
Circulating Structurally Related (-)-Epicatechin Metabolite Species and Levels after Sustained Intake of a Cocoa Powder High in Polyphenols Are Comparable to Those Achieved after a Single Dose

Abstract
:1. Introduction
2. Materials and Methods
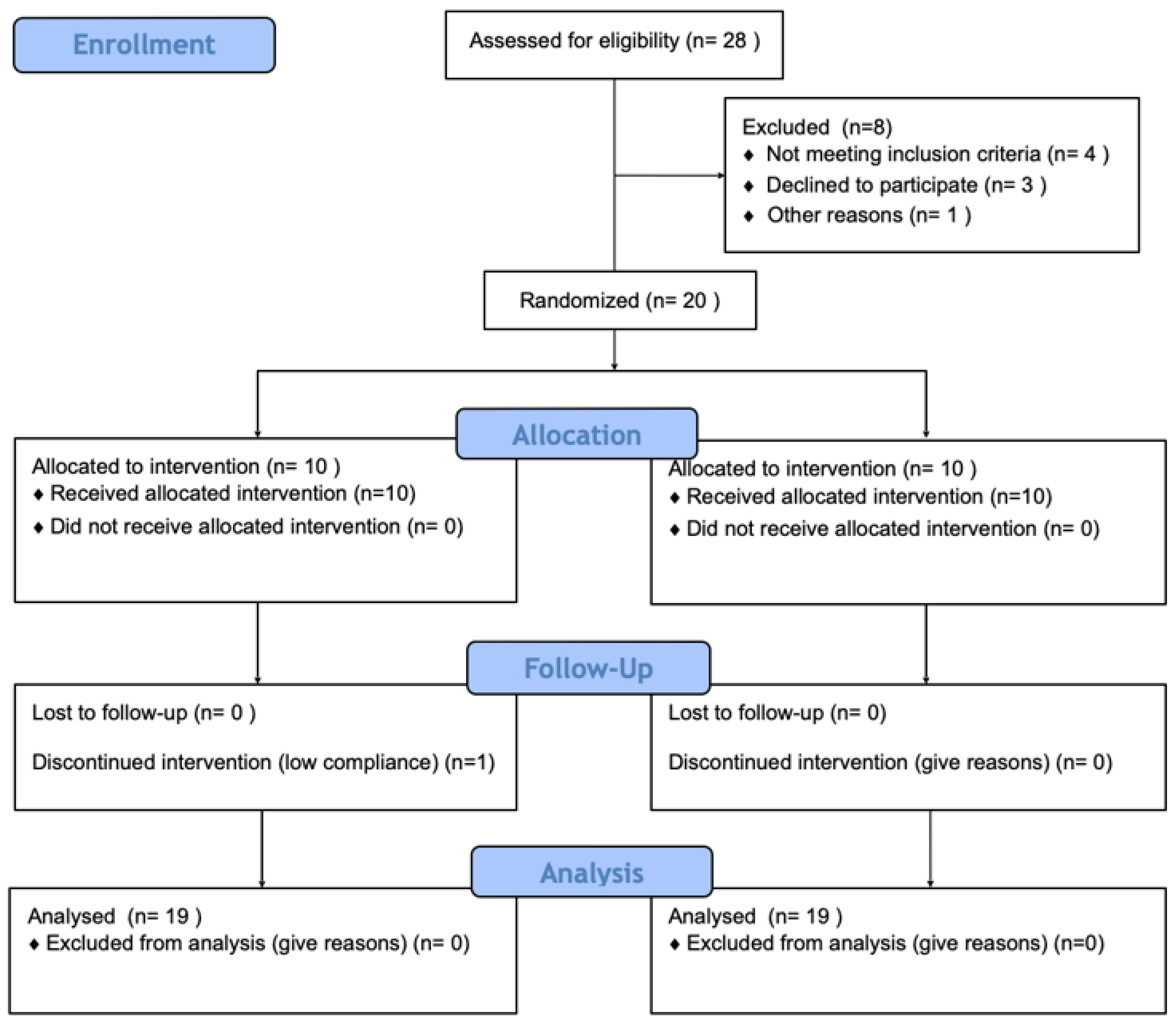
2.1. Participants
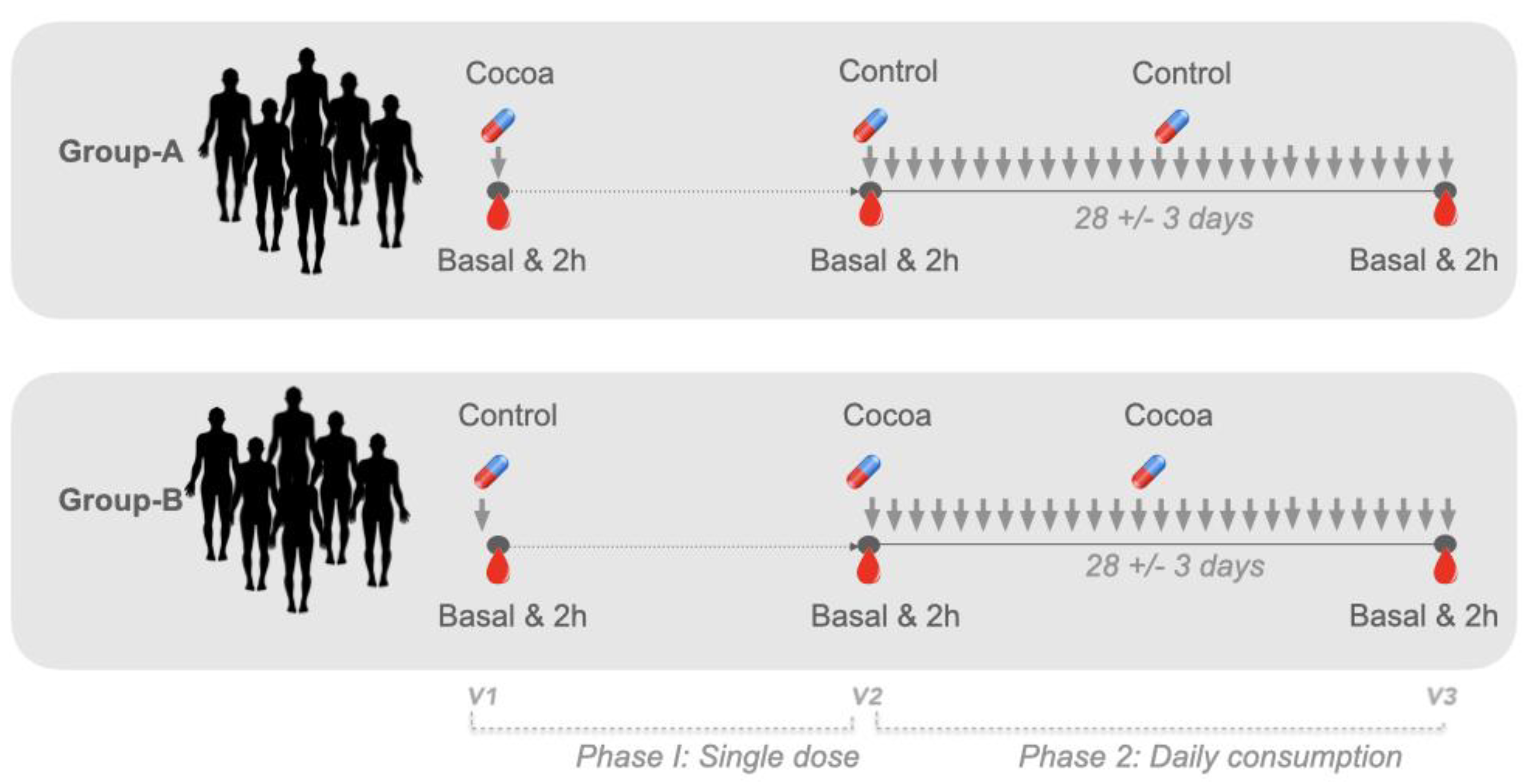
2.2. Study Design
2.3. Intervention and Data Collection
2.4. Study Product
2.5. (−)-Epicatechin Metabolites Profiling
2.6. Quantification of Total Antioxidant Activity of Plasma
3. Results
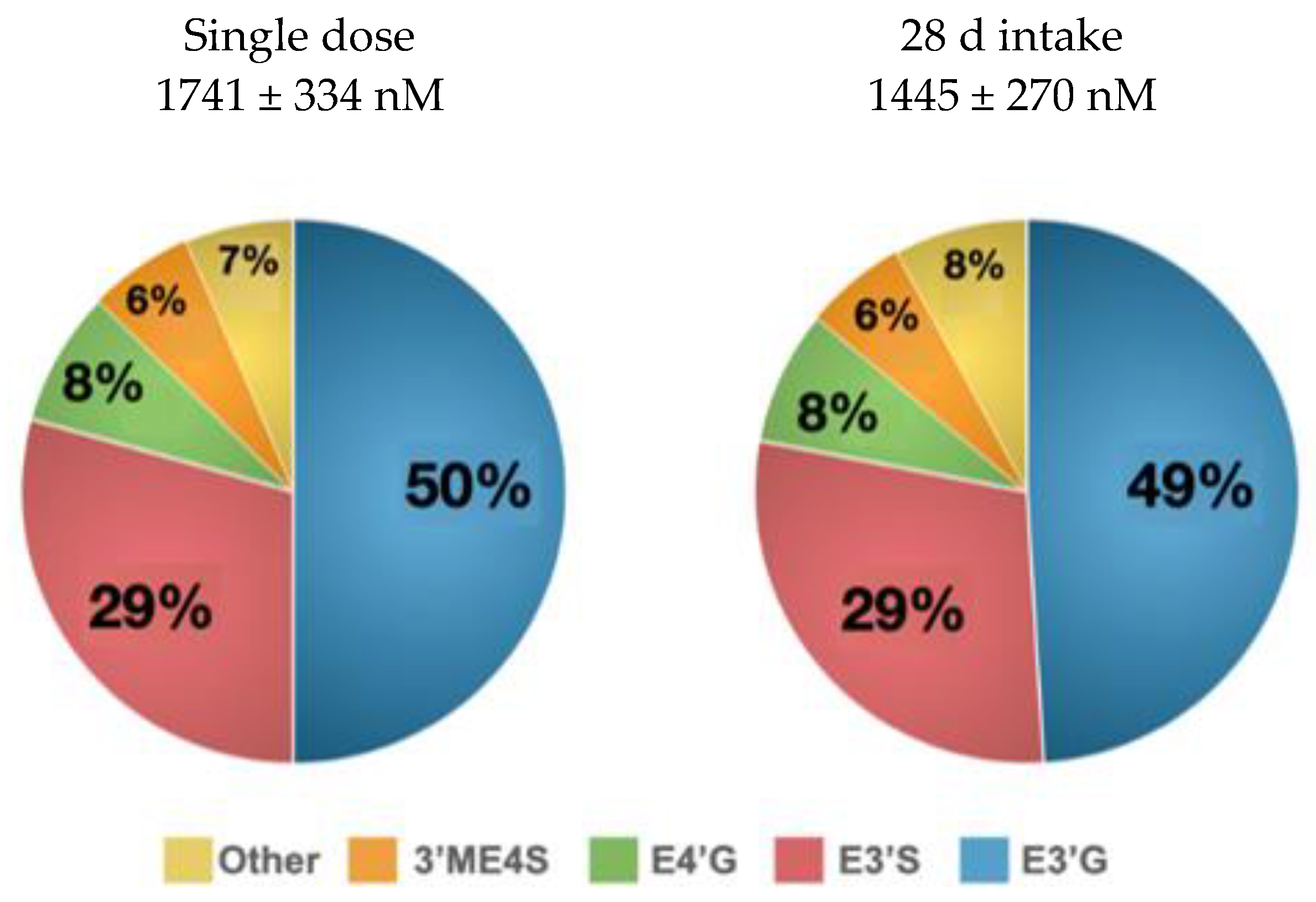
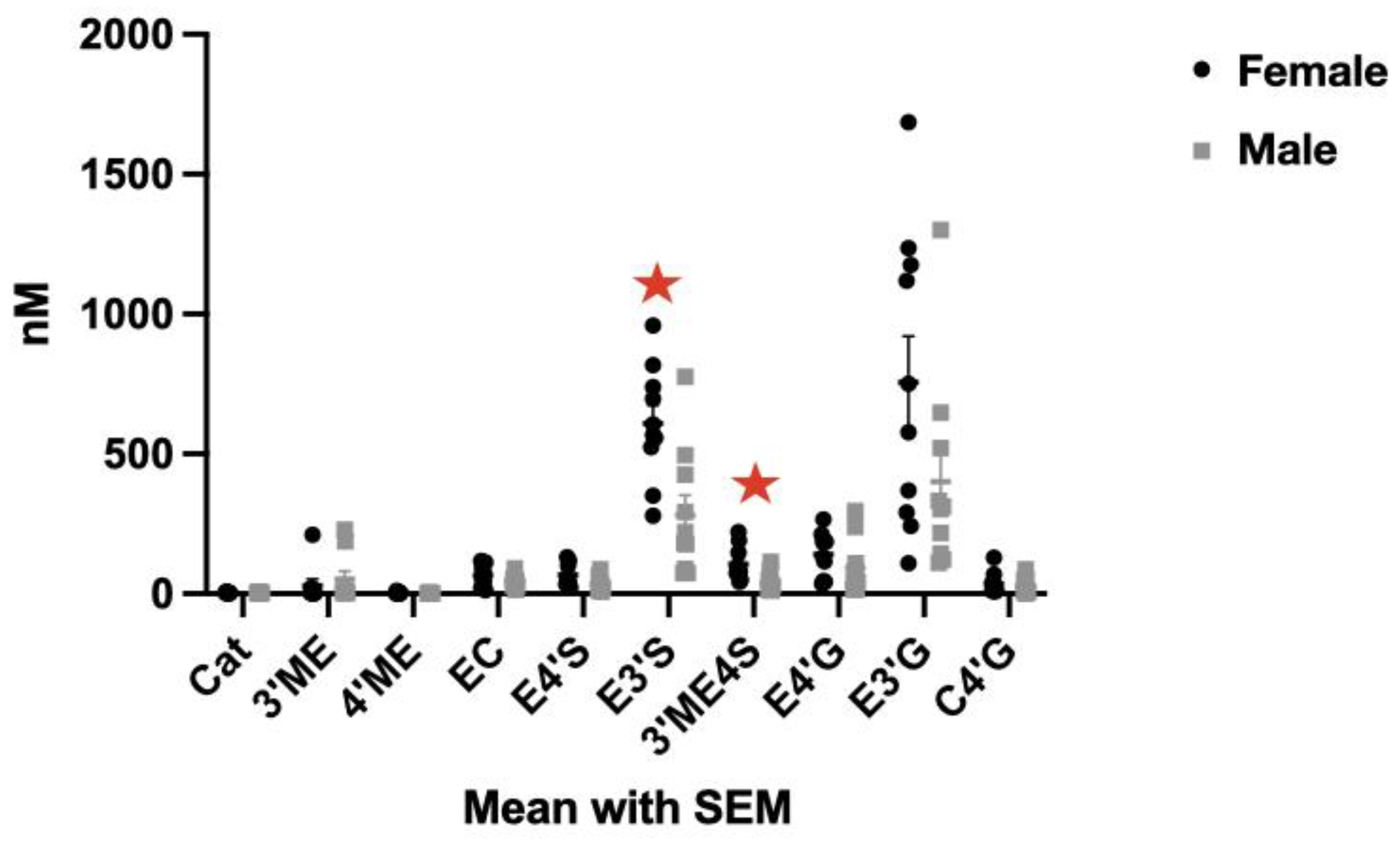
3.1. (-)-Epicatechin Metabolites
3.2. Antioxidant Capacity
4. Discussion
- i.
- The production of phase II metabolites of (-)-epicatechin in the liver and small intestine remained relatively stable after repeated doses with no significant differences in the production of the biotransformed metabolites.
- ii.
- The continuous excretion of SREMs does not allow for accumulation in the body, albeit biological effects have been observed after chronic interventions [2,29,30,31]; most of these studies have attributed the observed effects to SREMs. Recent data suggest, however, that cocoa polyphenols may also influence biological functions through different mechanisms, including the contribution of colonic metabolites [7].
- iii.
- Long-term interventions using the same vehicles and doses may predispose to a more homogeneous response by reducing the initial variability across individuals, which is influenced by former dietary habits [32]. Future studies should evaluate the pharmacokinetics of SREMs at longer interventions, taking into consideration the effect of other colonic metabolites.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Corti, R.; Flammer, A.J.; Hollenberg, N.K.; Luscher, T.F. Cocoa and cardiovascular health. Circulation 2009, 119, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and chocolate in human health and disease. Antioxid. Redox Signal. 2011, 15, 2779–2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, A.; Dodd, G.F.; Spencer, J.P.E. The Effects of Flavonoids on Cardiovascular Health: A Review of Human Intervention Trials and Implications for Cerebrovascular Function. Nutrients 2018, 10, 1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Z.; Liu, A.; Li, P.; Liu, C.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Advances in physiological functions and mechanisms of (-)-epicatechin. Crit. Rev. Food Sci. Nutr. 2021, 61, 211–233. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Oteiza, P.I. Dietary flavonoids: Role of (-)-epicatechin and related procyanidins in cell signaling. Free Radic. Biol. Med. 2011, 51, 813–823. [Google Scholar] [CrossRef]
- Oracz, J.; Nebesny, E.; Zyzelewicz, D.; Budryn, G.; Luzak, B. Bioavailability and metabolism of selected cocoa bioactive compounds: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1947–1985. [Google Scholar] [CrossRef]
- Sorrenti, V.; Ali, S.; Mancin, L.; Davinelli, S.; Paoli, A.; Scapagnini, G. Cocoa Polyphenols and Gut Microbiota Interplay: Bioavailability, Prebiotic Effect, and Impact on Human Health. Nutrients 2020, 12, 1908. [Google Scholar] [CrossRef] [PubMed]
- Jalil, A.M.; Ismail, A. Polyphenols in cocoa and cocoa products: Is there a link between antioxidant properties and health? Molecules 2008, 13, 2190–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludovici, V.; Barthelmes, J.; Nagele, M.P.; Enseleit, F.; Ferri, C.; Flammer, A.J.; Ruschitzka, F.; Sudano, I. Cocoa, Blood Pressure, and Vascular Function. Front. Nutr. 2017, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Actis-Goretta, L.; Leveques, A.; Giuffrida, F.; Romanov-Michailidis, F.; Viton, F.; Barron, D.; Duenas-Paton, M.; Gonzalez-Manzano, S.; Santos-Buelga, C.; Williamson, G.; et al. Elucidation of (-)-epicatechin metabolites after ingestion of chocolate by healthy humans. Free Radic. Biol. Med. 2012, 53, 787–795. [Google Scholar] [CrossRef]
- Ottaviani, J.I.; Borges, G.; Momma, T.Y.; Spencer, J.P.; Keen, C.L.; Crozier, A.; Schroeter, H. The metabolome of [2-(14)C](-)-epicatechin in humans: Implications for the assessment of efficacy, safety, and mechanisms of action of polyphenolic bioactives. Sci. Rep. 2016, 6, 29034. [Google Scholar] [CrossRef] [Green Version]
- Ottaviani, J.I.; Fong, R.; Kimball, J.; Ensunsa, J.L.; Gray, N.; Vogiatzoglou, A.; Britten, A.; Lucarelli, D.; Luben, R.; Grace, P.B.; et al. Evaluation of (-)-epicatechin metabolites as recovery biomarker of dietary flavan-3-ol intake. Sci. Rep. 2019, 9, 13108. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.; Zhang, Y.; Shen, X.; Cao, Y.; Shi, J.; Ye, X.; Chen, S. Rethinking the Mechanism of the Health Benefits of Proanthocyanidins: Absorption, Metabolism, and Interaction with Gut Microbiota. Compr. Rev. Food Sci. Food Saf. 2019, 18, 971–985. [Google Scholar] [CrossRef] [Green Version]
- Borges, G.; Ottaviani, J.I.; van der Hooft, J.J.J.; Schroeter, H.; Crozier, A. Absorption, metabolism, distribution and excretion of (-)-epicatechin: A review of recent findings. Mol. Aspects Med. 2018, 61, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Reyes, P.K.; Hernandez-Ramirez, N.; Cortes, J.; Poquet, L.; Redeuil, K.; Rangel-Escareno, C.; Kussmann, M.; Silva-Zolezzi, I.; Tejero, M.E. Gene expression changes by high-polyphenols cocoa powder intake: A randomized crossover clinical study. Eur. J. Nutr. 2019, 58, 1887–1898. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Mateos, A.; Cifuentes-Gomez, T.; Gonzalez-Salvador, I.; Ottaviani, J.I.; Schroeter, H.; Kelm, M.; Heiss, C.; Spencer, J.P. Influence of age on the absorption, metabolism, and excretion of cocoa flavanols in healthy subjects. Mol. Nutr. Food Res. 2015, 59, 1504–1512. [Google Scholar] [CrossRef]
- Actis-Goretta, L.; Leveques, A.; Rein, M.; Teml, A.; Schafer, C.; Hofmann, U.; Li, H.; Schwab, M.; Eichelbaum, M.; Williamson, G. Intestinal absorption, metabolism, and excretion of (-)-epicatechin in healthy humans assessed by using an intestinal perfusion technique. Am. J. Clin. Nutr. 2013, 98, 924–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manach, C.; Milenkovic, D.; Van de Wiele, T.; Rodriguez-Mateos, A.; de Roos, B.; Garcia-Conesa, M.T.; Landberg, R.; Gibney, E.R.; Heinonen, M.; Tomas-Barberan, F.; et al. Addressing the inter-individual variation in response to consumption of plant food bioactives: Towards a better understanding of their role in healthy aging and cardiometabolic risk reduction. Mol. Nutr. Food Res. 2017, 61, 1600557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellinger, R.W.; Garcia, A.M.; Meyskens, F.L., Jr. Differences in the glucuronidation of resveratrol and pterostilbene: Altered enzyme specificity and potential gender differences. Drug Metab. Pharm. 2014, 29, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrera-Reyes, P.K.; de Lara, J.C.; Gonzalez-Soto, M.; Tejero, M.E. Effects of Cocoa-Derived Polyphenols on Cognitive Function in Humans. Systematic Review and Analysis of Methodological Aspects. Plant. Foods Hum. Nutr. 2020, 75, 1–11. [Google Scholar] [CrossRef]
- Milenkovic, D.; Jude, B.; Morand, C. miRNA as molecular target of polyphenols underlying their biological effects. Free Radic. Biol. Med. 2013, 64, 40–51. [Google Scholar] [CrossRef]
- Zhang, X.; Sandhu, A.; Edirisinghe, I.; Burton-Freeman, B.M. Plasma and Urinary (Poly)phenolic Profiles after 4-Week Red Raspberry (Rubus idaeus L.) Intake with or without Fructo-Oligosaccharide Supplementation. Molecules 2020, 25, 4777. [Google Scholar] [CrossRef] [PubMed]
- Craig, C.L.; Marshall, A.L.; Sjostrom, M.; Bauman, A.E.; Booth, M.L.; Ainsworth, B.E.; Pratt, M.; Ekelund, U.; Yngve, A.; Sallis, J.F.; et al. International physical activity questionnaire: 12-country reliability and validity. Med. Sci. Sports Exerc. 2003, 35, 1381–1395. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Avila, M.; Romieu, I.; Parra, S.; Hernández-Avila, J.; Madrigal, H.; Willett, W. Validity and reproducibility ofa food frequency questionnaire to assessdietary intake of women living in Mexico City. Salud Publica Mex 1998, 40, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lohman, T.; Roache, A.; Martorell, R. Anthropometric Standardisation Reference Manual” Contains Complete Procedures for 45 Anthropometric Measurements; Human Kinetics Books: Champaign, IL, USA, 1991. [Google Scholar]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Monagas, M.; Khan, N.; Andres-Lacueva, C.; Casas, R.; Urpi-Sarda, M.; Llorach, R.; Lamuela-Raventos, R.M.; Estruch, R. Effect of cocoa powder on the modulation of inflammatory biomarkers in patients at high risk of cardiovascular disease. Am. J. Clin. Nutr. 2009, 90, 1144–1150. [Google Scholar] [CrossRef] [Green Version]
- Hollenberg, N.K.; Fisher, N.D.; McCullough, M.L. Flavanols, the Kuna, cocoa consumption, and nitric oxide. J. Am. Soc. Hypertens. 2009, 3, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.A.; Goya, L.; Ramos, S. Protective effects of tea, red wine and cocoa in diabetes. Evidences from human studies. Food Chem. Toxicol. 2017, 109, 302–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento-Silva, A.; Koistinen, V.M.; Mena, P.; Bronze, M.R.; Hanhineva, K.; Sahlstrom, S.; Kitryte, V.; Moco, S.; Aura, A.M. Factors affecting intake, metabolism and health benefits of phenolic acids: Do we understand individual variability? Eur. J. Nutr. 2020, 59, 1275–1293. [Google Scholar] [CrossRef] [Green Version]
- Ottaviani, J.I.; Momma, T.Y.; Kuhnle, G.K.; Keen, C.L.; Schroeter, H. Structurally related (-)-epicatechin metabolites in humans: Assessment using de novo chemically synthesized authentic standards. Free Radic. Biol. Med. 2012, 52, 1403–1412. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Mateos, A.; Oruna-Concha, M.J.; Kwik-Uribe, C.; Vidal, A.; Spencer, J.P. Influence of sugar type on the bioavailability of cocoa flavanols. Br. J. Nutr. 2012, 108, 2243–2250. [Google Scholar] [CrossRef] [Green Version]
- Davison, K.; Coates, A.M.; Buckley, J.D.; Howe, P.R. Effect of cocoa flavanols and exercise on cardiometabolic risk factors in overweight and obese subjects. Int. J. Obes. 2008, 32, 1289–1296. [Google Scholar] [CrossRef] [Green Version]
- van Velzen, E.J.; Westerhuis, J.A.; van Duynhoven, J.P.; van Dorsten, F.A.; Hoefsloot, H.C.; Jacobs, D.M.; Smit, S.; Draijer, R.; Kroner, C.I.; Smilde, A.K. Multilevel data analysis of a crossover designed human nutritional intervention study. J. Proteome Res. 2008, 7, 4483–4491. [Google Scholar] [CrossRef]
- Chow, H.H.; Cai, Y.; Hakim, I.A.; Crowell, J.A.; Shahi, F.; Brooks, C.A.; Dorr, R.T.; Hara, Y.; Alberts, D.S. Pharmacokinetics and safety of green tea polyphenols after multiple-dose administration of epigallocatechin gallate and polyphenon E in healthy individuals. Clin. Cancer Res. 2003, 9, 3312–3319. [Google Scholar]
- Feliciano, R.P.; Istas, G.; Heiss, C.; Rodriguez-Mateos, A. Plasma and Urinary Phenolic Profiles after Acute and Repetitive Intake of Wild Blueberry. Molecules 2016, 21, 1120. [Google Scholar] [CrossRef]
- Jacobs, D.M.; Fuhrmann, J.C.; van Dorsten, F.A.; Rein, D.; Peters, S.; van Velzen, E.J.; Hollebrands, B.; Draijer, R.; van Duynhoven, J.; Garczarek, U. Impact of short-term intake of red wine and grape polyphenol extract on the human metabolome. J. Agric. Food Chem. 2012, 60, 3078–3085. [Google Scholar] [CrossRef]
- Lotito, S.B.; Frei, B. Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: Cause, consequence, or epiphenomenon? Free Radic. Biol. Med. 2006, 41, 1727–1746. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Jia, Z.; Battino, M.; Miron, A.; Yu, Z.; Cao, H.; Xiao, J. Intracellular signaling pathways of inflammation modulated by dietary flavonoids: The most recent evidence. Crit. Rev. Food Sci. Nutr. 2018, 58, 2908–2924. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Mg/g |
|---|---|
| Total polyphenol Content | 500 |
| Epicatechin a | 78 |
| Flavan-3-ols b | 200 |
| Flavanols | 100 |
| Theobromine | 50 |
| Variable | Group A n = 10 | Group B n = 10 | p Value * |
|---|---|---|---|
| Sex (male) (%) | 4 (36%) | 6 (67%) | |
| Age (years) | 27.4 (4.0) | 26.7 (5.2) | 0.68 |
| BMI (kg/m2) | 23.7 (2.9) | 23.0 (3.1) | 0.65 |
| Physical activity (METs) | 2512 (2449) | 2197 (1203) | 0.42 |
| Glucose (mg/dL) | 90.9 (6.6) | 90.5 (6.5) | 0.88 |
| Cholesterol (mg/dL) | 158.4 (37.6) | 189.8 (37.8) | 0.06 |
| HDL (mg/dL) | 52.2 (13.0) | 47.6 (12.3) | 0.99 |
| LDL (mg/dL) | 90.2 (34.0) | 117.5(34.7) | 0.13 |
| Triglycerides (mg/dL) | 80.2 (27.2) | 120.7 (56.4) | 0.13 |
| Energy intake (kcal) | 1814 (518) | 2103 (807) | 0.29 |
| Fat mass (%) | 23.5 (5.7) | 25.5 (5.7) | 0.27 |
| Lean mass (kg) | 31.2 (9.5) | 33.2 (12.6) | 0.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrera-Reyes, P.K.; Cortés-Fernández de Lara, J.; Poquet, L.; Redeuil, K.; Kussmann, M.; Silva-Zolezzi, I.; Tejero, E.M. Circulating Structurally Related (-)-Epicatechin Metabolite Species and Levels after Sustained Intake of a Cocoa Powder High in Polyphenols Are Comparable to Those Achieved after a Single Dose. Nutrients 2021, 13, 3829. https://doi.org/10.3390/nu13113829
Barrera-Reyes PK, Cortés-Fernández de Lara J, Poquet L, Redeuil K, Kussmann M, Silva-Zolezzi I, Tejero EM. Circulating Structurally Related (-)-Epicatechin Metabolite Species and Levels after Sustained Intake of a Cocoa Powder High in Polyphenols Are Comparable to Those Achieved after a Single Dose. Nutrients. 2021; 13(11):3829. https://doi.org/10.3390/nu13113829
Chicago/Turabian StyleBarrera-Reyes, Paloma K., Josué Cortés-Fernández de Lara, Laure Poquet, Karine Redeuil, Martin Kussmann, Irma Silva-Zolezzi, and Elizabeth M. Tejero. 2021. "Circulating Structurally Related (-)-Epicatechin Metabolite Species and Levels after Sustained Intake of a Cocoa Powder High in Polyphenols Are Comparable to Those Achieved after a Single Dose" Nutrients 13, no. 11: 3829. https://doi.org/10.3390/nu13113829
APA StyleBarrera-Reyes, P. K., Cortés-Fernández de Lara, J., Poquet, L., Redeuil, K., Kussmann, M., Silva-Zolezzi, I., & Tejero, E. M. (2021). Circulating Structurally Related (-)-Epicatechin Metabolite Species and Levels after Sustained Intake of a Cocoa Powder High in Polyphenols Are Comparable to Those Achieved after a Single Dose. Nutrients, 13(11), 3829. https://doi.org/10.3390/nu13113829