC, Donaldson PJ. Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies. Nutrients. 2019; 11(5):1186. https://doi.org/10.3390/nu11051186
Chicago/Turabian StyleBraakhuis, Andrea J, Caitlin I Donaldson, Julie C Lim, and Paul J Donaldson. 2019. "Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies" Nutrients 11, no. 5: 1186. https://doi.org/10.3390/nu11051186
APA StyleBraakhuis, A. J., Donaldson, C. I., Lim, J. C., & Donaldson, P. J. (2019). Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies. Nutrients, 11(5), 1186. https://doi.org/10.3390/nu11051186