Vitamin D Effects on the Immune System from Periconception through Pregnancy


Abstract
:1. Introduction
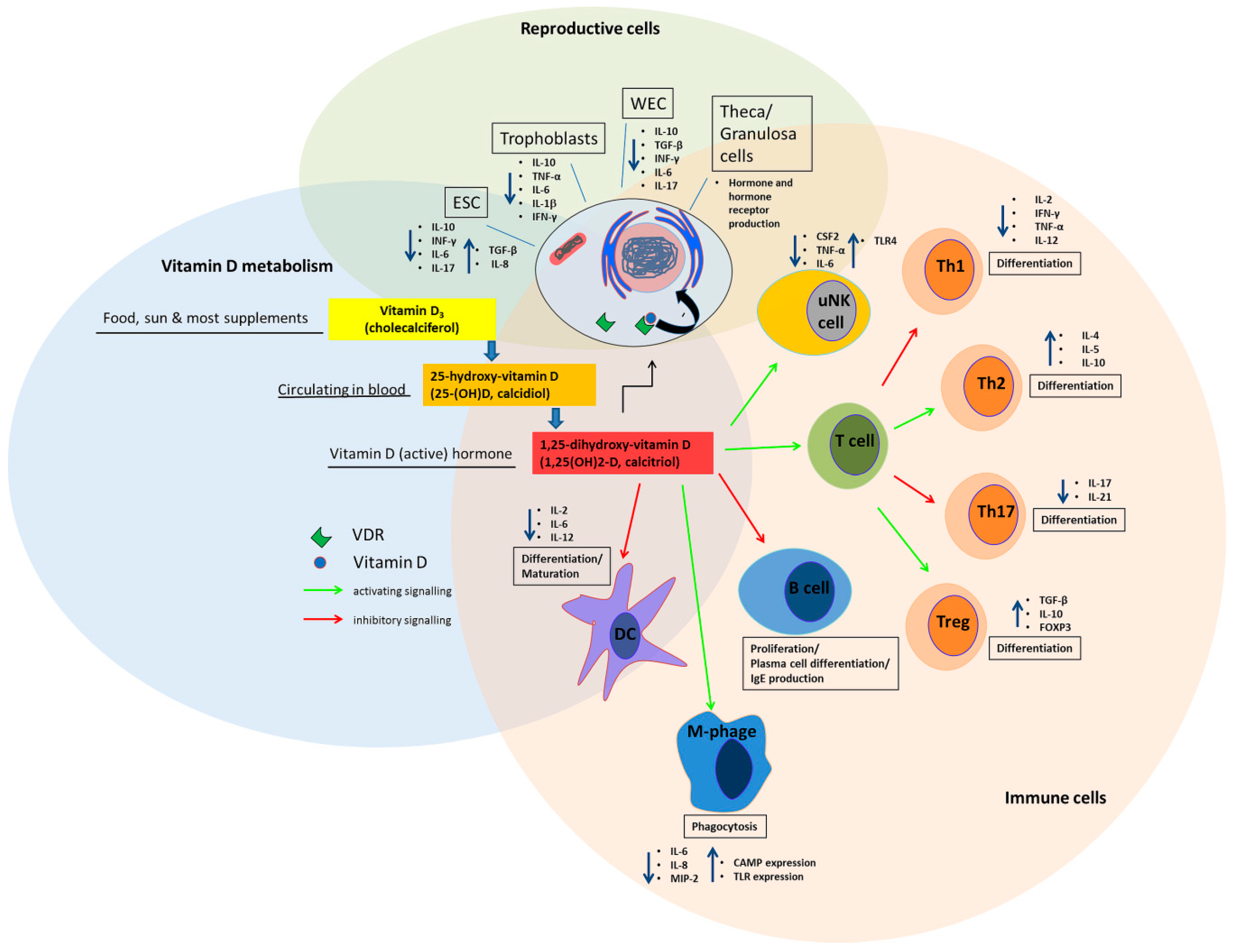
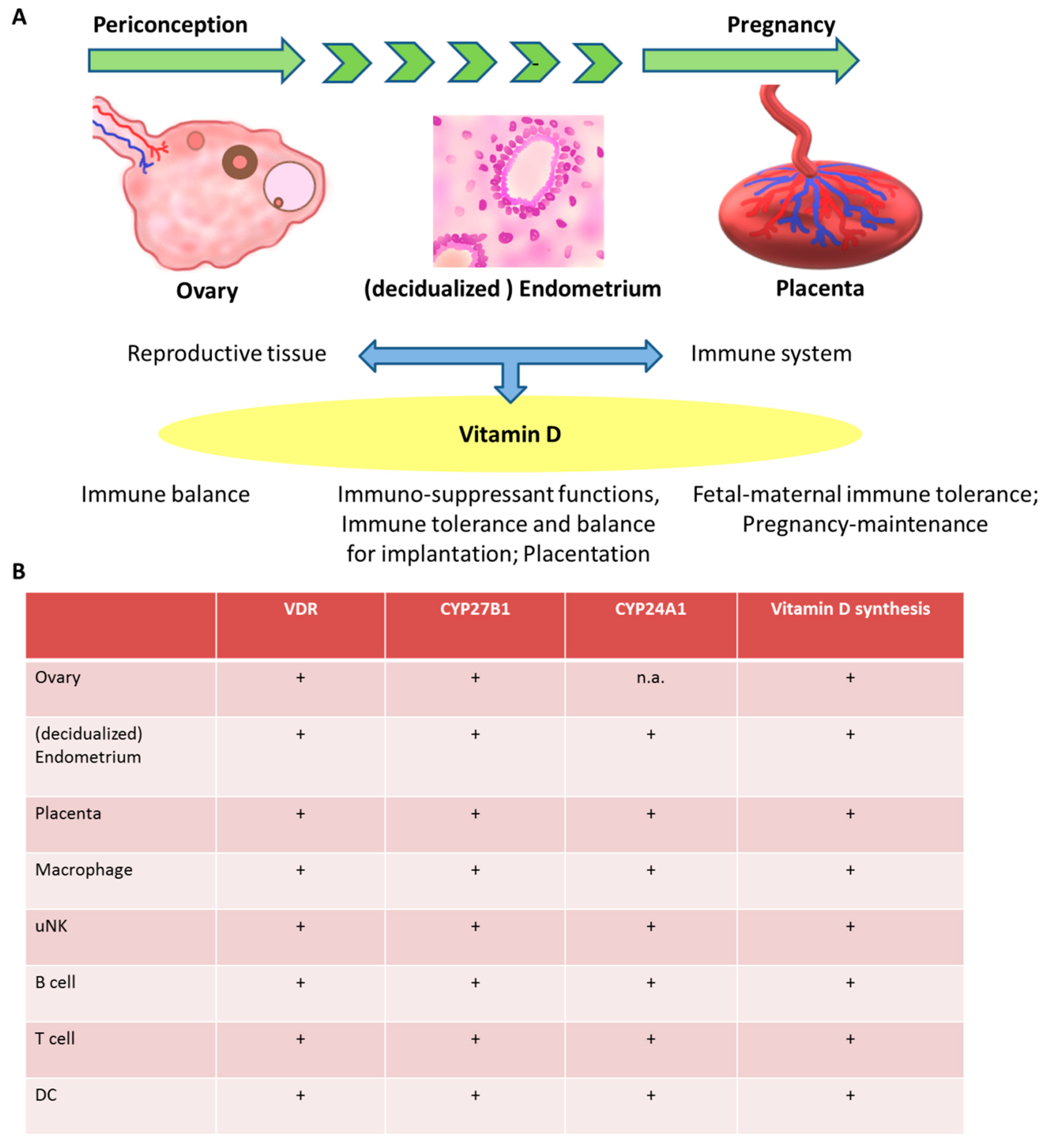
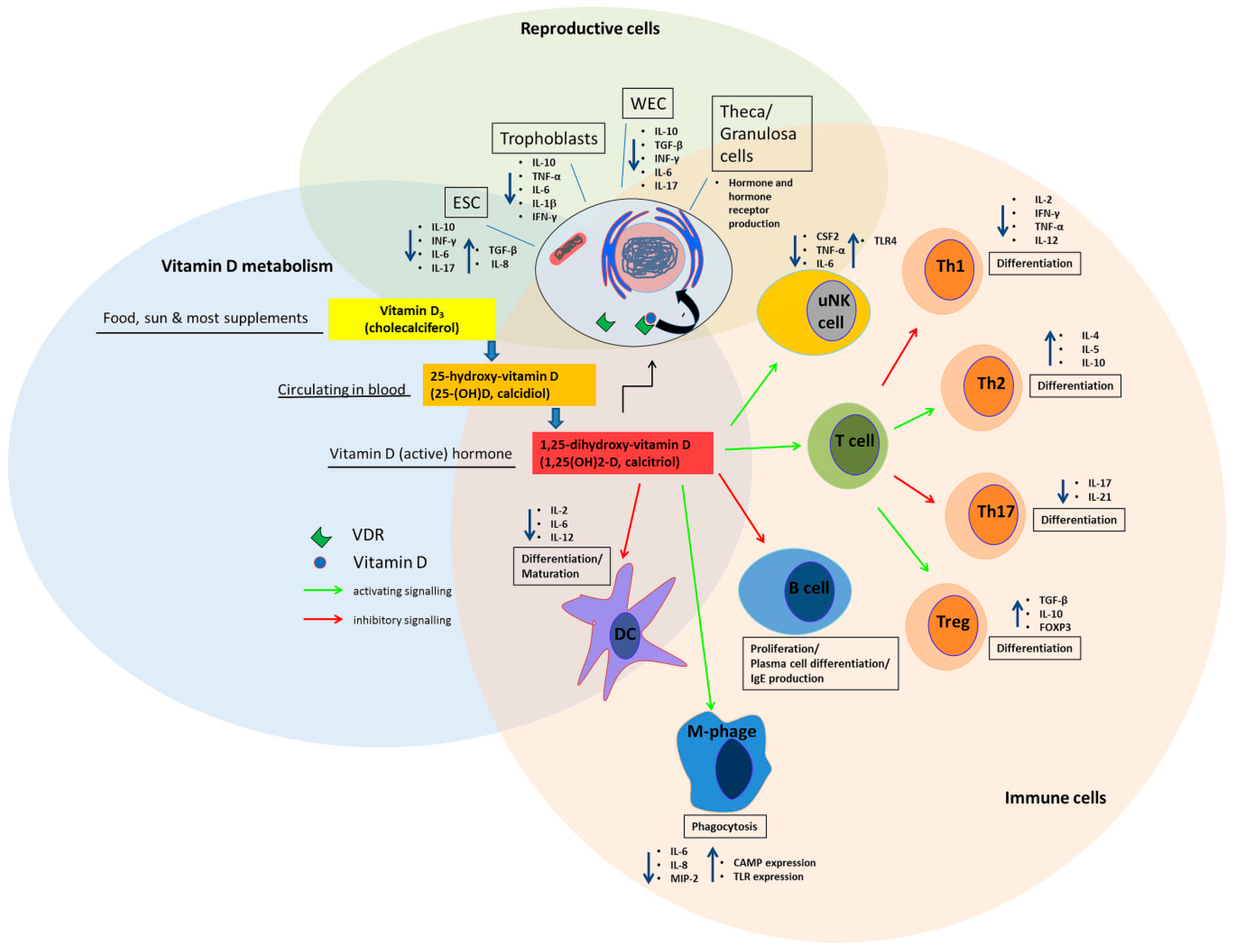
2. Vitamin D metabolism and Synthesis in Reproductive Tissue
3. The Effect of Vitamin D on the Immune System in the Periconception Period and in Pregnancy
4. Vitamin D, the Immune System and Adverse Reproductive Outcomes
4.1. Recurrent Pregnancy Loss
4.2. Recurrent Implantation Failure
4.3. ART Outcomes
5. Pregnancy Complications
5.1. Preeclampsia
5.2. Preterm Birth
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Schröder-Heurich, B.; Von Versen-Höynck, F. Famine, Starvation and Nuttrient Deprivation: From Biology to Policy. In Vitamin D Deficiency and Fertility-An Overview; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Aleyasin, A.; Hosseini, M.A.; Mahdavi, A.; Safdarian, L.; Fallahi, P.; Mohajeri, M.R.; Abbasi, M.; Esfahani, F. Predictive value of the level of vitamin D in follicular fluid on the outcome of assisted reproductive technology. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 159, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D status: Measurement, interpretation, and clinical application. Ann. Epidemiol. 2009, 19, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keane, K.N.; Cruzat, V.F.; Calton, E.K.; Hart, P.H.; Soares, M.J.; Newsholme, P.; Yovich, J.L. Molecular actions of vitamin D in reproductive cell biology. Reproduction 2017, 153, R29–R42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranow, C. Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.L.; Muyayalo, K.P.; Zhang, Y.H.; Hu, X.H.; Liao, A.H. Immunological function of vitamin D during human pregnancy. Am. J. Reprod. Immunol. 2017, 78. [Google Scholar] [CrossRef]
- Schröder-Heurich, B.; von Hardenberg, S.; Brodowski, L.; Kipke, B.; Meyer, N.; Borns, K.; von Kaisenberg, C.S.; Brinkmann, H.; Claus, P.; von Versen-Höynck, F. Vitamin D improves endothelial barrier integrity and counteracts inflammatory effects on endothelial progenitor cells. FASEB J. 2019, 33, 9142–9153. [Google Scholar] [CrossRef]
- Wei, S.Q. Vitamin D and pregnancy outcomes. Curr. Opin. Obstet. Gynecol. 2014, 26, 438–447. [Google Scholar] [CrossRef]
- Wöbke, T.K.; Sorg, B.L.; Steinhilber, D. Vitamin D in inflammatory diseases. Front. Physiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Bhalla, A.K.; Amento, E.P.; Serog, B.; Glimcher, L.H. 1,25-Dihydroxyvitamin D3 inhibits antigen-induced T cell activation. J. Immunol. 1984, 133, 1748–1754. [Google Scholar]
- Lemire, J.M.; Adams, J.S.; Sakai, R.; Jordan, S.C. 1 alpha,25-dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J. Clin. Investig. 1984, 74, 657–661. [Google Scholar] [CrossRef] [Green Version]
- Mattner, F.; Smiroldo, S.; Galbiati, F.; Muller, M.; Di Lucia, P.; Poliani, P.L.; Martino, G.; Panina-Bordignon, P.; Adorini, L. Inhibition of Th1 development and treatment of chronic-relapsing experimental allergic encephalomyelitis by a non-hypercalcemic analogue of 1,25-dihydroxyvitamin D(3). Eur. J. Immunol. 2000, 30, 498–508. [Google Scholar] [CrossRef]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.; O’Garra, A. 1alpha,25-Dihydroxyvitamin D3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; Chen, S.; Lipsky, P.E. Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J. Immunol. 2007, 179, 1634–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, C.; Sartory, N.A.; Zahn, N.; Radeke, H.H.; Stein, J.M. Immune modulatory treatment of trinitrobenzene sulfonic acid colitis with calcitriol is associated with a change of a T helper (Th) 1/Th17 to a Th2 and regulatory T cell profile. J. Pharmacol. Exp. Ther. 2008, 324, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Zhou, R.; Luger, D.; Zhu, W.; Silver, P.B.; Grajewski, R.S.; Su, S.B.; Chan, C.C.; Adorini, L.; Caspi, R.R. Calcitriol suppresses antiretinal autoimmunity through inhibitory effects on the Th17 effector response. J. Immunol. 2009, 182, 4624–4632. [Google Scholar] [CrossRef]
- Gregori, S.; Casorati, M.; Amuchastegui, S.; Smiroldo, S.; Davalli, A.M.; Adorini, L. Regulatory T cells induced by 1 alpha,25-dihydroxyvitamin D3 and mycophenolate mofetil treatment mediate transplantation tolerance. J. Immunol. 2001, 167, 1945–1953. [Google Scholar] [CrossRef] [Green Version]
- Barrat, F.J.; Cua, D.J.; Boonstra, A.; Richards, D.F.; Crain, C.; Savelkoul, H.F.; de Waal-Malefyt, R.; Coffman, R.L.; Hawrylowicz, C.M.; O’Garra, A. In vitro generation of interleukin 10-producing regulatory CD4(+) T cells is induced by immunosuppressive drugs and inhibited by T helper type 1 (Th1)- and Th2-inducing cytokines. J. Exp. Med. 2002, 195, 603–616. [Google Scholar] [CrossRef]
- Piemonti, L.; Monti, P.; Sironi, M.; Fraticelli, P.; Leone, B.E.; Dal Cin, E.; Allavena, P.; Di Carlo, V. Vitamin D3 affects differentiation, maturation, and function of human monocyte-derived dendritic cells. J. Immunol. 2000, 164, 4443–4451. [Google Scholar] [CrossRef] [Green Version]
- Harant, H.; Wolff, B.; Lindley, I.J. 1Alpha,25-dihydroxyvitamin D3 decreases DNA binding of nuclear factor-kappaB in human fibroblasts. FEBS Lett. 1998, 436, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Lahav, M.; Shany, S.; Tobvin, D.; Chaimovitz, C.; Douvdevani, A. Vitamin D decreases NFkappaB activity by increasing IkappaBalpha levels. Nephrol. Dial. Transpl. 2006, 21, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Vigano, P.; Lattuada, D.; Mangioni, S.; Ermellino, L.; Vignali, M.; Caporizzo, E.; Panina-Bordignon, P.; Besozzi, M.; Di Blasio, A.M. Cycling and early pregnant endometrium as a site of regulated expression of the vitamin D system. J. Mol. Endocrinol. 2006, 36, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.P.; Nierstenhoefer, M.; Yoo, S.W.; Penzias, A.S.; Tobiasch, E.; Usheva, A. The bile acid synthesis pathway is present and functional in the human ovary. PLoS ONE 2009, 4, e7333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thill, M.; Becker, S.; Fischer, D.; Cordes, T.; Hornemann, A.; Diedrich, K.; Salehin, D.; Friedrich, M. Expression of prostaglandin metabolising enzymes COX-2 and 15-PGDH and VDR in human granulosa cells. Anticancer Res. 2009, 29, 3611–3618. [Google Scholar] [PubMed]
- Shahbazi, M.; Jeddi-Tehrani, M.; Zareie, M.; Salek-Moghaddam, A.; Akhondi, M.M.; Bahmanpoor, M.; Sadeghi, M.R.; Zarnani, A.H. Expression profiling of vitamin D receptor in placenta, decidua and ovary of pregnant mice. Placenta 2011, 32, 657–664. [Google Scholar] [CrossRef]
- Lerchbaum, E.; Obermayer-Pietsch, B. Vitamin D and fertility: A systematic review. Eur. J. Endocrinol. 2012, 166, 765–778. [Google Scholar] [CrossRef]
- Chu, J.; Gallos, I.; Tobias, A.; Tan, B.; Eapen, A.; Coomarasamy, A. Vitamin D and assisted reproductive treatment outcome: A systematic review and meta-analysis. Hum. Reprod. 2018, 33, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Zelenko, Z.; Aghajanova, L.; Irwin, J.C.; Giudice, L.C. Nuclear receptor, coregulator signaling, and chromatin remodeling pathways suggest involvement of the epigenome in the steroid hormone response of endometrium and abnormalities in endometriosis. Reprod. Sci. 2012, 19, 152–162. [Google Scholar] [CrossRef]
- Bergada, L.; Pallares, J.; Maria Vittoria, A.; Cardus, A.; Santacana, M.; Valls, J.; Cao, G.; Fernandez, E.; Dolcet, X.; Dusso, A.S.; et al. Role of local bioactivation of vitamin D by CYP27A1 and CYP2R1 in the control of cell growth in normal endometrium and endometrial carcinoma. Lab. Investig. 2014, 94, 608–622. [Google Scholar] [CrossRef] [Green Version]
- Weisman, Y.; Harell, A.; Edelstein, S.; David, M.; Spirer, Z.; Golander, A. 1 alpha, 25-Dihydroxyvitamin D3 and 24,25-dihydroxyvitamin D3 in vitro synthesis by human decidua and placenta. Nature 1979, 281, 317–319. [Google Scholar] [CrossRef]
- Liu, N.Q.; Kaplan, A.T.; Lagishetty, V.; Ouyang, Y.B.; Ouyang, Y.; Simmons, C.F.; Equils, O.; Hewison, M. Vitamin D and the regulation of placental inflammation. J. Immunol. 2011, 186, 5968–5974. [Google Scholar] [CrossRef] [Green Version]
- Vijayendra Chary, A.; Hemalatha, R.; Seshacharyulu, M.; Vasudeva Murali, M.; Jayaprakash, D.; Dinesh Kumar, B. Vitamin D deficiency in pregnant women impairs regulatory T cell function. J. Steroid Biochem. Mol. Biol. 2015, 147, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Eapen, M.S.; Zosky, G.R. Vitamin D both facilitates and attenuates the cellular response to lipopolysaccharide. Sci. Rep. 2017, 7, 45172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puthuraya, S.; Karnati, S.; Kazzi, S.N.J.; Qureshi, F.; Jacques, S.M.; Thomas, R. Does vitamin D deficiency affect placental inflammation or infections among very low birth weight infants? J. Matern Fetal Neonatal Med. 2018, 31, 1906–1912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, H.; Wang, Y.; Zhang, C.; Tang, Z.; Li, H.; Huang, X.; Ouyang, F.; Huang, H.; Liu, Z. Severe vitamin D deficiency in the first trimester is associated with placental inflammation in high-risk singleton pregnancy. Clin. Nutr. 2019, 38, 1921–1926. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef] [Green Version]
- Evans, K.N.; Nguyen, L.; Chan, J.; Innes, B.A.; Bulmer, J.N.; Kilby, M.D.; Hewison, M. Effects of 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 on cytokine production by human decidual cells. Biol. Reprod. 2006, 75, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Kaplan, A.T.; Low, J.; Nguyen, L.; Liu, G.Y.; Equils, O.; Hewison, M. Vitamin D induces innate antibacterial responses in human trophoblasts via an intracrine pathway. Biol. Reprod. 2009, 80, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, Y.H.; Fu, L.; Yu, Z.; Xia, M.Z.; Hu, X.G.; Wang, H.; Xu, D.X. Vitamin D3 pretreatment protects against lipopolysaccharide-induced early embryo loss through its anti-inflammatory effects. Am. J. Reprod. Immunol. 2017, 77. [Google Scholar] [CrossRef]
- Yates, N.; Crew, R.C.; Wyrwoll, C.S. Vitamin D deficiency and impaired placental function: Potential regulation by glucocorticoids? Reproduction 2017, 153, R163–R171. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S.A. Immune regulation of conception and embryo implantation-all about quality control? J. Reprod. Immunol. 2010, 85, 51–57. [Google Scholar] [CrossRef]
- Miller, D.J. Review: The epic journey of sperm through the female reproductive tract. Animal 2018, 12, s110–s120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Pillai, V.V.; Weber, D.M.; Phinney, B.S.; Selvaraj, V. Profiling of proteins secreted in the bovine oviduct reveals diverse functions of this luminal microenvironment. PLoS ONE 2017, 12, e0188105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neira, J.A.; Tainturier, D.; Pena, M.A.; Martal, J. Effect of the association of IGF-I, IGF-II, bFGF, TGF-beta1, GM-CSF, and LIF on the development of bovine embryos produced in vitro. Theriogenology 2010, 73, 595–604. [Google Scholar] [CrossRef]
- Ptak, G.; Lopes, F.; Matsukawa, K.; Tischner, M.; Loi, P. Leukaemia inhibitory factor enhances sheep fertilization in vitro via an influence on the oocyte. Theriogenology 2006, 65, 1891–1899. [Google Scholar] [CrossRef]
- Robertson, S.A.; Moldenhauer, L.M. Immunological determinants of implantation success. Int. J. Dev. Biol. 2014, 58, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Arici, A.; Seli, E.; Zeyneloglu, H.B.; Senturk, L.M.; Oral, E.; Olive, D.L. Interleukin-8 induces proliferation of endometrial stromal cells: A potential autocrine growth factor. J. Clin. Endocrinol. Metab. 1998, 83, 1201–1205. [Google Scholar] [CrossRef]
- Cyprian, F.; Lefkou, E.; Varoudi, K.; Girardi, G. Immunomodulatory Effects of Vitamin D in Pregnancy and Beyond. Front. Immunol. 2019, 10, 2739. [Google Scholar] [CrossRef] [Green Version]
- Tavakoli, M.; Jeddi-Tehrani, M.; Salek-Moghaddam, A.; Rajaei, S.; Mohammadzadeh, A.; Sheikhhasani, S.; Kazemi-Sefat, G.E.; Zarnani, A.H. Effects of 1,25(OH)2 vitamin D3 on cytokine production by endometrial cells of women with recurrent spontaneous abortion. Fertil Steril. 2011, 96, 751–757. [Google Scholar] [CrossRef]
- Rajaei, S.; Mirahmadian, M.; Jeddi-Tehrani, M.; Tavakoli, M.; Zonoobi, M.; Dabbagh, A.; Zarnani, A.H. Effect of 1,25(OH)2 vitamin D3 on cytokine production by endometrial cells of women with repeated implantation failure. Gynecol. Endocrinol. 2012, 28, 906–911. [Google Scholar] [CrossRef]
- Penna, G.; Adorini, L. 1 Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J. Immunol. 2000, 164, 2405–2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.S.; Choi, M.Y.; Longtine, M.S.; Nelson, D.M. Vitamin D effects on pregnancy and the placenta. Placenta 2010, 31, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darrasse-Jeze, G.; Klatzmann, D.; Charlotte, F.; Salomon, B.L.; Cohen, J.L. CD4+CD25+ regulatory/suppressor T cells prevent allogeneic fetus rejection in mice. Immunol Lett 2006, 102, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Adorini, L.; Penna, G. Control of autoimmune diseases by the vitamin D endocrine system. Nat. Clin. Pract. Rheumatol. 2008, 4, 404–412. [Google Scholar] [CrossRef]
- Kamen, D.L.; Tangpricha, V. Vitamin D and molecular actions on the immune system: Modulation of innate and autoimmunity. J. Mol. Med. (Berl.) 2010, 88, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, D.; Evans, K.N.; Kilby, M.D.; Bulmer, J.N.; Innes, B.A.; Stewart, P.M.; Hewison, M. The ontogeny of 25-hydroxyvitamin D(3) 1alpha-hydroxylase expression in human placenta and decidua. Am. J. Pathol. 2002, 161, 105–114. [Google Scholar] [CrossRef]
- Ganguly, A.; Tamblyn, J.A.; Finn-Sell, S.; Chan, S.Y.; Westwood, M.; Gupta, J.; Kilby, M.D.; Gross, S.R.; Hewison, M. Vitamin D, the placenta and early pregnancy: Effects on trophoblast function. J. Endocrinol. 2018, 236, R93–R103. [Google Scholar] [CrossRef]
- Ikemoto, Y.; Kuroda, K.; Nakagawa, K.; Ochiai, A.; Ozaki, R.; Murakami, K.; Jinushi, M.; Matsumoto, A.; Sugiyama, R.; Takeda, S. Vitamin D Regulates Maternal T-Helper Cytokine Production in Infertile Women. Nutrients 2018, 10, 902. [Google Scholar] [CrossRef] [Green Version]
- Balogh, G.; de Boland, A.R.; Boland, R.; Barja, P. Effect of 1,25(OH)(2)-vitamin D(3) on the activation of natural killer cells: Role of protein kinase C and extracellular calcium. Exp. Mol. Pathol. 1999, 67, 63–74. [Google Scholar] [CrossRef]
- Liu, N.Q.; Larner, D.P.; Yao, Q.; Chun, R.F.; Ouyang, Y.; Zhou, R.; Tamblyn, J.A.; Wagner, C.L.; Hewison, M. Vitamin D-deficiency and sex-specific dysregulation of placental inflammation. J. Steroid Biochem. Mol. Biol. 2018, 177, 223–230. [Google Scholar] [CrossRef]
- Ota, K.; Dambaeva, S.; Kim, M.W.; Han, A.R.; Fukui, A.; Gilman-Sachs, A.; Beaman, K.; Kwak-Kim, J. 1,25-Dihydroxy-vitamin D3 regulates NK-cell cytotoxicity, cytokine secretion, and degranulation in women with recurrent pregnancy losses. Eur. J. Immunol. 2015, 45, 3188–3199. [Google Scholar] [CrossRef] [PubMed]
- Barrera, D.; Noyola-Martinez, N.; Avila, E.; Halhali, A.; Larrea, F.; Diaz, L. Calcitriol inhibits interleukin-10 expression in cultured human trophoblasts under normal and inflammatory conditions. Cytokine 2011, 57, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.; Noyola-Martinez, N.; Barrera, D.; Hernandez, G.; Avila, E.; Halhali, A.; Larrea, F. Calcitriol inhibits TNF-alpha-induced inflammatory cytokines in human trophoblasts. J. Reprod. Immunol. 2009, 81, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martinez, N.; Diaz, L.; Avila, E.; Halhali, A.; Larrea, F.; Barrera, D. Calcitriol downregulates TNF-alpha and IL-6 expression in cultured placental cells from preeclamptic women. Cytokine 2013, 61, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Yan, X.T.; Yan, C.F.; Zhang, X.W.; Hui, L.Y.; Xue, M.; Yu, X.W. Women with Recurrent Miscarriage Have Decreased Expression of 25-Hydroxyvitamin D3-1alpha-Hydroxylase by the Fetal-Maternal Interface. PLoS ONE 2016, 11, e0165589. [Google Scholar] [CrossRef]
- Ota, K.; Dambaeva, S.; Han, A.R.; Beaman, K.; Gilman-Sachs, A.; Kwak-Kim, J. Vitamin D deficiency may be a risk factor for recurrent pregnancy losses by increasing cellular immunity and autoimmunity. Hum. Reprod. 2014, 29, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wu, H.M.; Hang, F.; Zhang, Y.S.; Li, M.J. Women with recurrent spontaneous abortion have decreased 25(OH) vitamin D and VDR at the fetal-maternal interface. Braz. J. Med. Biol. Res. 2017, 50, e6527. [Google Scholar] [CrossRef] [Green Version]
- Fukui, A.; Kwak-Kim, J.; Ntrivalas, E.; Gilman-Sachs, A.; Lee, S.K.; Beaman, K. Intracellular cytokine expression of peripheral blood natural killer cell subsets in women with recurrent spontaneous abortions and implantation failures. Fertil. Steril. 2008, 89, 157–165. [Google Scholar] [CrossRef]
- Wu, L.; Vendiola, J.A.; Salazar Garcia, M.D.; Sung, N.; Skariah, A.; Gilman-Sachs, A.; Dambaeva, S.; Kwak-Kim, J. Poor ovarian response is associated with serum vitamin D levels and pro-inflammatory immune responses in women undergoing in-vitro fertilization. J. Reprod. Immunol. 2019, 136, 102617. [Google Scholar] [CrossRef]
- Chen, X.; Yin, B.; Lian, R.C.; Zhang, T.; Zhang, H.Z.; Diao, L.H.; Li, Y.Y.; Huang, C.Y.; Liang, D.S.; Zeng, Y. Modulatory effects of vitamin D on peripheral cellular immunity in patients with recurrent miscarriage. Am. J. Reprod. Immunol. 2016, 76, 432–438. [Google Scholar] [CrossRef]
- Ji, J.; Zhai, H.; Zhou, H.; Song, S.; Mor, G.; Liao, A. The role and mechanism of vitamin D-mediated regulation of Treg/Th17 balance in recurrent pregnancy loss. Am. J. Reprod. Immunol. 2019, 81, e13112. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, M.; Gharagozloo, M.; Ghahiri, A.; Mehrabian, F.; Maracy, M.R.; Kouhpayeh, S.; Pieper, I.L.; Rezaei, A. Altered Th17/Treg Ratio in Recurrent Miscarriage after Treatment with Paternal Lymphocytes and Vitamin D3: A Double-Blind Placebo-Controlled Study. Iran. J. Immunol. 2015, 12, 252–262. [Google Scholar] [PubMed]
- Ibrahim, Z.; Madany, E.; Abdel Aal, R.; El Biely, M. Role of 1,25-dihydroxyvitamin D (vitamin D3) as immunomodulator in recurrent missed miscarriage. Middle East Fertil. Soc. J. 2013, 18, 171–176. [Google Scholar] [CrossRef]
- Samimi, M.; Foroozanfard, F.; Amini, F.; Sehat, M. Effect of vitamin D supplementation on unexplained recurrent spontaneous abor- tion: A double-blind randomized controlled trial. Glob. J. Health Sci. 2016, 9, 95. [Google Scholar] [CrossRef]
- ESHRE Guideline Group on RPL; Bender Atik, R.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; Nelen, W.; Peramo, B.; Quenby, S.; et al. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2018, hoy004. [Google Scholar] [CrossRef]
- Practice Committee of the American Society for Reproductive Medicine. Evaluation and treatment of recurrent pregnancy loss: A committee opinion. Fertil. Steril. 2012, 98, 1103–1111. [Google Scholar] [CrossRef]
- Practice Committee of the American Society for Reproductive Medicine. Definitions of infertility and recurrent pregnancy loss. Fertil. Steril. 2008, 89, 1603. [Google Scholar] [CrossRef]
- Toth, B.; Würfel, W.; Bohlmann, M.; Zschocke, J.; Rudnik-Schöneborn, S.; Nawroth, F.; Schleußner, E.; Rogenhofer, N.; Wischmann, T.; von Wolff, M.; et al. Recurrent Miscarriage: Diagnostic and Therapeutic Procedures. Guideline of the DGGG, SGGG and OEGGG (S2k-Level, AWMF Registry No.015/050). Geburtshilfe Frauenheilkd. 2018, 78, 364–381. [Google Scholar] [CrossRef] [Green Version]
- Piccinni, M.P.; Scaletti, C.; Vultaggio, A.; Maggi, E.; Romagnani, S. Defective production of LIF, M-CSF and Th2-type cytokines by T cells at fetomaternal interface is associated with pregnancy loss. J. Reprod. Immunol. 2001, 52, 35–43. [Google Scholar] [CrossRef]
- Ng, S.C.; Gilman-Sachs, A.; Thaker, P.; Beaman, K.D.; Beer, A.E.; Kwak-Kim, J. Expression of intracellular Th1 and Th2 cytokines in women with recurrent spontaneous abortion, implantation failures after IVF/ET or normal pregnancy. Am. J. Reprod. Immunol. 2002, 48, 77–86. [Google Scholar] [CrossRef]
- Kwak-Kim, J.Y.; Chung-Bang, H.S.; Ng, S.C.; Ntrivalas, E.I.; Mangubat, C.P.; Beaman, K.D.; Beer, A.E.; Gilman-Sachs, A. Increased T helper 1 cytokine responses by circulating T cells are present in women with recurrent pregnancy losses and in infertile women with multiple implantation failures after IVF. Hum. Reprod. 2003, 18, 767–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak-Kim, J.; Lee, S.K.; Gilman-Sachs, A. Elevated Th1/Th2 cell ratios in a pregnant woman with a history of RSA, secondary Sjogren’s syndrome and rheumatoid arthritis complicated with one fetal demise of twin pregnancy. Am. J. Reprod. Immunol. 2007, 58, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, J.; Huang, S.Y.; Sun, X. Characterization of the subsets of human NKT-like cells and the expression of Th1/Th2 cytokines in patients with unexplained recurrent spontaneous abortion. J. Reprod. Immunol. 2015, 110, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, H.; Sugi, T.; Arai, T.; Kondo, A.; Suzuki, T.; Izumi, S.; McIntyre, J.A.; Makino, T. Different antiphospholipid antibody specificities are found in association with early repeated pregnancy loss versus recurrent IVF-failure patients. Am. J. Reprod. Immunol. 2001, 46, 323–329. [Google Scholar] [CrossRef]
- Ticconi, C.; Rotondi, F.; Veglia, M.; Pietropolli, A.; Bernardini, S.; Ria, F.; Caruso, A.; Di Simone, N. Antinuclear autoantibodies in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2010, 64, 384–392. [Google Scholar] [CrossRef]
- Lee, S.K.; Na, B.J.; Kim, J.Y.; Hur, S.E.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. Determination of clinical cellular immune markers in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2013, 70, 398–411. [Google Scholar] [CrossRef]
- Piosik, Z.M.; Goegebeur, Y.; Klitkou, L.; Steffensen, R.; Christiansen, O.B. Plasma TNF-alpha levels are higher in early pregnancy in patients with secondary compared with primary recurrent miscarriage. Am. J. Reprod. Immunol. 2013, 70, 347–358. [Google Scholar] [CrossRef]
- Goncalves, D.R.; Braga, A.; Braga, J.; Marinho, A. Recurrent pregnancy loss and vitamin D: A review of the literature. Am. J. Reprod. Immunol. 2018, 80, e13022. [Google Scholar] [CrossRef]
- Tavakoli, M.; Salek-Moghaddam, A.; Jeddi-Tehrani, M.; Talebi, S.; Kazemi-Sefat, G.E.; Vafaei, S.; Mohammadzadeh, A.; Sheikhhassani, S.; Zarnani, A.H. Comparable vitamin D3 metabolism in the endometrium of patients with recurrent spontaneous abortion and fertile controls. Mol. Reprod. Dev. 2015, 82, 356–364. [Google Scholar] [CrossRef]
- Yan, X.; Wang, L.; Yan, C.; Zhang, X.; Hui, L.; Sheng, Q.; Xue, M.; Yu, X. Decreased expression of the vitamin D receptor in women with recurrent pregnancy loss. Arch. Biochem. 2016, 606, 128–133. [Google Scholar] [CrossRef]
- Macklon, N. Recurrent implantation failure is a pathology with a specific transcriptomic signature. Fertil. Steril. 2017, 108, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Aflatoonian, A.; Arabjahvani, F.; Eftekhar, M.; Sayadi, M. Effect of vitamin D insufficiency treatment on fertility outcomes in frozen-thawed embryo transfer cycles: A randomized clinical trial. Iran. J. Reprod. Med. 2014, 12, 595–600. [Google Scholar] [PubMed]
- Adamson G, D.S.; Chambers, G.; Ishihara, O.; Mansour, R.; Banker, M. International Committee for Monitoring Assisted Reproductive Technologies (ICMART) Preliminary World Report on ART, 2015. Abstract ESHRE, Vienna 2019, 34: i65. Available online: https://academic.oup.com/humrep/article-abstract/34/Supplement_1/i1/5528444?redirectedFrom=fulltext (accessed on 11 May 2020).
- Dressler, N.; Chandra, A.; Aguirre Davila, L.; Spineli, L.M.; Schippert, C.; von Versen-Hoynck, F. BMI and season are associated with vitamin D deficiency in women with impaired fertility: A two-centre analysis. Arch. Gynecol. Obstet. 2016, 293, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, S.; Jindal, S.; Greenseid, K.; Shu, J.; Zeitlian, G.; Hickmon, C.; Pal, L. Replete vitamin D stores predict reproductive success following in vitro fertilization. Fertil. Steril. 2010, 94, 1314–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firouzabadi, R.D.; Rahmani, E.; Rahsepar, M.; Firouzabadi, M.M. Value of follicular fluid vitamin D in predicting the pregnancy rate in an IVF program. Arch. Gynecol. Obstet. 2014, 289, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Rudick, B.; Ingles, S.; Chung, K.; Stanczyk, F.; Paulson, R.; Bendikson, K. Characterizing the influence of vitamin D levels on IVF outcomes. Hum. Reprod. 2012, 27, 3321–3327. [Google Scholar] [CrossRef]
- Garbedian, K.; Boggild, M.; Moody, J.; Liu, K.E. Effect of vitamin D status on clinical pregnancy rates following in vitro fertilization. CMAJ Open 2013, 1, E77–E82. [Google Scholar] [CrossRef] [Green Version]
- Paffoni, A.; Ferrari, S.; Vigano, P.; Pagliardini, L.; Papaleo, E.; Candiani, M.; Tirelli, A.; Fedele, L.; Somigliana, E. Vitamin D deficiency and infertility: Insights from in vitro fertilization cycles. J. Clin. Endocrinol. Metab. 2014, 99, E2372–E2376. [Google Scholar] [CrossRef]
- Hypertension in Pregnancy. Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy. Obstet. Gynecol. 2013, 122, 1122–1131. [Google Scholar] [CrossRef]
- Makhseed, M.; Raghupathy, R.; Azizieh, F.; Al-Azemi, M.M.; Hassan, N.A.; Bandar, A. Mitogen-induced cytokine responses of maternal peripheral blood lymphocytes indicate a differential Th-type bias in normal pregnancy and pregnancy failure. Am. J. Reprod. Immunol. 1999, 42, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Sakai, M.; Sasaki, Y.; Tanebe, K.; Tsuda, H.; Michimata, T. Quantitative analysis of peripheral blood Th0, Th1, Th2 and the Th1:Th2 cell ratio during normal human pregnancy and preeclampsia. Clin. Exp. Immunol. 1999, 117, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Conrad, K.P.; Miles, T.M.; Benyo, D.F. Circulating levels of immunoreactive cytokines in women with preeclampsia. Am. J. Reprod. Immunol. 1998, 40, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Sakai, M. Th1/Th2 balance in preeclampsia. J. Reprod. Immunol. 2003, 59, 161–173. [Google Scholar] [CrossRef]
- Szarka, A.; Rigo, J., Jr.; Lazar, L.; Beko, G.; Molvarec, A. Circulating cytokines, chemokines and adhesion molecules in normal pregnancy and preeclampsia determined by multiplex suspension array. BMC Immunol. 2010, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Yao, M.Z.; Liu, J.B.; Xiong, L.K. A meta-analysis of tumor necrosis factor-alpha, interleukin-6, and interleukin-10 in preeclampsia. Cytokine 2011, 56, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.Y.; Guild, S.J.; Barrett, C.J.; Chen, Q.; McCowan, L.; Jordan, V.; Chamley, L.W. Tumor necrosis factor-alpha, interleukin-6, and interleukin-10 levels are altered in preeclampsia: A systematic review and meta-analysis. Am. J. Reprod. Immunol. 2013, 70, 412–427. [Google Scholar] [CrossRef]
- Sacks, G.P.; Studena, K.; Sargent, K.; Redman, C.W. Normal pregnancy and preeclampsia both produce inflammatory changes in peripheral blood leukocytes akin to those of sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86. [Google Scholar] [CrossRef]
- Rusterholz, C.; Hahn, S.; Holzgreve, W. Role of placentally produced inflammatory and regulatory cytokines in pregnancy and the etiology of preeclampsia. Semin. Immunopathol. 2007, 29, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Bauer, S.; Pollheimer, J.; Hartmann, J.; Husslein, P.; Aplin, J.D.; Knofler, M. Tumor necrosis factor-alpha inhibits trophoblast migration through elevation of plasminogen activator inhibitor-1 in first-trimester villous explant cultures. J. Clin. Endocrinol. Metab. 2004, 89, 812–822. [Google Scholar] [CrossRef]
- Xu, B.; Nakhla, S.; Makris, A.; Hennessy, A. TNF-alpha inhibits trophoblast integration into endothelial cellular networks. Placenta 2011, 32, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Pober, J.S.; Cotran, R.S. Cytokines and endothelial cell biology. Physiol. Rev. 1990, 70, 427–451. [Google Scholar] [CrossRef]
- Bodnar, L.M.; Catov, J.M.; Roberts, J.M. Racial/ethnic differences in the monthly variation of preeclampsia incidence. Am. J. Obstet. Gynecol. 2007, 196, 324.e1–324.e5. [Google Scholar] [CrossRef] [PubMed]
- TePoel, M.R.; Saftlas, A.F.; Wallis, A.B. Association of seasonality with hypertension in pregnancy: A systematic review. J. Reprod. Immunol. 2011, 89, 140–152. [Google Scholar] [CrossRef] [PubMed]
- August, P.; Marcaccio, B.; Gertner, J.M.; Druzin, M.L.; Resnick, L.M.; Laragh, J.H. Abnormal 1,25-dihydroxyvitamin D metabolism in preeclampsia. Am. J. Obstet. Gynecol. 1992, 166, 1295–1299. [Google Scholar] [CrossRef]
- Bodnar, L.M.; Catov, J.M.; Simhan, H.N.; Holick, M.F.; Powers, R.W.; Roberts, J.M. Maternal vitamin D deficiency increases the risk of preeclampsia. J. Clin. Endocrinol. Metab. 2007, 92, 3517–3522. [Google Scholar] [CrossRef] [Green Version]
- Tabesh, M.; Salehi-Abargouei, A.; Esmaillzadeh, A. Maternal vitamin D status and risk of pre-eclampsia: A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2013, 98, 3165–3173. [Google Scholar] [CrossRef] [Green Version]
- Hypponen, E.; Cavadino, A.; Williams, D.; Fraser, A.; Vereczkey, A.; Fraser, W.D.; Banhidy, F.; Lawlor, D.; Czeizel, A.E. Vitamin D and Pre-Eclampsia: Original Data, Systematic Review and Meta-Analysis. Ann. Nutr. Metab. 2014, 63, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Akbari, S.; Khodadadi, B.; Ahmadi, S.A.Y.; Abbaszadeh, S.; Shahsavar, F. Association of vitamin D level and vitamin D deficiency with risk of preeclampsia: A systematic review and updated meta-analysis. Taiwan J. Obstet. Gynecol. 2018, 57, 241–247. [Google Scholar] [CrossRef]
- Hamedanian, L.; Badehnoosh, B.; Razavi-Khorasani, N.; Mohammadpour, Z.; Mozaffari-Khosravi, H. Evaluation of vitamin D status, parathyroid hormone, and calcium among Iranian pregnant women with preeclampsia: A case-control study. Int. J. Reprod. Biomed. (Yazd.) 2019, 17, 831–840. [Google Scholar] [CrossRef]
- Baker, A.M.; Haeri, S.; Camargo, C.A., Jr.; Espinola, J.A.; Stuebe, A.M. A Nested Case-Control Study of Midgestation Vitamin D Deficiency and Risk of Severe Preeclampsia. J. Clin. Endocrinol. Metab. 2010, 95, 5105–5109. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.J.; Wagner, C.L.; Hollis, B.W.; Baatz, J.E.; Johnson, D.D. Maternal vitamin D and fetal growth in early-onset severe preeclampsia. Am. J. Obstet. Gynecol. 2011, 204, 556.e1–556.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodnar, L.M.; Simhan, H.N.; Catov, J.M.; Roberts, J.M.; Platt, R.W.; Diesel, J.C.; Klebanoff, M.A. Maternal vitamin d status and the risk of mild and severe preeclampsia. Epidemiology 2014, 25, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Villa, A.R.; Madrazo, E.; Soria, M.C.; Mercado, E.; Diaz, L.; Avila, E.; Garabedian, M.; Larrea, F. Longitudinal changes in maternal serum 1,25-dihydroxyvitamin D and insulin like growth factor I levels in pregnant women who developed preeclampsia: Comparison with normotensive pregnant women. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Seely, E.W.; Rana, S.; Bhan, I.; Ecker, J.; Karumanchi, S.A.; Thadhani, R. First trimester vitamin D, vitamin D binding protein, and subsequent preeclampsia. Hypertension 2010, 56, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Shand, A.W.; Nassar, N.; Von Dadelszen, P.; Innis, S.M.; Green, T.J. Maternal vitamin D status in pregnancy and adverse pregnancy outcomes in a group at high risk for pre-eclampsia. BJOG 2010, 117, 1593–1598. [Google Scholar] [CrossRef]
- Olsen, S.F.; Secher, N.J. A possible preventive effect of low-dose fish oil on early delivery and pre-eclampsia: Indications from a 50-year-old controlled trial. Br. J. Nutr. 1990, 64, 599–609. [Google Scholar] [CrossRef]
- Haugen, M.; Brantsaeter, A.L.; Trogstad, L.; Alexander, J.; Roth, C.; Magnus, P.; Meltzer, H.M. Vitamin D supplementation and reduced risk of preeclampsia in nulliparous women. Epidemiology 2009, 20, 720–726. [Google Scholar] [CrossRef]
- Behjat Sasan, S.; Zandvakili, F.; Soufizadeh, N.; Baybordi, E. The Effects of Vitamin D Supplement on Prevention of Recurrence of Preeclampsia in Pregnant Women with a History of Preeclampsia. Obstet. Gynecol. Int. 2017, 2017, 8249264. [Google Scholar] [CrossRef] [Green Version]
- Marya, R.K.; Rathee, S.; Manrow, M. Effect of calcium and vitamin D supplementation on toxaemia of pregnancy. Gynecol. Obstet. Investig 1987, 24, 38–42. [Google Scholar] [CrossRef]
- Hollis, B.W.; Wagner, C.L. Randomized controlled trials to determine the safety of vitamin D supplementation during pregnancy and lactation. In Proceedings of the Fourteenth Workshop on Vitamin D, Brugge, Belgium, 4–8 October 2009; p. 134. [Google Scholar]
- Hewison, M. Vitamin D and the immune system: New perspectives on an old theme. Endocrinol. Metab. Clin. N. Am. 2012, 39, 365–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrera, D.; Avila, E.; Hernandez, G.; Mendez, I.; Gonzalez, L.; Halhali, A.; Larrea, F.; Morales, A.; Diaz, L. Calcitriol affects hCG gene transcription in cultured human syncytiotrophoblasts. Reprod. Biol. Endocrinol. 2008, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemire, J.M.; Archer, D.C.; Beck, L.; Spiegelberg, H.L. Immunosuppressive actions of 1,25-dihydroxyvitamin D3: Preferential inhibition of Th1 functions. J. Nutr. 1995, 125, 1704S–1708S. [Google Scholar] [PubMed]
- Dong, M.; He, J.; Wang, Z.; Xie, X.; Wang, H. Placental imbalance of Th1- and Th2-type cytokines in preeclampsia. Acta Obstet. Gynecol. Scand. 2005, 84, 788–793. [Google Scholar] [CrossRef]
- Diaz, L.; Arranz, C.; Avila, E.; Halhali, A.; Vilchis, F.; Larrea, F. Expression and activity of 25-hydroxyvitamin D-1 alpha-hydroxylase are restricted in cultures of human syncytiotrophoblast cells from preeclamptic pregnancies. J. Clin. Endocrinol. Metab. 2002, 87, 3876–3882. [Google Scholar]
- Halhali, A.; Diaz, L.; Barrera, D.; Avila, E.; Larrea, F. Placental calcitriol synthesis and IGF-I levels in normal and preeclamptic pregnancies. J. Steroid Biochem. Mol. Biol. 2013. [Google Scholar] [CrossRef]
- Cardus, A.; Panizo, S.; Encinas, M.; Dolcet, X.; Gallego, C.; Aldea, M.; Fernandez, E.; Valdivielso, J.M. 1,25-dihydroxyvitamin D3 regulates VEGF production through a vitamin D response element in the VEGF promoter. Atherosclerosis 2009, 204, 85–89. [Google Scholar] [CrossRef]
- Ajabshir, S.; Asif, A.; Nayer, A. The effects of vitamin D on the renin-angiotensin system. J. Nephropathol. 2014, 3, 41–43. [Google Scholar] [CrossRef]
- Brodowski, L.; Burlakov, J.; Myerski, A.C.; von Kaisenberg, C.S.; Grundmann, M.; Hubel, C.A.; von Versen-Höynck, F. Vitamin D prevents endothelial progenitor cell dysfunction induced by sera from women with preeclampsia or conditioned media from hypoxic placenta. PLoS ONE 2014, 9, e98527. [Google Scholar] [CrossRef] [Green Version]
- von Versen-Höynck, F.; Brodowski, L.; Dechend, R.; Myerski, A.C.; Hubel, C.A. Vitamin D antagonizes negative effects of preeclampsia on fetal endothelial colony forming cell number and function. PLoS ONE 2014, 9, e98990. [Google Scholar] [CrossRef]
- Chawanpaiboon, S.; Vogel, J.P.; Moller, A.B.; Lumbiganon, P.; Petzold, M.; Hogan, D.; Landoulsi, S.; Jampathong, N.; Kongwattanakul, K.; Laopaiboon, M.; et al. Global, regional, and national estimates of levels of preterm birth in 2014: A systematic review and modelling analysis. Lancet Glob. Health 2019, 7, e37–e46. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lopez, N.; StLouis, D.; Lehr, M.A.; Sanchez-Rodriguez, E.N.; Arenas-Hernandez, M. Immune cells in term and preterm labor. Cell. Mol. Immunol. 2014, 11, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.S.; Tao, Y.H.; Huang, K.; Zhu, B.B.; Tao, F.B. Vitamin D and risk of preterm birth: Up-to-date meta-analysis of randomized controlled trials and observational studies. J. Obstet. Gynaecol. Res. 2017, 43, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, C.L.; McNeil, R.B.; Johnson, D.D.; Hulsey, T.C.; Ebeling, M.; Robinson, C.; Hamilton, S.A.; Hollis, B.W. Health characteristics and outcomes of two randomized vitamin D supplementation trials during pregnancy: A combined analysis. J. Steroid Biochem. Mol. Biol. 2013, 136, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C.L.; Baggerly, C.; McDonnell, S.; Baggerly, K.A.; French, C.B.; Baggerly, L.; Hamilton, S.A.; Hollis, B.W. Post-hoc analysis of vitamin D status and reduced risk of preterm birth in two vitamin D pregnancy cohorts compared with South Carolina March of Dimes 2009-2011 rates. J. Steroid Biochem. Mol. Biol. 2016, 155, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Palacios, C.; Kostiuk, L.K.; Pena-Rosas, J.P. Vitamin D supplementation for women during pregnancy. Cochrane Database Syst. Rev. 2019, 7, Cd008873. [Google Scholar] [CrossRef]
- Delorme, A.C.; Danan, J.L.; Acker, M.G.; Ripoche, M.A.; Mathieu, H. In rat uterus 17 beta-estradiol stimulates a calcium-binding protein similar to the duodenal vitamin D-dependent calcium-binding protein. Endocrinology 1983, 113, 1340–1347. [Google Scholar] [CrossRef]
- Tribe, R.M. Regulation of human myometrial contractility during pregnancy and labour: Are calcium homeostatic pathways important? Exp. Physiol. 2001, 86, 247–254. [Google Scholar] [CrossRef]
- Hewison, M. Antibacterial effects of vitamin D. Nat. Rev. Endocrinol. 2011, 7, 337–345. [Google Scholar] [CrossRef]
- Liu, N.Q.; Hewison, M. Vitamin D, the placenta and pregnancy. Arch. Biochem. Biophys. 2012, 523, 37–47. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Reference | Main Findings |
|---|---|
| In vivo studies | |
| [31] | Compared to wild-type mouse placentas, placentas of VDR and CYP27B1 knock-out mice show enhanced proinflammatory cytokine and chemokine expression. |
| [39] | Cholecalciferol supplementation reduces the rate of LPS-induced abortions in mice. Additionally, cholecalciferol inhibits immunological modulations induced by LPS. |
| [61] | Vitamin D deficient mice challenged with LPS in pregnancy have higher IP-10, MCP-1, SAP, TIMP-1, VCAM-1, vWF and lower GCP-2 levels than vitamin D sufficient mice. |
| In vitro studies | |
| [22] | IL-1β induces CYP27B1 mRNA expression in human decidua cells. |
| [37] | Decidual NK cells treated with 1,25-(OH)2D or 25(OH)D synthesize less cytokines, but more CAMP. |
| [38] | In human trophoblasts, 1,25-(OH)2D induces CAMP expression. A 3A trophoblast cell line treated with 1,25-(OH)2D shows decreased colony forming units, when infected with E. coli. |
| [50] | After 1,25-(OH)2D treatment, cytokine expression in WECs from patients with unexplained RPL are reduced and shifted toward a Th2 phenotype. In ESCs, cytokine production is overall down-reguated, but TGF-β production is stimulated. |
| [51] | In WECs from RIF and normal patients, 1,25-(OH)2D reduces most cytokine production, whereas IL-8 is elevated. In ESCs, similar 1,25-(OH)2D effects are observed, except for an up-regulation of TGF-β in the RIF group. |
| [62] | In women with RPL, 1,25-(OH)2D has immune regulatory effects on NK cell cytotoxicity, cytokine secretion, degranulation process and TLR4 expression. |
| [63] | 1,25-(OH)2D reduces IL-10 production in trophoblasts from normal and preeclamptic placentas. |
| [64] | TNF-α-induced immune response and cytokine production in human trophoblasts is inhibited by 11,25-(OH)2D. |
| [65] | TNF-α and IL-6 secretion and mRNA expression in human trophoblasts are reduced by 1,25-(OH)2D. |
| Observational studies | |
| [32] | Treg cell population is lower in maternal blood and cord blood in 25(OH)D3 deficient pregnant women. CD23 and CD21 B cell population is higher in maternal blood and cord blood in 25(OH)D deficient pregnant women. TGF-β and IL-10 levels are lower. |
| [35] | Maternal serum 25(OH)D deficiency is associated with placental inflammation. |
| [66] | Women with RPL have a lower level of CYP27B1 expression in chorionic villi and decidua compared with normal pregnant women. CYP27B1 and cytokine expression (IL-10, IFN-γ, TNF-α and IL-2) co-localize in chorionic villi and decidua cells. |
| [67] | In women with RPL, low 25(OH)D3 levels are associated with abnormalities in cellular immunity and cytokine production. |
| [68] | Decidual tissues of patients with RPL show less 25(OH)D, TGF-β and VDR expression and significant increase in IL-23 and IL-17. |
| [69] | Natural killer-1 shift in peripheral blood NK cells was identified in nonpregnant women with RPL and implantation failures. |
| [70] | In women with POR and low serum 25(OH)D, NK cell levels and cytotoxicity are higher. CD19+ B cell levels are higher, as well as the Th1/Th2 cell ratio. |
| Interventional studies | |
| [59] | Vitamin D supplementation in infertile women with insufficient 25(OH)D decreases Th1/Th2 ratio. In endometrial biopsies, 1,25(OH)2D3treatment reduces IFN-γ. |
| [71] | Higher percentages of CD19+ B cells and NK cytotoxicity, as well as a higher percentage of TNF-α-expressing Th cells are observed in RPL patients with low serum 25(OH)D levels and can be regulated to some extent with 1,25(OH)2D supplementation. |
| [72] | 25(OH)D levels and Treg/Th17 ratios are decreased in women with RPL. Vitamin D supplementation increases Treg/Th17 ratio, VDR and CYP24A1 expression. |
| RCTs | |
| [73] | Vitamin D supplementation reduces Th17 cell population in peripheral blood from women with RM and reduces Th17/Treg ratio. |
| [74] | Patients with a history of RPL who recieved vitamin D reveal lower serum IFN-γ levels. The risk of miscarriage is reduced by 15% compared to untreated patients. |
| [75] | Vitamin D supplementation in women with unexplained RPL decreases serum IL-23 levels and reduces the frequency of miscarriages. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder-Heurich, B.; Springer, C.J.P.; von Versen-Höynck, F. Vitamin D Effects on the Immune System from Periconception through Pregnancy. Nutrients 2020, 12, 1432. https://doi.org/10.3390/nu12051432
Schröder-Heurich B, Springer CJP, von Versen-Höynck F. Vitamin D Effects on the Immune System from Periconception through Pregnancy. Nutrients. 2020; 12(5):1432. https://doi.org/10.3390/nu12051432
Chicago/Turabian StyleSchröder-Heurich, Bianca, Clara Juliane Pacifica Springer, and Frauke von Versen-Höynck. 2020. "Vitamin D Effects on the Immune System from Periconception through Pregnancy" Nutrients 12, no. 5: 1432. https://doi.org/10.3390/nu12051432