The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota



 ,
,
Abstract
:1. Introduction
Intestinal Microflora
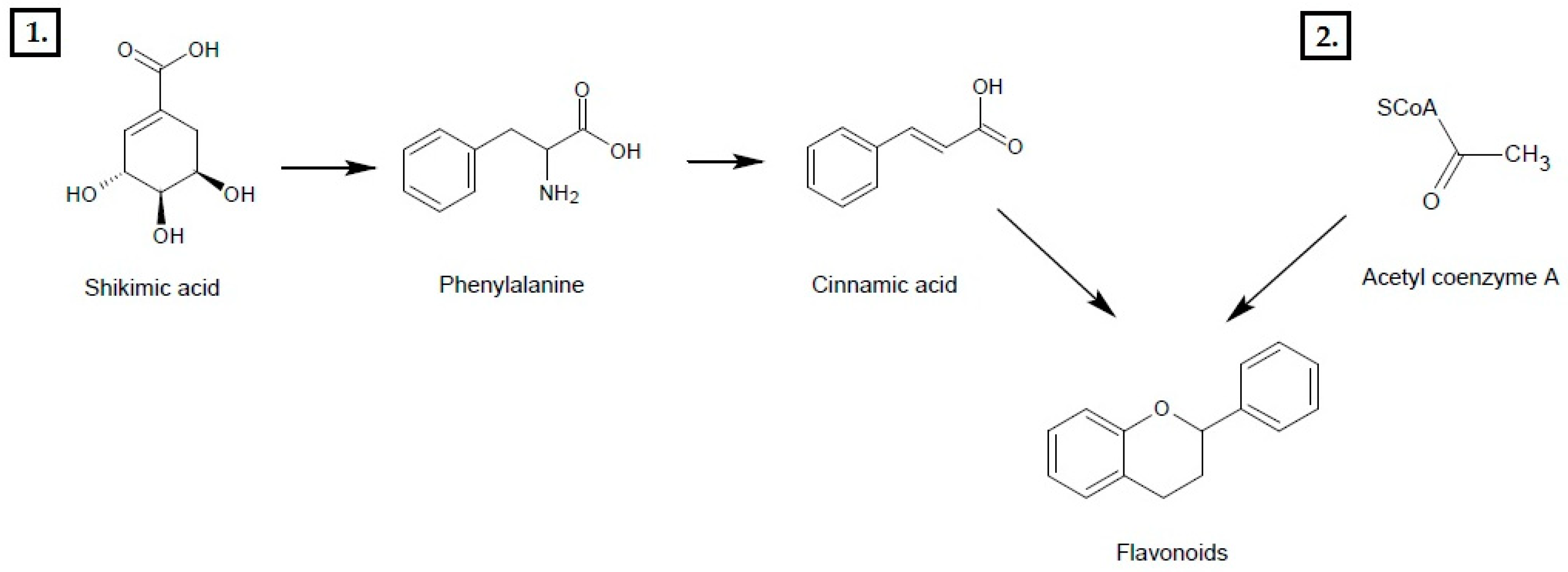
2. Phenol Compounds
3. The Influence of Polyphenol Compounds Contained in Tea on Intestinal Microflora.
4. The Influence of Polyphenols Present in Plant Extracts on Microbiota
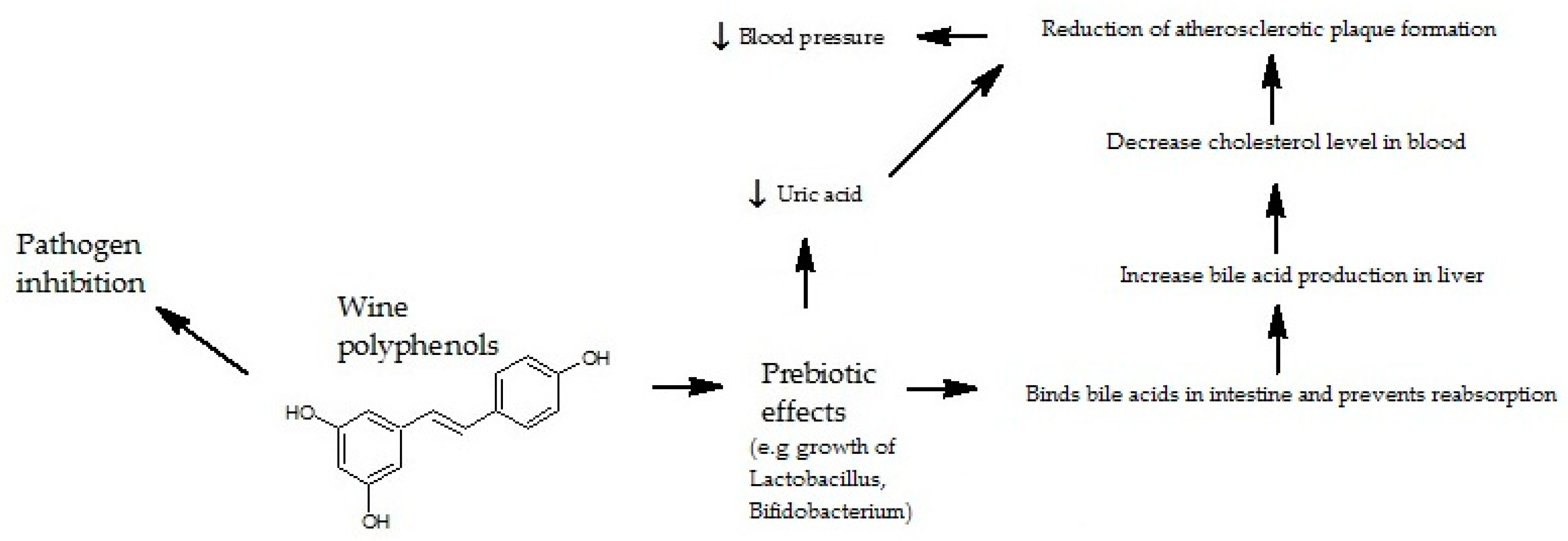
5. The Effect of Polyphenols Contained in Wine on Intestinal Microflora
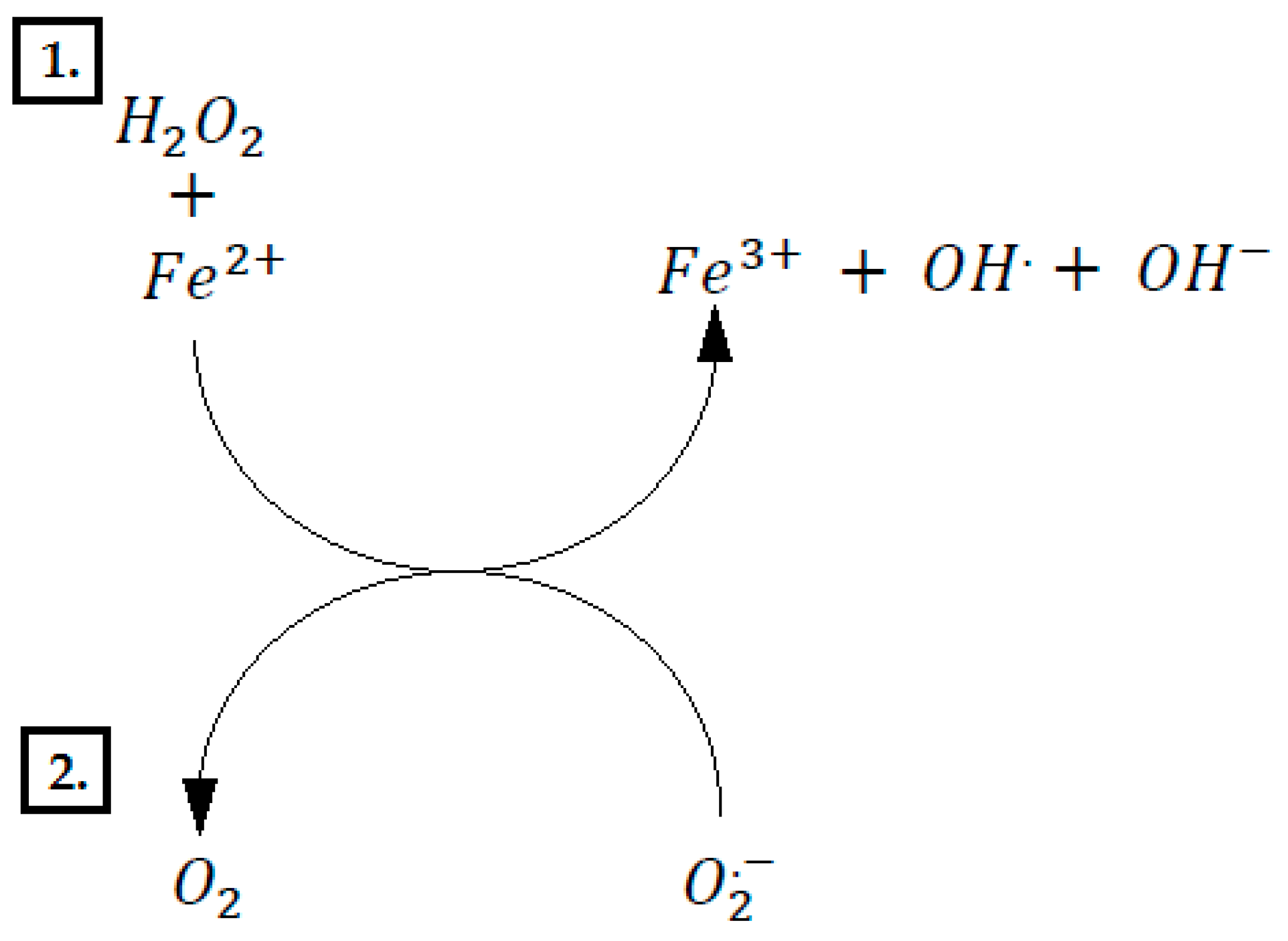
6. Polyphenols’ Negative Impact on Microflora of Intestines

{kind=link}
{kind=link}
{kind=link}
| Examined Polyphenols | Dose | Duration | Type of Experiment | Subjects | Diet | Diseases | Medications Taken | Stimulation of Bacterial Growth | Inhibition of Bacterial Growth | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| (+) – catechin (-) - epicatechin | 150 mg/1000 mg | 48 h | In vitro | Escherichia coli Bifidobacterium spp. Group Clostridium coccoides- Eubacterium rectale | Clostridium histolyticum | [38] | ||||
| Tea catechins | 300 mg | 6 weeks | In vivo [Open label pilot] | 31 females and 4 males from 66 to 98 years of age | 38.7 g proteins, 21.1 g lipids, 191 g carbohydrates, minerals, and vitamins. | Hypertension, cerebral stroke, senile dementia | None antibiotics | Bifidobacterium Lactobacillus | [46] | |
| (+) – epicatechin (-) – catechin 3-O-methylgallic acid Gallic acid Caffeic acid Phloretic acid 3-Phenylpropionic acid 4-Hydroxyphenylacetic acid | 1 mg/mL | 24 h | In vitro | Clostridium spp., Bifidobacterium spp., Lactobacillus spp. | Clostridium perfringens, Clostridium difficile, Escherichia coli, Salmonella, Staphylococcus spp.. | [47] | ||||
| Extract from grapes’ seeds containing proanthocyanidins | 0,19 g 0,38 g | 2 weeks | In vivo [Open label pilot] | 9 healthy adults from 37 to 42 years of age 8 elderly inpatients from 67 to 98 years of age | None red wine, green tea, products prepared using lactic bacteria 62,5±4,7 g/d proteins, 38,0 ± 4,7 g/d lipids, 212,0 ± 15,0 g/d carbohydrate 9,4 ± 1,3 g/d sodium, 803,9 ± 105,8 mg/d moisture, 568,1 ± 89,9 mg/d calcium | Cerebral stroke, bone fracture, senile dementia, articular rheumatism | None antibiotics None antibiotics | Bifidobacterium spp. | Enterobacteriaceae | [48] |
| Cocoa flavonoids | 23 mg 494 mg | 4 weeks | In vivo (Randomized, double-blind, placebo- controlled) | 22 healthy volunteers (12 male and 10 female - premenopausal ) from 18 to 50 years of age | The patients did not change their dietary habits | None antibiotics | Bifidobacterium spp. Lactobacillus spp. | Clostridium spp. | [50] | |
| Polyphenols in wine | 272 mL/d | 4 weeks | In vivo (Randomized, crossover, controlled, intervention) | 10 healthy adult men from 45–50 years of age | The patients did not change their dietary habits None alcohol and red wine | None antibiotics and supplements | Enterococcus, Prevotella, Bacteroides, Bifidobacterium, Bacteroides uniformis, Eggerthella lenta, Blautia coccoides–Eubacterium rectale group | [52] | ||
| Polyphenols in wine | 272 mL/d | 30 days | In vivo (Randomized, crossover, controlled intervention) | 20 adult men (10 obese and 10 healthy) from 45 to 50 years old: | The American Heart Association dietary guidelines | None antibiotics, prebiotic, probiotics, symbiotics, vitamin supplements and any medical treatment influencing intestinal microbiota | Bifidobacterium spp. Lactobacillus spp. | Enterobacter cloacae Escherichia coli | [56] | |
| Resveratrol Epigallocatechin-3-gallate | 80 mg/d 282 mg/d | 12 week | In vivo (Randomized, double-blind, placebo- controlled) | 18 males and 19 females from 20 to 50 years of age | < 600 mg caffeine, < 3 cups green tea, < 20 g alcohol | Overweight and obese | None antibiotics, medication, supplements | Bacteroidetes | [57] | |
| Anthocyanins Gallic acid | 24 h | In vitro | Bifidobacterium spp. Lactobacillus spp. | Clostridium histolyticum | [64] | |||||
| Quercetin | 12.5, 25, 50, 75μg/mL | 24 h | In vitro | Bifidobacterium catenulatum, Enterococcus caccae | [73] | |||||
| Naringenin, Naringin, Hesperetin, Hesperidin, Rutin, Quercetin Catechin | 20, 100, 250 μg/mL* * [for quercetin 4, 20, 50 μg/mL] | 24 h | In vitro | Lactic acid bacteria | [74] | |||||
| Grapes’ polyphenols | 0.25, 0.5, 1 mg/mL | 24 h | In vitro | Bifidobacterium spp. Lactobacillus spp. | [75] | |||||
| Extracts from: blackcurrant blueberry cranberry cloudberry lingonberry raspberry berry of sea-buckthorn strawberry Polyphenols: Apigenin, Caffeic acid, (+) - Catechin, Chlorogenic acid, Coumarin-3-carboxylic acid, Cyanidin chloride, Delphinidin chloride, Ferulic acid, Isoquercetin, Kaempferol, Cyanidin-3-O-glucoside, Luteolin, Myricetin, Pelargonidin chloride, Quercetin dihydrate, Rutin trihydrate, Trans-cinnamic acid | 0,5, 1, 5 mg/mL | 24 h | In vitro | Lactobacillus spp. | [76] |
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bibbo, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar] [PubMed]
- Nicholson, J.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, M.; Ambalam, P.; Doble, M. Probiotics and Bioactive Carbohydrates in Colon Cancer Management. In Probiotics and Bioactive Carbohydrates in Colon Cancer Management; Springer: New Delhi, India, 2016. [Google Scholar]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Littman, D.R.; MacPherson, A.J. Interactions Between the Microbiota and the Immune System. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, P. Molecular mechanistic pathway of colorectal carcinogenesis associated with intestinal microbiota. Anaerobe 2018, 49, 63–70. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [Green Version]
- Modi, S.R.; Collins, J.J.; Relman, D.A. Antibiotics and the gut microbiota. J. Clin. Investig. 2014, 124, 4212–4218. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.A.; Prudencio, A.; Aichelmann-Reidy, M.E.; Woodward, K.; Uhrich, K. Non-Steroidal Anti-inflammatory Drug (NSAID)-Derived Poly(anhydrideesters) in Bone and Periodontal Regeneration. Curr. Drug Deliv. 2007, 4, 233–239. [Google Scholar] [CrossRef]
- Hirata, S.; Kunisawa, J. Gut microbiome, metabolome, and allergic diseases. Allergol Int. 2017, 66, 523–528. [Google Scholar] [CrossRef]
- Cenit, M.C.; Matzaraki, V.; Tigchelaar, E.; Zhernakova, A. Rapidly expanding knowledge on the role of the gut microbiome in health and disease. Biochim. et Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 1981–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oriach, C.S.; Robertson, R.C.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Food for thought: The role of nutrition in the microbiota-gut–brain axis. Clin. Nutr. Exp. 2016, 6, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.A.; Ramos-Lopez, O.; Riezu-Boj, J.-I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, and Obesity: Links with Host Genetics and Epigenetics and Potential Applications. Adv. Nutr. 2019, 10, S17–S30. [Google Scholar]
- Ozdal, T.; Sela, D.; Ulrih, N.P.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Colomer, R.; Sarrats, A.; Lupu, R.; Puig, T. Natural Polyphenols and their Synthetic Analogs as Emerging Anticancer Agents. Curr. Drug Targets 2017, 18, 147–159. [Google Scholar] [CrossRef]
- Wang, T.-Y.; Li, Q.; Bi, K. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytother. Res. 2018, 33, 13–40. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Liang, F.; Liu, K.; Qaiser, S.; Pan, S.; Xu, X. Structure characteristics for intestinal uptake of flavonoids in Caco-2 cells. Food Res. Int. 2017, 105, 353–360. [Google Scholar] [CrossRef]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteiza, P.I.; Fraga, C.; Mills, D.A.; Taft, D. Flavonoids and the gastrointestinal tract: Local and systemic effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Fruit Polyphenols: A Review of Anti-inflammatory Effects in Humans. Crit. Rev. Food Sci. Nutr. 2015, 56, 419–444. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Neveu, V.; Voß, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the Phenol-Explorer database. Eur. J. Clin. Nutr. 2010, 64, S112–S120. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Leopoldini, M.; Nino, R.; Toscano, M. The molecular basis of working mechanism of natural polyphenolic antioxidants. Food Chem. 2011, 125, 288–306. [Google Scholar] [CrossRef]
- Wang, T.; Jonsdottir, R.; Olafsdottir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Brillas, E.; Sirés, I.; Oturan, M.A. Electro-Fenton Process and Related Electrochemical Technologies Based on Fenton’s Reaction Chemistry. Chem. Rev. 2009, 109, 6570–6631. [Google Scholar] [CrossRef]
- Trujillo, D.; Font, X.; Sanchez, A. Use of Fenton reaction for the treatment of leachate from composting of different wastes. J. Hazard. Mater. 2006, 138, 201–204. [Google Scholar] [CrossRef] [Green Version]
- Ajiboye, T.O.; Aliyu, M.; Isiaka, I.; Haliru, F.; Ibitoye, O.; Uwazie, J.; Muritala, H.; Bello, S.; Yusuf, I.; Mohammed, A. Contribution of reactive oxygen species to (+)-catechin-mediated bacterial lethality. Chem. Interactions 2016, 258, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, N.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Knaze, V.; Zamora-Ros, R. Polyphenols: Dietary assessment and role in the prevention of cancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Lecci, R.M.; Logrieco, A.; Leone, A. Pro-oxidative action of polyphenols as action mechanism for their pro-apoptotic activity. Anti-Cancer Agents Med. Chem. 2014, 14, 1363–1375. [Google Scholar] [CrossRef]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2007, 99, 782–792. [Google Scholar] [CrossRef] [Green Version]
- Shortt, C.; Hasselwander, O.; Meynier, A.; Nauta, A.; Fernández, E.N.; Putz, P.; Rowland, I.; Swann, J.; Türk, J.; Vermeiren, J.; et al. Systematic review of the effects of the intestinal microbiota on selected nutrients and non-nutrients. Eur. J. Nutr. 2017, 57, 25–49. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M. Short-chain fatty acids. In Microbial Inhabitants of Humans; Cambridge University Press: Cambridge, UK, 2005; pp. 388–390. [Google Scholar]
- Redondo, N.; Gheorghe, A.; Díaz, L.E.; Villavisencio, B.; Marcos, A.; Nova, E. Associations of Probiotic Fermented Milk (PFM) and Yogurt Consumption with Bifidobacterium and Lactobacillus Components of the Gut Microbiota in Healthy Adults. Nutrients 2019, 11, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, N.; Tian, Y.; Wu, Y.; Ma, X. Contributions of the Interaction between Dietary Protein and Gut Microbiota to Intestinal Health. Curr. Protein Pept. Sci. 2017, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Lubudisz, Z. Karcynogenna aktywność mikroorganizmów jelitowych. Żywność Nauka Technologia Jakość 2008, 6, 25–39. [Google Scholar]
- Goto, K.; Kanaya, S.; Nishikawa, T.; Hara, H.; Terada, A.; Ishigami, T.; Hara, Y. The effects of tea catechins on fecal flora of elderly residents in long-term care facilities. J. Nutr. Sci. Vitaminol. (Tokyo). 1999, 45, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.-K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Yamakoshi, J.; Tokutake, S.; Kikuchi, M. Effect of Proanthocyanidin-Rich Extract from Grape Seeds on Human Fecal Flora and Fecal Odor. Microb Ecol Health Dis. 2001, 13, 25–31. [Google Scholar]
- Lee, E.-S.; Song, E.-J.; Nam, Y.-D.; Lee, S.-Y. Probiotics in human health and disease: From nutribiotics to pharmabiotics. J. Microbiol. 2018, 56, 773–782. [Google Scholar] [CrossRef]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2010, 93, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Requena, T.; Monagas, M.; Pozo-Bayon, M.A.; Martin-Alvarez, P.; Bartolomé, B.; Del Campo, R.; Ávila, M.; Martínez-Cuesta, M.C.; Peláez, C.; Moreno-Arribas, M. Perspectives of the potential implications of wine polyphenols on human oral and gut microbiota. Trends Food Sci. Technol. 2010, 21, 332–344. [Google Scholar] [CrossRef]
- Ortuño, M.I.Q.; Boto-Ordóñez, M.; Murri, M.; Gómez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Diaz, F.C.; Andres-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef]
- Melanie, H.; Susilowati, A.; Maryati, Y. Fermented inulin hydrolysate by Bifidobacterium breve as cholesterol binder in functional food application. Int. Symp. Appl. Chem. 2016, 1803, 259–266. [Google Scholar]
- Guo, Z.; Zhang, J.; Wang, Z.; Ang, K.Y.; Huang, S.; Hou, Q.; Su, X.; Qiao, J.; Zheng, Y.; Wang, L.; et al. Intestinal Microbiota Distinguish Gout Patients from Healthy Humans. Sci. Rep. 2016, 6, 20602. [Google Scholar] [CrossRef] [Green Version]
- Melander, O.; Newton-Cheh, C.; Almgren, P.; Hedblad, B.; Berglund, G.; Engström, G.; Persson, M.; Smith, J.G.; Magnusson, M.; Christensson, A.; et al. Novel and Conventional Biomarkers for Prediction of Incident Cardiovascular Events in the Community. JAMA. 2009, 302, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Indias, I.; Sanchez-Alcoholado, L.; Pérez-Martínez, P.; Andres-Lacueva, C.; Cardona, F.; Tinahones, F.J.; Ortuño, M.I.Q. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Most, J.; Penders, J.; Lucchesi, M.; Goossens, G.H.; E Blaak, E. Gut microbiota composition in relation to the metabolic response to 12-week combined polyphenol supplementation in overweight men and women. Eur. J. Clin. Nutr. 2017, 71, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Halder, C.V.; Faria, A.V.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, L.; Wang, Y.; Dorfman, R.G.; Tang, D.; Xu, L.; Pan, Y.; Zhou, Q.; Li, Y.; Yin, Y.; et al. Faecalibacterium prausnitzii produces butyrate to decrease c-Myc-related metabolism and Th17 differentiation by inhibiting histone deacetylase 3. Int. Immunol. 2019, 31, 499–514. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Enrich-Capó, N.; Aldeguer, X.; Sabat-Mir, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Alterations in the Abundance and Co-occurrence of Akkermansia muciniphila and Faecalibacterium prausnitzii in the Colonic Mucosa of Inflammatory Bowel Disease Subjects. Front. Microbiol. 2018, 8, 281. [Google Scholar] [CrossRef]
- Biedermann, L.; Mwinyi, J.; Scharl, M.; Frei, P.; Zeitz, J.; Kullak- Ublick, G.A.; Vavricka, S.R.; Fried, M.; Weber, A.; Humpf, H.U.; et al. Bilberry ingestion improves disease activity in mild to moderate ulcerative colitis – An open pilot study. J. Crohns Colitis. 2013, 7, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Dryden, G.; Lam, A.; Beatty, K.; Qazzaz, H.H.; McClain, C.J. A Pilot Study to Evaluate the Safety and Efficacy of an Oral Dose of (−)-Epigallocatechin-3-Gallate–Rich Polyphenon E in Patients With Mild to Moderate Ulcerative Colitis. Inflamm. Bowel Dis. 2013, 19, 1904–1912. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Concha, M.J.O.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.E.; Gibson, G.R.; De Pascual-Teresa, S. Metabolism of Anthocyanins by Human Gut Microflora and Their Influence on Gut Bacterial Growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Eslami, M.; Yousefi, B.; Kokhaei, P.; Hemati, M.; Nejad, Z.R.; Arabkari, V.; Namdar, A. Importance of probiotics in the prevention and treatment of colorectal cancer. J. Cell. Physiol. 2019, 234, 17127–17143. [Google Scholar] [CrossRef] [PubMed]
- Hervert-Hernández, D.; Goñi, I. Dietary Polyphenols and Human Gut Microbiota: A Review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Hartmann-Schmidlin, S.; Kähkönen, M.; Heinonen, M.; Maatta-Riihinen, K.; Oksman-Caldentey, K.-M. Berry phenolics selectively inhibit the growth of intestinal pathogens. J. Appl. Microbiol. 2005, 98, 991–1000. [Google Scholar] [CrossRef]
- Kemperman, R.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analysing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiol. 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [Green Version]
- Anna, V.S.; Biondo, E.; Kolchinski, E.M.; Da Silva, L.F.S.; Correa, A.P.F.; Bach, E.; Brandelli, A. Total Polyphenols, Antioxidant, Antimicrobial and Allelopathic Activities of Spend Coffee Ground Aqueous Extract. Waste Biomass Valorization 2016, 8, 439–442. [Google Scholar]
- Coppo, E.; Marchese, A. Antibacterial activity of polyphenols. Curr. Pharm. Biotechnol. 2014, 15, 380–390. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum Sensing and Phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef] [Green Version]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Ortuño, M.I.Q. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Firrman, J.; Liu, L.; Zhang, L.; Argoty, G.A.; Wang, M.; Tomasula, P.; Kobori, M.; Pontious, S.; Xiao, W. The effect of quercetin on genetic expression of the commensal gut microbes Bifidobacterium catenulatum, Enterococcus caccae and Ruminococcus gauvreauii. Anaerobe 2016, 42, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2012, 63, 497–503. [Google Scholar]
- Tabasco, R.; Sánchez-Patán, F.; Monagas, M.; Bartolomé, B.; Moreno-Arribas, M.V.; Peláez, C.; Requena, T. Effect of grape polyphenols on lactic acid bacteria and bifidobacteria growth: Resistance and metabolism. Food Microbiol. 2011, 28, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Nohynek, L.; Meier, C.; Kahkonen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.-M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef] [PubMed]



| Class | Subclass | Examples of Compounds | Source |
|---|---|---|---|
| Phenolic acids | Hydroxycinnamic acids | Curcumin Caffeic acid Ferulic acid | Fruit and cereals |
| Hydroxybenzoic acids | Gallic acid Protocatechuic acid Vanillic acid | Onion, raspberry, blackberry, strawberry | |
| Favonoids | Flavonols | Kaempferol, Quercetin, Myricetin | Onions, tea, lettuce, broccoli, apples |
Flavanones | Naringenin, Hesperetin | Oranges, grapefruits | |
Flavanols | Gallocatechin Catechins | Tea, red wine, chocolate | |
Isoflavones | Genistein, Glycitein, Daidzein | Soybeans, legumes | |
Anthocyanins | Pelargonidin, Delphinidin, Malvidin | Blackcurrant, strawberries, red wine, chokeberry | |
Flavones | Apigenin, Luteolin, | Parsley, celery, red pepper, lemon, thyme | |
| Stilbenes | Resvertrol | Red wine | |
| Lignans | Pinoresinol, Lariciresinol, Secoisolariciresinol, Sesamin | Flax seed, sesame seed, red wine |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiciński, M.; Gębalski, J.; Mazurek, E.; Podhorecka, M.; Śniegocki, M.; Szychta, P.; Sawicka, E.; Malinowski, B. The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota. Nutrients 2020, 12, 350. https://doi.org/10.3390/nu12020350
Wiciński M, Gębalski J, Mazurek E, Podhorecka M, Śniegocki M, Szychta P, Sawicka E, Malinowski B. The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota. Nutrients. 2020; 12(2):350. https://doi.org/10.3390/nu12020350
Chicago/Turabian StyleWiciński, Michał, Jakub Gębalski, Ewelina Mazurek, Marta Podhorecka, Maciej Śniegocki, Paweł Szychta, Ewelina Sawicka, and Bartosz Malinowski. 2020. "The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota" Nutrients 12, no. 2: 350. https://doi.org/10.3390/nu12020350
APA StyleWiciński, M., Gębalski, J., Mazurek, E., Podhorecka, M., Śniegocki, M., Szychta, P., Sawicka, E., & Malinowski, B. (2020). The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota. Nutrients, 12(2), 350. https://doi.org/10.3390/nu12020350