Bovine Colostrum: Its Constituents and Uses

Abstract
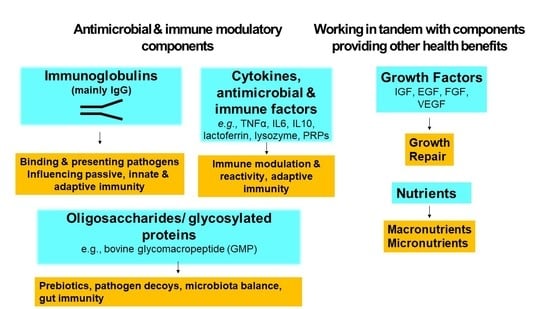
:
1. Introduction
2. Constituents of BC
2.1. Macronutrients and Micronutrients
2.1.1. Proteins and Peptides
2.1.2. Carbohydrates
2.1.3. Fats and Lipids
2.1.4. Vitamins and Minerals
2.2. Bioactive Components
2.2.1. Antimicrobial Factors
Immunoglobulins
Other Antimicrobials
2.2.2. Cytokines and Immune Regulators
Cytokines
Other Immune Regulators
2.2.3. Growth Factors
Insulin-Like Growth Factors (Somatomedins) and Their Binding Process
Epidermal Growth Factor
Transforming Growth Factor α
Transforming Growth Factor β Family
Platelet-Derived Growth Factor
Vascular Endothelial Growth Factor
Milk Fat Globule-Epidermal Growth Factor 8 (MFG-E8)
2.2.4. Hormones
Growth Hormone and Its Releasing Factor
Leptin
3. Assessment of BC for Human Use
Use of BC in Combination with Other Nutraceuticals
4. BC Use in Human and Veterinary Health
4.1. BC Therapy for Human Gastrointestinal (GI) Health and Disease
4.2. BC and Immune Function in Health and Disease
4.3. BC and Skin
4.4. BC and Bone Density
4.5. BC, Diabetes, Hypercholesterolemia, and Non-Alcoholic Fatty Liver Disease (NAFLD)
4.6. BC in Veterinary Practice
4.6.1. Use of BC in Production Animals
4.6.2. Use of BC in Canine and Equine Animals
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Panahi, Y.; Falahi, G.; Falahpour, M.; Moharamzad, Y.; Khorasgani, M.R.; Beiraghdar, F.; Naghizadeh, M.M. Bovine colostrum in the management of nonorganic failure to thrive: A randomized clinical trial. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 551–554. [Google Scholar] [CrossRef]
- Playford, R.J.; Macdonald, C.E.; Calnan, D.P.; Floyd, D.N.; Podas, T.; Johnson, W.; Wicks, A.C.; Bashir, O.; Marchbank, T. Co-administration of the health food supplement, bovine colostrum, reduces the acute non-steroidal anti-inflammatory drug-induced increase in intestinal permeability. Clin. Sci. 2001, 100, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Buckley, J.D.; Brinkworth, G.D.; Abbott, M.J. Effect of bovine colostrum on anaerobic exercise performance and plasma insulin-like growth factor I. J. Sports Sci. 2003, 21, 577–588. [Google Scholar] [CrossRef]
- Kotsis, Y.; Mikellidi, A.; Aresti, C.; Persia, E.; Sotiropoulos, A.; Panagiotakos, D.B.; Antonopoulou, S.; Nomikos, T. A low-dose, 6-week bovine colostrum supplementation maintains performance and attenuates inflammatory indices following a Loughborough Intermittent Shuttle Test in soccer players. Eur. J. Nutr. 2018, 57, 1181–1195. [Google Scholar] [CrossRef] [Green Version]
- Fenger, C.K.; Tobin, T.; Casey, P.J.; Roualdes, E.A.; Langemeier, J.L.; Haines, D.M. Bovine colostrum supplementation optimises earnings, performance and recovery in racing thoroughbreds. Comp. Exerc. Physiol. 2014, 10, 233–238. [Google Scholar] [CrossRef]
- Fenger, C.K.; Tobin, T.; Casey, P.J.; Roualdes, E.A.; Langemeier, J.L.; Cowles, R.; Haines, D.M. Enhanced Bovine Colostrum Supplementation Shortens the Duration of Respiratory Disease in Thoroughbred Yearlings. J. Equine Vet. Sci. 2016, 42, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Satyaraj, E.; Reynolds, A.; Pelker, R.; Labuda, J.; Zhang, P.; Sun, P. Supplementation of diets with bovine colostrum influences immune function in dogs. Br. J. Nutr. 2013, 110, 2216–2221. [Google Scholar] [CrossRef] [Green Version]
- Hadorn, U.; Hammon, H.; Bruckmaier, R.M.; Blum, J.W. Delaying Colostrum Intake by One Day Has Important Effects on Metabolic Traits and on Gastrointestinal and Metabolic Hormones in Neonatal Calves. J. Nutr. 1997, 127, 2011–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yvon, M.; Levieux, D.; Valluy, M.C.; Pelissier, J.P.; Mirand, P.P. Colostrum protein digestion in newborn lambs. J. Nutr. 1993, 123, 586–596. [Google Scholar] [CrossRef]
- Ginger, M.R.; Grigor, M.R. Comparative aspects of milk caseins. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1999, 124, 133–145. [Google Scholar] [CrossRef]
- Daniel, H.; Vohwinkel, M.; Rehner, G. Effect of casein and β-casomorphins on gastrointestinal motility in rats. J. Nutr. 1990, 120, 252–257. [Google Scholar] [CrossRef]
- Pessi, T.; Isolauri, E.; Sütas, Y.; Kankaanranta, H.; Moilanen, E.; Hurme, M. Suppression of T-cell activation by Lactobacillus rhamnosus GG-degraded bovine casein. Int. Immunopharmacol. 2001, 1, 211–218. [Google Scholar] [CrossRef]
- Meister, D.; Bode, J.; Shand, A.; Ghosh, S. Anti-inflammatory effects of enteral diet components on Crohn’s disease-affected tissues in vitro. Digest. Liver Dis. 2002, 34, 430–438. [Google Scholar] [CrossRef]
- Playford, R.J.; Woodman, A.C.; Vesey, D.; Deprez, P.H.; Calam, J.; Watanapa, P.; Williamson, R.C.N.; Clark, P. Effect of luminal growth factor preservation on intestinal growth. Lancet 1993, 341, 843–848. [Google Scholar] [CrossRef]
- Godden, S.M.; Lombard, J.E.; Woolums, A.R. Colostrum Management for Dairy Calves. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 535–556. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Murakawa, Y.; Ohno, M.; Ohtani, S.; Higaki, K. Gastrointestinal absorption of recombinant human insulin-like growth factor-I in rats. J. Pharmacol. Exp. Ther. 1997, 283, 611–618. [Google Scholar] [PubMed]
- Kanwar, J.; Kanwar, R.; Sun, X.; Punj, V.; Matta, H.; Morley, S.; Parratt, A.; Puri, M.; Sehgal, R. Molecular and Biotechnological Advances in Milk Proteins in Relation to Human Health. Curr. Protein Peptide Sci. 2009, 10, 308–338. [Google Scholar] [CrossRef] [PubMed]
- Bastian, S.E.P.; Dunbar, A.J.; Priebe, I.K.; Owens, P.C.; Goddard, C. Measurement of betacellulin levels in bovine serum, colostrum and milk. J. Endocrinol. 2001, 168, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuiken, K.A.; Pearson, P.B. The essential amino acid (except tryptophan) content of colostrum and milk of the cow and ewe. J. Nutr. 1949, 39, 167–176. [Google Scholar] [CrossRef]
- Matsumoto, H.; Shimokawa, Y.; Ushida, Y.; Toida, T.; Hayasawa, H. New Biological Function of Bovine α-Lactalbumin: Protective Effect against Ethanol- and Stress-induced Gastric Mucosal Injury in Rats. Biosci. Biotechnol. Biochem. 2001, 65, 1104–1111. [Google Scholar] [CrossRef]
- Nciuc, N.S.T.Ă.; Râpeanu, G. An overview of bovine α-lactalbumin stucture and functionality. Ann. Univ. Dunarea Jos Galati Fascicle VI Food Technol. 2010, 34, 82–93. [Google Scholar]
- Kiewiet, M.B.G.; Dekkers, R.; Gros, M.; van Neerven, R.J.J.; Groeneveld, A.; de Vos, P.; Faas, M.M. Toll-like receptor mediated activation is possibly involved in immunoregulating properties of cow’s milk hydrolysates. PLoS ONE 2017, 12, e0178191. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, S.I.; Jayarao, B.M.; Heinrichs, A.J. A survey of bovine colostrum composition and colostrum management practices on Pennsylvania dairy farms. J. Dairy Sci. 2007, 90, 4108–4116. [Google Scholar] [CrossRef] [Green Version]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of milk and colostrum in non-human mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef]
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 422–427. [Google Scholar] [CrossRef]
- Ten Bruggencate, S.J.; Bovee-Oudenhoven, I.M.; Feitsma, A.L.; van Hoffen, E.; Schoterman, M.H. Functional role and mechanisms of sialyllactose and other sialylated milk oligosaccharides. Nutr. Rev. 2014, 72, 377–389. [Google Scholar] [CrossRef] [Green Version]
- Zivkovic, A.M.; Barile, D. Bovine milk as a source of functional oligosaccharides for improving human health. Adv. Nutr. 2011, 2, 284–289. [Google Scholar] [CrossRef]
- Gopal, P.K.; Gill, H.S. Oligosaccharides and glycoconjugates in bovine milk and colostrum. Br. J. Nutr. 2000, 84, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Martín-Sosa, S.; Martín, M.J.; García-Pardo, L.A.; Hueso, P. Sialyloligosaccharides in human and bovine milk and in infant formulas: Variations with the progression of lactation. J. Dairy Sci. 2003, 86, 52–59. [Google Scholar] [CrossRef]
- Underwood, M.A.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Bifidobacterium longum subspecies infantis: Champion colonizer of the infant gut. Pediatr. Res. 2015, 77, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Bondue, P.; Crèvecoeur, S.; Brose, F.; Daube, G.; Seghaye, M.C.; Griffiths, M.W.; LaPointe, G.; Delcenserie, V. Cell-free spent media obtained from Bifidobacterium bifidum and Bifidobacterium crudilactis grown in media supplemented with 3′-sialyllactose modulate virulence gene expression in Escherichia coli O157: H7 and Salmonella Typhimurium. Front. Microbiol. 2016, 7, 1460. [Google Scholar] [CrossRef] [Green Version]
- O’Riordan, N.; O’Callaghan, J.; Buttò, L.F.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Bovine glycomacropeptide promotes the growth of Bifidobacterium longum ssp. infantis and modulates its gene expression. J. Dairy Sci. 2018, 101, 6730–6741. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M.; Pelizzola, V.; Monti, L.; Bruni, A.; Passolungo, L.; Abeni, F.; Degano, L. Bovine colostrum: Changes in lipid constituents in the first 5 days after parturition. J. Dairy Sci. 2014, 97, 5065–5072. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, T.F.; O’Donovan, M.; Murphy, J.P.; Sugrue, K.; Mannion, D.; McCarthy, W.P.; Timlin, M.; Kilcawley, K.N.; Hickey, R.M.; Tobin, J.T. Evolution of the bovine milk fatty acid profile—From colostrum to milk five days post parturition. Int. Dairy J. 2020, 104, 8721–8731. [Google Scholar] [CrossRef]
- Verardo, V.; Gómez-Caravaca, A.M.; Arráez-Román, D.; Hettinga, K. Recent advances in phospholipids from colostrum, milk and dairy by-products. Int. J. Mol. Sci. 2017, 18, 173. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.A.; Calder, P.C. The influence of the position of palmitate in infant formula triacylglycerols on health outcomes. Nutr. Res. 2017, 44, 1–8. [Google Scholar] [CrossRef]
- Bar-Yoseph, F.; Lifshitz, Y.; Cohen, T. Review of sn-2 palmitate oil implications for infant health. Prostaglandins Leukot. Essent. Fatty Acids 2013, 89, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Sales-Campos, H.; Reis de Souza, P.; Crema Peghini, B.; Santana da Silva, J.; Ribeiro Cardoso, C. An Overview of the Modulatory Effects of Oleic Acid in Health and Disease. Mini Rev. Med. Chem. 2013, 13, 201–210. [Google Scholar]
- Cleminson, J.S.; Zalewski, S.P.; Embleton, N.D. Nutrition in the preterm infant: What’s new? Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 220–225. [Google Scholar]
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef]
- Bagwe, S.; Tharappel, L.J.P.; Kaur, G.; Buttar, H.S. Bovine colostrum: An emerging nutraceutical. J. Complement. Integr. Med. 2015, 12, 175–185. [Google Scholar] [CrossRef]
- Bagwe-Parab, S.; Yadav, P.; Kaur, G.; Tuli, H.S.; Buttar, H.S. Therapeutic Applications of Human and Bovine Colostrum in the Treatment of Gastrointestinal Diseases and Distinctive Cancer Types: The Current Evidence. Front. Pharmacol. 2020, 11, 1100. [Google Scholar] [CrossRef]
- Pakkanen, R.; Aalto, J. Growth factors and antimicrobial factors of bovine colostrum. Int. Dairy J. 1997, 7, 285–297. [Google Scholar] [CrossRef]
- Kelly, G.S. Bovine colostrum: A review of clinical uses. Altern. Med. Rev. 2003, 8, 378–394. [Google Scholar]
- Stelwagen, K.; Carpenter, E.; Haigh, B.; Hodgkinson, A.; Wheeler, T.T. Immune components of bovine colostrum and milk. J. Anim. Sci. 2009, 87, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Ulfman, L.H.; Leusen, J.H.W.; Savelkoul, H.F.J.; Warner, J.O.; van Neerven, R.J.J. Effects of Bovine Immunoglobulins on Immune Function, Allergy, and Infection. Front. Nutr. 2018, 5, 1–20. [Google Scholar] [CrossRef]
- Tacket, C.O.; Losonsky, G.; Link, H.; Hoang, Y.; Guesry, P.; Hilpert, H.; Levine, M.M. Protection by Milk Immunoglobulin Concentrate against Oral Challenge with Enterotoxigenic Escherichia coli. N. Engl. J. Med. 1988, 318, 1240–1243. [Google Scholar] [CrossRef]
- Ebina, T.; Ohta, M.; Kanamaru, Y.; Yamamoto-Osumi, Y.; Baba, K. Passive immunizations of suckling mice and infants with bovine colostrum containing antibodies to human rotavirus. J. Med. Virol. 1992, 38, 117–123. [Google Scholar] [CrossRef]
- Hodgkinson, A.J.; Cannon, R.D.; Holmes, A.R.; Fischer, F.J.; Willix-Payne, D.J. Production from dairy cows of semi-industrial quantities of milk-protein concentrate (MPC) containing efficacious anti-Candida albicans IgA antibodies. J. Dairy Res. 2007, 74, 269–275. [Google Scholar] [CrossRef]
- Loimaranta, V.; Laine, M.; Söderling, E.; Vasara, E.; Rokka, S.; Marnila, P.; Korhonen, H.; Tossavainen, O.; Tenovuo, J. Effects of bovine immune and non-immune whey preparations on the composition and pH response of human dental plaque. Eur. J. Oral Sci. 1999, 107, 244–250. [Google Scholar] [CrossRef]
- Wilson, D.; Evans, M.; Weaver, E.; Shaw, A.L.; Klein, G.L. Evaluation of serum-derived bovine immunoglobulin protein isolate in subjects with diarrhea-predominant irritable bowel syndrome. Clin. Med. Insights Gastroenterol. 2013, 6, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Morrin, S.T.; Lane, J.A.; Marotta, M.; Bode, L.; Carrington, S.D.; Irwin, J.A.; Hickey, R.M. Bovine colostrum-driven modulation of intestinal epithelial cells for increased commensal colonisation. Appl. Microbiol. Biotechnol. 2019, 103, 2745–2758. [Google Scholar] [CrossRef] [PubMed]
- Douëllou, T.; Montel, M.C.; Thevenot Sergentet, D. Invited review: Anti-adhesive properties of bovine oligosaccharides and bovine milk fat globule membrane-associated glycoconjugates against bacterial food enteropathogens. J. Dairy Sci. 2017, 100, 3348–3359. [Google Scholar] [CrossRef] [PubMed]
- Seifu, E.; Buys, E.M.; Donkin, E.F. Significance of the lactoperoxidase system in the dairy industry and its potential applications: A review. Trends Food Sci. Technol. 2005, 16, 137–145. [Google Scholar] [CrossRef]
- Wheeler, T.T.; Hodgkinson, A.J.; Prosser, C.G.; Davis, S.R. Immune components of colostrum and milk—A historical perspective. J. Mammary Gland Biol. Neoplasia 2007, 12, 237–247. [Google Scholar] [CrossRef]
- Clare, D.; Catignani, G.; Swaisgood, H. Biodefense Properties of Milk: The Role of Antimicrobial Proteins and Peptides. Curr. Pharm. Des. 2005, 9, 1239–1255. [Google Scholar] [CrossRef]
- Lönnerdal, B. Nutritional and physiologic significance of human milk proteins. Am. J. Clin. Nutr. 2003, 77, 1537S–1543S. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F. Lactoferrin in milk from different species. Comparat. Biochem. Physiol. Part B Biochem. Physiol. 1971, 39, IN11–IN13. [Google Scholar] [CrossRef]
- Sanchez, L.; Aranda, P.; Perez, M.; Calvo, M. Concentration of Lactoferrin and Transferrin throughout Lactation in Cow’s Colostrum and Milk. Biol. Chem. Hoppe-Seyler 1988, 369, 1005–1008. [Google Scholar] [CrossRef]
- Arnold, R.R.; Brewer, M.; Gauthier, J.J. Bactericidal activity of human lactoferrin: Sensitivity of a variety of microorganisms. Infect. Immun. 1980, 28, 893–898. [Google Scholar]
- Aisen, P.; Listowsky, I. Iron transport and storage proteins. Ann. Rev. Biochem. 1980, 49, 357–393. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xu, X.-X.; Liu, Y.; Xi, E.-Z.; An, J.-J.; Tabys, D.; Liu, N. The In Vitro Protective Role of Bovine Lactoferrin on Intestinal Epithelial Barrier. Molecules 2019, 24, 148. [Google Scholar] [CrossRef] [PubMed]
- King, J.C.; Cummings, G.E.; Guo, N.; Trivedi, L.; Readmond, B.X.; Keane, V.; Feigelman, S.; de Waard, R. A double-blind, placebo-controlled, pilot study of bovine lactoferrin supplementation in bottle-fed infants. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa, T.J.; Chea-Woo, E.; Campos, M.; Pecho, I.; Prada, A.; McMahon, R.J.; Cleary, T.G. Impact of lactoferrin supplementation on growth and prevalence of Giardia colonization in children. Clin. Infect. Dis. 2008, 46, 1881–1883. [Google Scholar] [CrossRef] [Green Version]
- Kruzel, M.L.; Bacsi, A.; Choudhury, B.; Sur, S.; Boldogh, I. Lactoferrin decreases pollen antigen-induced allergic airway inflammation in a murine model of asthma. Immunology 2006, 119, 159–166. [Google Scholar] [CrossRef]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 3, CD007137. [Google Scholar] [CrossRef]
- Tacket, C.O.; Binion, S.B.; Bostwick, E.; Losonsky, G.; Roy, M.J.; Edelman, R. Efficacy of bovine milk immunoglobulin concentrate in preventing illness after Shigella flexneri challenge. Am. J. Trop. Med. Hyg. 1992, 47, 276–283. [Google Scholar] [CrossRef]
- Jones, A.W.; March, D.S.; Thatcher, R.; Diment, B.; Walsh, N.P.; Davison, G. The effects of bovine colostrum supplementation on in vivo immunity following prolonged exercise: A randomised controlled trial. Eur. J. Nutr. 2019, 58, 335–344. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Tuomola, E.; Arvilommi, H.; Salminen, S. Modulation of human humoral immune response through orally administered bovine colostrum. FEMS Immunol. Med. Microbiol. 2001, 31, 93–96. [Google Scholar] [CrossRef]
- Dinarello, C.A. The interleukin-1 family: 10 years of discovery. FASEB J. 1994, 8, 1314–1325. [Google Scholar] [CrossRef] [Green Version]
- Rathe, M.; Müller, K.; Sangild, P.T.; Husby, S. Clinical applications of bovine colostrum therapy: A systematic review. Nutr. Rev. 2014, 72, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, M.J.; Cheon, J.H.; Kim, S.W.; Park, J.J.; Moon, C.M.; Han, S.Y.; Kim, E.S.; Kim, T.I.; Kim, W.H. Bovine colostrum inhibits nuclear factor κB-mediated proinflammatory cytokine expression in intestinal epithelial cells. Nutr. Res. 2009, 29, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.J.; Byron, K.; Gibson, P.R. Interleukin-8 stimulates the migration of human colonic epithelial cells in vitro. Clin. Sci. 1999, 97, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Barrington, G.M.; Besser, T.E.; Davis, W.C.; Gay, C.C.; Reeves, J.J.; McFadden, T.B. Expression of Immunoglobulin G1 Receptors by Bovine Mammary Epithelial Cells and Mammary Leukocytes. J. Dairy Sci. 1997, 80, 86–93. [Google Scholar] [CrossRef]
- Van Hese, K.; Goossens, L.; Vandaele, G. Opsomer, Invited review: MicroRNAs in bovine colostrum—Focus on their origin and potential health benefits for the calf. J. Dairy Sci. 2020, 103, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Beshay, E.; Prud’homme, G.J. Towards an understanding of biological role of colostrinin peptides. J. Mol. Neurosci. 2001, 17, 379–389. [Google Scholar]
- Rattray, M. Technology evaluation: Colostrinin, ReGen. Curr. Opin. Mol. Ther. 2005, 7, 78–84. [Google Scholar]
- Zabłocka, A.; Sokołowska, A.; Macała, J.; Bartoszewska, M.; Mitkiewicz, M.; Janusz, M.; Wilusz, T.; Polanowski, A. Colostral Proline-Rich Polypeptide Complexes. Comparative Study of the Antioxidant Properties, Cytokine-Inducing Activity, and Nitric Oxide Release of Preparations Produced by a Laboratory and a Large-Scale Method. Int. J. Peptide Res. Ther. 2020, 26, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Boldogh, I.; Aguilera-Aguirre, L.; Bacsi, A.; Choudhury, B.K.; Saavedra-Molina, A.; Kruzel, M. Colostrinin decreases hypersensitivity and allergic responses to common allergens. Int. Arch. Allergy Immunol. 2008, 146, 289–306. [Google Scholar] [CrossRef]
- Ragab, S.S.; Ibrahim, M.K. Role of Bovine Colostrum and its Biofunctional Fraction PRP in Oral Treatment of Enterogenic Endotoxaemia in Rats. Int. J. Agric. Biol. 2004, 6, 576–580. [Google Scholar]
- Hagiwara, T.; Shinoda, I.; Fukuwatari, Y.; Shimamura, S. Effects of lactoferrin and its peptides on proliferation of rat intestinal epithelial cell line, IEC-18, in the presence of epidermal growth factor. Biosci. Biotechnol. Biochem. 1995, 59, 1875–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
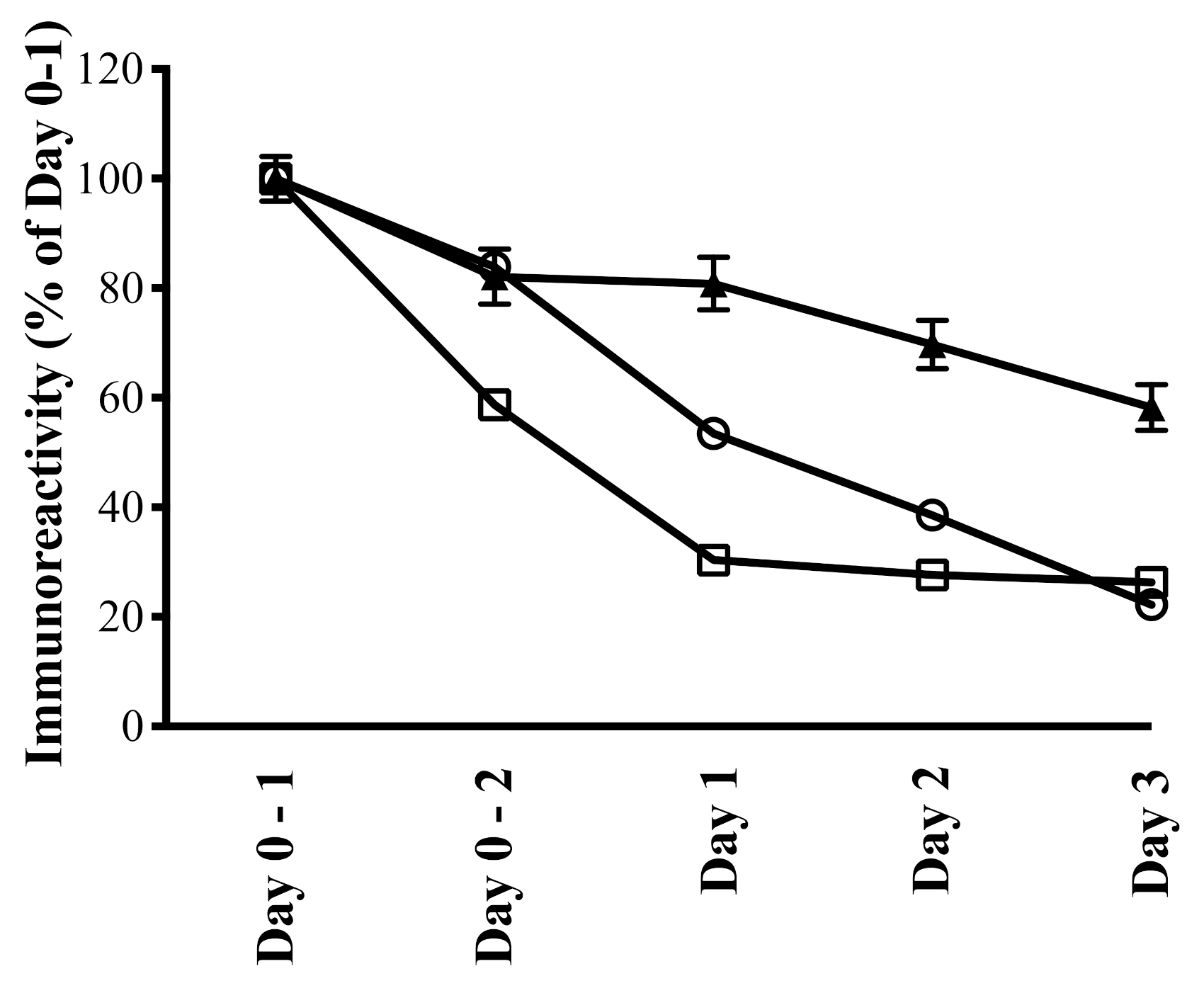
- Playford, R.J.; Cattell, M.; Marchbank, T. Marked variability in bioactivity between commercially available bovine colostrum for human use; implications for clinical trials. PLoS ONE 2020, 15, e0240392. [Google Scholar]
- Daughaday, W.H.; Rotwein, P.; Rotwein, P. Insulin-like growth factors I and II. Peptide, messenger ribonucleic acid and gene structures, serum, and tissue concentrations. Endocr. Rev. 1989, 10, 68–91. [Google Scholar] [CrossRef] [PubMed]
- Lund, P.K.; Zimmermann, E.M. Insulin-like growth factors and inflammatory bowel disease. Bailliere’s Clin. Gastroenterol. 1996, 10, 83–96. [Google Scholar] [CrossRef]
- Baxter, R.C.; Zaltsman, Z.; Turtle, J.R. Immunoreactive somatomedin-c/insulin-like growth factor I and its binding protein in human milk. J. Clin. Endocrinol. Metab. 1984, 58, 955–959. [Google Scholar] [CrossRef]
- Vacher, P.Y.; Blum, J.W. Age dependency of insulin like growth factor 1, insulin protein and immunoglobulin concentrations and gamma glutamyl transferase activity in first colostrum of dairy cows. Milchwissenschaft 1993, 48, 423–425. [Google Scholar]
- Collier, R.J.; Miller, M.A.; Hildebrandt, J.R.; Torkelson, A.R.; White, T.C.; Madsen, K.S.; Vicini, J.L.; Eppard, P.J.; Lanza, G.M. Factors Affecting Insulin-Like Growth Factor-I Concentration in Bovine Milk. J. Dairy Sci. 1991, 74, 2905–2911. [Google Scholar] [CrossRef]
- Lowe, W.L. Biological actions of the insulin-like growth factors. In Insulin-like Growth Factors, Molecular and Cellular Aspects; LeRoith, D., Ed.; CRC Press: Boca Raton, FL, USA, 1991; pp. 49–85. [Google Scholar]
- Lo, H.C.; Hinton, P.S.; Yang, H.; Unterman, T.G.; Ney, D.M. Insulin-like growth factor-I but not growth hormone attenuates dexamethasone-induced catabolism in parenterally fed rats. J. Parenter. Enter. Nutr. 1996, 20, 171–177. [Google Scholar] [CrossRef]
- Schams, D.; Einspanier, R. Growth hormone, IGF-I and insulin in mammary gland secretion before and after parturition and possibility of their transfer into the calf. Endocr. Regul. 1991, 25, 139–143. [Google Scholar]
- Yan, J.; Charles, J.F. Gut Microbiota and IGF-1. Calcif. Tissue Int. 2018, 102, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Heidegger, I.; Ofer, P.; Doppler, W.; Rotter, V.; Klocker, H.; Massoner, P. Diverse functions of IGF/insulin signaling in malignant and noncancerous prostate cells: Proliferation in cancer cells and differentiation in noncancerous cells. Endocrinology 2012, 153, 4633–4643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, G.; Jones, A.W.; Marchbank, T.; Playford, R.J. Oral bovine colostrum supplementation does not increase circulating insulin-like growth factor-1 concentration in healthy adults: Results from short- and long-term administration studies. Eur. J. Nutr. 2019, 59, 1473–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, M.R.; Brent Polk, D. ErbB receptors and their growth factor ligands in pediatric intestinal inflammation. Pediatr. Res. 2014, 75, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koldovsky, O.; Britton, J.; Davis, D.; Davis, T.; Grimes, J.; Kong, W.; Rao, R.; Schaudies, P. The developing gastrointestinal tract and milk-borne epidermal growth factor. Adv. Exp. Med. Biol. 1991, 310, 99–105. [Google Scholar] [PubMed]
- Playford, R.J.; Marchbank, T.; Calnan, D.P.; Calam, J.; Royston, P.; Batten, J.J.; Hansen, H.F. Epidermal growth factor is digested to smaller, less active forms in acidic gastric juice. Gastroenterology 1995, 108, 92–101. [Google Scholar] [CrossRef]
- Playford, R.J.; Hanby, A.M.; Gschmeissner, S.; Peiffer, L.P.; Wright, N.A.; McGarrity, T. The epidermal growth factor receptor (EGF-R) is present on the basolateral, but not the apical, surface of enterocytes in the human gastrointestinal tract. Gut 1996, 39, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Playford, R.J. Peptides and gastrointestinal mucosal integrity. Gut 1995, 37, 595–597. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.F.; van den Berg, M.; Stokkers, P.C.F. Developmental regulation of epidermal growth factor receptor kinase in rat intestine. Gastroenterology 1994, 107, 1278–1287. [Google Scholar] [CrossRef]
- Okuyama, H.; Urao, M.; Lee, D.; Drongowski, R.A.; Coran, A.G. The effect of epidermal growth factor on bacterial translocation in newborn rabbits. Proc. J. Pediatr. Surg. 1998, 33, 225–228. [Google Scholar] [CrossRef]
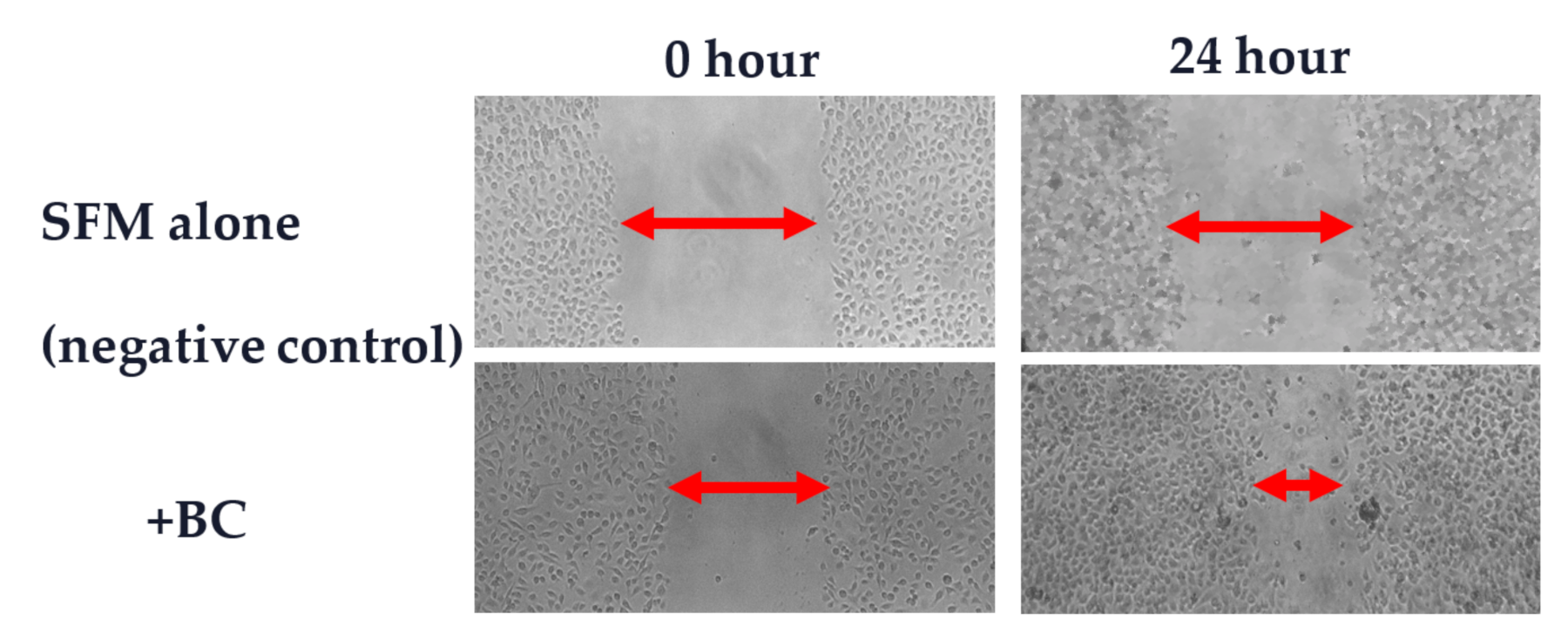
- Yu, M.; Kim, J.; Ahn, J.H.; Moon, Y. Nononcogenic restoration of the intestinal barrier by E. coli-delivered human EGF. JCI Insight 2019, 4, e125166. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Ohmura, E.; Kamiya, Y.; Murakami, H.; Onoda, N.; Iwashita, M.; Wakai, K.; Tsushima, T.; Shizume, K. Transforming growth factor (TGF)—α in human milk. Life Sci. 1991, 48, 1151–1156. [Google Scholar] [CrossRef]
- Cartlidge, S.A.; Elder, J.B. Transforming growth factor α and epidermal growth factor levels in normal human gastrointestinal mucosa. Br. J. Cancer 1989, 60, 657–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, J.A.; Daniel Beauchamp, R.; Russell, W.E.; Dubois, R.N.; Coffey, R.J. Epidermal growth factor-related peptides and their relevance to gastrointestinal pathophysiology. Gastroenterology 1995, 108, 564–580. [Google Scholar] [CrossRef]
- Dignass, A.U.; Podolsky, D.K. Cytokine modulation of intestinal epithelial cell restitution: Central role of transforming growth factor β. Gastroenterology 1993, 105, 1323–1332. [Google Scholar] [CrossRef]
- Playford, R.J.; Floyd, D.N.; Macdonald, C.E.; Calnan, D.P.; Adenekan, R.O.; Johnson, W.; Goodlad, R.A.; Marchbank, T. Bovine colostrum is a health food supplement which prevents NSAID induced gut damage. Gut 1999, 44, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Walker, A. Breast Milk as the Gold Standard for Protective Nutrients. J. Pediatr. 2010, 156, S3–S7. [Google Scholar] [CrossRef]
- McEntee, C.P.; Gunaltay, S.; Travis, M.A. Regulation of barrier immunity and homeostasis by integrin-mediated transforming growth factor β activation. Immunology 2020, 160, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Shing, Y.; Davidson, S.; Klagsbrun, M. Purification of Polypeptide Growth Factors from Milk. Methods Enzymol. 1987, 146, 42–48. [Google Scholar]
- Shing, Y.W.; Klagsbrun, M. Human and bovine milk contain different sets of growth factors. Endocrinology 1984, 115, 273–282. [Google Scholar] [CrossRef]
- Shing, Y.; Klagsbrun, M. Purification and characterization of a bovine colostrum-derived growth factor. Mol. Endocrinol. 1987, 1, 335–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keck, P.J.; Hauser, S.D.; Krivi, G.; Sanzo, K.; Warren, T.; Feder, J.; Connolly, D.T. Vascular permeability factor, an endothelial cell mitogen related to PDGF. Science 1989, 246, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Siafakas, C.G.; Anatolitou, F.; Fusunyan, R.D.; Walker, W.A.; Sanderson, I.R. Vascular endothelial growth factor (VEGF) is present in human breast milk and its receptor is present on intestinal epithelial cells. Pediatr. Res. 1999, 45, 652–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterton, D.E.W.; Nguyen, D.N.; Bering, S.B.; Sangild, P.T. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int. J. Biochem. Cell Biol. 2013, 45, 1730–1747. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.S. Functional role of milk fat globule-epidermal growth factor VIII in macrophage-mediated inflammatory responses and inflammatory/autoimmune diseases. Mediat. Inflamm. 2016, 2016, 5628486. [Google Scholar] [CrossRef] [Green Version]
- Baram, T.; Koch, Y.; Hazum, E.; Fridkin, M. Gonadotropin-releasing hormone in milk. Science 1977, 198, 300–302. [Google Scholar] [CrossRef] [Green Version]
- Amit, T.; Dibner, C.; Barkey, R.J. Characterization of prolactin- and growth hormone-binding proteins in milk and their diversity among species. Mol. Cell. Endocrinol. 1997, 130, 167–180. [Google Scholar] [CrossRef]
- Grosvenor, C.E.; Picciano, M.F.; Baumrucker, C.R. Hormones and growth factors in milk. Endocr. Rev. 1993, 14, 710–728. [Google Scholar] [CrossRef]
- Ulshen, M.H.; Dowling, R.H.; Fuller, C.R.; Zimmermann, E.M.; Lund, P.K. Enhanced growth of small bowel in transgenic mice overexpressing bovine growth hormone. Gastroenterology 1993, 104, 973–980. [Google Scholar] [CrossRef]
- Delehaye-Zervas, M.C.; Mertani, H.; Martini, J.F.; Nihoul-Feketé, C.; Morel, G.; Postel-Vinay, M.C. Expression of the growth hormone receptor gene in human digestive tissues. J. Clin. Endocrinol. Metab. 1994, 78, 1473–1480. [Google Scholar]
- Jensen, E.A.; Young, J.A.; Mathes, S.C.; List, E.O.; Carroll, R.K.; Kuhn, J.; Onusko, M.; Kopchick, J.J.; Murphy, E.R.; Berryman, D.E. Crosstalk between the growth hormone/insulin-like growth factor-1 axis and the gut microbiome: A new frontier for microbial endocrinology. Growth Horm. IGF Res. 2020, 53–54, 101333. [Google Scholar] [CrossRef] [PubMed]
- Pinotti, L.; Rosi, F. Leptin in bovine colostrum and milk. Horm. Metab. Res. 2006, 38, 89–93. [Google Scholar] [CrossRef]
- Hedbacker, K.; Birsoy, K.; Wysocki, R.W.; Asilmaz, E.; Ahima, R.S.; Farooqi, I.S.; Friedman, J.M. Antidiabetic Effects of IGFBP2, a Leptin-Regulated Gene. Cell Metab. 2010, 11, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterton, D.E.W.; Aagaard, S.; Hesselballe Hansen, T.; Nguyen, D.N.; de Gobba, C.; Lametsch, R.; Sangild, P.T. Bioactive proteins in bovine colostrum and effects of heating, drying and irradiation. Food Funct. 2020, 11, 2309–2327. [Google Scholar] [CrossRef] [PubMed]
- McGuirk, S.M.; Collins, M. Managing the production, storage, and delivery of colostrum. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 593–603. [Google Scholar] [CrossRef]
- Morrill, K.M.; Conrad, E.; Lago, A.; Campbell, J.; Quigley, J.; Tyler, H. Nationwide evaluation of quality and composition of colostrum on dairy farms in the United States. J. Dairy Sci. 2012, 95, 3997–4005. [Google Scholar] [CrossRef] [Green Version]
- McGrath, B.A.; Fox, P.F.; McSweeney, P.L.H.; Kelly, A.L. Composition and properties of bovine colostrum: A review. Dairy Sci. Technol. 2016, 96, 133–158. [Google Scholar] [CrossRef]
- Hałasa, M.; Maciejewska-Markiewicz, D.; Baśkiewicz-Hałasa, M.; Safranow, K.; Stachowska, E. Post-Delivery Milking Delay Influence on the Effect of Oral Supplementation with Bovine Colostrum as Measured with Intestinal Permeability Test. Medicina 2020, 56, 495. [Google Scholar] [CrossRef]
- Davison, G.; Marchbank, T.; March, D.S.; Thatcher, R.; Playford, R.J. Zinc carnosine works with bovine colostrum in truncating heavy exercise-induced increase in gut permeability in healthy volunteers. Am. J. Clin. Nutr. 2016, 104, 526–536. [Google Scholar] [CrossRef] [Green Version]
- Playford, R.J.; Garbowsky, M.; Marchbank, T. Pasteurized Chicken Egg Powder Stimulates Proliferation and Migration of AGS, RIE1, and Caco-2 Cells and Reduces NSAID-Induced Injury in Mice and Colitis in Rats. J. Nutr. 2020, 150, 1434–1442. [Google Scholar] [CrossRef]
- Gleeson, M. Immunological aspects of sport nutrition. Immunol. Cell Biol. 2016, 94, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathe, M.; de Pietri, S.; Wehner, P.S.; Frandsen, T.L.; Grell, K.; Schmiegelow, K.; Sangild, P.T.; Husby, S.; Müller, K. Bovine Colostrum Against Chemotherapy-Induced Gastrointestinal Toxicity in Children With Acute Lymphoblastic Leukemia: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Parenter. Enter. Nutr. 2019, 2, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Filipescu, I.E.; Leonardi, L.; Menchetti, L.; Guelfi, G.; Traina, G.; Casagrande-Proietti, P.; Piro, F.; Quattrone, A.; Barbato, O.; Brecchia, G. Preventive effects of bovine colostrum supplementation in TNBS-induced colitis in mice. PLoS ONE 2018, 13, e0202929. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Macdonald, C.; Wicks, A.C.; Holt, M.P.; Floyd, D.; Ghosh, S.; Wright, N.A.; Playford, R.J. Use of the “nutriceutical”, bovine colostrum, for the treatment of distal colitis: Results from an initial study. Aliment. Pharmacol. Ther. 2002, 16, 1917–1922. [Google Scholar] [CrossRef]
- Hutton, M.L.; Cunningham, B.A.; Mackin, K.E.; Lyon, S.A.; James, M.L.; Rood, J.I.; Lyras, D. Bovine antibodies targeting primary and recurrent Clostridium difficile disease are a potent antibiotic alternative. Sci. Rep. 2017, 7, 5–9. [Google Scholar] [CrossRef]
- Gritz Emily, C.; Bhandari, V. The human neonatal gut microbiome: A brief review. Front. Pediatr. 2015, 3, 17. [Google Scholar]
- Sun, J.; Li, Y.; Pan, X.; Nguyen, D.N.; Brunse, A.; Bojesen, A.M.; Rudloff, S.; Mortensen, M.S.; Burrin, D.G.; Sangild, P.T. Human Milk Fortification with Bovine Colostrum Is Superior to Formula-Based Fortifiers to Prevent Gut Dysfunction, Necrotizing Enterocolitis, and Systemic Infection in Preterm Pigs. J. Parenter. Enter. Nutr. 2019, 43, 252–262. [Google Scholar] [CrossRef]
- Bierut, T.; Duckworth, L.; Grabowsky, M.; Ordiz, M.I.; Laury, M.L.; Callaghan-Gillespie, M.; Maleta, K.; Manary, M.J. The effect of bovine colostrum/egg supplementation compared with corn/soy flour in young Malawian children: A randomized, controlled clinical trial. Am. J. Clin. Nutr. 2020, nqaa325. [Google Scholar] [CrossRef]
- Hałasa, M.; Maciejewska, D.; Baśkiewicz-Hałasa, M.; Machaliński, B.; Safranow, K.; Stachowska, E. Oral Supplementation with Bovine Colostrum Decreases Intestinal Permeability and Stool Concentrations of Zonulin in Athletes. Nutrients 2017, 9, 370. [Google Scholar] [CrossRef]
- Cesarone, M.R.; Belcaro, G.; di Renzo, A.; Dugall, M.; Cacchio, M.; Ruffini, I.; Pellegrini, L.; del Boccio, G.; Fano, F.; Ledda, A.; et al. Prevention of influenza episodes with colostrum compared with vaccination in healthy and high-risk cardiovascular subjects: The epidemiologic study in San Valentino. Clin. Appl. Thromb. Hemost. 2007, 13, 130–136. [Google Scholar] [CrossRef]
- Patel, K.; Rana, R. Pedimune in recurrent respiratory infection and diarrhoea- The Indian experience-the pride study. Indian J. Pediatr. 2006, 73, 585–591. [Google Scholar] [CrossRef]
- Saad, K.; Abo-Elela, M.G.M.; El-Baseer, K.A.A.; Ahmed, A.E.; Ahmad, F.A.; Tawfeek, M.S.K.; El-Houfey, A.A.; Khair, M.D.A.; Abdel-Salam, A.M.; Abo-Elgheit, A.; et al. Effects of bovine colostrum on recurrent respiratory tract infections and diarrhea in children. Medicine 2016, 95, 4–8. [Google Scholar] [CrossRef]
- Nederend, M.; van Stigt, A.H.; Jansen, J.H.M.; Jacobino, S.R.; Brugman, S.; de Haan, C.A.M.; Bont, L.J.; van Neerven, R.J.J.; Leusen, J.H.W. Bovine IgG Prevents Experimental Infection With RSV and Facilitates Human T Cell Responses to RSV. Front. Immunol. 2020, 11, 1701. [Google Scholar] [CrossRef]
- Jones, A.W.; March, D.S.; Curtis, F.; Bridle, C. Bovine colostrum supplementation and upper respiratory symptoms during exercise training: A systematic review and meta-analysis of randomised controlled trials. BMC Sports Sci. Med. Rehabil. 2016, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, D.; Maresca, V.; Flori, E.; Mastrofrancesco, A.; Picardo, M.; Cardinali, G. Bovine colostrum induces the differentiation of human primary keratinocytes. FASEB J. 2020, 34, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Hartog, A.; Leenders, I.; van der Kraan, P.M.; Garssen, J. Anti-inflammatory effects of orally ingested lactoferrin and glycine in different zymosan-induced inflammation models: Evidence for synergistic activity. Int. Immunopharmacol. 2007, 7, 1784–1792. [Google Scholar] [CrossRef]
- Hong, J.P.; Park, S.W. The combined effect of recombinant human epidermal growth factor and erythropoietin on full-thickness wound healing in diabetic rat model. Int. Wound J. 2014, 11, 373–378. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Cumberbatch, M.; Tucker, S.C.; Dearman, R.J.; Andrew, S.; Headon, D.R.; Kimber, I. Exogenous topical lactoferrin inhibits allergen-induced Langerhans cell migration and cutaneous inflammation in humans. Br. J. Dermatol. 2001, 144, 715–725. [Google Scholar] [CrossRef]
- Lee, J.; Kwon, S.H.; Kim, H.M.; Fahey, S.N.; Knighton, D.R.; Sansom, A. Effect of a growth protein-colostrum fraction* on bone development in juvenile rats. Biosci. Biotechnol. Biochem. 2008, 72, 1–6. [Google Scholar] [CrossRef]
- Duff, W.R.D.; Chilibeck, P.D.; Rooke, J.J.; Kaviani, M.; Krentz, J.R.; Haines, D.M. The effect of bovine colostrum supplementation in older adults during resistance training. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 276–285. [Google Scholar] [CrossRef]
- Hwang, K.A.; Hwang, Y.J.; Ha, W.; Choo, Y.K.; Ko, K. Oral administration of insulin-like growth factor-I from colostral whey reduces blood glucose in streptozotocin-induced diabetic mice. Br. J. Nutr. 2012, 108, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Jung, W.S.; Choi, N.J.; Kim, D.O.; Shin, D.H.; Kim, Y.J. Health-promoting effects of bovine colostrum in Type 2 diabetic patients can reduce blood glucose, cholesterol, triglyceride and ketones. J. Nutr. Biochem. 2009, 20, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Golay, A.; Ferrara, J.M.; Felber, J.P.; Schneider, H. Cholesterol-lowering effect of skim milk from immunized cows in hypercholesterolemic patients. Am. J. Clin. Nutr. 1990, 52, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, M.; Shabat, Y.; Ben Ya’acov, A.; Lalazar, G.; Adar, T.; Wong, V.; Muller, B.; Rawlin, G.; Ilan, Y. Alleviation of insulin resistance and liver damage by oral administration of Imm124-E is mediated by increased Tregs and associated with increased serum GLP-1 and adiponectin: Results of a phase I/II clinical trial in NASH. J. Inflamm. Res. 2012, 5, 141–150. [Google Scholar]
- Adar, T.; Ben Ya’acov, A.; Lalazar, G.; Lichtenstein, Y.; Nahman, D.; Mizrahi, M.; Wong, V.; Muller, B.; Rawlin, G.; Ilan, Y. Oral administration of immunoglobulin G-enhanced colostrum alleviates insulin resistance and liver injury and is associated with alterations in natural killer T cells. Clin. Exp. Immunol. 2012, 167, 252–260. [Google Scholar] [CrossRef]
- Faber, S.N.; Faber, N.E.; Mccauley, T.C.; Ax, R.L. Case Study: Effects of Colostrum Ingestion on Lactational Performance. Profess. Anim. Sci. 2005, 21, 420–425. [Google Scholar] [CrossRef]
- Wells, S.J.; Dargatz, D.A.; Ott, S.L. Factors associated with mortality to 21 days of life in dairy heifers in the United States. Prev. Vet. Med. 1996, 29, 9–19. [Google Scholar] [CrossRef]
- Boudry, C. Bovine colostrum as a natural growth promoter for newly weaned piglets: A review. Anim. Sci. Biotechnol. 2008, 66, 1–2. [Google Scholar]
- King, M.R.; Morel, P.C.H.; Pluske, J.R.; Hendriks, W.H. A comparison of the effects of dietary spray-dried bovine colostrum and animal plasma on growth and intestinal histology in weaner pigs. Livest. Sci. 2008, 119, 167–173. [Google Scholar] [CrossRef]
- Huërou-Luron, I.L.; Huguet, A.; Callarec, J.; Leroux, T.; Dividich, J.L. Supplementation of a weaning diet with bovine colostrum increases feed intake and growth of weaned piglets. J. Rech. Porcine France 2004, 36, 33–38. [Google Scholar]
- King, M.R.; Ravindran, V.; Morel, P.C.H.; Thomas, D.v.; Birtles, M.J.; Pluske, J.R. Effects of spray-dried colostrum and plasmas on the performance and gut morphology of broiler chickens. Aust. J. Agric. Res. 2005, 56, 811–817. [Google Scholar] [CrossRef]
- Parapary, M.H.G.; Nobakht, A.; Mehmannavaz, Y. Co-supplementation of colostrum powder on performance, intestinal morphology, blood biochemical parameters and antioxidant status of broilers in heat stress. Semina Cien. Agrar. 2020, 41, 3419–3427. [Google Scholar] [CrossRef]
- Giffard, C.J.; Seino, M.M.; Markwell, P.J.; Bektash, R.M. Benefits of bovine colostrum on fecal quality in recently weaned puppies. J. Nutr. 2004, 134, 2126S–2127S. [Google Scholar] [CrossRef] [PubMed]
- Dequenne, M.; Robaye, V.; Dotreppe, O.; Neizer, C.; Delhalle, L.; Thonart, P.; Bertrand, C.; Robert, B.; Moinet, B.; Diez, M. A supplement of bovine colostrum and probiotics increased protein digestibility in dogs but did not influence microbiome. In Proceedings of the 18th Congress of the European Society of Veterinary and Comparative Nutrition, Utrecht, The Netherlands, 11–13 September 2014; Available online: http://hdl.handle.net/2268/200872 (accessed on 1 July 2020).
- Holmes, M.A.; Lunn, D.P. A study of bovine and equine immunoglobulin levels in pony foals fed bovine colostrum. Equine Vet. J. 1991, 2, 116–118. [Google Scholar] [CrossRef]
- Fenger, C.; Tobin, T.; Casey, P.; Langemeier, J.; Haines, D. Bovine colostrum supplementation does not influence serum insulin-like growth factor-1 in horses in race training. J. Equine Vet. Sci. 2014, 34, 1025–1027. [Google Scholar] [CrossRef]





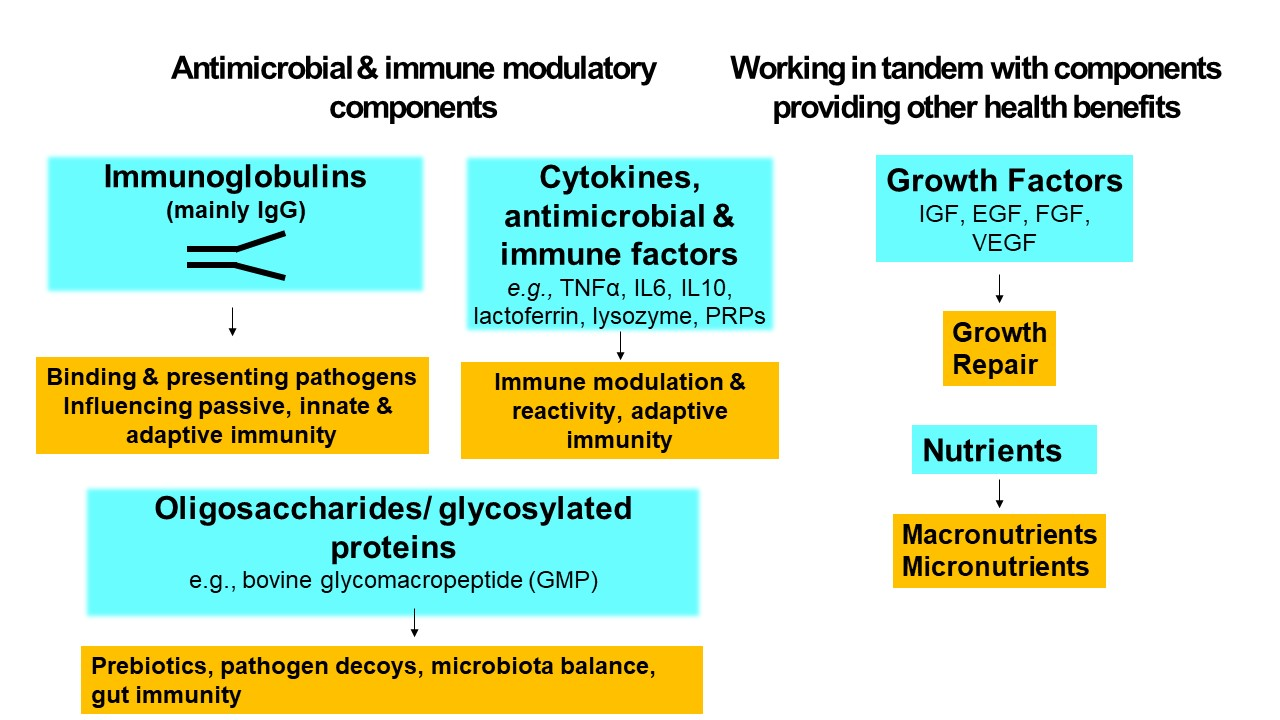{kind=link}
{kind=link}
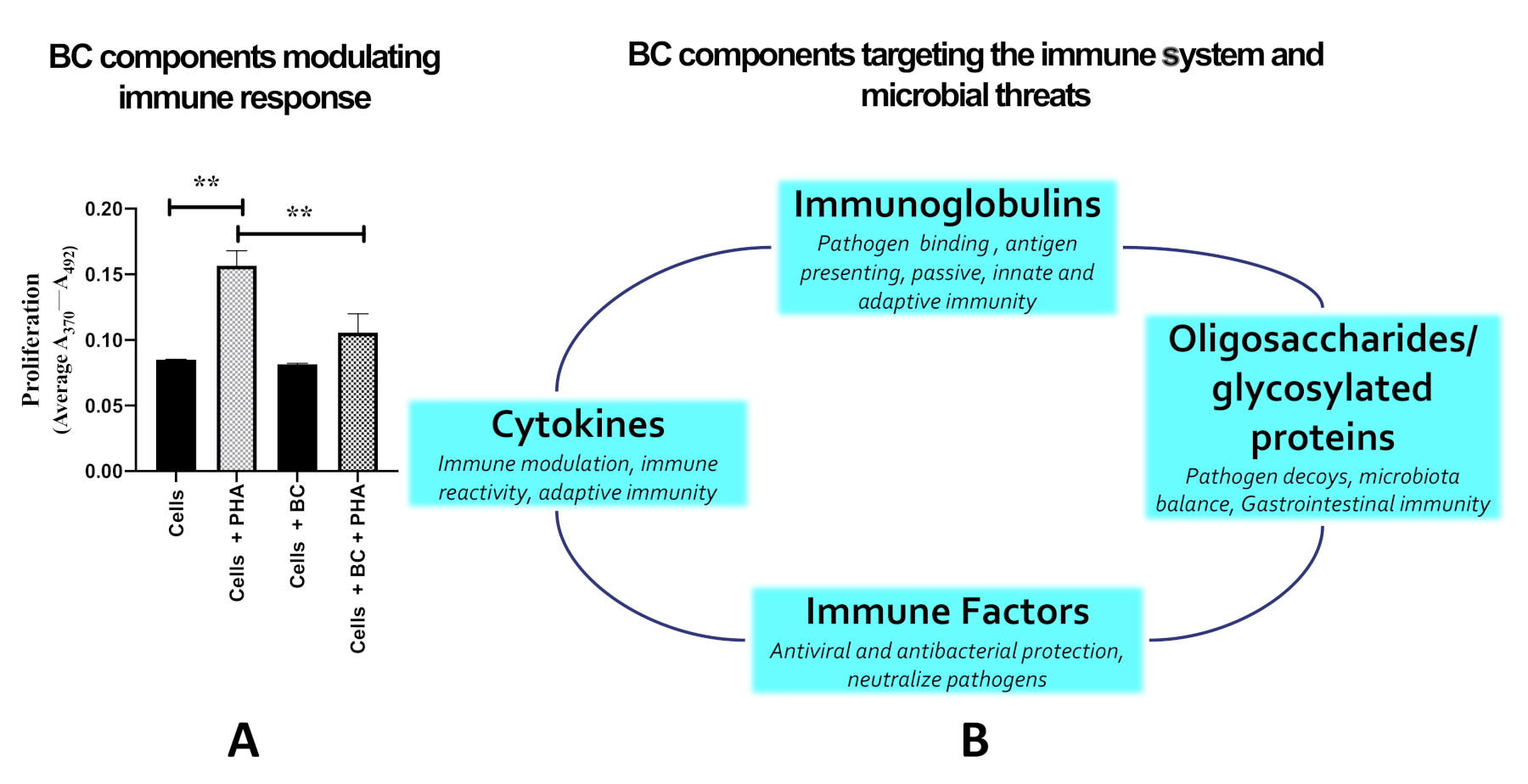{kind=link}
{kind=link}
{kind=link}
| Component | BC | Mature Milk |
|---|---|---|
| Total solids (%) | 24–28 | 12.9 |
| Fat (%) | 6–7 | 3.6–4.0 |
| Protein (%) | 14–16 | 3.1–3.2 |
| Casein (%) | 4.8 | 2.5–2.6 |
| Albumin (%) | 6.0 | 0.4–0.5 |
| Total immunoglobulin (mg/mL) | 42–90 | 0.4–0.9 |
| Lactose (%) | 2–3 | 4.7–5.0 |
| Minerals | ||
| Calcium (g/kg) | 2.6–4.7 | 1.2–1.3 |
| Phosphorus (g/kg) | 4.5 | 0.9–1.2 |
| Potassium (g/kg) | 1.4–2.8 | 1.5–1.7 |
| Sodium (g/kg) | 0.7–1.1 | 0.4 |
| Magnesium (g/kg) | 0.4–0.7 | 0.1 |
| Zinc (mg/kg) | 11.6–38.1 | 3.0–6.0 |
| Vitamins | ||
| Thiamin (B1) (µg/mL) | 0.58–0.90 | 0.4–0.5 |
| Riboflavin (B2) (µg/mL) | 4.55–4.83 | 1.5–1.7 |
| Niacin (B3) (µg/mL) | 0.34–0.96 | 0.8–0.9 |
| Cobalamin (B12) (µg/mL) | 0.05–0.60 | 0.004–0.006 |
| Vitamin A (µg/100 mL) | 25 | 34 |
| Vitamin D (IU/g fat) | 0.89–1.81 | 0.41 |
| Tocopherol (E) (µg/g) | 2.92–5.63 | 0.06 |
| Immunoglobulins | ||
| IgG1 (g/L) | 34.0–87.0 | 0.31–0.40 |
| IgG2 (g/L) | 1.6–6.0 | 0.03–0.08 |
| IgA (g/L) | 3.2–6.2 | 0.04–0.06 |
| IgM (g/L) | 3.7–6.1 | 0.03–0.06 |
| Antimicrobials | ||
| Lactoferrin (g/L) | 1.5–5 | 0.02–0.75 |
| Lactoperoxidase (mg/L) | 11–45 | 13–30 |
| Lysozyme (mg/L) | 0.14–0.7 | 0.07–0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Playford, R.J.; Weiser, M.J. Bovine Colostrum: Its Constituents and Uses. Nutrients 2021, 13, 265. https://doi.org/10.3390/nu13010265
Playford RJ, Weiser MJ. Bovine Colostrum: Its Constituents and Uses. Nutrients. 2021; 13(1):265. https://doi.org/10.3390/nu13010265
Chicago/Turabian StylePlayford, Raymond John, and Michael James Weiser. 2021. "Bovine Colostrum: Its Constituents and Uses" Nutrients 13, no. 1: 265. https://doi.org/10.3390/nu13010265
APA StylePlayford, R. J., & Weiser, M. J. (2021). Bovine Colostrum: Its Constituents and Uses. Nutrients, 13(1), 265. https://doi.org/10.3390/nu13010265