Calorie Restriction Modulates Reproductive Development and Energy Balance in Pre-Pubertal Male Rats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rats, Housing and Experimental Design
2.2. Metabolic Measurements
2.3. Blood Collection and Hormone Analyses
2.4. Testis Evaluations
2.5. RNA Isolation, cDNA Synthesis and Gene Expression Analyses by Real-time Polymerase Chain Reaction (RTPCR)
2.6. Statistical Analyses
3. Results
3.1. Feed Intake, Energy Expenditure and Respiratory Quotient
3.2. Body Weight and Composition
3.3. Plasma Hormone and Glucose Concentrations
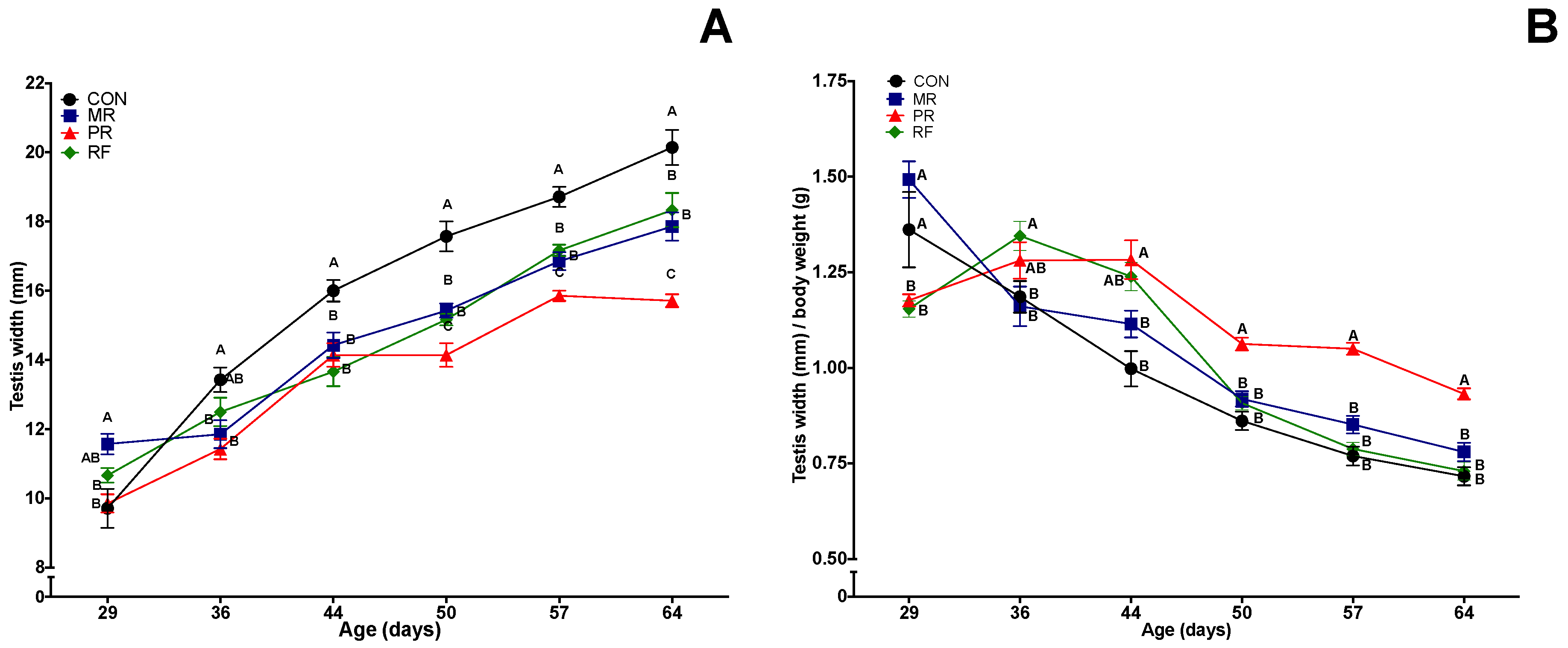
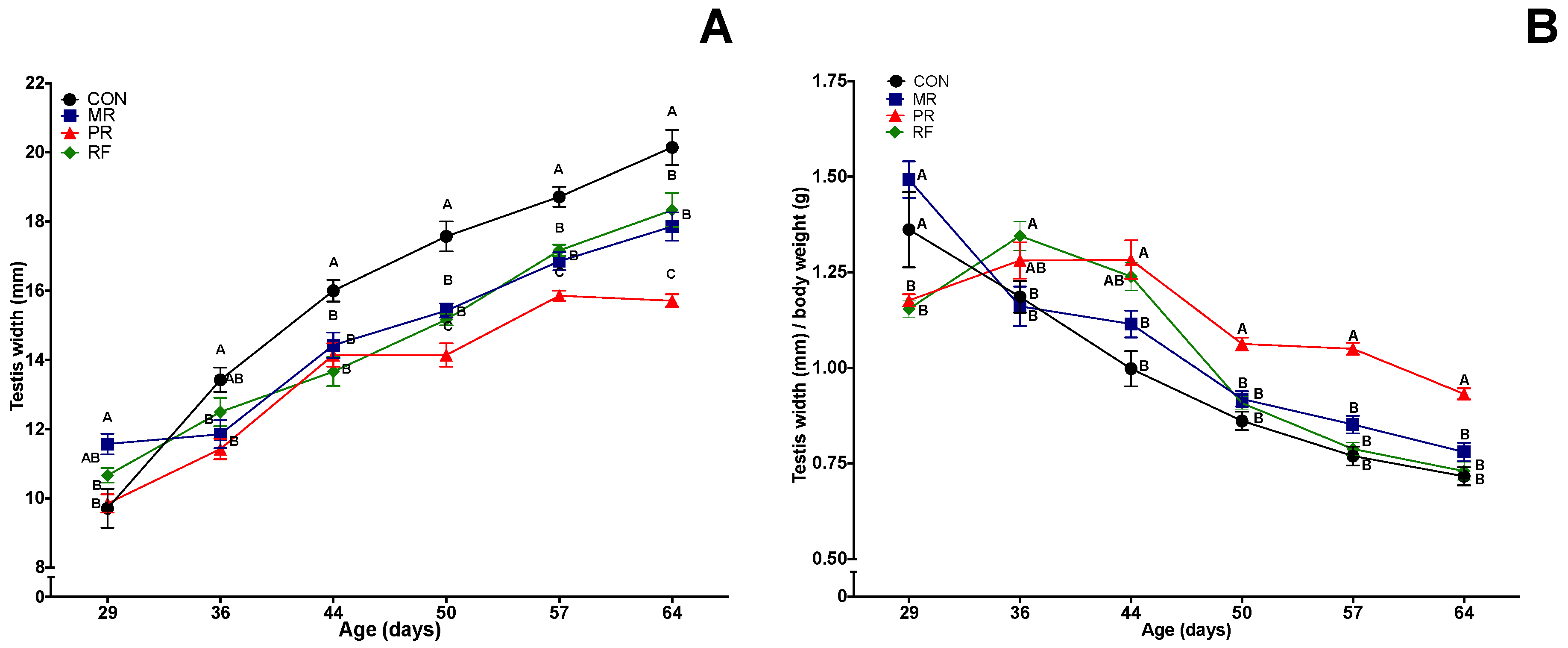
3.4. Testes Width and Weight
3.5. Sperm Motility, Morphology and Acrosome Integrity
3.6. mRNA Abundance of IGF-1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cantó, C.; Auwerx, J. Caloric restriction, SIRT1 and longevity. Trends Endocrinol. Metab. 2009, 20, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquero, A.; Reinberg, D. Calorie restriction and the exercise of chromatin. Genes Dev. 2009, 23, 1849–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCay, C.M.; Crowell, M.F.; Maynard, L.A. The effect of retarded growth upon the length of life span and upon the ultimate body size. J. Nutr. 1935, 10, 63–79. [Google Scholar] [CrossRef]
- Glass, A.R.; Herbert, D.C.; Anderson, J. Fertility onset, spermatogenesis, and pubertal development in male rats: Effect of graded underfeeding. Pediatr. Res. 1986, 20, 1161–1167. [Google Scholar] [CrossRef]
- Dance, A.; Kastelic, J.; Thundathil, J. A combination of insulin-like growth factor I (IGF-I) and FSH promotes proliferation of prepubertal bovine Sertoli cells isolated and cultured in vitro. Reprod. Fertil. Dev. 2017, 29, 1635–1641. [Google Scholar] [CrossRef]
- Brito, L.F.C.; Barth, A.D.; Rawlings, N.C.; Wilde, R.E.; Crews, D.H., Jr.; Mir, P.S.; Kastelic, J.P. Circulating metabolic hormones during the peripubertal period and their association with testicular development in bulls. Reprod. Domest. Anim. 2007, 42, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Dance, A.; Thundathil, J.; Wilde, R.; Blondin, P.; Kastelic, J. Enhanced early-life nutrition promotes hormone production and reproductive development in Holstein bulls. J. Dairy Sci. 2015, 98, 987–998. [Google Scholar] [CrossRef]
- Brecchia, G.; Bonanno, A.; Galeati, G.; Federici, C.; Maranesi, M.; Gobbetti, A.; Zerani, M.; Boiti, C. Hormonal and metabolic adaptation to fasting: Effects on the hypothalamic–pituitary–ovarian axis and reproductive performance of rabbit does. Domest. Anim. Endocrinol. 2006, 31, 105–122. [Google Scholar] [CrossRef]
- Compagnucci, C.V.; Compagnucci, G.E.; Lomniczi, A.; Mohn, C.; Vacas, I.; Cebral, E.; Elverdin, J.C.; Friedman, S.M.; Rettori, V.; Boyer, P.M. Effect of nutritional stress on the hypothalamo-pituitary-gonadal axis in the growing male rat. Neuroimmunomodulation 2002, 10, 153–162. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, R.; Martini, A.C.; Navarro, V.M.; Castellano, J.M.; Dieguez, C.; Aguilar, E.; Pinilla, L.; Tena-Sempere, M. Novel signals for the integration of energy balance and reproduction. Mol. Cell. Endocrinol. 2006, 254–255, 127–132. [Google Scholar] [CrossRef]
- Roa, J.; Tena-Sempere, M. Energy balance and puberty onset: Emerging role of central mTOR signaling. Trends Endocrinol. Metab. 2010, 21, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.F.; Sousa, M.; Silva, B.M.; Monteiro, M.P.; Alves, M.G. Obesity, energy balance and spermatogenesis. Reproduction 2017, 153, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.L.; Amatoury, M.; Steinbeck, K. Energy expenditure and intake during puberty in healthy nonobese adolescents: A systematic review. Am. J. Clin. Nutr. 2016, 104, 1061–1074. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.M. Endocrinology of growth and growth factors. Connect. Tissue Res. 1995, 31, 3–7. [Google Scholar] [CrossRef]
- Kelsey, M.M.; Zeitler, P.S. Insulin resistance of puberty. Curr. Diab. Rep. 2016, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gong, M. Review of the role of leptin in the regulation of male reproductive function. Andrologia 2018, 50, e12965. [Google Scholar] [CrossRef]
- Pezeshki, A.; Fahim, A.; Chelikani, P.K. Dietary whey and casein differentially affect energy balance, gut hormones, glucose metabolism, and taste preference in diet-induced obese rats. J. Nutr. 2015, 145, 2236–2244. [Google Scholar] [CrossRef]
- Vetter-O’Hagen, C.S.; Spear, L.P. The effects of gonadectomy on age and sex-typical patterns of ethanol consumption in sprague-dawley rats. Alcohol. Clin. Exp. Res. 2011, 35, 2039–2049. [Google Scholar] [CrossRef]
- Tu, L.; Yu, L.; Zhang, H. Morphology of rat testis preserved in three different fixatives. J. Huazhong Univ. Sci. Technol. Med. Sci. 2011, 31, 178–180. [Google Scholar] [CrossRef]
- Barth, A.; Oko, R. Abnormal Morphology of Bovine Spermatozoa; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Lybaert, P.; Danguy, A.; Leleux, F.; Meuris, S.; Lebrun, P. Improved methodology for the detection and quantification of the acrosome reaction in mouse spermatozoa. Histol. Histopathol. 2009, 24, 999–1007. [Google Scholar]
- Johnson, L.; May, M.R.; Busbee, D.L.; Williams, J.D. Effect of age and dietary restriction on daily sperm production and number and transit time of epididymal spermatozoa in the mouse. Age 1992, 15, 65–71. [Google Scholar] [CrossRef]
- Robb, G.W.; Amann, R.P.; Killian, G.J. Daily sperm production and epididymal sperm reserves of pubertal and adult rats. J. Reprod. Fertil. 1978, 54, 103–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndtson, W.E.; Neefus, C.; Foote, R.H.; Amann, R.P. Optimal replication for histometric analyses of testicular function in rats or rabbits. Toxicol. Sci. 1989, 12, 291–302. [Google Scholar] [CrossRef]
- Nausheen, S.; Shah, I.H.; Pezeshki, A.; Sigalet, D.L.; Chelikani, P.K. Effects of sleeve gastrectomy and ileal transposition, alone and in combination, on food intake, body weight, gut hormones, and glucose metabolism in rats. Am. J. Physiol. Endocrinol. Metab. 2013, 305, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Even, P.C.; Nadkarni, N.A. Indirect calorimetry in laboratory mice and rats: Principles, practical considerations, interpretation and perspectives. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R459–R476. [Google Scholar] [CrossRef]
- Dance, A.; Thundathil, J.; Blondin, P.; Kastelic, J. Enhanced early-life nutrition of Holstein bulls increases sperm production potential without decreasing postpubertal semen quality. Theriogenology 2016, 86, 687–694. [Google Scholar] [CrossRef]
- Brown, B.W. A review of nutritional influences on reproduction in boars, bulls and rams. Reprod. Nutr. Dev. 1994, 34, 89–114. [Google Scholar] [CrossRef] [Green Version]
- Eng, L.A.; Lee, J.H.; Sponenberg, D.P. Food restriction in adult male rats: Effects on mating, pregnancy, litter size and offspring growth rate. Nutr. Res. 1987, 7, 957–963. [Google Scholar] [CrossRef]
- Nelson, R.J. Photoperiod influences reproduction in the prairie vole (Microtus Ochrogaster). Biol. Reprod. 1985, 33, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, K.E.; Riggs, L.; Stetson, M.H. Food availability and photoperiod affect reproductive development and maintenance in the marsh rice rat (Oryzomys palustris). Physiol. Behav. 2003, 78, 41–49. [Google Scholar] [CrossRef]
- Chapin, R.E.; Gulati, D.K.; Barnes, L.H.; Teague, J.L. The effects of feed restriction on reproductive function in sprague-dawley rats. Toxicol. Sci. 1993, 20, 23–29. [Google Scholar] [CrossRef]
- Mouritsen, A.; Aksglaede, L.; Soerensen, K.; Hagen, C.P.; Petersen, J.H.; Main, K.M.; Juul, A. The pubertal transition in 179 healthy Danish children: Associations between pubarche, adrenarche, gonadarche, and body composition. Eur. J. Endocrinol. 2013, 168, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Loomba-Albrecht, L.A.; Styne, D.M. Effect of puberty on body composition. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 10–15. [Google Scholar] [CrossRef]
- Even, P.C.; Nicolaïdis, S. Adaptive changes in energy expenditure during mild and severe feed restriction in the rat. Br. J. Nutr. 1993, 70, 421–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostazir, M.; Jeffery, A.; Voss, L.; Wilkin, T. Childhood obesity: Evidence for distinct early and late environmental determinants a 12-year longitudinal cohort study. Int. J. Obes. 2015, 39, 1057. [Google Scholar] [CrossRef]
- Schneider, J.E. Energy balance and reproduction. Physiol. Behav. 2004, 81, 289–317. [Google Scholar] [CrossRef]
- Abreu, A.P.; Kaiser, U.B. Pubertal development and regulation. Lancet Diabetes Endocrinol. 2016, 4, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Barash, I.A.; Cheung, C.C.; Weigle, D.S.; Ren, H.O.; Kabigting, E.B.; Kuijper, J.L.; Clifton, D.K.; Steiner, R.A. Leptin is a metabolic signal to the reproductive system. Endocrinology 1996, 137, 3144–3147. [Google Scholar] [CrossRef]
- Schoeller, E.L.; Schon, S.; Moley, K.H. The effects of type 1 diabetes on the hypothalamic, pituitary and testes axis. Cell Tissue Res. 2012, 349, 839–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | Location on Template (bp) | GenBank Accession No. | Annealing Temperature (°C) | qPCR Efficiency (%) |
|---|---|---|---|---|---|
| IGF1 | Forward: TCTGCCTCTGTGACTTCTTGA | 321–341 | NM_001082478.1 | 61.4 | 98 |
| Reverse: AGCCTGTGGGCTTGTTGAAG | 515–496 | ||||
| IGF1R | Forward: TCGGCATCAAACTCCTCCTC | 1903–1922 | NM_052807 | 61.3 | 93.8 |
| Reverse: GATGGGTATTTTGTCTTTGGAGCA | 2058–2035 | ||||
| B-Actin | Forward: GGATCAGCAAGCAGGAGTACGA | 1148–1169 | NM_031144 | 60 | 113.3 |
| Reverse: AACGCAGCTCAGTAACAGTCCG | 1229–1208 |
| End Point | r | p Value |
|---|---|---|
| Testes weight | 0.70 | <0.0001 |
| Total sperm/testes | 0.78 | <0.0001 |
| Sperm/g testis | 0.537 | <0.01 |
| Daily sperm production | 0.537 | <0.01 |
| Tubule diameter | 0.364 | <0.07 |
| CON | MR | PR | RF | |
|---|---|---|---|---|
| Paired testis weight (g) | 3.25 ± 0.11 a | 3.02 ± 0.12 a | 2.52 ± 0.04 b | 2.76 ± 0.09 a |
| Testis (g) relative to body weight (100 g) | 1.14 ± 0.04 a | 1.32 ± 0.07 ab | 1.5 ± 0.07 b | 1.12 ± 0.16 a |
| Pubertal at Day 50 (testosterone >1 ng/mL) | 5/6 a | 4/6 ab | 1/6 b | 4/5 a |
| Total sperm/g testis (107) | 10.70 ± 0.20 a | 11.0 ± 0.30 a | 8.60 ± 0.20 b | 10.40 ± 0.30 a |
| Total sperm/testes (107) | 34.80 ± 1.10 a | 33.30 ± 2.0 a | 22.10 ± 5.20 b | 30.10 ± 1.0 a |
| Daily sperm production (106/g testis) | 17.60 ± 0.40 a | 18.0 ± 0.60 a | 14.10 ± 0.30 b | 17.0 ± 0.50 a |
| Seminiferous tubule diameter (µm) | 436.80 ± 1.60 | 432.10 ± 1.76 a | 409.50 ± 1.52 b | 433.10 ± 1.98 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzoto, G.; Sekhar, D.; Thundathil, J.C.; Chelikani, P.K.; Kastelic, J.P. Calorie Restriction Modulates Reproductive Development and Energy Balance in Pre-Pubertal Male Rats. Nutrients 2019, 11, 1993. https://doi.org/10.3390/nu11091993
Rizzoto G, Sekhar D, Thundathil JC, Chelikani PK, Kastelic JP. Calorie Restriction Modulates Reproductive Development and Energy Balance in Pre-Pubertal Male Rats. Nutrients. 2019; 11(9):1993. https://doi.org/10.3390/nu11091993
Chicago/Turabian StyleRizzoto, Guilherme, Deepa Sekhar, Jacob C. Thundathil, Prasanth K. Chelikani, and John P. Kastelic. 2019. "Calorie Restriction Modulates Reproductive Development and Energy Balance in Pre-Pubertal Male Rats" Nutrients 11, no. 9: 1993. https://doi.org/10.3390/nu11091993
APA StyleRizzoto, G., Sekhar, D., Thundathil, J. C., Chelikani, P. K., & Kastelic, J. P. (2019). Calorie Restriction Modulates Reproductive Development and Energy Balance in Pre-Pubertal Male Rats. Nutrients, 11(9), 1993. https://doi.org/10.3390/nu11091993