Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress

 , , , and
, , , and
Abstract
1. Introduction
2. GLP-1 Receptor and Signaling Pathways
2.1. Molecular Effects of GLP1 in the Hypothalamic Area
2.1.1. Voltage-Gated Na+ Channels
2.1.2. Voltage-Gated Ca2+ Channels and Nonselective Cationic Current
2.1.3. K+ Channels
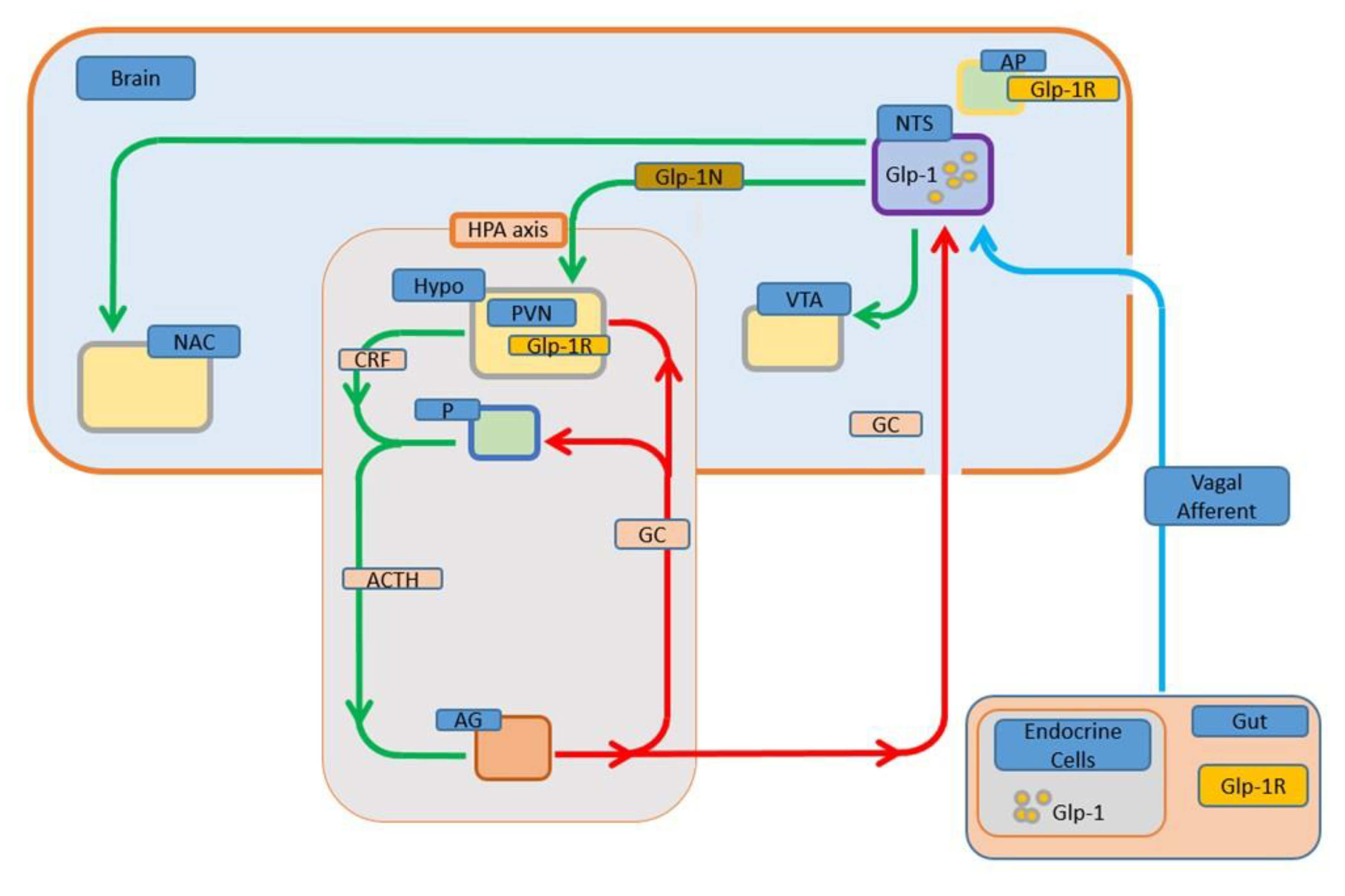
3. GLP-1 and the Stress Responses
3.1. GLP-1 Activates Hypothalamic-Pituitary-Adrenal Axis
3.2. GLP-1 Activates the Sympathetic Nervous System (SNS)
3.3. GLP-1 Mediates Multiple Responses to Stressors
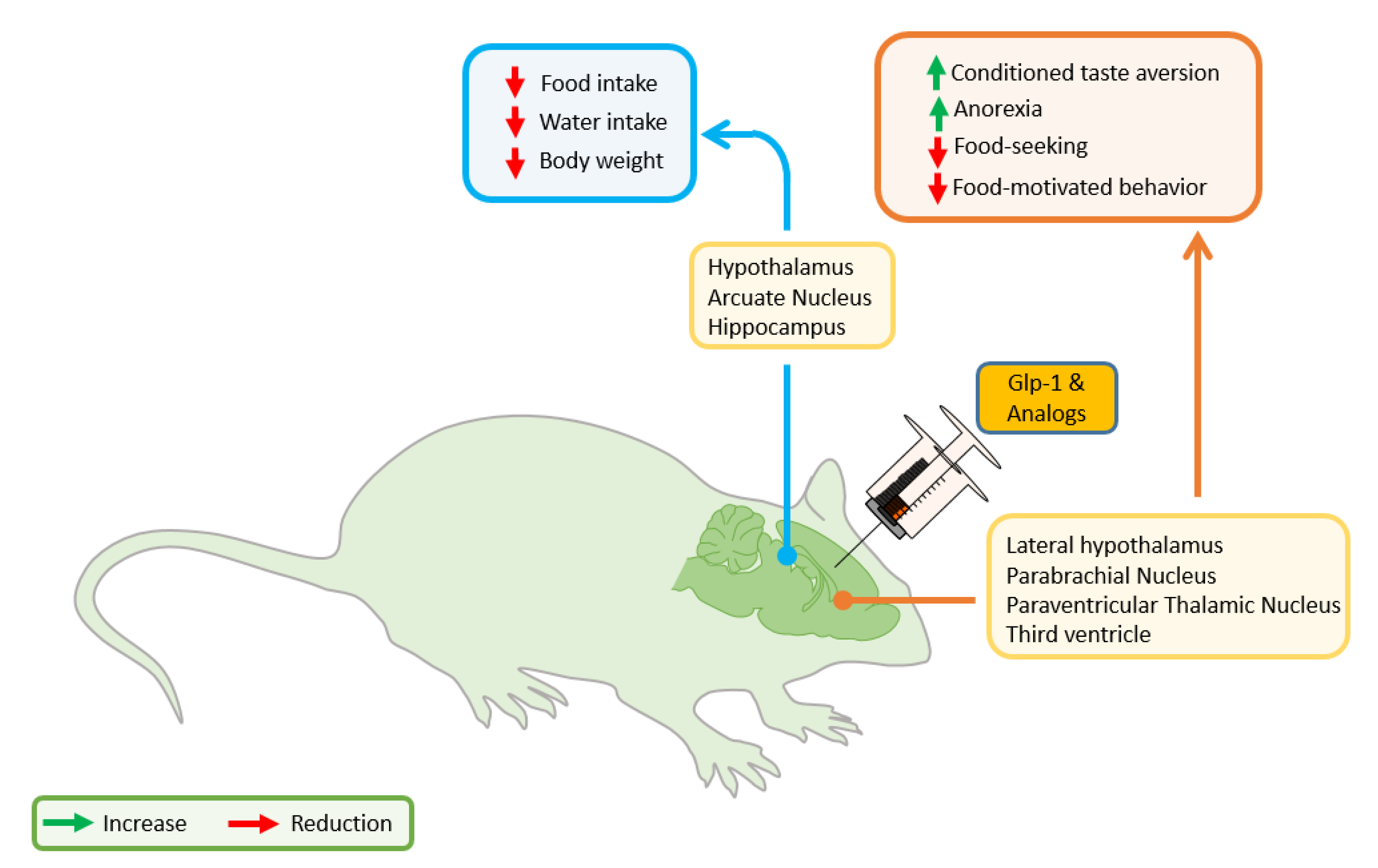
4. GLP-1 in the Control of Food Intake. Crosstalk with the Stress System
4.1. GLP-1 Anorectic Action and HPA Activation
4.2. GLP-1 Anorectic Effect: Hypothalamic Actions
4.3. GLP-1 Anorectic Effect: Forebrain/Hindbrain/Mesolimbic Actions
4.4. Stress in Obesity: GLP-1 and the Motivation to Feed
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sinha, R.; Jastreboff, A.M. Stress as a common risk factor for obesity and addiction. Biol. Psychiatry 2013, 73, 827–835. [Google Scholar]
- Dallman, M.F. Stress-induced obesity and the emotional nervous system. Trends Endocrinol. Metab. 2010, 21, 159–165. [Google Scholar]
- Lenoir, M.; Serre, F.; Cantin, L.; Ahmed, S.H. Intense sweetness surpasses cocaine reward. PLoS ONE 2007, 2, e698. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.J.; Baler, R.D. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. 2011, 15, 37–46. [Google Scholar]
- Kenny, P.J. Reward Mechanisms in obesity: New insights and future directions. Neuron 2011, 69, 664–679. [Google Scholar] [PubMed]
- Lutter, M.; Nestler, E.J. Homeostatic and hedonic signals interact in the regulation of food intake. J. Nutr. 2009, 139, 629–632. [Google Scholar] [PubMed]
- Vainik, U.; García-García, I.; Dagher, A. Uncontrolled eating: A unifying heritable trait linked with obesity, overeating, personality and the brain. Eur. J. Neurosci. 2019, 50, 2430–2445. [Google Scholar] [PubMed]
- Sweeney, P.; Yang, Y. Neural circuit mechanisms underlying emotional regulation of homeostatic feeding. Trends Endocrinol. Metab. 2017, 28, 437–448. [Google Scholar] [PubMed]
- Wise, R.A. Role of brain dopamine in food reward and reinforcement. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1149–1158. [Google Scholar]
- Kelley, A.E.; Baldo, B.A.; Pratt, W.E. A proposed hypothalamic-thalamic-striatal axis for the integration of energy balance, arousal, and food reward. J. Comp. Neurol. 2005, 493, 72–85. [Google Scholar] [PubMed]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [PubMed]
- Baggio, L.L.; Drucker, D.J. Glucagon-like peptide-1 receptors in the brain: Controlling food intake and body weight. J. Clin. Investig. 2014, 124, 4223–4226. [Google Scholar] [PubMed]
- Holt, M.K.; Richards, J.E.; Cook, D.R.; Brierley, D.I.; Williams, D.L.; Reimann, F.; Gribble, F.M.; Trapp, S. Preproglucagon neurons in the nucleus of the solitary tract are the main source of brain GLP-1, mediate stress-induced hypophagia, and limit unusually large intakes of food. Diabetes 2019, 68, 21–33. [Google Scholar] [PubMed]
- Drucker, D.J. Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar]
- Eissele, R.; Göke, R.; Willemer, S.; Harthus, H.-P.; Vermeer, H.; Arnold, R.; Göke, B. Glucagon-like peptide-1 cells in the gastrointestinal tract and pancreas of rat, pig and man. Eur. J. Clin. Investig. 1992, 22, 283–291. [Google Scholar]
- Larsen, P.J.; Holst, J.J. Glucagon-related peptide 1 (GLP-1): Hormone and neurotransmitter. Regul. Pept. 2005, 128, 97–107. [Google Scholar]
- Kuhre, R.E.; Holst, J.J.; Kappe, C. The regulation of function, growth and survival of GLP-1-producing L-cells. Clin. Sci. 2015, 130, 79–91. [Google Scholar]
- Mojsov, S.; Heinrich, G.; Wilson, I.B.; Ravazzola, M.; Orci, L.; Habener, J.F. Preproglucagon gene expression in pancreas and intestine diversifies at the level of post-translational processing. J. Biol. Chem. 1986, 261, 11880–11889. [Google Scholar]
- Ugleholdt, R.; Zhu, X.; Deacon, C.F.; Ørskov, C.; Steiner, D.F.; Holst, J.J. Impaired intestinal proglucagon processing in mice lacking prohormone convertase 1. Endocrinology 2004, 145, 1349–1355. [Google Scholar]
- Rouillé, Y.; Martin, S.; Steiner, D.F. Differential processing of proglucagon by the subtilisin-like prohormone convertases PC2 and PC3 to generate either glucagon or glucagon-like peptide. J. Biol. Chem. 1995, 270, 26488–26496. [Google Scholar]
- Larsen, P.J.; Tang-Christensen, M.; Holst, J.J.; Ørskov, C. Distribution of glucagon-like peptide-1 and other preproglucagon-derived peptides in the rat hypothalamus and brainstem. Neuroscience 1997, 77, 257–270. [Google Scholar]
- Llewellyn-Smith, I.J.; Reimann, F.; Gribble, F.M.; Trapp, S. Preproglucagon neurons project widely to autonomic control areas in the mouse brain. Neuroscience 2011, 180, 111–121. [Google Scholar]
- Merchenthaler, I.; Lane, M.; Shughrue, P. Distribution of pre-pro-glucagon and glucagon-like peptide-1 receptor messenger RNAs in the rat central nervous system. J. Comp. Neurol. 1999, 403, 261–280. [Google Scholar] [PubMed]
- Hisadome, K.; Reimann, F.; Gribble, F.M.; Trapp, S. Leptin directly depolarizes preproglucagon neurons in the nucleus tractus dolitarius electrical properties of glucagon-like peptide 1 neurons. Diabetes 2010, 59, 1890–1898. [Google Scholar] [PubMed]
- Vrang, N.; Grove, K. The brainstem preproglucagon system in a non-human primate (Macaca mulatta). Brain Res. 2011, 1397, 28–37. [Google Scholar] [PubMed]
- Zheng, H.; Cai, L.; Rinaman, L. Distribution of glucagon-like peptide 1-immunopositive neurons in human caudal medulla. Brain Struct. Funct. 2015, 220, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Thiebaud, N.; Llewellyn-Smith, I.J.; Gribble, F.; Reimann, F.; Trapp, S.; Fadool, D.A. The incretin hormone glucagon-like peptide 1 increases mitral cell excitability by decreasing conductance of a voltage-dependent potassium channel. J. Physiol. 2016, 594, 2607–2628. [Google Scholar] [PubMed]
- Thiebaud, N.; Gribble, F.; Reimann, F.; Trapp, S.; Fadool, D.A. A unique olfactory bulb microcircuit driven by neurons expressing the precursor to glucagon-like peptide 1. Sci. Rep. 2019, 9, 1–16. [Google Scholar]
- Llewellyn-Smith, I.J.; Gnanamanickam, G.J.E.; Reimann, F.; Gribble, F.M.; Trapp, S. Preproglucagon (PPG) neurons innervate neurochemicallyidentified autonomic neurons in the mouse brainstem. Neuroscience 2013, 229, 130–143. [Google Scholar]
- Vrang, N.; Hansen, M.; Larsen, P.J.; Tang-Christensen, M. Characterization of brainstem preproglucagon projections to the paraventricular and dorsomedial hypothalamic nuclei. Brain Res. 2007, 1149, 118–126. [Google Scholar]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 neurons in the nucleus of the solitary tract project directly to the ventral tegmental area and nucleus accumbens to control for food intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [PubMed]
- Vrang, N.; Larsen, P.J. Preproglucagon derived peptides GLP-1, GLP-2 and oxyntomodulin in the CNS: Role of peripherally secreted and centrally produced peptides. Prog. Neurobiol. 2010, 92, 442–462. [Google Scholar] [PubMed]
- Williams, D.L.; Lilly, N.A.; Edwards, I.J.; Yao, P.; Richards, J.E.; Trapp, S. GLP-1 action in the mouse bed nucleus of the stria terminalis. Neuropharmacology 2018, 131, 83–95. [Google Scholar] [PubMed]
- Zheng, H.; Stornetta, R.L.; Agassandian, K.; Rinaman, L. Glutamatergic phenotype of glucagon-like peptide 1 neurons in the caudal nucleus of the solitary tract in rats. Brain Struct. Funct. 2015, 220, 3011–3022. [Google Scholar] [PubMed]
- Alhadeff, A.L.; Baird, J.-P.; Swick, J.C.; Hayes, M.R.; Grill, H.J. Glucagon-like peptide-1 receptor signaling in the lateral parabrachial nucleus contributes to the control of food intake and motivation to feed. Neuropsychopharmacology 2014, 39, 2233–2243. [Google Scholar] [PubMed]
- Ong, Z.Y.; Liu, J.J.; Pang, Z.P.; Grill, H.J. Paraventricular thalamic control of food intake and reward: Role of glucagon-like peptide-1 receptor signaling. Neuropsychopharmacology 2017, 42, 2387–2397. [Google Scholar] [PubMed]
- López-Ferreras, L.; Eerola, K.; Shevchouk, O.T.; Richard, J.E.; Nilsson, F.H.; Jansson, L.E.; Hayes, M.R.; Skibicka, K.P. The supramammillary nucleus controls anxiety-like behavior; key role of GLP-1R. Psychoneuroendocrinology 2020, 119, 104720. [Google Scholar]
- Ulrich-Lai, Y.M.; Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn-Smith, I.J.; Marina, N.; Manton, R.N.; Reimann, F.; Gribble, F.M.; Trapp, S. Spinally projecting preproglucagon axons preferentially innervate sympathetic preganglionic neurons. Neuroscience 2015, 284, 872–887. [Google Scholar]
- Alvarez, E.; Martinez, M.D.; Roncero, I.; Chowen, J.A.; Garcia-Cuartero, B.; Gispert, J.D.; Sanz, C.; Vazquez, P.; Maldonado, A.; de Caceres, J.; et al. The expression of GLP-1 receptor mRNA and protein allows the effect of GLP-1 on glucose metabolism in the human hypothalamus and brainstem. J. Neurochem. 2005, 92, 798–806. [Google Scholar]
- Chowen, J.A.; de Fonseca, F.R.; Alvarez, E.; Navarro, M.; García-Segura, L.M.; Blázquez, E. Increased glucagon-like peptide-1 receptor expression in glia after mechanical lesion of the rat brain. Neuropeptides 1999, 33, 212–215. [Google Scholar] [PubMed]
- Reiner, D.J.; Mietlicki-Baase, E.G.; McGrath, L.E.; Zimmer, D.J.; Bence, K.K.; Sousa, G.L.; Konanur, V.R.; Krawczyk, J.; Burk, D.H.; Kanoski, S.E.; et al. Astrocytes regulate GLP-1 receptor-mediated effects on energy balance. J. Neurosci. 2016, 36, 3531–3540. [Google Scholar]
- Fandiño, J.; Vaz, A.A.; Toba, L.; Romaní-Pérez, M.; González-Matías, L.; Mallo, F.; Diz-Chaves, Y. Liraglutide enhances the activity of the ACE-2/Ang(1–7)/Mas receptor pathway in lungs of male pups from food-restricted mothers and prevents the reduction of SP-A. Int. J. Endocrinol. 2018, 2018, 6920620. [Google Scholar]
- Diz-Chaves, Y.; Toba, L.; Fandiño, J.; González-Matías, L.C.; Garcia-Segura, L.M.; Mallo, F. The GLP-1 analog, liraglutide prevents the increase of proinflammatory mediators in the hippocampus of male rat pups submitted to maternal perinatal food restriction. J. Neuroinflamm. 2018, 15, 337. [Google Scholar] [PubMed]
- Romaní-Pérez, M.; Outeiriño-Iglesias, V.; Gil-Lozano, M.; González-Matías, L.C.; Mallo, F.; Vigo, E. Pulmonary GLP-1 receptor increases at birth and exogenous GLP-1 receptor agonists augmented surfactant-protein levels in litters from normal and nitrofen-treated pregnant rats. Endocrinology 2013, 154, 1144–1155. [Google Scholar] [CrossRef]
- Outeiriño-Iglesias, V.; Romaní-Pérez, M.; González-Matías, L.C.; Vigo, E.; Mallo, F. GLP-1 increases preovulatory LH source and the number of mature follicles, as well as synchronizing the onset of puberty in female rats. Endocrinology 2015, 156, 4226–4237. [Google Scholar] [CrossRef]
- Skibicka, K.P. The central GLP-1: Implications for food and drug reward. Front. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Ghosal, S.; Myers, B.; Herman, J.P. Role of central glucagon-like peptide-1 in stress regulation. Physiol. Behav. 2013, 122, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Holt, M.K.; Trapp, S. The physiological role of the brain GLP-1 system in stress. Cogent Biol. 2016, 2, 1229086. [Google Scholar]
- Terrill, S.J.; Maske, C.B.; Williams, D.L. Endogenous GLP-1 in lateral septum contributes to stress-induced hypophagia. Physiol. Behav. 2018, 192, 17–22. [Google Scholar]
- Terrill, S.J.; Holt, M.K.; Maske, C.B.; Abrams, N.; Reimann, F.; Trapp, S.; Williams, D.L. Endogenous GLP-1 in lateral septum promotes satiety and suppresses motivation for food in mice. Physiol. Behav. 2019, 206, 191–199. [Google Scholar] [CrossRef]
- Zheng, H.; Reiner, D.J.; Hayes, M.R.; Rinaman, L. Chronic suppression of glucagon-like peptide-1 receptor (GLP1R) mRNA translation in the rat bed nucleus of the stria terminalis reduces anxiety-like behavior and stress-induced hypophagia, but prolongs stress-induced elevation of plasma corticosterone. J. Neurosci. 2019, 39, 2649–2663. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. From the incretin concept and the discovery of GLP-1 to today’s diabetes therapy. Front. Endocrinol. 2019, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Madsbad, S. Review of head-to-head comparisons of glucagon-like peptide-1 receptor agonists. Diabetes Obes. Metab. 2016, 18, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Hoare, S.R.J. Mechanisms of peptide and nonpeptide ligand binding to Class B G-protein-coupled receptors. Drug Discov. Today 2005, 10, 417–427. [Google Scholar] [CrossRef]
- Thorens, B. Expression cloning of the pancreatic beta cell receptor for the gluco-incretin hormone glucagon-like peptide 1. Proc. Natl. Acad. Sci. USA 1992, 89, 8641–8645. [Google Scholar] [CrossRef]
- Bullock, B.P.; Heller, R.S.; Habener, J.F. Tissue distribution of messenger ribonucleic acid encoding the rat glucagon-like peptide-1 receptor. Endocrinology 1996, 137, 2968–2978. [Google Scholar] [CrossRef]
- Romaní-Pérez, M.; Outeiriño-Iglesias, V.; Moya, C.M.; Santisteban, P.; González-Matías, L.C.; Vigo, E.; Mallo, F. Activation of the GLP-1 receptor by liraglutide increases ACE2 expression, reversing right ventricle hypertrophy, and improving the production of SP-A and SP-B in the lungs of type 1 diabetes rats. Endocrinology 2015, 156, 3559–3569. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, H.; Ma, X.; Zhang, Y.; Lu, S.; Wang, Y.; Zong, C.; Qin, D.; Wang, Y.; Yingfeng Yang, Y.; et al. GLP-1/GLP-1R Signaling in Regulation of Adipocyte Differentiation and Lipogenesis. Cell. Physiol. Biochem. 2017, 42, 1165–1176. [Google Scholar] [CrossRef]
- Iacobellis, G.; Camarena, V.; Sant, D.; Wang, G. Human epicardial fat expresses glucagon-like peptide 1 and 2 receptors genes. Horm. Metab. Res. 2017, 49, 625–630. [Google Scholar] [CrossRef]
- Ejarque, M.; Guerrero-Pérez, F.; de la Morena, N.; Casajoana, A.; Virgili, N.; López-Urdiales, R.; Maymó-Masip, E.; Pujol Gebelli, J.; Garcia Ruiz de Gordejuela, A.; Perez-Maraver, M.; et al. Role of adipose tissue GLP-1R expression in metabolic improvement after bariatric surgery in patients with type 2 diabetes. Sci. Rep. 2019, 9, 6274. [Google Scholar] [CrossRef] [PubMed]
- Vendrell, J.; El Bekay, R.; Peral, B.; García-Fuentes, E.; Megia, A.; Macias-Gonzalez, M.; Real, J.F.; Jimenez-Gomez, Y.; Escoté, X.; Pachón, G.; et al. Study of the potential association of adipose tissue GLP-1 receptor with obesity and insulin resistance. Endocrinology 2011, 152, 4072–4079. [Google Scholar] [CrossRef]
- Segerstolpe, Å.; Palasantza, A.; Eliasson, P.; Andersson, E.M.; Andréasson, A.C.; Sun, X.; Picelli, S.; Sabirsh, A.; Clausen, M.; Bjursell, M.K.; et al. Single-cell transcriptome profiling of human pancreatic islets in health and type 2 diabetes. Cell Metab. 2016, 24, 593–607. [Google Scholar] [CrossRef]
- Zhang, Y.; Parajuli, K.R.; Fava, G.E.; Gupta, R.; Xu, W.; Nguyen, L.U.; Zakaria, A.F.; Fonseca, V.A.; Wang, H.; Mauvais-Jarvis, F.; et al. GLP-1 receptor in pancreatic A-cells regulates glucagon secretion in a glucose-dependent bidirectional manner. Diabetes 2019, 68, 34–44. [Google Scholar] [CrossRef]
- Ast, J.; Arvaniti, A.; Fine, N.H.F.; Nasteska, D.; Ashford, F.B.; Stamataki, Z.; Koszegi, Z.; Bacon, A.; Jones, B.J.; Lucey, M.A.; et al. Super-resolution microscopy compatible fluorescent probes reveal endogenous glucagon-like peptide-1 receptor distribution and dynamics. Nat. Commun. 2020, 11, 467. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.; Parker, H.E.; Adriaenssens, A.E.; Hodgson, J.M.; Cork, S.C.; Trapp, S.; Gribble, F.M.; Reimann, F. Identification and characterization of GLP-1 receptor-expressing cells using a new transgenic mouse model. Diabetes 2014, 63, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, H.; Wiesenthal, S.R.; MacDonald, P.E.; McCall, R.H.; Tchipashvili, V.; Rashid, S.; Satkunarajah, M.; Irwin, D.M.; Shi, Z.Q.; Brubaker, P.L.; et al. Glucagon-like peptide 1 increases insulin sensitivity in depancreatized dogs. Diabetes 1999, 48, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.V.; Lee, Y.C.; Drucker, D.J. Divergent tissue-specific and developmental expression of receptors for glucagon and glucagon-like peptide-1 in the mouse. Endocrinology 1994, 134, 2156–2164. [Google Scholar] [CrossRef]
- Uttenthal, L.O.; Toledano, A.; Blázquez, E. Autoradiographic localization of receptors for glucagon-like peptide-1(7-36) amide in rat brain. Neuropeptides 1992, 21, 143–146. [Google Scholar] [CrossRef]
- Shimizu, I.; Hirota, M.; Ohboshi, C.; Shima, K. Identification and localization of glucagon-like peptide-1 and its receptor in rat Brain. Endocrinology 1987, 121, 1076–1082. [Google Scholar] [CrossRef]
- Cork, S.C.; Richards, J.E.; Holt, M.K.; Gribble, F.M.; Reimann, F.; Trapp, S. Distribution and characterisation of Glucagon-like peptide-1 receptor expressing cells in the mouse brain. Mol. Metab. 2015, 4, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Göke, R.; Larsen, P.J.; Mikkelsen, J.D.; Sheikh, S.P. Identification of specific binding sites for glucagon-like peptide-1 on the posterior lobe of the rat pituitary. Neuroendocrinology 1995, 62, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.L.; Durai, H.H.; Trammell, T.S.; Noble, B.L.; Mortlock, D.P.; Galli, A.; Stanwood, G.D. A novel mouse model of glucagon-like peptide-1 receptor expression: A look at the brain. J. Comp. Neurol. 2020, 528, 2445–2470. [Google Scholar] [CrossRef]
- López-Ferreras, L.; Eerola, K.; Mishra, D.; Shevchouk, O.T.; Richard, J.E.; Nilsson, F.H.; Hayes, M.R.; Skibicka, K.P. GLP-1 modulates the supramammillary nucleus-lateral hypothalamic neurocircuit to control ingestive and motivated behavior in a sex divergent manner. Mol. Metab. 2019, 20, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Heppner, K.M.; Kirigiti, M.; Secher, A.; Paulsen, S.J.; Buckingham, R.; Pyke, C.; Knudsen, L.B.; Vrang, N.; Grove, K.L. Expression and distribution of glucagon-like peptide-1 receptor mRNA, protein and binding in the male nonhuman primate (Macaca mulatta) brain. Endocrinology 2015, 156, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Hölscher, C. Receptors for the incretin glucagon-like peptide-1 are expressed on neurons in the central nervous system. Neuroreport 2009, 20, 1161–1166. [Google Scholar] [CrossRef]
- Yoon, G.; Kim, Y.K.; Song, J. Glucagon-like peptide-1 suppresses neuroinflammation and improves neural structure. Pharmacol. Res. 2020, 152, 104615. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.P.; Kam, T.I.; Panicker, N.; Kim, S.; Oh, Y.; Park, J.S.; Kwon, S.H.; Park, Y.J.; Karuppagounder, S.S.; Park, H.; et al. Block of A1 astrocyte conversion by microglia is neuroprotective in models of Parkinson’s disease. Nat. Med. 2018, 24, 931–938. [Google Scholar] [CrossRef]
- Timper, K.; del Río-Martín, A.; Cremer, A.L.; Bremser, S.; Alber, J.; Giavalisco, P.; Varela, L.; Heilinger, C.; Nolte, H.; Trifunovic, A.; et al. GLP-1 receptor signaling in astrocytes regulates fatty acid oxidation, mitochondrial integrity, and function. Cell Metab. 2020, 31, 1189–1205. [Google Scholar] [CrossRef]
- Wu, H.-Y.; Tang, X.-Q.; Mao, X.-F.; Wang, Y.-X. Autocrine interleukin-10 mediates glucagon-like peptide-1 receptor-induced spinal microglial β-endorphin expression. J. Neurosci. 2017, 37, 11701–11714. [Google Scholar] [CrossRef]
- Pyke, C.; Knudsen, L.B. The Glucagon-like peptide-1 receptor—Or not? Endocrinology 2013, 154, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Kakei, M.; Yada, T.; Nakagawa, A.; Nakabayashi, H. Glucagon-like peptide-1 evokes action potentials and increases cytosolic Ca2+in rat nodose ganglion neurons. Auton. Neurosci. Basic Clin. 2002, 102, 39–44. [Google Scholar] [CrossRef]
- Perry, T.; Haughey, N.J.; Mattson, M.P.; Egan, J.M.; Greig, N.H. Protection and reversal of excitotoxic neuronal damage by glucagon-like peptide-1 and exendin-4. J. Pharmacol. Exp. Ther. 2002, 302, 881–888. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, P.E.; Wang, X.; Xia, F.; El-kholy, W.; Targonsky, E.D.; Tsushima, R.G.; Wheeler, M.B. Antagonism of rat β-cell voltage-dependent K + currents by exendin 4 requires dual activation of the cAMP/Protein kinase A and phosphatidylinositol 3-kinase signaling pathways. J. Biol. Chem. 2003, 278, 52446–52453. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Philippe, J.; Mojsov, S.; Chick, W.L.; Habener, J.F. Glucagon-like peptide I stimulates insulin gene expression and increases cyclic AMP levels in a rat islet cell line. Proc. Natl. Acad. Sci. USA 1987, 84, 3434–3438. [Google Scholar] [CrossRef]
- Wang, C.; Mao, R.; Van De Casteele, M.; Pipeleers, D.; Ling, Z. Glucagon-like peptide-1 stimulates GABA formation by pancreatic β-cells at the level of glutamate decarboxylase. Am. J. Physiol. Metab. 2007, 292, E1201–E1206. [Google Scholar] [CrossRef] [PubMed]
- Light, P.E.; Manning Fox, J.E.; Riedel, M.J.; Wheeler, M.B. Glucagon-like peptide-1 inhibits pancreatic ATP-sensitive potassium channels via a protein kinase A- and ADP-dependent mechanism. Mol. Endocrinol. 2002, 16, 2135–2144. [Google Scholar] [CrossRef]
- Psichas, A.; Glass, L.L.; Sharp, S.J.; Reimann, F.; Gribble, F.M. Galanin inhibits GLP-1 and GIP secretion via the GAL 1 receptor in enteroendocrine L and K cells. Br. J. Pharmacol. 2016, 173, 888–898. [Google Scholar] [CrossRef]
- Gromada, J.; Anker, C.; Bokvist, K.; Knudsen, L.B.; Wahl, P. Glucagon-like peptide-1 receptor expression in Xenopus oocytes stimulates inositol trisphosphate-dependent intracellular Ca2+ mobilization. FEBS Lett. 1998, 425, 277–280. [Google Scholar] [CrossRef]
- Shigeto, M.; Cha, C.Y.; Rorsman, P.; Kaku, K. A role of PLC/PKC-dependent pathway in GLP-1-stimulated insulin secretion. J. Mol. Med. 2017, 95, 361–368. [Google Scholar] [CrossRef]
- Bos, J.L. Epac proteins: Multi-purpose cAMP targets. Trends Biochem. Sci. 2006, 31, 680–686. [Google Scholar] [CrossRef]
- Kang, G.; Joseph, J.W.; Chepurny, O.G.; Monaco, M.; Wheeler, M.B.; Bos, J.L.; Schwede, F.; Genieser, H.-G.; Holz, G.G. Epac-selective cAMP analog 8-pCPT-2′- O -Me-cAMP as a stimulus for Ca2+ -induced Ca2+ release and exocytosis in pancreatic β-Cells. J. Biol. Chem. 2003, 278, 8279–8285. [Google Scholar] [CrossRef] [PubMed]
- Jacobo, S.M.P.; Guerra, M.L.; Hockerman, G.H. Cav1.2 and Cav1.3 are differentially coupled to glucagon-like peptide-1 potentiation of glucose-stimulated insulin secretion in the pancreatic β-Cell line INS-1. J. Pharmacol. Exp. Ther. 2009, 331, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Hällbrink, M.; Holmqvist, T.; Olsson, M.; Östenson, C.G.; Efendic, S.; Langel, Ü. Different domains in the third intracellular loop of the GLP-1 receptor are responsible for Gαs and Gαi/Gαo activation. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2001, 1546, 79–86. [Google Scholar] [CrossRef]
- Gromada, J.; Ding, W.G.; Barg, S.; Renström, E.; Rorsman, P. Multisite regulation of insulin secretion by cAMP-increasing agonists: Evidence that glucagon-like peptide 1 and glucagon act via distinct receptors. Pflug. Arch. Eur. J. Physiol. 1997, 434, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Acuna-Goycolea, C.; van den Pol, A. Glucagon-like peptide 1 excites hypocretin/orexin neurons by direct and indirect mechanisms: Implications for viscera-mediated arousal. J. Neurosci. 2004, 24, 8141–8152. [Google Scholar] [CrossRef] [PubMed]
- Farkas, I.; Vastagh, C.; Farkas, E.; Bálint, F.; Skrapits, K.; Hrabovszky, E.; Fekete, C.; Liposits, Z. Glucagon-like peptide-1 excites firing and increases GABAergic miniature postsynaptic currents (mPSCs) in gonadotropin-releasing hormone (GnRH) neurons of the male mice via activation of nitric oxide (NO) and suppression of endocannabinoid signaling pathways. Front. Cell. Neurosci. 2016, 10, 214. [Google Scholar]
- Holz, G.G., IV; Leech, C.A.; Habener, J.F. Activation of a cAMP-regulated Ca2+-signaling pathway in pancreatic β-cells by the insulinotropic hormone glucagon-like peptide-1. J. Biol. Chem. 1995, 270, 17749–17757. [Google Scholar] [CrossRef]
- McIntosh, C.H.S.; Widenmaier, S.; Kim, S.J. Pleiotropic actions of the incretin hormones. Vitam. Horm. 2010, 84, 21–79. [Google Scholar]
- Shigeto, M.; Ramracheya, R.; Tarasov, A.I.; Cha, C.Y.; Chibalina, M.V.; Hastoy, B.; Philippaert, K.; Reinbothe, T.; Rorsman, N.; Salehi, A.; et al. GLP-1 stimulates insulin secretion by PKC-dependent TRPM4 and TRPM5 activation. J. Clin. Investig. 2015, 125, 4714–4728. [Google Scholar] [CrossRef]
- Gromada, J.; Holst, J.J.; Rorsman, P. Cellular regulation of islet hormone secretion by the incretin hormone glucagon-like peptide 1. Pflügers Arch. Eur. J. Physiol. 1998, 435, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Leech, C.A.; Holz, G.G.; Habener, J.F. Signal transduction of PACAP and GLP-1 in pancreatic beta cells. Ann. N. Y. Acad. Sci. 1996, 805, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Gromada, J.; Dissing, S.; Bokvist, K.; Renström, E.; Frøkjser-Jensen, J.; Wulff, B.S.; Rorsman, P. Glucagon-like peptide I increases cytoplasmic calcium in insulin-secreting βTC3-cells by enhancement of intracellular calcium mobilization. Diabetes 1995, 44, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Gaisano, G.G.; Park, S.J.; Daly, D.M.; Beyak, M.J. Glucagon-like peptide-1 inhibits voltage-gated potassium currents in mouse nodose ganglion neurons. Neurogastroenterol. Motil. 2010, 22, 470-e111. [Google Scholar] [CrossRef]
- Lamas, J.A. Mechanically gated channels and their regulation. In Mechanically Gated Channels and Their Regulation; Kamkin, A., Lozinsky, I., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2012; pp. 35–68. ISBN 978-94-007-5072-2. [Google Scholar]
- Lesage, F.; Terrenoire, C.; Romey, G.; Lazdunski, M. Human TREK2, a 2P domain mechano-sensitive K+ channel with multiple regulations by polyunsaturated fatty acids, lysophospholipids, and Gs, Gi, and Gq protein-coupled receptors. J. Biol. Chem. 2000, 275, 28398–28405. [Google Scholar] [CrossRef]
- Patel, A.J.; Honoré, E.; Maingret, F.; Lesage, F.; Fink, M.; Duprat, F.; Lazdunski, M. A mammalian two pore domain mechano-gated S-like K+ channel. EMBO J. 1998, 17, 4283–4290. [Google Scholar] [CrossRef]
- Koh, S.D.; Monaghan, K.; Sergeant, G.P.; Ro, S.; Walker, R.L.; Sanders, K.M.; Horowitz, B. TREK-1 regulation by nitric oxide and cGMP-dependent protein kinase. J. Biol. Chem. 2001, 276, 44338–44346. [Google Scholar] [CrossRef]
- Heurteaux, C.; Lucas, G.; Guy, N.; El Yacoubi, M.; Thümmler, S.; Peng, X.-D.; Noble, F.; Blondeau, N.; Widmann, C.; Borsotto, M.; et al. Deletion of the background potassium channel TREK-1 results in a depression-resistant phenotype. Nat. Neurosci. 2006, 9, 1134–1141. [Google Scholar] [CrossRef]
- Mayo, K.E.; Miller, L.J.; Bataille, D.; Dalle, S.; Göke, B.; Thorens, B.; Drucker, D.J. The glucagon receptor family. Pharmacol. Rev. 2003, 55, 167–194. [Google Scholar] [CrossRef]
- Fine, N.H.F.; Doig, C.L.; Elhassan, Y.S.; Vierra, N.C.; Marchetti, P.; Bugliani, M.; Nano, R.; Piemonti, L.; Rutter, G.A.; Jacobson, D.A.; et al. Glucocorticoids reprogram β-Cell signaling to preserve insulin secretion. Diabetes 2018, 67, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Kennard, L.E.; Chumbley, J.R.; Ranatunga, K.M.; Armstrong, S.J.; Veale, E.L.; Mathie, A. Inhibition of the human two-pore domain potassium channel, TREK-1, by fluoxetine and its metabolite norfluoxetine. Br. J. Pharmacol. 2005, 144, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Cadaveira-Mosquera, A.; Ribeiro, S.J.; Reboreda, A.; Perez, M.; Lamas, J.A. Activation of TREK currents by the neuroprotective agent riluzole in mouse sympathetic neurons. J. Neurosci. 2011, 31, 1375–1385. [Google Scholar] [CrossRef]
- Joseph, A.; Thuy, T.T.T.; Thanh, L.T.; Okada, M. Antidepressive and anxiolytic effects of ostruthin, a TREK-1 channel activator. PLoS ONE 2018, 13, e0201092. [Google Scholar] [CrossRef] [PubMed]
- Mazella, J.; Pétrault, O.; Lucas, G.; Deval, E.; Béraud-Dufour, S.; Gandin, C.; El-Yacoubi, M.; Widmann, C.; Guyon, A.; Chevet, E.; et al. Spadin, a sortilin-derived peptide, targeting rodent TREK-1 channels: A new concept in the antidepressant drug design. PLoS Biol. 2010, 8, e1000355. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, R.H.; Richard, J.E.; Hansson, C.; Nissbrandt, H.; Bergquist, F.; Skibicka, K.P. GLP-1 is both anxiogenic and antidepressant; divergent effects of acute and chronic GLP-1 on emotionality. Psychoneuroendocrinology 2016, 65, 54–66. [Google Scholar] [CrossRef]
- Liu, W.; Liu, J.; Huang, Z.; Cui, Z.; Li, L.; Liu, W.; Qi, Z. Possible role of GLP-1 in antidepressant effects of metformin and exercise in CUMS mice. J. Affect. Disord. 2019, 246, 486–497. [Google Scholar] [CrossRef]
- Sharma, A.N.; Ligade, S.S.; Sharma, J.N.; Shukla, P.; Elased, K.M.; Lucot, J.B. GLP-1 receptor agonist liraglutide reverses long-term atypical antipsychotic treatment associated behavioral depression and metabolic abnormalities in rats. Metab. Brain Dis. 2015, 30, 519–527. [Google Scholar] [CrossRef]
- Kinzig, K.P.; D’Alessio, D.A.; Herman, J.P.; Sakai, R.R.; Vahl, T.P.; Figueiredo, H.F.; Murphy, E.K.; Seeley, R.J. CNS glucagon-like peptide-1 receptors mediate endocrine and anxiety responses to interoceptive and psychogenic stressors. J. Neurosci. 2003, 23, 6163–6170. [Google Scholar] [CrossRef]
- Gil-Lozano, M.; Pérez-Tilve, D.; Alvarez-Crespo, M.; Martís, A.; Fernandez, A.M.; Catalina, P.A.F.; Gonzalez-Matias, L.C.; Mallo, F. GLP-1(7-36)-amide and exendin-4 stimulate the HPA axis in rodents and humans. Endocrinology 2010, 151, 2629–2640. [Google Scholar] [CrossRef]
- Maniscalco, J.W.; Zheng, H.; Gordon, P.J.; Rinaman, L. Negative energy balance blocks neural and behavioral responses to acute stress by “silencing” central glucagon-like peptide 1 signaling in rats. J. Neurosci. 2015, 35, 10701–10714. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- de Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: Central role of the brain. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef]
- de Kloet, E.R.; Vreugdenhil, E.; Oitzl, M.S.; Joëls, M. Brain corticosteroid receptor balance in health and disease. Endocr. Rev. 1998, 19, 269–301. [Google Scholar] [CrossRef]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar]
- Herman, J.P. Regulation of hypothalamo-pituitary-adrenocortical responses to stressors by the nucleus of the solitary tract/dorsal vagal complex. Cell. Mol. Neurobiol. 2018, 38, 25–35. [Google Scholar] [CrossRef]
- Cunningham, E.T.; Sawchenko, P.E. Anatomical specificity of noradrenergic inputs to the paraventricular and supraoptic nuclei of the rat hypothalamus. J. Comp. Neurol. 1988, 274, 60–76. [Google Scholar] [CrossRef]
- Katsurada, K.; Maejima, Y.; Nakata, M.; Kodaira, M.; Suyama, S.; Iwasaki, Y.; Kario, K.; Yada, T. Endogenous GLP-1 acts on paraventricular nucleus to suppress feeding: Projection from nucleus tractus solitarius and activation of corticotropin-releasing hormone, nesfatin-1 and oxytocin neurons. Biochem. Biophys. Res. Commun. 2014, 451, 276–281. [Google Scholar] [CrossRef]
- Li, C.; Navarrete, J.; Liang-Guallpa, J.; Lu, C.; Funderburk, S.C.; Chang, R.B.; Liberles, S.D.; Olson, D.P.; Krashes, M.J. Defined paraventricular hypothalamic populations exhibit differential responses to food contingent on caloric State. Cell Metab. 2019, 29, 681–694.e5. [Google Scholar] [CrossRef] [PubMed]
- Tauchi, M.; Zhang, R.; D’Alessio, D.A.; Stern, J.E.; Herman, J.P. Distribution of glucagon-like peptide-1 immunoreactivity in the hypothalamic paraventricular and supraoptic nuclei. J. Chem. Neuroanat. 2008, 36, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Zueco, J.A.; Esquifino, A.I.; Chowen, J.A.; Alvarez, E.; Castrillón, P.O.; Blázquez, E. Coexpression of glucagon-like peptide-1 (GLP-1) receptor, vasopressin, and oxytocin mRNAs in neurons of the rat hypothalamic supraoptic and paraventricular nuclei: Effect of GLP-1(7-36)amide on vasopressin and oxytocin release. J. Neurochem. 1999, 72, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.J.; Tang-Christensen, M.; Jessop, D. Central administration of glucagon-like peptide-1 activates hypothalamic neuroendocrine neurons in the rat. Endocrinology 1997, 138, 4445–4455. [Google Scholar] [CrossRef]
- Rowland, N.E.; Crews, E.C.; Gentry, R.M. Comparison of Fos induced in rat brain by GLP-1 and amylin. Regul. Pept. 1997, 71, 171–174. [Google Scholar] [CrossRef]
- Ghosal, S.; Packard, A.E.B.; Mahbod, P.; McKlveen, J.M.; Seeley, R.J.; Myers, B.; Ulrich-Lai, Y.; Smith, E.P.; D’Alessio, D.A.; Herman, J.P. Disruption of glucagon-like peptide 1 signaling in Sim1 neurons reduces physiological and behavioral reactivity to acute and chronic stress. J. Neurosci. 2017, 37, 184–193. [Google Scholar] [CrossRef]
- Gil-Lozano, M.; Romaní-Pérez, M.; Outeiriño-Iglesias, V.; Vigo, E.; González-Matías, L.C.; Brubaker, P.L.; Mallo, F. Corticotropin-releasing hormone and the sympathoadrenal system are major mediators in the effects of peripherally administered exendin-4 on the hypothalamic-pituitary-adrenal axis of male rats. Endocrinology 2014, 155, 2511–2523. [Google Scholar] [CrossRef]
- Yamamoto, H.; Kishi, T.; Lee, C.E.; Choi, B.J.; Fang, H.; Hollenberg, A.N.; Drucker, D.J.; Elmquist, J.K. Glucagon-like peptide-1-responsive catecholamine neurons in the area postrema link peripheral glucagon-like peptide-1 with central autonomic control sites. J. Neurosci. 2003, 23, 2939–2946. [Google Scholar] [CrossRef]
- Zoccal, D.B.; Furuya, W.I.; Bassi, M.; Colombari, D.S.A.; Colombari, E.; Hildreth, C.; Putnam, R.W. The nucleus of the solitary tract and the coordination of respiratory and sympathetic activities. Front. Physiol. 2014, 5, 238. [Google Scholar] [CrossRef]
- Holt, M.K.; Pomeranz, L.E.; Beier, K.T.; Reimann, F.; Gribble, F.M.; Rinaman, L. Synaptic inputs to the mouse dorsal vagal complex and its resident preproglucagon neurons. J. Neurosci. 2019, 39, 9767–9781. [Google Scholar] [CrossRef]
- Williams, E.K.K.; Chang, R.B.B.; Strochlic, D.E.E.; Umans, B.D.D.; Lowell, B.B.B.; Liberles, S.D.D. Sensory neurons that detect stretch and nutrients in the digestive system. Cell 2016, 166, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Mesgarzadeh, S.; Ramesh, K.S.; Huey, E.L.; Liu, Y.; Gray, L.A.; Aitken, T.J.; Chen, Y.; Beutler, L.R.; Ahn, J.S.; et al. Genetic identification of vagal sensory neurons that control feeding. Cell 2019, 179, 1129–1143. [Google Scholar] [CrossRef] [PubMed]
- Abbott, S.B.G.; Stornetta, R.L.; Socolovsky, C.S.; West, G.H.; Guyenet, P.G. Photostimulation of channelrhodopsin-2 expressing ventrolateral medullary neurons increases sympathetic nerve activity and blood pressure in rats. J. Physiol. 2009, 587, 5613–5631. [Google Scholar] [CrossRef] [PubMed]
- Guyenet, P.G. The sympathetic control of blood pressure. Nat. Rev. Neurosci. 2006, 7, 335–346. [Google Scholar] [CrossRef]
- Ray, R.S.; Corcoran, A.E.; Brust, R.D.; Kim, J.C.; Richerson, G.B.; Nattie, E.; Dymecki, S.M. Impaired respiratory and body temperature control upon acute serotonergic neuron inhibition. Science 2011, 333, 637–642. [Google Scholar] [CrossRef]
- Lee, S.J.; Diener, K.; Kaufman, S.; Krieger, J.-P.; Pettersen, K.G.; Jejelava, N.; Arnold, M.; Watts, A.G.; Langhans, W. Limiting glucocorticoid secretion increases the anorexigenic property of Exendin-4. Mol. Metab. 2016, 5, 552–565. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Arnhold, M.M.; Engeland, W.C. Adrenal splanchnic innervation contributes to the diurnal rhythm of plasma corticosterone in rats by modulating adrenal sensitivity to ACTH. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R1128–R1135. [Google Scholar] [CrossRef]
- Pérez-Tilve, D.; González-Matías, L.; Aulinger, B.A.; Alvarez-Crespo, M.; Gil-Lozano, M.; Alvarez, E.; Andrade-Olivie, A.M.; Tschöp, M.H.; D’Alessio, D.A.; Mallo, F. Exendin-4 increases blood glucose levels acutely in rats by activation of the sympathetic nervous system. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1088–E1096. [Google Scholar] [CrossRef]
- Barragán, J.M.; Eng, J.; Rodríguez, R.; Blázquez, E. Neural contribution to the effect of glucagon-like peptide-1-(7-36) amide on arterial blood pressure in rats. Am. J. Physiol. 1999, 277, E784–E791. [Google Scholar] [CrossRef]
- Yamamoto, H.; Lee, C.E.; Marcus, J.N.; Williams, T.D.; Overton, J.M.; Lopez, M.E.; Hollenberg, A.N.; Baggio, L.; Saper, C.B.; Drucker, D.J.; et al. Glucagon-like peptide-1 receptor stimulation increases blood pressure and heart rate and activates autonomic regulatory neurons. J. Clin. Investig. 2002, 110, 43–52. [Google Scholar] [CrossRef]
- Griffioen, K.J.; Wan, R.; Okun, E.; Wang, X.; Lovett-Barr, M.R.; Li, Y.; Mughal, M.R.; Mendelowitz, D.; Mattson, M.P. GLP-1 receptor stimulation depresses heart rate variability and inhibits neurotransmission to cardiac vagal neurons. Cardiovasc. Res. 2011, 89, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Holt, M.K.; Cook, D.R.; Brierley, D.I.; Richards, J.E.; Reimann, F.; Gourine, A.V.; Marina, N.; Trapp, S. PPG neurons in the nucleus of the solitary tract modulate heart rate but do not mediate GLP-1 receptor agonist-induced tachycardia in mice. Mol. Metab. 2020, 39, 101024. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Ussher, J.R.; McLean, B.A.; Cao, X.; Kabir, M.G.; Mulvihill, E.E.; Mighiu, A.S.; Zhang, H.; Ludwig, A.; Seeley, R.J.; et al. The autonomic nervous system and cardiac GLP-1 receptors control heart rate in mice. Mol. Metab. 2017, 6, 1339–1349. [Google Scholar] [CrossRef]
- Smits, M.M.; Muskiet, M.H.A.; Tonneijck, L.; Hoekstra, T.; Kramer, M.H.H.; Diamant, M.; Van Raalte, D.H. Exenatide acutely increases heart rate in parallel with augmented sympathetic nervous system activation in healthy overweight males. Br. J. Clin. Pharmacol. 2016, 81, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Heuvelman, V.D.; Van Raalte, D.H.; Smits, M.M. Cardiovascular effects of glucagon-like peptide 1 receptor agonists: From mechanistic studies in humans to clinical outcomes. Cardiovasc. Res. 2020, 116, 916–930. [Google Scholar] [CrossRef]
- Smits, M.M.; Tonneijck, L.; Muskiet, M.H.A.; Hoekstra, T.; Kramer, M.H.H.; Diamant, M.; Van Raalte, D.H. Heart rate acceleration with GLP-1 receptor agonists in type 2 diabetes patients: An acute and 12-week randomised, double-blind, placebo-controlled trial. Eur. J. Endocrinol. 2017, 176, 77–86. [Google Scholar] [CrossRef]
- Baraboi, E.-D.; St-Pierre, D.H.; Shooner, J.; Timofeeva, E.; Richard, D. Brain activation following peripheral administration of the GLP-1 receptor agonist exendin-4. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R1011–R1024. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Lockie, S.H.; Heppner, K.M.; Chaudhary, N.; Chabenne, J.R.; Morgan, D.A.; Veyrat-Durebex, C.; Ananthakrishnan, G.; Rohner-Jeanrenaud, F.; Drucker, D.J.; Dimarchi, R.; et al. Direct control of brown adipose tissue thermogenesis by central nervous system glucagon-like Peptide-1 receptor signaling. Diabetes 2012, 61, 2753–2762. [Google Scholar] [CrossRef]
- Beiroa, D.; Imbernon, M.; Gallego, R.; Senra, A.; Herranz, D.; Villarroya, F.; Serrano, M.; Fernø, J.; Salvador, J.; Escalada, J.; et al. GLP-1 agonism stimulates brown adipose tissue thermogenesis and browning through hypothalamic AMPK. Diabetes 2014, 63, 3346–3358. [Google Scholar] [CrossRef]
- Lee, S.J.; Sanchez-Watts, G.; Krieger, J.P.; Pignalosa, A.; Norell, P.N.; Cortella, A.; Pettersen, K.G.; Vrdoljak, D.; Hayes, M.R.; Kanoski, S.; et al. Loss of dorsomedial hypothalamic GLP-1 signaling reduces BAT thermogenesis and increases adiposity. Mol. Metab. 2018, 11, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Gaykema, R.P.; Newmyer, B.A.; Ottolini, M.; Raje, V.; Warthen, D.M.; Lambeth, P.S.; Niccum, M.; Yao, T.; Huang, Y.; Schulman, I.G.; et al. Activation of murine pre-proglucagon-producing neurons reduces food intake and body weight. J. Clin. Investig. 2017, 127, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A. Distributed circuits underlying anxiety. Front. Behav. Neurosci. 2014, 8, 112. [Google Scholar] [CrossRef]
- Calhoon, G.G.; Tye, K.M. Resolving the neural circuits of anxiety. Nat. Neurosci. 2015, 18, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Gulec, G.; Isbil-Buyukcoskun, N.; Kahveci, N. Effects of centrally-injected glucagon-like peptide-1 on pilocarpine-induced seizures, anxiety and locomotor and exploratory activity in rat. Neuropeptides 2010, 44, 285–291. [Google Scholar] [CrossRef]
- Möller, C.; Sommer, W.; Thorsell, A.; Rimondini, R.; Heilig, M. Anxiogenic-like action of centrally administered glucagon-like peptide-1 in a punished drinking test. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2002, 26, 119–122. [Google Scholar] [CrossRef]
- Zhang, R.; Jankord, R.; Flak, J.N.; Solomon, M.B.; D’Alessio, D.A.; Herman, J.P. Role of glucocorticoids in tuning hindbrain stress integration. J. Neurosci. 2010, 30, 14907–14914. [Google Scholar] [CrossRef]
- Tauchi, M.; Zhang, R.; D’Alessio, D.A.; Seeley, R.J.; Herman, J.P. Role of central glucagon-like peptide-1 in hypothalamo-pituitary-adrenocortical facilitation following chronic stress. Exp. Neurol. 2008, 210, 458–466. [Google Scholar] [CrossRef]
- Gil-Lozano, M.; Romaní-Pérez, M.; Outeiriño-Iglesias, V.; Vigo, E.; Brubaker, P.L.; González-Matías, L.C.; Mallo, F. Effects of prolonged exendin-4 administration on hypothalamic-pituitary-adrenal axis activity and water balance. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1105–E1117. [Google Scholar] [CrossRef]
- Rinaman, L. Interoceptive stress activates glucagon-like peptide-1 neurons that project to the hypothalamus. Am. J. Physiol. 1999, 277, R582–R590. [Google Scholar] [CrossRef]
- Rinaman, L. A functional role for central glucagon-like peptide-1 receptors in lithium chloride-induced anorexia. Am. J. Physiol. Integr. Comp. Physiol. 1999, 277, R1537–R1540. [Google Scholar] [CrossRef] [PubMed]
- Seeley, R.J.; Blake, K.; Rushing, P.A.; Benoit, S.; Eng, J.; Woods, S.C.; D’Alessio, D. The role of CNS glucagon-like peptide-1 (7-36) amide receptors in mediating the visceral illness effects of lithium chloride. J. Neurosci. 2000, 20, 1616–1621. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tang-Christensen, M. Central administration of GLP-1(7[ndash]36)amide inhibits food and water intake in rats. Am. J. Phys. 1996, 271, 848–856. [Google Scholar]
- Thiele, T.E. Central infusion of GLP-1, but not leptin, produces conditioned taste aversions in rats. Am. J. Physiol. 1997, 272, R726–R730. [Google Scholar] [CrossRef] [PubMed]
- Lachey, J.L.; D’Alessio, D.A.; Rinaman, L.; Elmquist, J.K.; Drucker, D.J.; Seeley, R.J. The role of central glucagon-like peptide-1 in mediating the effects of visceral illness: Differential effects in rats and mice. Endocrinology 2005, 146, 458–462. [Google Scholar] [CrossRef]
- Kinzig, K.P.; D’Alessio, D.A.; Seeley, R.J. The diverse roles of specific GLP-1 receptors in the control of food intake and the response to visceral illness. J. Neurosci. 2002, 22, 10470–10476. [Google Scholar] [CrossRef]
- Vrang, N.; Phifer, C.B.; Corkern, M.M.; Berthoud, H.-R. Gastric distension induces c-Fos in medullary GLP-1/2-containing neurons. Am. J. Physiol. Integr. Comp. Physiol. 2003, 285, R470–R478. [Google Scholar] [CrossRef]
- Gülpinar, M.A.; Bozkurt, A.; Coşkun, T.; Ulusoy, N.B.; Yeǧen, B.Ç. Glucagon-like peptide (GLP-1) is involved in the central modulation of fecal output in rats. Am. J. Physiol. Liver Physiol. 2000, 278, G924–G929. [Google Scholar] [CrossRef]
- Steinert, R.E.; Feinle-Bisset, C.; Asarian, L.; Horowitz, M.; Beglinger, C.; Geary, N. Ghrelin, CCK, GLP-1, and PYY(3-36): Secretory controls and physiological roles in eating and glycemia in health, obesity, and after RYGB. Physiol. Rev. 2017, 97, 411–463. [Google Scholar] [CrossRef]
- Williams, D.L.; Baskin, D.G.; Schwartz, M.W. Evidence that intestinal glucagon-like peptide-1 plays a physiological role in satiety. Endocrinology 2009, 150, 1680–1687. [Google Scholar] [CrossRef]
- Krieger, J.P. Intestinal glucagon-like peptide-1 effects on food intake: Physiological relevance and emerging mechanisms. Peptides 2020, 131. [Google Scholar] [CrossRef]
- Flint, A.; Raben, A.; Astrup, A.; Holst, J.J. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J. Clin. Investig. 1998, 101, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Gutzwiller, J.P.; Göke, B.; Drewe, J.; Hildebrand, P.; Ketterer, S.; Handschin, D.; Winterhalder, R.; Conen, D.; Beglinger, C. Glucagon-like peptide-1: A potent regulator of food intake in humans. Gut 1999, 44, 81–86. [Google Scholar] [CrossRef]
- Ronveaux, C.C.; de Lartigue, G.; Raybould, H.E. Ability of GLP-1 to decrease food intake is dependent on nutritional status. Physiol. Behav. 2014, 135, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, D.; Barrera, J.G.; Stefater, M.A.; Sisley, S.; Woods, S.C.; D’Alessio, D.D.; Seeley, R.J. The Anorectic effect of GLP-1 in rats is nutrient dependent. PLoS ONE 2012, 7, e51870. [Google Scholar] [CrossRef] [PubMed]
- Punjabi, M.; Arnold, M.; Rüttimann, E.; Graber, M.; Geary, N.; Pacheco-López, G.; Langhans, W. Circulating glucagon-like peptide-1 (GLP-1) inhibits eating in male rats by acting in the hindbrain and without inducing avoidance. Endocrinology 2014, 155, 1690–1699. [Google Scholar] [CrossRef]
- Cabou, C.; Campistron, G.; Marsollier, N.; Leloup, C.; Cruciani-Guglielmacci, C.; Penicaud, L.; Drucker, D.J.; Magnan, C.; Burcelin, R. Brain glucagon-like peptide-1 regulates arterial blood flow, heart rate, and insulin sensitivity. Diabetes 2008, 57, 2577–2587. [Google Scholar] [CrossRef]
- Berthoud, H.-R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. GLP-1 and weight loss: Unraveling the diverse neural circuitry. Am. J. Physiol. Integr. Comp. Physiol. 2016, 310, R885–R895. [Google Scholar] [CrossRef]
- Krieger, J.-P.; Arnold, M.; Pettersen, K.G.; Lossel, P.; Langhans, W.; Lee, S.J. Knockdown of GLP-1 receptors in vagal afferents affects normal food intake and glycemia. Diabetes 2016, 65, 34–43. [Google Scholar] [CrossRef]
- Abbott, C.R.; Monteiro, M.; Small, C.J.; Sajedi, A.; Smith, K.L.; Parkinson, J.R.C.; Ghatei, M.A.; Bloom, S.R. The inhibitory effects of peripheral administration of peptide YY3–36 and glucagon-like peptide-1 on food intake are attenuated by ablation of the vagal–brainstem–hypothalamic pathway. Brain Res. 2005, 1044, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ritter, R.C. Circulating GLP-1 and CCK-8 reduce food intake by capsaicin-insensitive, nonvagal mechanisms. Am. J. Physiol. Integr. Comp. Physiol. 2012, 302, R264–R273. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sisley, S.; Gutierrez-Aguilar, R.; Scott, M.; D’Alessio, D.A.; Sandoval, D.A.; Seeley, R.J. Neuronal GLP1R mediates liraglutide’s anorectic but not glucose-lowering effect. J. Clin. Investig. 2014, 124, 2456–2463. [Google Scholar] [CrossRef] [PubMed]
- Secher, A.; Jelsing, J.; Baquero, A.F.; Hecksher-Sørensen, J.; Cowley, M.A.; Dalbøge, L.S.; Hansen, G.; Grove, K.L.; Pyke, C.; Raun, K.; et al. The arcuate nucleus mediates GLP-1 receptor agonist liraglutide-dependent weight loss. J. Clin. Investig. 2014, 124, 4473–4488. [Google Scholar] [CrossRef] [PubMed]
- Orskov, C.; Poulsen, S.S.; Møller, M.; Holst, J.J. Glucagon-like peptide I receptors in the subfornical organ and the area postrema are accessible to circulating glucagon-like peptide I. Diabetes 1996, 45, 832–835. [Google Scholar] [CrossRef]
- Kastin, A.J.; Akerstrom, V.; Pan, W. Interactions of glucagon-like peptide-1 (GLP-1) with the blood-brain barrier. J. Mol. Neurosci. 2002, 18, 7–14. [Google Scholar] [CrossRef]
- Varin, E.M.; Mulvihill, E.E.; Baggio, L.L.; Koehler, J.A.; Cao, X.; Seeley, R.J.; Drucker, D.J. Distinct neural sites of GLP-1R expression mediate physiological versus pharmacological control of incretin action. Cell Rep. 2019, 27, 3371–3384. [Google Scholar] [CrossRef]
- Hisadome, K.; Reimann, F.; Gribble, F.M.; Trapp, S. CCK stimulation of GLP-1 neurons involves α 1-adrenoceptor- mediated increase in glutamatergic synaptic inputs. Diabetes 2011, 60, 2701–2709. [Google Scholar] [CrossRef]
- Maniscalco, J.W.; Edwards, C.M.; Rinaman, L. Ghrelin signaling contributes to fasting-induced attenuation of hindbrain neural activation and hypophagic responses to systemic cholecystokinin in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R1014–R1023. [Google Scholar] [CrossRef]
- Goldstone, A.P.; Mercer, J.G.; Gunn, I.; Moar, K.M.; Edwards, C.M.B.; Rossi, M.; Howard, J.K.; Rasheed, S.; Turton, M.D.; Small, C.; et al. Leptin interacts with glucagon-like peptide-1 neurons to reduce food intake and body weight in rodents. FEBS Lett. 1997, 415, 134–138. [Google Scholar] [CrossRef]
- Huo, L.; Gamber, K.M.; Grill, H.J.; Bjørbæk, C. Divergent leptin signaling in proglucagon neurons of the nucleus of the solitary tract in mice and rats. Endocrinology 2008, 149, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Maniscalco, J.W.; Rinaman, L. Systemic leptin dose-dependently increases STAT3 phosphorylation within hypothalamic and hindbrain nuclei. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R576–R585. [Google Scholar] [CrossRef]
- Cheng, W.; Ndoka, E.; Hutch, C.; Roelofs, K.; MacKinnon, A.; Khoury, B.; Magrisso, J.; Kim, K.S.; Rhodes, C.J.; Olson, D.P.; et al. Leptin receptor–expressing nucleus tractus solitarius neurons suppress food intake independently of GLP1 in mice. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Turton, M.D. A role for glucagon-like peptide-1 in the central regulation of feeding. Nature 1996, 379, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Donahey, J.C.; van Dijk, G.; Woods, S.C.; Seeley, R.J. Intraventricular GLP-1 reduces short- but not long-term food intake or body weight in lean and obese rats. Brain Res. 1998, 779, 75–83. [Google Scholar] [CrossRef]
- Williams, D.L. Neural integration of satiation and food reward: Role of GLP-1 and orexin pathways. Physiol. Behav. 2014, 136, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Scrocchi, L.A.; Brown, T.J.; MacLusky, N.; Brubaker, P.L.; Auerbach, A.B.; Joyner, A.L.; Drucker, D.J. Glucose intolerance but normal satiety in mice with a null mutation in the glucagon-like peptide 1 receptor gene. Nat. Med. 1996, 2, 1254–1258. [Google Scholar] [CrossRef] [PubMed]
- Mietlicki-Baase, E.G.; Ortinski, P.I.; Rupprecht, L.E.; Olivos, D.R.; Alhadeff, A.L.; Christopher Pierce, R.; Hayes, M.R. The food intake-suppressive effects of glucagon-like peptide-1 receptor signaling in the ventral tegmental area are mediated by AMPA/kainate receptors. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1367–E1374. [Google Scholar] [CrossRef] [PubMed]
- Mietlicki-Baase, E.G.; Ortinski, P.I.; Reiner, D.J.; Sinon, C.G.; McCutcheon, J.E.; Pierce, R.C.; Roitman, M.F.; Hayes, M.R. Glucagon-like peptide-1 receptor activation in the nucleus accumbens core suppresses feeding by increasing glutamatergic AMPA/kainate signaling. J. Neurosci. 2014, 34, 6985–6992. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Iwai, T.; Sasaki-Hamada, S.; Kamanaka, G.; Oka, J.I. Exendin (5-39), an antagonist of GLP-1 receptor, modulates synaptic transmission via glutamate uptake in the dentate gyrus. Brain Res. 2013, 1505, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zanotto, C.; Hansen, F.; Galland, F.; Batassini, C.; Federhen, B.C.; da Silva, V.F.; Leite, M.C.; Nardin, P.; Gonçalves, C.A. Glutamatergic alterations in STZ-induced diabetic rats are reversed by exendin-4. Mol. Neurobiol. 2018, 1–14. [Google Scholar] [CrossRef]
- Liu, J.; Conde, K.; Zhang, P.; Lilascharoen, V.; Xu, Z.; Lim, B.K.; Seeley, R.J.; Zhu, J.J.; Scott, M.M.; Pang, Z.P. Enhanced AMPA receptor trafficking mediates the anorexigenic effect of endogenous glucagon-like peptide-1 in the paraventricular hypothalamus. Neuron 2017, 96, 897–909. [Google Scholar] [CrossRef] [PubMed]
- McKay, N.J.; Kanoski, S.E.; Hayes, M.R.; Daniels, D. Glucagon-like peptide-1 receptor agonists suppress water intake independent of effects on food intake. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R1755–R1764. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Pei, H.; Sandoval, D.A.; Seeley, R.J.; Chang, R.B.; Liberles, S.D.; Olson, D.P. Liraglutide modulates appetite and body weight through glucagon-like peptide 1 receptor-expressing glutamatergic neurons. Diabetes 2018, 67, 1538–1548. [Google Scholar] [CrossRef] [PubMed]
- Maniscalco, J.W.; Kreisler, A.D.; Rinaman, L. Satiation and stress-induced hypophagia: Examining the role of hindbrain neurons expressing prolactin-releasing peptide or glucagon-like peptide 1. Front. Neurosci. 2013, 6, 199. [Google Scholar] [CrossRef] [PubMed]
- Kreisler, A.D.; Rinaman, L. Hindbrain glucagon-like peptide-1 neurons track intake volume and contribute to injection stress-induced hypophagia in meal-entrained rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R906–R916. [Google Scholar] [CrossRef] [PubMed]
- Maniscalco, J.W.; Rinaman, L. Interoceptive modulation of neuroendocrine, emotional, and hypophagic responses to stress. Physiol. Behav. 2017, 176, 195–206. [Google Scholar] [CrossRef]
- Anesten, F.; Holt, M.K.; Schéle, E.; Pálsdóttir, V.; Reimann, F.; Gribble, F.M.; Safari, C.; Skibicka, K.P.; Trapp, S.; Jansson, J.O. Preproglucagon neurons in the hindbrain have IL-6 receptor-α and show Ca2+ influx in response to IL-6. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R115–R123. [Google Scholar] [CrossRef]
- Nieuwenhuizen, A.G.; Rutters, F. The hypothalamic-pituitary-adrenal-axis in the regulation of energy balance. Physiol. Behav. 2008, 94, 169–177. [Google Scholar] [CrossRef]
- Morton, G.J.; Meek, T.H.; Schwartz, M.W. Neurobiology of food intake in health and disease. Nat. Rev. Neurosci. 2014, 15, 367–378. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Fulton, S.; Wilson, M.; Petrovich, G.; Rinaman, L. Stress exposure, food intake and emotional state. Stress 2015, 18, 381–399. [Google Scholar] [PubMed]
- Dallman, M.F.; Pecoraro, N.; Akana, S.F.; La Fleur, S.E.; Gomez, F.; Houshyar, H.; Bell, M.E.; Bhatnagar, S.; Laugero, K.D.; Manalo, S. Chronic stress and obesity: A new view of “comfort food”. Proc. Natl. Acad. Sci. USA 2003, 100, 11696–11701. [Google Scholar] [CrossRef]
- Dabrowska, J.; Hazra, R.; Guo, J.D.; DeWitt, S.; Rainnie, D.G. Central CRF neurons are not created equal: Phenotypic differences in CRF-containing neurons of the rat paraventricular hypothalamus and the bed nucleus of the stria terminalis. Front. Neurosci. 2013, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Long, B.; Yuan, J.; Peng, X.; Ni, H.; Li, X.; Gong, H.; Luo, Q.; Li, A. A quantitative analysis of the distribution of CRH neurons in whole mouse brain. Front. Neuroanat. 2017, 11. [Google Scholar] [CrossRef]
- Koob, G.F. The role of CRF and CRF-related peptides in the dark side of addiction. Brain Res. 2010, 1314, 3–14. [Google Scholar] [CrossRef]
- Zorrilla, E.P.; Logrip, M.L.; Koob, G.F. Corticotropin releasing factor: A key role in the neurobiology of addiction. Front. Neuroendocrinol. 2014, 35, 234–244. [Google Scholar] [CrossRef]
- Diz-Chaves, Y.; Gil-Lozano, M.; Toba, L.; Fandiño, J.; Ogando, H.; González-Matías, L.C.; Mallo, F. Stressing diabetes? The hidden links between insulinotropic peptides and the HPA axis. J. Endocrinol. 2016, 230, R77–R94. [Google Scholar] [CrossRef] [PubMed]
- Tachi, M.; Kondo, F.; Gosho, M.; Okada, S.; Fukayama, M.; Yoshikawa, K. Changes in hypothalamic neurotransmitter and prostanoid levels in response to NMDA, CRF, and GLP-1 stimulation. Anal. Bioanal. Chem. 2015, 407, 5261–5272. [Google Scholar]
- Korol, S.V.; Jin, Z.; Babateen, O.; Birnir, B. GLP-1 and exendin-4 transiently enhance gabaa receptor-mediated synaptic and tonic currents in rat hippocampal ca3 pyramidal neurons. Diabetes 2015, 64, 79–89. [Google Scholar] [CrossRef]
- Korol, S.V.; Jin, Z.; Birnir, B. The GLP-1 receptor agonist exendin-4 and diazepam differentially regulate GABAA receptor-mediated tonic currents in rat hippocampal CA3 pyramidal neurons. PLoS ONE 2015, 10, e0124765. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Xu, J.; Sun, Q.; Yu, F.; Cheng, J.; Peng, B.; Liu, W.; Xiao, Z.; Yin, J.; et al. Inhibition of DPP4 enhances inhibitory synaptic transmission through activating the GLP-1/GLP-1R signaling pathway in a rat model of febrile seizures. Biochem. Pharmacol. 2018, 156, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Grill, H.J.; Carmody, J.S.; Amanda Sadacca, L.; Williams, D.L.; Kaplan, J.M. Attenuation of lipopolysaccharide anorexia by antagonism of caudal brain stem but not forebrain GLP-1-R. Am. J. Physiol. Integr. Comp. Physiol. 2004, 287, R1190–R1193. [Google Scholar] [CrossRef] [PubMed]
- Dallman, M.F.; Akana, S.F.; Bhatnagar, S.; Bell, M.E.; Choi, S.; Chu, A.; Horsley, C.; Levin, N.; Meijer, O.; Soriano, L.R.; et al. Starvation: Early signals, sensors, and sequelae. Endocrinology 1999, 140, 4015–4023. [Google Scholar] [CrossRef]
- Chacón, F.; Esquifino, A.I.; Perelló, M.; Cardinali, D.P.; Spinedi, E.; Alvarez, M.P. 24-hour changes in ACTH, corticosterone, growth hormone, and leptin levels in young male rats subjected to calorie restriction. Chronobiol. Int. 2005, 22, 253–265. [Google Scholar] [CrossRef]
- Parker, J.A.; McCullough, K.A.; Field, B.C.T.; Minnion, J.S.; Martin, N.M.; Ghatei, M.A.; Bloom, S.R. Glucagon and GLP-1 inhibit food intake and increase c-fos expression in similar appetite regulating centres in the brainstem and amygdala. Int. J. Obes. 2013, 37, 1391–1398. [Google Scholar] [CrossRef]
- Ronveaux, C.C.; Tomé, D.; Raybould, H.E. Glucagon-like peptide 1 interacts with ghrelin and leptin to regulate glucose metabolism and food intake through vagal afferent neuron signaling. J. Nutr. 2015, 145, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Betley, J.N.; Cao, Z.F.H.; Ritola, K.D.; Sternson, S.M. Parallel, redundant circuit organization for homeostatic control of feeding behavior. Cell 2013, 55, 1337–1350. [Google Scholar] [CrossRef]
- Rosen, A.M.; Victor, J.D.; Di Lorenzo, P.M. Temporal coding of taste in the parabrachial nucleus of the pons of the rat. J. Neurophysiol. 2011, 105, 1889–1896. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, D.A.; Bagnol, D.; Woods, S.C.; D’Alessio, D.A.; Seeley, R.J. Arcuate glucagon-like peptide 1 receptors regulate glucose homeostasis but not food intake. Diabetes 2008, 57, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Stuber, G.D.; Wise, R.A. Lateral hypothalamic circuits for feeding and reward. Nat. Neurosci. 2016, 19, 198–205. [Google Scholar] [CrossRef]
- López-Ferreras, L.; Richard, J.E.; Noble, E.E.; Eerola, K.; Anderberg, R.H.; Olandersson, K.; Taing, L.; Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. Lateral hypothalamic GLP-1 receptors are critical for the control of food reinforcement, ingestive behavior and body weight. Mol. Psychiatry 2018, 23, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Schick, R.R.; Zimmermann, J.P.; vorm Walde, T.; Schusdziarra, V. Glucagon-like peptide 1-(7–36) amide acts at lateral and medial hypothalamic sites to suppress feeding in rats. Am. J. Physiol. Integr. Comp. Physiol. 2003, 284, R1427–R1435. [Google Scholar] [CrossRef]
- Burmeister, M.A.; Brown, J.D.; Ayala, J.E.; Stoffers, D.A.; Sandoval, D.A.; Seeley, R.J.; Ayala, J.E. The glucagon-like peptide-1 receptor in the ventromedial hypothalamus reduces short-term food intake in male mice by regulating nutrient sensor activity. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E651–E662. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, M.A.; Ayala, J.E.; Smouse, H.; Landivar-Rocha, A.; Brown, J.D.; Drucker, D.J.; Stoffers, D.A.; Sandoval, D.A.; Seeley, R.J.; Ayala, J.E. The hypothalamic glucagon-like peptide 1 receptor is sufficient but not necessary for the regulation of energy balance and glucose homeostasis in mice. Diabetes 2017, 66, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Ho, C.Y.; Richard, J.M.; Difeliceantonio, A.G. The tempted brain eats: Pleasure and desire circuits in obesity and eating disorders. Brain Res. 2010, 1350, 43–64. [Google Scholar] [CrossRef]
- Li, M.M.; Madara, J.C.; Steger, J.S.; Krashes, M.J.; Balthasar, N.; Campbell, J.N.; Resch, J.M.; Conley, N.J.; Garfield, A.S.; Lowell, B.B. The paraventricular hypothalamus regulates satiety and prevents obesity via two genetically distinct circuits. Neuron 2019, 102, 653–667. [Google Scholar] [CrossRef]
- Wu, Q.; Clark, M.S.; Palmiter, R.D. Deciphering a neuronal circuit that mediates appetite. Nature 2012, 483, 594–597. [Google Scholar] [CrossRef]
- Richard, J.E.; Farkas, I.; Anesten, F.; Anderberg, R.H.; Dickson, S.L.; Gribble, F.M.; Reimann, F.; Jansson, J.-O.; Liposits, Z.; Skibicka, K.P. GLP-1 Receptor stimulation of the lateral parabrachial nucleus reduces food intake: Neuroanatomical, electrophysiological, and behavioral evidence. Endocrinology 2014, 155, 4356–4367. [Google Scholar] [CrossRef]
- Kirouac, G.J. Placing the paraventricular nucleus of the thalamus within the brain circuits that control behavior. Neurosci. Biobehav. Rev. 2015, 56, 315–329. [Google Scholar] [CrossRef]
- Dossat, A.M.; Lilly, N.; Kay, K.; Williams, D.L. Glucagon-like peptide 1 receptors in nucleus accumbens affect food intake. J. Neurosci. 2011, 31, 14453–14457. [Google Scholar] [CrossRef]
- Dickson, S.L.; Shirazi, R.H.; Hansson, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. The glucagon-like peptide 1 (GLP-1) analogue, exendin-4, decreases the rewarding value of food: A new role for mesolimbic GLP-1 receptors. J. Neurosci. 2012, 32, 4812–4820. [Google Scholar] [CrossRef]
- Dossat, A.M.; Diaz, R.; Gallo, L.; Panagos, A.; Kay, K.; Williams, D.L. Nucleus accumbens GLP-1 receptors influence meal size and palatability. Am. J. Physiol. Metab. 2013, 304, E1314–E1320. [Google Scholar] [CrossRef]
- Hurley, M.M.; Maunze, B.; Block, M.E.; Frenkel, M.M.; Reilly, M.J.; Kim, E.; Chen, Y.; Li, Y.; Baker, D.A.; Liu, Q.S.; et al. Pituitary adenylate-cyclase activating polypeptide regulates hunger- and palatability-induced binge eating. Front. Neurosci. 2016, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, X.; Leng, H.; Luan, X.; Guo, F.; Sun, X.; Gao, S.; Liu, X.; Qin, H.; Xu, L. Zona incerta projection neurons and GABAergic and GLP-1 mechanisms in the nucleus accumbens are involved in the control of gastric function and food intake. Neuropeptides 2020, 80, 102018. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Grill, H.J. Hippocampus contributions to food intake control: Mnemonic, neuroanatomical, and endocrine mechanisms. Biol. Psychiatry 2017, 81, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Suarez, A.N.; Hsu, T.M.; Liu, C.M.; Noble, E.E.; Cortella, A.M.; Nakamoto, E.M.; Hahn, J.D.; De Lartigue, G.; Kanoski, S.E. Gut vagal sensory signaling regulates hippocampus function through multi-order pathways. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.M.; Hahn, J.D.; Konanur, V.R.; Noble, E.E.; Suarez, A.N.; Thai, J.; Nakamoto, E.M.; Kanoski, S.E. Hippocampus ghrelin signaling mediates appetite through lateral hypothalamic orexin pathways. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Suarez, A.N.; Liu, C.M.; Cortella, A.M.; Noble, E.E.; Kanoski, S.E. Ghrelin and orexin interact to increase meal size through a descending hippocampus to hindbrain signaling pathway. Biol. Psychiatry 2020, 87, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.M.; Hahn, J.D.; Konanur, V.R.; Lam, A.; Kanoski, S.E. Hippocampal GLP-1 receptors influence food intake, meal size, and effort-based responding for food through volume transmission. Neuropsychopharmacology 2015, 40, 327–337. [Google Scholar] [CrossRef]
- Born, J.M.; Lemmens, S.G.T.; Rutters, F.; Nieuwenhuizen, A.G.; Formisano, E.; Goebel, R.; Westerterp-Plantenga, M.S. Acute stress and food-related reward activation in the brain during food choice during eating in the absence of hunger. Int. J. Obes. 2010, 34, 172–181. [Google Scholar] [CrossRef]
- Sinha, R. Role of addiction and stress neurobiology on food intake and obesity. Biol. Psychol. 2018, 131, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Cottone, P.; Sabino, V.; Roberto, M.; Bajo, M.; Pockros, L.; Frihauf, J.B.; Fekete, E.M.; Steardo, L.; Rice, K.C.; Grigoriadis, D.E.; et al. CRF system recruitment mediates dark side of compulsive eating. Proc. Natl. Acad. Sci. USA 2009, 106, 20016–20020. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.F.; Sabino, V.; Koob, G.F.; Cottone, P. Neuroscience of compulsive eating behavior. Front. Neurosci. 2017, 11, 469. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Hum, A.; Gustafson, T.J.; Mietlicki-Baase, E.G. Binge-like palatable food intake in rats reduces preproglucagon in the nucleus tractus solitarius. Physiol. Behav. 2020, 219, 112830. [Google Scholar] [CrossRef] [PubMed]
- Maske, C.B.; Coiduras, I.I.; Ondriezek, Z.E.; Terrill, S.J.; Williams, D.L. Intermittent high-fat diet intake reduces sensitivity to intragastric nutrient infusion and exogenous amylin in female rats. Obesity 2020, 28, 942–952. [Google Scholar] [CrossRef]
- Small, D.M.; Jones-Gotman, M.; Dagher, A. Feeding-induced dopamine release in dorsal striatum correlates with meal pleasantness ratings in healthy human volunteers. Neuroimage 2003, 19, 1709–1715. [Google Scholar] [CrossRef]
- Johnson, P.M.; Kenny, P.J. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat. Neurosci. 2010, 13, 635–641. [Google Scholar] [CrossRef]
- Wang, G.J.; Volkow, N.D.; Thanos, P.K.; Fowler, J.S. Similarity between obesity and drug addiction as assessed by neurofunctional imaging: A concept review. J. Addict. Dis. 2004, 23, 39–53. [Google Scholar] [CrossRef]
- Petrovich, G.D.; Ross, C.A.; Mody, P.; Holland, P.C.; Gallagher, M. Central, but not basolateral, amygdala is critical for control of feeding by aversive learned cues. J. Neurosci. 2009, 29, 15205–15212. [Google Scholar] [CrossRef]
- Anderberg, R.H.; Anefors, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. Dopamine signaling in the amygdala, increased by food ingestion and GLP-1, regulates feeding behavior. Physiol. Behav. 2014, 136, 135–144. [Google Scholar] [CrossRef]
- Teegarden, S.L.; Bale, T.L. Decreases in dietary preference produce increased emotionality and risk for dietary relapse. Biol. Psychiatry 2007, 61, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Barrera, J.G.; Sandoval, D.A.; D’Alessio, D.A.; Seeley, R.J. GLP-1 and energy balance: An integrated model of short-term and long-term control. Nat. Rev. Endocrinol. 2011, 7, 507–516. [Google Scholar] [CrossRef]
- Duca, F.A.; Swartz, T.D.; Sakar, Y.; Covasa, M. Decreased intestinal nutrient response in diet-induced obese rats: Role of gut peptides and nutrient receptors. Int. J. Obes. 2013, 37, 375–381. [Google Scholar] [CrossRef]
- Williams, D.L.; Hyvarinen, N.; Lilly, N.; Kay, K.; Dossat, A.; Parise, E.; Torregrossa, A.M. Maintenance on a high-fat diet impairs the anorexic response to glucagon-like-peptide-1 receptor activation. Physiol. Behav. 2011, 103, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Mul, J.D.; Begg, D.P.; Barrera, J.G.; Li, B.; Matter, E.K.; D’Alessio, D.A.; Woods, S.C.; Seeley, R.J.; Sandoval, D.A. High-fat diet changes the temporal profile of GLP-1 receptor-mediated hypophagia in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R68–R77. [Google Scholar] [CrossRef]
- Anandhakrishnan, A.; Korbonits, M. Glucagon-like peptide 1 in the pathophysiology and pharmacotherapy of clinical obesity. World J. Diabetes 2016, 7, 572. [Google Scholar] [CrossRef] [PubMed]
- Ranganath, L.R.; Beety, J.M.; Morgan, L.M.; Wright, J.W.; Howland, R.; Marks, V. Attenuated GLP-1 secretion in obesity: Cause or consequence? Gut 1996, 38, 916–919. [Google Scholar] [CrossRef]
- Torekov, S.S.; Madsbad, S.; Holst, J.J. Obesity—An indication for GLP-1 treatment? Obesity pathophysiology and GLP-1 treatment potential. Obes. Rev. 2011, 12, 593–601. [Google Scholar] [CrossRef]
- Gagnon, J.; Baggio, L.L.; Drucker, D.J.; Brubaker, P.L. Ghrelin is a novel regulator of GLP-1 secretion. Diabetes 2015, 64, 1513–1521. [Google Scholar] [CrossRef]
- Anini, Y.; Brubaker, P.L. Role of leptin in the regulation of glucagon-like peptide-1 secretion. Diabetes 2003, 52, 252–259. [Google Scholar] [CrossRef]
- Marzullo, P.; Verti, B.; Savia, G.; Walker, G.E.; Guzzaloni, G.; Tagliaferri, M.; Di Blasio, A.; Liuzzi, A. The relationship between active ghrelin levels and human obesity involves alterations in resting energy expenditure. J. Clin. Endocrinol. Metab. 2004, 89, 936–939. [Google Scholar] [CrossRef] [PubMed]
- Scarpace, P.J.; Zhang, Y. Leptin resistance: A prediposing factor for diet-induced obesity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R493–R500. [Google Scholar] [CrossRef] [PubMed]
- Muscelli, E.; Mari, A.; Casolaro, A.; Camastra, S.; Seghieri, G.; Gastaldelli, A.; Holst, J.J.; Ferrannini, E. Separate impact of obesity and glucose tolerance on the incretin effect in normal subjects and type 2 diabetic patients. Diabetes 2008, 57, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Madsbad, S. The role of glucagon-like peptide-1 impairment in obesity and potential therapeutic implications. Diabetes Obes. Metab. 2014, 16, 9–21. [Google Scholar] [CrossRef]
- Van Bloemendaal, L.; Veltman, D.J.; Ten Kulve, J.S.; Groot, P.F.C.; Ruhé, H.G.; Barkhof, F.; Sloan, J.H.; Diamant, M.; Ijzerman, R.G. Brain reward-system activation in response to anticipation and consumption of palatable food is altered by glucagon-like peptide-1 receptor activation in humans. Diabetes, Obes. Metab. 2015, 17, 878–886. [Google Scholar] [CrossRef]
- Van Bloemendaal, L.; IJzerman, R.G.; ten Kulve, J.S.; Barkhof, F.; Konrad, R.J.; Drent, M.L.; Veltman, D.J.; Diamant, M. GLP-1 receptor activation modulates appetite- and reward-related brain areas in humans. Diabetes 2014, 63, 4186–4196. [Google Scholar] [CrossRef]
- Ten Kulve, J.S.; Veltman, D.J.; Van Bloemendaal, L.; Groot, P.F.C.; Ruhé, H.G.; Barkhof, F.; Diamant, M.; Ijzerman, R.G. Endogenous GLP1 and GLP1 analogue alter CNS responses to palatable food consumption. J. Endocrinol. 2016, 229, 1–12. [Google Scholar] [CrossRef]
- Farr, O.M.; Upadhyay, J.; Rutagengwa, C.; DiPrisco, B.; Ranta, Z.; Adra, A.; Bapatla, N.; Douglas, V.P.; Douglas, K.A.A.; Nolen-Doerr, E.; et al. Longer-term liraglutide administration at the highest dose approved for obesity increases reward-related orbitofrontal cortex activation in response to food cues: Implications for plateauing weight loss in response to anti-obesity therapies. Diabetes, Obes. Metab. 2019, 21, 2459–2464. [Google Scholar] [CrossRef]
- Vegiopoulos, A.; Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 2007, 275, 43–61. [Google Scholar] [CrossRef]
- Rose, A.J.; Vegiopoulos, A.; Herzig, S. Role of glucocorticoids and the glucocorticoid receptor in metabolism: Insights from genetic manipulations. J. Steroid Biochem. Mol. Biol. 2010, 122, 10–20. [Google Scholar] [CrossRef]
- Sarkar, S.; Fekete, C.; Légrádi, G.; Lechan, R.M. Glucagon like peptide-1 (7-36) amide (GLP-1) nerve terminals densely innervate corticotropin-releasing hormone neurons in the hypothalamic paraventricular nucleus. Brain Res. 2003, 985, 163–168. [Google Scholar] [CrossRef]
- Arase, K.; York, D.A.; Shimizu, H.; Shargill, N.; Bray, G.A. Effects of corticotropin-releasing factor on food intake and brown adipose tissue thermogenesis in rats. Am. J. Physiol. Endocrinol. Metab. 1988, 255, E255–E259. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Compound | Effect | Channel/ Current | Preparation | Dose | Tissue | Model |
|---|---|---|---|---|---|---|
| Ex-4 | activation | INav | in vitro | 1 µM | HPN | mouse |
| Ex-4 | activation | nonselective cationic | in vitro | 1 µM | HPN | mouse |
| Ex-4 | non effect | IA | in vitro | 1 µM | HPN | mouse |
| GLP-1 and analogs | ¿blockade? | K2P (TREK) 1 | ||||
| Ex-4 | depolarizing | in vitro | 1 µM | HPN | mouse | |
| Ex-4 | reduction | ICav | in vitro | 1 µM | HPN | mouse |
| Ex-4 | reduction | IAHP | 1 µM | HPN | mouse |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diz-Chaves, Y.; Herrera-Pérez, S.; González-Matías, L.C.; Lamas, J.A.; Mallo, F. Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress. Nutrients 2020, 12, 3304. https://doi.org/10.3390/nu12113304
Diz-Chaves Y, Herrera-Pérez S, González-Matías LC, Lamas JA, Mallo F. Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress. Nutrients. 2020; 12(11):3304. https://doi.org/10.3390/nu12113304
Chicago/Turabian StyleDiz-Chaves, Yolanda, Salvador Herrera-Pérez, Lucas C. González-Matías, José Antonio Lamas, and Federico Mallo. 2020. "Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress" Nutrients 12, no. 11: 3304. https://doi.org/10.3390/nu12113304
APA StyleDiz-Chaves, Y., Herrera-Pérez, S., González-Matías, L. C., Lamas, J. A., & Mallo, F. (2020). Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress. Nutrients, 12(11), 3304. https://doi.org/10.3390/nu12113304