
Effect of Dairy Matrix on the Postprandial Blood Metabolome


 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Test Meals
2.3. NMR Spectroscopy
2.4. Statistics
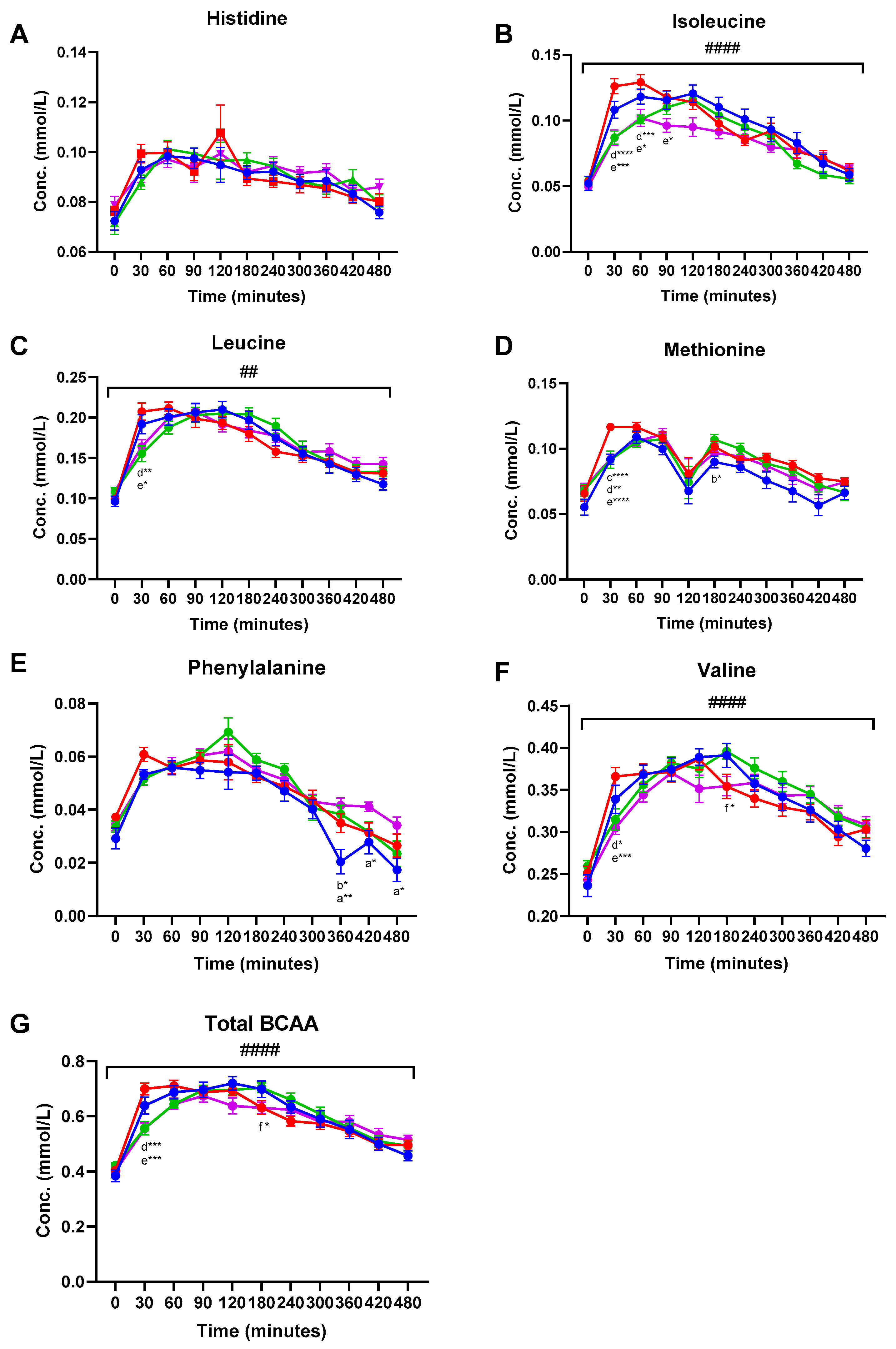
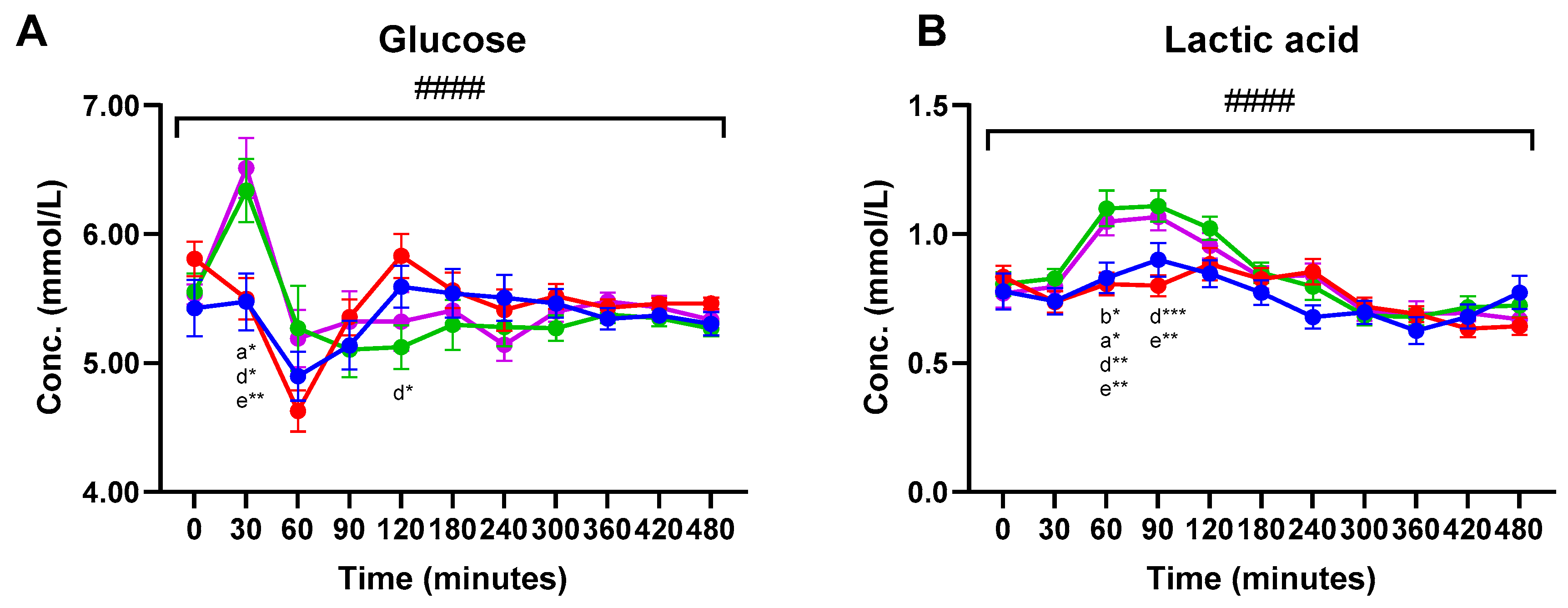
3. Results
3.1. Amino Acid Concentrations in Plasma
3.2. Plasma Metabolites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lovegrove, J.A. Dietary dilemmas over fats and cardiometabolic risk. Proc. Nutr. Soc. 2020, 79, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Hjerpsted, J.; Leedo, E.; Tholstrup, T. Cheese intake in large amounts lowers LDL-cholesterol concentrations compared with butter intake of equal fat content. Am. J. Clin. Nutr. 2011, 94, 1479–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeney, E.L.; Barron, R.; Dible, V.; Hamilton, Z.; Power, Y.; Tanner, L.; Flynn, C.; Bouchier, P.; Beresford, T.; Noronha, N.; et al. Dairy matrix effects: Response to consumption of dairy fat differs when eaten within the cheese matrix-a randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 667–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouin-Chartier, J.-P.; Tremblay, A.; Maltais-Giguère, J.; Charest, A.; Guinot, L.; Rioux, L.-E.; Labrie, S.; Britten, M.; Lamarche, B.; Turgeon, S.; et al. Differential impact of the cheese matrix on the postprandial lipid response: A randomized, crossover, controlled trial. Am. J. Clin. Nutr. 2017, 106, 1358–1365. [Google Scholar] [CrossRef] [Green Version]
- Thorning, T.K.; Bertram, H.C.; Bonjour, J.-P.; de Groot, L.; Dupont, D.; Feeney, E.; Ipsen, R.; Lecerf, J.M.; Mackie, A.; McKinley, M.C.; et al. Whole dairy matrix or single nutrients in assessment of health effects: Current evidence and knowledge gaps. Am. J. Clin. Nutr. 2017, 105, 1033–1045. [Google Scholar] [CrossRef]
- Parada, J.J.M. Aguilera, Food Microstructure Affects the Bioavailability of Several Nutrients. J. Food Sci. 2007, 72, R21–R32. [Google Scholar] [CrossRef]
- Grundy, M.M.; Grassby, T.; Mandalari, G.; Waldron, K.W.; Butterworth, P.J.; Berry, S.E.; Ellis, P.R. Effect of mastication on lipid bioaccessibility of almonds in a randomized human study and its implications for digestion kinetics, metabolizable energy, and postprandial lipemia. Am. J. Clin. Nutr. 2015, 101, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Fardet, A.; Dupont, D.; Rioux, L.E.; Turgeon, S.L. Influence of food structure on dairy protein, lipid and calcium bioavailability: A narrative review of evidence. Crit. Rev. Food Sci. Nutr. 2019, 59, 1987–2010. [Google Scholar] [CrossRef]
- Fang, X.; Rioux, L.-E.; Labrie, S.; Turgeon, S.L. Commercial cheeses with different texture have different disintegration and protein/peptide release rates during simulated in vitro digestion. Int. Dairy J. 2016, 56, 169–178. [Google Scholar] [CrossRef]
- Fang, X.; Rioux, L.-E.; Labrie, S.; Turgeon, S.L. Disintegration and nutrients release from cheese with different textural properties during in vitro digestion. Food Res. Int. 2016, 88, 276–283. [Google Scholar] [CrossRef]
- Lamothe, S.; Rémillard, N.; Tremblay, J.; Britten, M. Influence of dairy matrices on nutrient release in a simulated gastrointestinal environment. Food Res. Int. 2017, 92, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Barbé, F.; Ménard, O.; Le Gouar, Y.; Buffière, C.; Famelart, M.-H.; Laroche, B.; Le Feunteun, S.; Dupont, D.; Rémond, D. The heat treatment and the gelation are strong determinants of the kinetics of milk proteins digestion and of the peripheral availability of amino acids. Food Chem. 2013, 136, 1203–1212. [Google Scholar] [CrossRef]
- Horstman, A.M.H.; Ganzevles, R.A.; Kudla, U.; Kardinaal, A.F.M.; van den Borne, J.J.G.C.; Huppertz, T. Postprandial blood amino acid concentrations in older adults after consumption of dairy products: The role of the dairy matrix. Int. Dairy J. 2021, 113, 104890. [Google Scholar] [CrossRef]
- Kjølbæk, L.; Schmidt, J.M.; Rouy, E.; Jensen, K.J.; Astrup, A.; Bertram, H.C.; Hammershøj, M.; Raben, A. Matrix structure of dairy products results in different postprandial lipid responses: A randomized crossover trial. Am. J. Clin. Nutr. 2021, 114, 1729–1742. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.M.; Kjølbæk, L.; Jensen, K.J.; Rouy, E.; Bertram, H.C.; Larsen, T.; Raben, A.; Astrup, A.; Hammershøj, M. Influence of type of dairy matrix micro- and macrostructure on in vitro lipid digestion. Food Funct. 2020, 11, 4960–4972. [Google Scholar] [CrossRef]
- Thøgersen, R.; Lindahl, I.E.I.; Khakimov, B.; Kjølbæk, L.; Juhl Jensen, K.; Astrup, A.; Hammershøj, M.; Raben, A.; Bertram, H.C. Progression of Postprandial Blood Plasma Phospholipids Following Acute Intake of Different Dairy Matrices: A Randomized Crossover Trial. Metabolites 2021, 11, 454. [Google Scholar] [CrossRef]
- Jiménez, B.; Holmes, E.; Heude, C.; Tolson, R.F.; Harvey, N.; Lodge, S.L.; Chetwynd, A.J.; Cannet, C.; Fang, F.; Pearce, J.T.M.; et al. Quantitative Lipoprotein Subclass and Low Molecular Weight Metabolite Analysis in Human Serum and Plasma by 1H NMR Spectroscopy in a Multilaboratory Trial. Anal. Chem. 2018, 90, 11962–11971. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Sanggaard, K.M.; Holst, J.J.; Rehfeld, J.F.; Sandström, B.; Raben, A.; Tholstrup, T. Different effects of whole milk and a fermented milk with the same fat and lactose content on gastric emptying and postprandial lipaemia, but not on glycaemic response and appetite. Br. J. Nutr. 2004, 92, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Gaudichon, C.; Mahé, S.; Roos, N.; Benamouzig, R.; Luengo, C.; Huneau, J.F.; Sick, H.; Bouley, C.; Rautureau, J.; Tome, D. Exogenous and endogenous nitrogen flow rates and level of protein hydrolysis in the human jejunum after [15N]milk and [15N]yoghurt ingestion. Br. J. Nutr. 1995, 74, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Hermans, W.J.H.; Senden, J.M.; Churchward-Venne, T.A.; Paulussen, K.J.M.; Fuchs, C.J.; Smeets, J.S.J.; van Loon, J.J.A.; Verdijk, L.B.; van Loon, L.J.C. Insects are a viable protein source for human consumption: From insect protein digestion to postprandial muscle protein synthesis in vivo in humans: A double-blind randomized trial. Am. J. Clin. Nutr. 2021, 114, 934–944. [Google Scholar] [CrossRef] [PubMed]
- de Kruif, C.G. Skim Milk Acidification. J. Colloid Interface Sci. 1997, 185, 19–25. [Google Scholar] [CrossRef]
- Tsukahara, T.; Koyama, H.; Okada, M.; Ushida, K. Stimulation of butyrate production by gluconic acid in batch culture of pig cecal digesta and identification of butyrate-producing bacteria. J. Nutr. 2002, 132, 2229–2234. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Yuasa, K.; Kunugita, K.; Teraji, T.; Mitsuoka, T. Effects of Gluconic Acid on Human Faecal Bacteria. Microb. Ecol. Health Dis. 1994, 7, 247–256. [Google Scholar]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Trapecar, M.; Communal, C.; Velazquez, J.; Maass, C.A.; Huang, Y.J.; Schneider, K.; Wright, C.W.; Butty, V.; Eng, G.; Yilmaz, O.; et al. Gut-Liver Physiomimetics Reveal Paradoxical Modulation of IBD-Related Inflammation by Short-Chain Fatty Acids. Cell Syst. 2020, 10, 223–239. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Time (min) | 0 | 30 | 60 | 90 | 120 | 180 | 240 | 300 | 360 | 420 | 480 | qmeal-time | qmeal | qtime |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acetic acid | ||||||||||||||
| Cheese | 0.070 | 0.063 | 0.036 | 0.029 | 0.030 ab | 0.025 a | 0.018 a | 0.030 a | 0.038 a | 0.036 a | 0.027 a | <0.001 | <0.001 | <0.001 |
| Hom. Cheese | 0.057 | 0.046 | 0.029 | 0.017 | 0.023 a | 0.026 a | 0.026 a | 0.034 a | 0.032 a | 0.033 a | 0.034 ab | |||
| MCI Drink | 0.057 | 0.044 | 0.024 | 0.027 | 0.033 ab | 0.023 a | 0.020 a | 0.028 a | 0.029 a | 0.031 a | 0.027 a | |||
| MCI Gel | 0.066 | 0.050 | 0.037 | 0.032 | 0.050 b | 0.056 b | 0.073 b | 0.084 b | 0.095 b | 0.066 b | 0.057 b | |||
| Acetoacetic acid | ||||||||||||||
| Cheese | 0.070 | 0.063 | 0.036 | 0.029 | 0.030 | 0.025 | 0.018 | 0.030 | 0.038 ab | 0.036 | 0.027 | <0.001 | 0.831 | 0.003 |
| Hom. Cheese | 0.057 | 0.046 | 0.029 | 0.017 | 0.023 | 0.026 | 0.026 | 0.034 | 0.032 ac | 0.033 | 0.034 | |||
| MCI Drink | 0.057 | 0.044 | 0.024 | 0.027 | 0.033 | 0.023 | 0.020 | 0.028 | 0.029 c | 0.031 | 0.027 | |||
| MCI Gel | 0.066 | 0.050 | 0.037 | 0.032 | 0.050 | 0.056 | 0.073 | 0.084 | 0.095 b | 0.066 | 0.057 | |||
| Acetone | ||||||||||||||
| Cheese | 0.035 | 0.035 | 0.033 | 0.037 | 0.036 | 0.039 | 0.036 | 0.031 | 0.032 | 0.035 | 0.036 | 0.112 | 0.992 | 0.001 |
| Hom. Cheese | 0.035 | 0.033 | 0.038 | 0.036 | 0.034 | 0.038 | 0.031 | 0.032 | 0.031 | 0.035 | 0.032 | |||
| MCI Drink | 0.032 | 0.044 | 0.039 | 0.042 | 0.036 | 0.042 | 0.035 | 0.037 | 0.035 | 0.035 | 0.032 | |||
| MCI Gel | 0.024 | 0.032 | 0.037 | 0.037 | 0.030 | 0.039 | 0.035 | 0.030 | 0.036 | 0.034 | 0.038 | |||
| Citric acid | ||||||||||||||
| Cheese | 0.148 | 0.157 | 0.156 | 0.151 | 0.142 | 0.139 | 0.153 | 0.148 | 0.145 | 0.147 | 0.162 | 0.061 | 0.992 | <0.001 |
| Hom. Cheese | 0.143 | 0.157 | 0.167 | 0.161 | 0.126 | 0.150 | 0.141 | 0.139 | 0.139 | 0.162 | 0.150 | |||
| MCI Drink | 0.142 | 0.161 | 0.149 | 0.141 | 0.149 | 0.137 | 0.156 | 0.141 | 0.141 | 0.153 | 0.158 | |||
| MCI Gel | 0.130 | 0.166 | 0.157 | 0.148 | 0.127 | 0.150 | 0.148 | 0.158 | 0.148 | 0.165 | 0.168 | |||
| Creatine | ||||||||||||||
| Cheese | 0.008 | 0.013 | 0.011 | 0.010 | 0.007 | 0.007 | 0.005 | 0.005 | 0.005 | 0.005 | 0.005 | 0.963 | 0.992 | <0.001 |
| Hom. Cheese | 0.007 | 0.014 | 0.010 | 0.012 | 0.005 | 0.009 | 0.007 | 0.008 | 0.008 | 0.006 | 0.006 | |||
| MCI Drink | 0.010 | 0.014 | 0.012 | 0.011 | 0.006 | 0.006 | 0.007 | 0.005 | 0.005 | 0.006 | 0.003 | |||
| MCI Gel | 0.007 | 0.013 | 0.012 | 0.011 | 0.009 | 0.008 | 0.008 | 0.005 | 0.007 | 0.006 | 0.006 | |||
| Creatinine | ||||||||||||||
| Cheese | 0.098 | 0.107 | 0.103 | 0.105 | 0.104 | 0.099 | 0.099 | 0.094 | 0.095 | 0.094 | 0.094 | 0.691 | 0.992 | <0.001 |
| Hom. Cheese | 0.098 | 0.106 | 0.111 | 0.101 | 0.104 | 0.102 | 0.096 | 0.095 | 0.094 | 0.095 | 0.093 | |||
| MCI Drink | 0.099 | 0.109 | 0.105 | 0.102 | 0.103 | 0.097 | 0.096 | 0.094 | 0.095 | 0.093 | 0.093 | |||
| MCI Gel | 0.092 | 0.109 | 0.110 | 0.102 | 0.107 | 0.098 | 0.096 | 0.094 | 0.089 | 0.094 | 0.090 | |||
| Dimethylsulfone | ||||||||||||||
| Cheese | 0.008 | 0.007 | 0.009 | 0.010 | 0.007 | 0.008 | 0.008 | 0.007 | 0.007 | 0.007 | 0.007 | 0.649 | 0.992 | 0.043 |
| Hom. Cheese | 0.008 | 0.010 | 0.008 | 0.009 | 0.007 | 0.009 | 0.008 | 0.008 | 0.010 | 0.008 | 0.008 | |||
| MCI Drink | 0.010 | 0.011 | 0.009 | 0.006 | 0.004 | 0.007 | 0.008 | 0.009 | 0.007 | 0.009 | 0.009 | |||
| MCI Gel | 0.006 | 0.007 | 0.007 | 0.005 | 0.004 | 0.006 | 0.007 | 0.006 | 0.008 | 0.007 | 0.008 | |||
| Formic acid | ||||||||||||||
| Cheese | 0.020 | 0.021 | 0.024 | 0.024 | 0.015 | 0.019 ab | 0.023 a | 0.021 | 0.021 ab | 0.022 ab | 0.014 a | 0.212 | 0.041 | 0.061 |
| Hom. Cheese | 0.019 | 0.026 | 0.024 | 0.021 | 0.015 | 0.020 ab | 0.021 a | 0.022 | 0.019 ab | 0.021 a | 0.019 a | |||
| MCI Drink | 0.022 | 0.022 | 0.019 | 0.026 | 0.018 | 0.016 a | 0.022 ab | 0.018 | 0.015 a | 0.020 a | 0.021 ab | |||
| MCI Gel | 0.024 | 0.021 | 0.024 | 0.021 | 0.021 | 0.027 b | 0.030 b | 0.028 | 0.028 b | 0.030 b | 0.029 b | |||
| Glucose | ||||||||||||||
| Cheese | 5.533 | 6.514 a | 5.191 | 5.322 | 5.322 ab | 5.408 | 5.140 | 5.398 | 5.480 | 5.432 | 5.337 ab | <0.001 | 0.992 | <0.001 |
| Hom. Cheese | 5.554 | 6.338 ac | 5.270 | 5.103 | 5.123 a | 5.296 | 5.279 | 5.270 | 5.376 | 5.349 | 5.265 a | |||
| MCI Drink | 5.808 | 5.500 b | 4.629 | 5.355 | 5.830 b | 5.564 | 5.411 | 5.518 | 5.428 | 5.462 | 5.465 b | |||
| MCI Gel | 5.427 | 5.476 bc | 4.898 | 5.137 | 5.591 ab | 5.541 | 5.507 | 5.463 | 5.344 | 5.368 | 5.304 ab | |||
| 3-hydroxybutyric acid | ||||||||||||||
| Cheese | 0.088 | 0.073 | 0.046 | 0.040 | 0.042 | 0.058 | 0.053 | 0.078 | 0.098 ab | 0.113 | 0.139 | 0.037 | 0.992 | <0.001 |
| Hom. Cheese | 0.136 | 0.075 | 0.047 | 0.043 | 0.042 | 0.060 | 0.073 | 0.072 | 0.082 ab | 0.129 | 0.120 | |||
| MCI Drink | 0.074 | 0.096 | 0.061 | 0.056 | 0.062 | 0.052 | 0.049 | 0.050 | 0.059 a | 0.085 | 0.084 | |||
| MCI Gel | 0.074 | 0.094 | 0.059 | 0.042 | 0.049 | 0.063 | 0.080 | 0.079 | 0.121 b | 0.109 | 0.139 | |||
| Lactic acid | ||||||||||||||
| Cheese | 0.771 | 0.796 | 1.048 a | 1.067 a | 0.955 | 0.832 | 0.841 | 0.701 | 0.691 | 0.695 | 0.670 | <0.001 | 0.438 | <0.001 |
| Hom. Cheese | 0.807 | 0.828 | 1.100 a | 1.110 a | 1.023 | 0.850 | 0.797 | 0.685 | 0.678 | 0.717 | 0.724 | |||
| MCI Drink | 0.835 | 0.737 | 0.807 b | 0.800 b | 0.884 | 0.826 | 0.854 | 0.720 | 0.690 | 0.634 | 0.643 | |||
| MCI Gel | 0.778 | 0.740 | 0.831 b | 0.900 ab | 0.846 | 0.773 | 0.678 | 0.697 | 0.626 | 0.680 | 0.773 | |||
| Pyruvic acid | ||||||||||||||
| Cheese | 0.061 | 0.064 | 0.100 a | 0.103 a | 0.078 | 0.073 | 0.062 | 0.057 | 0.053 | 0.049 | 0.044 | <0.001 | 0.992 | <0.001 |
| Hom. Cheese | 0.062 | 0.068 | 0.091 ab | 0.103 a | 0.076 | 0.075 | 0.064 | 0.057 | 0.046 | 0.048 | 0.043 | |||
| MCI Drink | 0.068 | 0.066 | 0.075 ab | 0.074 b | 0.073 | 0.076 | 0.073 | 0.065 | 0.054 | 0.045 | 0.045 | |||
| MCI Gel | 0.064 | 0.056 | 0.071 b | 0.083 ab | 0.065 | 0.070 | 0.058 | 0.052 | 0.047 | 0.046 | 0.049 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thøgersen, R.; Egsgaard, K.L.; Kjølbæk, L.; Jensen, K.J.; Astrup, A.; Hammershøj, M.; Raben, A.; Bertram, H.C. Effect of Dairy Matrix on the Postprandial Blood Metabolome. Nutrients 2021, 13, 4280. https://doi.org/10.3390/nu13124280
Thøgersen R, Egsgaard KL, Kjølbæk L, Jensen KJ, Astrup A, Hammershøj M, Raben A, Bertram HC. Effect of Dairy Matrix on the Postprandial Blood Metabolome. Nutrients. 2021; 13(12):4280. https://doi.org/10.3390/nu13124280
Chicago/Turabian StyleThøgersen, Rebekka, Kristian Leth Egsgaard, Louise Kjølbæk, Klaus Juhl Jensen, Arne Astrup, Marianne Hammershøj, Anne Raben, and Hanne Christine Bertram. 2021. "Effect of Dairy Matrix on the Postprandial Blood Metabolome" Nutrients 13, no. 12: 4280. https://doi.org/10.3390/nu13124280
APA StyleThøgersen, R., Egsgaard, K. L., Kjølbæk, L., Jensen, K. J., Astrup, A., Hammershøj, M., Raben, A., & Bertram, H. C. (2021). Effect of Dairy Matrix on the Postprandial Blood Metabolome. Nutrients, 13(12), 4280. https://doi.org/10.3390/nu13124280