Nutritional Composition and Microbial Communities of Two Non-alcoholic Traditional Fermented Beverages from Zambia: A Study of Mabisi and Munkoyo

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Measurement of Nutrient Value
2.2.1. Proximate Analysis
2.2.2. Atomic Absorption Spectrophotometry (AAS)
2.2.3. Quality Control (QC)
2.2.4. Analysis of B-Vitamins (B1, B2, B3, B6 and B12) by High Pressure Liquid Chromatography (HPLC)
2.2.5. Standard Preparation
2.2.6. Quality Control
2.3. Characterization of Microbial Composition of Traditonal Fermented Foods Mabisi and Munkoyo
2.3.1. Total Genomic DNA Extraction
2.3.2. 16SrRNA Amplicon Sequencing of DNA Samples and Analysis of Sequence Data
2.4. Statistical Analysis
3. Results
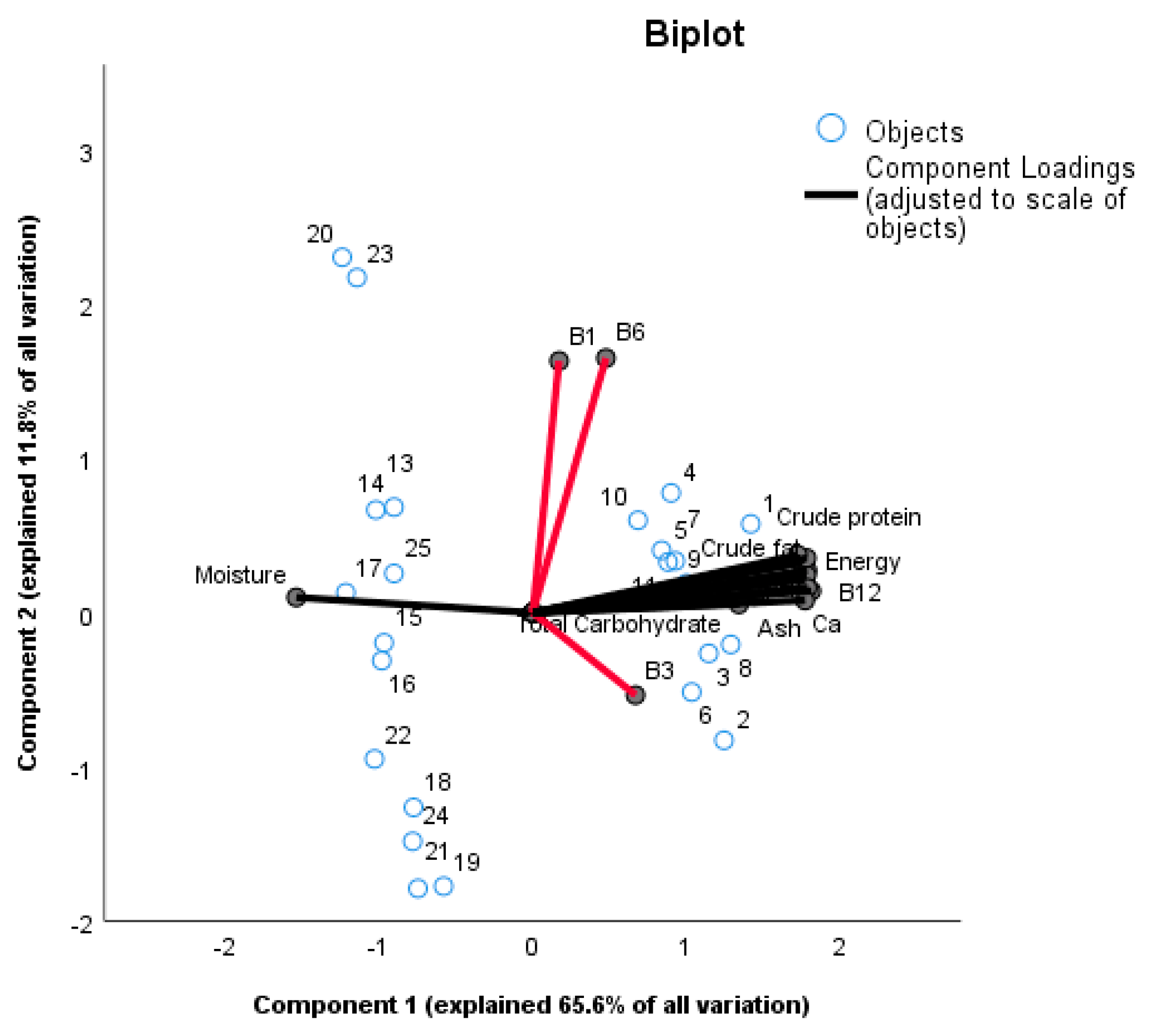
3.1. Nutritional Analyses
3.2. Microbial Analyses
4. Discussion
4.1. Nutritional Composition
4.2. Microbial Community Composition
4.3. Factors that Affect Microbial Community Composition
5. Conclusions and Significance
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nout, M.J.R. Rich nutrition from the poorest—Cereal fermentations in Africa and Asia. Food Microbiol. 2009, 26, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Battcock, M.; Azam-Ali, S. Fermented Fruits and Vegetables. A Global Perspective. United Nations Rome Fao Agricultural Services Bulletin 134; FAO: Rome, Italy, 1998; pp. 10–15. [Google Scholar]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Hotz, C.; Gibson, R.S. Traditional food-processing and preparation practices to enhance the bioavailability of micronutrients in plant-based diets. J. Nutr. 2007, 137, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Ansorena, D.; Astiasarán, I. Fermented Foods: Composition and Health Effects; Oxford Academic Press: Oxford, UK, 2016. [Google Scholar]
- Ceapa, C.; Wopereis, H.; Rezaiki, L.; Kleerebezem, M.; Knol, J.; Oozeer, R. Influence of fermented milk products, prebiotics and probiotics on microbiota composition and health. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.C.; Rubio-Texeira, M.; Polaina, J. Lactose: The Milk Sugar from a Biotechnological Perspective. Crit. Rev. Food Sci. Nutr. 2005, 44, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Agyei, D.; Owusu-Kwarteng, J.; Akabanda, F.; Akomea-Frempong, S. Indigenous African fermented dairy products: Processing technology, microbiology and health benefits. Crit. Rev. Food Sci. Nutr. 2019, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Pérez Montoro, B.; Casimiro-Soriguer, C.S.; Pérez Pulido, A.J.; Knapp, C.W.; Caballero Gómez, N.; Castillo-Gutiérrez, S.; Estudillo-Martínez, M.D.; Gálvez, A.; Benomar, N. Insight into Potential Probiotic Markers Predicted in Lactobacillus pentosus MP-10 Genome Sequence. Front. Microbiol. 2017, 8, 891. [Google Scholar] [CrossRef] [Green Version]
- Linares, D.M.; Gómez, C.; Renes, E.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Stanton, C. Lactic Acid Bacteria and Bifidobacteria with Potential to Design Natural Biofunctional Health-Promoting Dairy Foods. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Saez-Lara, M.J.; Gomez-Llorente, C.; Plaza-Diaz, J.; Gil, A. The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: A systematic review of randomized human clinical trials. BioMed. Res. Int. 2015, 2015, 505878. [Google Scholar] [CrossRef]
- Kapila, S.; Sinha, P.R.; Singh, S. Influence of feeding fermented milk and non-fermented milk containing Lactobacillus casei on immune response in mice. Food Agric. Immunol. 2007, 18, 75–82. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Kasase, C.; Toarta, C.; Kassen, R.; Poulain, A.J. Microbial Community Structure of Three Traditional Zambian Fermented Products: Mabisi, Chibwantu and Munkoyo. PLoS ONE 2013, 8, e63948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moonga, H.B.; Schoustra, S.E.; Linnemann, A.R.; Kuntashula, E.; Shindano, J.; Smid, E.J. The art of mabisi production: A traditional fermented milk. PLoS ONE 2019, 14, e0213541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foma, R.K.; Destain, J.; Mobinzo, P.K.; Kayisu, K.; Thonart, P. Study of physicochemical parameters and spontaneous fermentation during traditional production of munkoyo, an indigenous beverage produced in Democratic Republic of Congo. Food Contr. 2012, 25, 334–341. [Google Scholar] [CrossRef]
- Phiri, S.; Schoustra, S.E.; van den Heuvel, J.; Smid, E.J.; Shindano, J.; Linnemann, A. Fermented cereal-based Munkoyo beverage: Processing practices, microbial diversity and aroma compounds. PLoS ONE 2019, 14, e0223501. [Google Scholar] [CrossRef]
- Phiri, S.; Schoustra, S.E.; van den Heuvel, J.; Smid, E.J.; Shindano, J.; Linnemann, A.R. How processing methods affect the microbial community composition in a cereal-based fermented beverage. LWT 2020, 128, 109451. [Google Scholar] [CrossRef]
- Groenenboom, A.E.; Parker, M.E.; de Vries, A.; de Groot, S.; Zobrist, S.; Mansen, K.; Milani, P.; Kort, R.; Smid, E.J.; Schoustra, S.E. Bacterial community dynamics in lait caillé, a traditional product of spontaneous fermentation from Senegal. PLoS ONE 2019, 14, e0215658. [Google Scholar] [CrossRef]
- Hotz, C.; Palaniappan, U.; Chileshe, J.; Kafwembe, E.; Siamusantu, W. Nutrition Survey in Central and Eastern Provinces, Zambia 2009: Focus on Vitamin a and Maize Intakes, and Vitamin a Status among Women and Children; National Food and Nutrition Commission of Zambia: Lusaka, Zambia, 2011. [Google Scholar]
- World Health Organization; FAO. Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; FAO: Rome, Italy, 2004. [Google Scholar]
- Efsa, N.D.A. Efsa Panel on Dietetic Products. Allergies. Scientific Opinion on Dietary Reference Values for cobalamin (vitamin B12). EFSA J. 2015, 13, 4150. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Ferguson, E.; Chege, P.; Kimiywe, J.; Wiesmann, D.; Hotz, C. Zinc, iron and calcium are major limiting nutrients in the complementary diets of rural Kenyan children. Matern. Child. Nutr. 2015, 3, 6–20. [Google Scholar] [CrossRef]
- Zafra-Gómez, A.; Garballo, A.; Morales, J.C.; García-Ayuso, L.E. Simultaneous Determination of Eight Water-Soluble Vitamins in Supplemented Foods by Liquid Chromatography. J. Agric. Food Chem. 2006, 54, 4531–4536. [Google Scholar] [CrossRef]
- Moreno, P.; Salvadó, V. Determination of eight water- and fat-soluble vitamins in multi-vitamin pharmaceutical formulations by high-performance liquid chromatography. J. Chromatogr. A 2000, 870, 207–215. [Google Scholar] [CrossRef]
- Groenenboom, A.E. Microbial Community Dynamics in Traditionally Fermented Milk; Wageningen University: Wageningen, The Netherlands, 2019. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Guidelines of Food Fortification with Micronutrients; WHO: Geneva, Switzerland, 2006. [Google Scholar]
- European Food Safety Authority. Dietary Reference Values for nutrients Summary report. EFSA Support. Publ. 2017, 14, e15121E. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Rouf Shah, T.; Prasad, K.; Kumar, P. Maize—A potential source of human nutrition and health: A review. Cogent Food Agric. 2016, 2. [Google Scholar] [CrossRef]
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional Value and Technological Suitability of Milk from Various Animal Species Used for Dairy Production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- De Vrese, M.; Stegelmann, A.; Richter, B.; Fenselau, S.; Laue, C.; Schrezenmeir, J. Probiotics—Compensation for lactase insufficiency. Am. J. Clin. Nut. 2001, 73, 421s–429s. [Google Scholar] [CrossRef] [Green Version]
- Chileshe, J.; Talsma, E.F.; Schoustra, S.E.; Borgonjen-van den Berg, K.J.; Handema, R.; Zwaan, B.J.; Brouwer, I.D. Potential contribution of cereal and milk based fermented foods to dietary nutrient intake of 1–5 years old children in Central province in Zambia. PLoS ONE 2020, 15, e0232824. [Google Scholar] [CrossRef]
- Kaliwile, C.; Michelo, C.; Titcomb, T.J.; Moursi, M.; Donahue Angel, M.; Reinberg, C.; Bwembya, P.; Alders, R.; Tanumihardjo, S.A. Dietary Intake Patterns among Lactating and Non-Lactating Women of Reproductive Age in Rural Zambia. Nutrients 2019, 11, 288. [Google Scholar] [CrossRef] [Green Version]
- Du Plooy, Z.; Schönfeldt, H.C.; Hall, N. The role of traditional foods in food-based dietary guidelines—A South African case study on maas (cultured milk). Food Chem. 2018, 238, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nout, M.J.R. Fermented foods and food safety. Food Res. Int. 1994, 27, 291–298. [Google Scholar] [CrossRef]
- Hardin, G. The Competitive Exclusion Principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.-H.; Jost, L. Phylogenetic diversity measures based on Hill numbers. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3599–3609. [Google Scholar] [CrossRef]
- Shangpliang, H.N.J.; Rai, R.; Keisam, S.; Jeyaram, K.; Tamang, J.P. Bacterial community in naturally fermented milk products of Arunachal Pradesh and Sikkim of India analysed by high-throughput amplicon sequencing. Sci. Rep. 2018, 8, 1532. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-F.; Liu, C.-J.; Zhang, H.-Y.; Gong, F.-M.; Luo, Y.-Y.; Li, X.-R. The bacterial community structure of yond bap, a traditional fermented goat milk product, from distinct Chinese regions. Dairy Sci. Technol. 2015, 95, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Székely, A.J.; Langenheder, S. The importance of species sorting differs between habitat generalists and specialists in bacterial communities. FEMS Microbiol. Ecol. 2014, 87, 102–112. [Google Scholar] [CrossRef]
- Loeuille, N.; Leibold, M.A. Evolution in Metacommunities: On the Relative Importance of Species Sorting and Monopolization in Structuring Communities. Am. Nat. 2008, 171, 788–799. [Google Scholar] [CrossRef] [Green Version]
- Nout, M.J.R.; Rombouts, F.M. Fermentative preservation of plant foods. J. Appl. Bacteriol. 1992, 73, 136s–147s. [Google Scholar] [CrossRef]
- Melini, F.; Melini, V.; Luziatelli, F.; Ficca, A.G.; Ruzzi, M. Health-Promoting Components in Fermented Foods: An Up-to-Date Systematic Review. Nutrients 2019, 11, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollán, G.C.; Gerez, C.L.; LeBlanc, J.G. Lactic Fermentation as a Strategy to Improve the Nutritional and Functional Values of Pseudocereals. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekar Rajendran, S.C.; Chamlagain, B.; Kariluoto, S.; Piironen, V.; Saris, P.E.J. Biofortification of riboflavin and folate in idli batter, based on fermented cereal and pulse, by Lactococcus lactis N8 and Saccharomyces boulardii SAA655. J. Appl. Microbiol. 2017, 122, 1663–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebo, O.A.; Kayitesi, E.; Njobeh, P.B. Reduction of Mycotoxins during Fermentation of Whole Grain Sorghum to Whole Grain Ting (a Southern African Food). Toxins 2019, 11, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample | Moisture | Dry Matter | Ash | Fibre | Crude Protein | Crude Fat | Total Carbohydrate | Energy | pH | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | SD | CV% | % | SD | CV% | % | SD | CV% | % | SD | CV% | % | SD | CV% | % | SD | CV% | % | SD | CV% | Kcal/100 g | SD | CV% | ||
| Mabisi | |||||||||||||||||||||||||
| 1 | 67.4 | ±0.378 | 0.6 | 32.6 | 0.400 | 1.3 | 0.47 | ±0.023 | 4.9 | 0 | ±0.000 | 0 | 4.68 | ±0.045 | 1.0 | 4.2 | ±0.181 | 4.3 | 23.28 | ±0.567 | 2.4 | 149.67 | ±0.747 | 0.5 | 4.1 |
| 2 | 85.2 | ±5.622 | 6.6 | 14.8 | 0.033 | 0.3 | 0.65 | ±0.006 | 0.9 | 0 | ±0.000 | 0 | 3.17 | ±0.006 | 0.2 | 3.37 | ±0.275 | 8.2 | 7.6 | ±5.427 | 71.4 | 73.38 | ±23.537 | 32.1 | 4.13 |
| 3 | 88.1 | ±0.172 | 0.2 | 11.9 | 0.182 | 1.5 | 0.73 | ±0.01 | 1.4 | 0 | ±0.000 | 0 | 3.34 | ±0.175 | 5.2 | 3.83 | ±0.404 | 10.5 | 4.04 | ±0.546 | 13.5 | 64.03 | ±2.109 | 3.3 | 4.65 |
| 4 | 89.3 | ±0.323 | 0.4 | 10.8 | 0.086 | 0.8 | 0.71 | ±0.061 | 8.6 | 0 | ±0.000 | 0 | 3.45 | ±0.365 | 10.6 | 3.23 | ±0.308 | 9.5 | 3.35 | ±0.842 | 25.1 | 56.21 | ±0.994 | 1.8 | 4.1 |
| 5 | 89.6 | ±0.246 | 0.3 | 10.4 | 0.335 | 3.2 | 0.62 | ±0.015 | 2.4 | 0 | ±0.000 | 0 | 5.27 | ±0.170 | 3.2 | 3.08 | ±0.534 | 17.3 | 1.39 | ±0.38 | 27.3 | 54.34 | ±3.066 | 5.6 | 4.1 |
| 6 | 87.1 | ±0.530 | 0.6 | 12.9 | 0.531 | 4.1 | 0.72 | ±0.021 | 2.9 | 0 | ±0.000 | 0 | 4.46 | ±0.085 | 1.9 | 4.67 | ±0.117 | 2.5 | 3.08 | ±0.54 | 17.5 | 72.15 | ±1.466 | 2.0 | 4.05 |
| 7 | 88.6 | ±0.068 | 0.1 | 11.4 | 0.196 | 1.7 | 0.71 | ±0.02 | 2.8 | 0 | ±0.000 | 0 | 4.3 | ±0.091 | 2.1 | 4.07 | ±0.563 | 13.8 | 2.32 | ±0.566 | 24.4 | 63.08 | ±3.165 | 5.0 | 4.1 |
| 8 | 73.9 | ±0.559 | 0.8 | 26.1 | 0.580 | 2.2 | 0.63 | ±0.01 | 1.6 | 0 | ±0.000 | 0 | 3.38 | ±0.221 | 6.5 | 6.98 | ±0.472 | 6.8 | 15.08 | ±0.795 | 5.3 | 136.63 | ±1.041 | 0.8 | 4 |
| 9 | 89.5 | ±0.168 | 0.2 | 10.5 | 0.168 | 1.6 | 0.71 | ±0.01 | 1.4 | 0 | ±0.000 | 0 | 2.83 | ±0.179 | 6.3 | 4.5 | ±0.532 | 11.8 | 2.46 | ±0.551 | 22.4 | 61.68 | ±3.306 | 5.4 | 4.05 |
| 10 | 88.9 | ±0.246 | 0.3 | 11.1 | 0.374 | 3.4 | 0.68 | ±0.025 | 3.7 | 0 | ±0.000 | 0 | 4.55 | ±0.26 | 5.7 | 3.75 | ±0.462 | 12.3 | 2.11 | ±0.418 | 19.8 | 60.39 | ±1.725 | 2.9 | 4 |
| 11 | 89.1 | ±0.260 | 0.3 | 10.9 | 0.290 | 2.7 | 0.74 | ±0.049 | 6.6 | 0 | ±0.000 | 0 | 4.07 | ±0.071 | 1.7 | 3.19 | ±0.064 | 2.0 | 2.93 | ±0.262 | 8.9 | 56.73 | ±1.493 | 2.6 | 4.1 |
| 12 | 87.4 | ±0.355 | 0.4 | 12.6 | 0.259 | 2.1 | 0.65 | ±0.025 | 3.8 | 0 | ±0.000 | 0 | 2.93 | ±0.268 | 9.1 | 3.85 | ±0.137 | 3.6 | 5.18 | ±0.347 | 6.7 | 67.1 | ±1.379 | 2.1 | 4.1 |
| Mean | 85.34 | 3.61 | 4.2 | 14.67 | 1.81 | 12.3 | 0.67 | 0.074 | 11 | 0 | 0 | 3.87 | 0.786 | 20.3 | 4.06 | 1.053 | 25.9 | 6.07 | 6.57 | 108.2 | 76.28 | 31.901 | 41.8 | 4.12 | |
| Munkoyo | |||||||||||||||||||||||||
| 1 | 95.7 | ±0.000 | 0.0 | 4.3 | 0.07 | 1.7 | 0.19 | ±0.012 | 6.3 | 0.72 | ±0.062 | 8.6 | 0.39 | ±0.045 | 11.5 | 0.56 | ±0.231 | 41.3 | 2.46 | ±0.122 | 5.0 | 16.48 | ±1.432 | 8.7 | 3.1 |
| 2 | 92.5 | ±0.457 | 0.5 | 7.5 | 0.026 | 0.4 | 0.19 | ±0.006 | 3.2 | 0.69 | ±0.023 | 3.3 | 0.79 | ±0.082 | 10.4 | 1.02 | ±0.180 | 17.6 | 4.78 | ±0.396 | 8.3 | 31.49 | ±2.184 | 6.9 | 3.05 |
| 3 | 95.9 | ±0.000 | 0.0 | 4.1 | 0.001 | 0.1 | 0.16 | ±0.025 | 15.6 | 0.85 | ±0.121 | 14.2 | 0.51 | ±0.087 | 17.1 | 0.63 | ±0.164 | 26.0 | 1.99 | ±0.025 | 1.3 | 15.67 | ±1.206 | 7.7 | 3.1 |
| 4 | 95.4 | ±0.081 | 0.1 | 4.6 | 0.116 | 2.5 | 0.14 | ±0.006 | 4.3 | 0.7 | ±0.006 | 0.9 | 0.36 | ±0.017 | 4.7 | 0.36 | ±0.092 | 25.6 | 3.08 | ±0.056 | 1.8 | 17 | ±0.730 | 4.3 | 3.2 |
| 5 | 95.8 | ±0.044 | 0.0 | 4.2 | 0.05 | 1.2 | 0.11 | ±0.012 | 10.9 | 0.67 | ±0.031 | 4.6 | 0.23 | ±0.031 | 13.5 | 0.85 | ±0.015 | 1.8 | 2.3 | ±0.040 | 1.7 | 17.8 | ±0.110 | 0.6 | 2.9 |
| 6 | 92.5 | ±0.105 | 0.1 | 7.5 | 0.148 | 2 | 0.16 | ±0.031 | 19.4 | 0.5 | ±0.006 | 1.2 | 0.49 | ±0.045 | 9.2 | 0.93 | ±0.432 | 46.5 | 5.39 | ±0.408 | 7.6 | 31.85 | ±2.423 | 7.6 | 3.35 |
| 7 | 90.2 | ±0.062 | 0.1 | 9.8 | 0.086 | 0.9 | 0.16 | ±0.021 | 7.5 | 0.68 | ±0.006 | 0.9 | 0.21 | ±0.042 | 20.0 | 0.76 | ±0.071 | 9.3 | 7.96 | ±0.021 | 0.3 | 39.52 | ±0.542 | 1.4 | 3.4 |
| 8 | 94.5 | ±0.061 | 0.1 | 5.5 | 0.084 | 1.5 | 0.1 | ±0.006 | 6.0 | 0.41 | ±0.223 | 54.4 | 0.18 | ±0.080 | 44.4 | 0.86 | ±0.108 | 12.6 | 3.96 | ±0.267 | 6.7 | 24.32 | ±1.580 | 6.5 | 3.25 |
| 9 | 89 | ±0.050 | 0.1 | 11 | 0.034 | 0.3 | 0.1 | ±0.006 | 6.0 | 0.49 | ±0.221 | 45.1 | 0.26 | ±0.006 | 2.3 | 0.75 | ±0.092 | 12.3 | 9.45 | ±0.146 | 1.5 | 45.59 | ±1.353 | 3.0 | 3.3 |
| 10 | 95.5 | ±0.296 | 0.3 | 4.5 | 0.418 | 9.4 | 0.04 | ±0.000 | 0.0 | 0.57 | ±0.107 | 18.8 | 0.15 | ±0.042 | 28.0 | 0.95 | ±0.128 | 13.5 | 2.75 | ±0.029 | 1.1 | 20.15 | ±1.391 | 6.9 | 3 |
| 11 | 94.6 | ±0.030 | 0.0 | 5.4 | 0.042 | 0.8 | 0.06 | ±0.020 | 33.3 | 0.61 | ±0.059 | 9.7 | 0.46 | ±0.099 | 21.5 | 0.74 | ±0.096 | 13.0 | 3.57 | ±0.053 | 1.5 | 22.82 | ±0.673 | 4.3 | 3.3 |
| 12 | 92.4 | ±0.355 | 0.4 | 7.6 | 0.446 | 6 | 0.11 | ±0.010 | 9.1 | 0.54 | ±0.012 | 2.2 | 0.35 | ±0.006 | 1.7 | 1.46 | ±0.070 | 4.8 | 5.14 | ±0.425 | 8.3 | 35.09 | ±1.063 | 3.0 | 2.9 |
| 13 | 94.4 | ±1.267 | 1.3 | 4.6 | 0.57 | 12.6 | 0.13 | ±0.000 | 0.0 | 0.77 | ±0.010 | 1.3 | 0.27 | ±0.012 | 4.4 | 0.72 | ±0.056 | 7.8 | 3.75 | ±1.239 | 33.0 | 22.55 | ±5.323 | 23.6 | 3.2 |
| Mean | 93.72 | 2.24 | 2.4 | 6.20 | 2.28 | 36.8 | 0.13 | 0.046 | 38.3 | 0.63 | 0.124 | 20.0 | 0.36 | 0.175 | 48.6 | 0.81 | 0.262 | 31.2 | 4.35 | 2.23 | 49.4 | 26.18 | 9.651 | 35.6 | 3.16 |
| Sample | Vitamin B1 | Vitamin B2 | Vitamin B3 | Vitamin B6 | Vitamin B12 | Calcium | Iron | Zinc | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg/100 g | SD | CV% | mg/100 g | SD | CV% | mg/100 g | SD | CV% | mg/100 g | SD | CV% | µg/100 g | SD | CV% | mg/100 g | SD | CV% | mg/100 g | SD | CV% | mg/100 g | SD | CV% | |
| Mabisi | ||||||||||||||||||||||||
| 1 | 0.062 | ±0.002 | 3.2 | 0.133 | ±0.002 | 1.5 | 0.131 | ±0.008 | 6.1 | 0.017 | ±0.001 | 5.9 | 0.327 | ±0.018 | 5.5 | 58 | ±1.989 | 1.9 | 0.625 | ±0.017 | 2.7 | 0.748 | ±0.112 | 15 |
| 2 | 0.034 | ±0.003 | 8.8 | 0.151 | ±0.002 | 1.3 | 0.455 | ±0.008 | 1.8 | 0.016 | ±0.000 | 0 | 0.429 | ±0.016 | 3.7 | 104.8 | ±8.534 | 8.1 | 0.273 | ±0.013 | 4.8 | 0.625 | ±0.049 | 7.8 |
| 3 | 0.03 | ±0.002 | 6.7 | 0.127 | ±0.002 | 1.6 | 0.222 | ±0.013 | 5.9 | 0.017 | ±0.000 | 0 | 0.372 | ±0.014 | 3.8 | 116.5 | ±6.028 | 5.2 | 0.264 | ±0.010 | 3.8 | 0.85 | ±0.102 | 12 |
| 4 | 0.072 | ±0.016 | 22.2 | 0.136 | ±0.002 | 1.5 | 0.375 | ±0.016 | 4.3 | 0.019 | ±0.001 | 5.3 | 0.475 | ±0.018 | 3.8 | 109.6 | ±2.693 | 2.5 | 0.183 | ±0.015 | 8.2 | 0.778 | ±0.047 | 6 |
| 5 | 0.044 | ±0.002 | 4.5 | 0.129 | ±0.001 | 0.8 | 0.415 | ±0.012 | 2.9 | 0.021 | ±0.003 | 14.3 | 0.411 | ±0.018 | 4.4 | 106.2 | ±5.821 | 5.5 | 0.133 | ±0.015 | 11.3 | 0.586 | ±0.039 | 6.7 |
| 6 | 0.031 | ±0.001 | 3.2 | 0.151 | ±0.002 | 1.3 | 0.566 | ±0.005 | 0.9 | 0.023 | ±0.002 | 8.7 | 0.528 | ±0.016 | 3 | 107 | ±5.534 | 5.2 | 0.029 | ±0.002 | 6.9 | 0.458 | ±0.041 | 9 |
| 7 | 0.034 | ±0.002 | 5.9 | 0.148 | ±0.002 | 1.4 | 0.261 | ±0.004 | 1.5 | 0.023 | ±0.001 | 4.3 | 0.326 | ±0.004 | 1.2 | 104.8 | ±8.794 | 8.4 | 0.105 | ±0.012 | 11.4 | 0.789 | ±0.033 | 4.2 |
| 8 | 0.024 | ±0.001 | 4.2 | 0.126 | ±0.003 | 2.4 | 0.169 | ±0.005 | 3 | 0.022 | ±0.001 | 4.5 | 0.317 | ±0.010 | 3.2 | 68.5 | ±9.668 | 14.1 | 0.15 | ±0.014 | 9.3 | 0.588 | ±0.020 | 3.4 |
| 9 | 0.034 | ±0.003 | 8.8 | 0.131 | ±0.002 | 1.5 | 0.366 | ±0.004 | 1.1 | 0.026 | ±0.001 | 3.8 | 0.339 | ±0.024 | 7.1 | 110.1 | ±16.757 | 15.2 | 0.166 | ±0.011 | 6.6 | 0.684 | ±0.032 | 4.7 |
| 10 | 0.036 | ±0.001 | 2.8 | 0.121 | ±0.002 | 1.7 | 0.24 | ±0.012 | 5 | 0.028 | ±0.001 | 3.6 | 0.342 | ±0.014 | 4.1 | 96.2 | ±5.754 | 6 | 0.113 | ±0.010 | 8.8 | 0.648 | ±0.050 | 7.7 |
| 11 | 0.023 | ±0.002 | 8.7 | 0.132 | ±0.005 | 3.8 | 0.257 | ±0.007 | 2.7 | 0.03 | ±0.001 | 3.3 | 0.342 | ±0.013 | 3.8 | 101.8 | ±8.284 | 8.1 | 0.124 | ±0.005 | 4 | 0.74 | ±0.032 | 4.3 |
| 12 | 0.054 | ±0.005 | 9.3 | 0.101 | ±0.075 | 74.3 | 0.494 | ±0.008 | 1.6 | 0.017 | ±0.001 | 5.9 | 0.52 | ±0.018 | 3.5 | 98.5 | ±2.535 | 2.6 | 0.096 | ±0.010 | 10.4 | 0.609 | ±0.026 | 4.3 |
| Mean | 0.04 | 0.015 | 38.2 | 0.132 | 0.014 | 10.6 | 0.329 | 0.136 | 41.4 | 0.022 | 0.005 | 21.1 | 0.394 | 0.077 | 19.6 | 98.5 | 17.4 | 17.7 | 0.188 | 0.154 | 81.7 | 0.675 | 0.110 | 16.4 |
| Munkoyo | ||||||||||||||||||||||||
| 1 | 0.057 | ±0.003 | 5.3 | 0.045 | ±0.002 | 4.4 | 0.34 | ±0.039 | 11.5 | 0.019 | ±0.001 | 5.3 | 0 | ±0.000 | 2.4 | ±0.344 | 14.3 | 0.067 | ±0.004 | 6 | 0.267 | ±0.002 | 0.7 | |
| 2 | 0.043 | ±0.003 | 7 | 0.031 | ±0.001 | 3.2 | 0.244 | ±0.008 | 3.3 | 0.022 | ±0.001 | 4.5 | 0 | ±0.000 | 3.9 | ±0.105 | 2.7 | 0.092 | ±0.004 | 4.3 | 0.328 | ±0.003 | 0.9 | |
| 3 | 0.026 | ±0.001 | 3.8 | 0.045 | ±0.001 | 2.2 | 0.097 | ±0.007 | 7.2 | 0.014 | ±0.001 | 7.1 | 0 | ±0.000 | 2.9 | ±0.081 | 2.8 | 0.047 | ±0.005 | 10.6 | 0.318 | ±0.002 | 0.6 | |
| 4 | 0.032 | ±0.002 | 6.3 | 0.056 | ±0.002 | 3.6 | 0.082 | ±0.004 | 4.9 | 0.016 | ±0.001 | 6.3 | 0 | ±0.000 | 5.1 | ±0.098 | 1.9 | 0.023 | ±0.002 | 8.7 | 0.167 | ±0.003 | 1.8 | |
| 5 | 0.034 | ±0.002 | 5.9 | 0.027 | ±0.001 | 3.7 | 0.24 | ±0.006 | 2.5 | 0.018 | ±0.001 | 5.6 | 0 | ±0.000 | 3 | ±0.055 | 1.8 | 0.056 | ±0.002 | 3.6 | 0.243 | ±0.003 | 1.2 | |
| 6 | 0.022 | ±0.001 | 4.5 | 0.037 | ±0.001 | 2.7 | 0.301 | ±0.022 | 7.3 | 0.01 | ±0.001 | 10 | 0 | ±0.000 | 2.7 | ±0.154 | 5.7 | 0.019 | ±0.002 | 10.5 | 0.257 | ±0.003 | 1.2 | |
| 7 | 0.022 | ±0.001 | 4.5 | 0.039 | ±0.001 | 2.6 | 0.498 | ±0.020 | 4 | 0.006 | ±0.001 | 16.7 | 0 | ±0.000 | 8.7 | ±0.706 | 8.1 | 0.013 | ±0.001 | 7.7 | 0.269 | ±0.002 | 0.7 | |
| 8 | 0.074 | ±0.002 | 2.7 | 0.028 | ±0.002 | 7.1 | 0.094 | ±0.011 | 11.7 | 0.034 | ±0.002 | 5.9 | 0 | ±0.000 | 0.9 | ±0.067 | 7.4 | 0.026 | ±0.001 | 3.8 | 0.401 | ±0.002 | 0.5 | |
| 9 | 0.013 | ±0.001 | 7.7 | 0.025 | ±0.001 | 4 | 0.36 | ±0.019 | 5.3 | 0.004 | ±0.001 | 25 | 0 | ±0.000 | 2.9 | ±0.123 | 4.2 | 0.05 | ±0.002 | 4 | 0.252 | ±0.003 | 1.2 | |
| 10 | 0.023 | ±0.001 | 4.3 | 0.036 | ±0.001 | 2.8 | 0.059 | ±0.005 | 8.5 | 0.008 | ±0.001 | 12.5 | 0 | ±0.000 | 3.3 | ±0.214 | 6.5 | 0.09 | ±0.002 | 2.2 | 0.262 | ±0.003 | 1.1 | |
| 11 | 0.068 | ±0.002 | 2.9 | 0.053 | ±0.002 | 3.8 | 0.197 | ±0.016 | 8.1 | 0.035 | ±0.002 | 5.7 | 0 | ±0.000 | 2.7 | ±0.087 | 3.2 | 0.033 | ±0.001 | 3 | 0.182 | ±0.001 | 0.5 | |
| 12 | 0.017 | ±0.002 | 11.8 | 0.032 | ±0.002 | 6.3 | 0.227 | ±0.019 | 8.4 | 0.005 | ±0.001 | 20 | 0 | ±0.000 | 6.9 | ±0.331 | 4.8 | 0.129 | ±0.003 | 2.3 | 0.326 | ±0.003 | 0.9 | |
| 13 | 0.037 | ±0.002 | 5.4 | 0.044 | ±0.002 | 4.5 | 0.235 | ±0.004 | 1.7 | 0.017 | ±0.002 | 11.8 | 0 | ±0.000 | 2.1 | ±0.133 | 6.3 | 0.058 | ±0.002 | 3.4 | 0.291 | ±0.002 | 0.7 | |
| Mean | 0.036 | 0.019 | 53.9 | 0.038 | 0.01 | 25.9 | 0.229 | 0.127 | 55.6 | 0.016 | 0.01 | 62.6 | 0 | 0 | 3.654 | 2.106 | 57.6 | 0.054 | 0.034 | 62.6 | 0.274 | 0.062 | 22.6 | |
| EAR a | Women 19–50 Years Old | %EAR Mabisi | %EAR Munkoyo |
|---|---|---|---|
| Protein b (g) | 40 | 18% | 2.0% |
| Vitamin B1 (mg) | 0.90 | 8.0% | 7.3% |
| Vitamin B2 (mg) | 0.90 | 27% | 8.0% |
| Vitamin B3 (mg) | 11 | 5.5% | 4.0% |
| Vitamin B6 (mg) | 1.1 | 4.0% | 3.0% |
| Vitamin B12 (µg) | 2.0 | 3.0% | 0% |
| Calcium (mg) | 833 | 22% | 0.80% |
| Iron c (mg) | 59 | 0.60% | 0.20% |
| Zinc d (mg) | 8.2 | 15% | 6.0% |
| Variable | Anosim R | Number of Treatment Groups | p-Value |
|---|---|---|---|
| Product and processing variables | |||
| Product type Mabisi or Munkoyo | 0.816 | 2 | <0.001 |
| Fermentation vessel used | 0.605 | 4 | <0.001 |
| Nutritional variables | |||
| Protein | 0.816 | 2 | <0.001 |
| Fat | 0.78 | 3 | <0.001 |
| Vitamin B1 | −0.0683 | 2 | 0.718 |
| Vitamin B2 | 0.532 | 3 | <0.001 |
| Vitamin B3 | −0.0374 | 2 | 0.436 |
| Vitamin B12 | 0.816 | 2 | <0.001 |
| Calcium | 0.772 | 3 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chileshe, J.; van den Heuvel, J.; Handema, R.; Zwaan, B.J.; Talsma, E.F.; Schoustra, S. Nutritional Composition and Microbial Communities of Two Non-alcoholic Traditional Fermented Beverages from Zambia: A Study of Mabisi and Munkoyo. Nutrients 2020, 12, 1628. https://doi.org/10.3390/nu12061628
Chileshe J, van den Heuvel J, Handema R, Zwaan BJ, Talsma EF, Schoustra S. Nutritional Composition and Microbial Communities of Two Non-alcoholic Traditional Fermented Beverages from Zambia: A Study of Mabisi and Munkoyo. Nutrients. 2020; 12(6):1628. https://doi.org/10.3390/nu12061628
Chicago/Turabian StyleChileshe, Justin, Joost van den Heuvel, Ray Handema, Bas J Zwaan, Elise F. Talsma, and Sijmen Schoustra. 2020. "Nutritional Composition and Microbial Communities of Two Non-alcoholic Traditional Fermented Beverages from Zambia: A Study of Mabisi and Munkoyo" Nutrients 12, no. 6: 1628. https://doi.org/10.3390/nu12061628
APA StyleChileshe, J., van den Heuvel, J., Handema, R., Zwaan, B. J., Talsma, E. F., & Schoustra, S. (2020). Nutritional Composition and Microbial Communities of Two Non-alcoholic Traditional Fermented Beverages from Zambia: A Study of Mabisi and Munkoyo. Nutrients, 12(6), 1628. https://doi.org/10.3390/nu12061628