
The Gut Microbiome and Metabolomics Profiles of Restricting and Binge-Purging Type Anorexia Nervosa

 , ,
, ,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbiome Analysis
2.2. Metabolomic Analysis
2.3. Statistical Analysis
3. Results
3.1. Clinical Characteristics
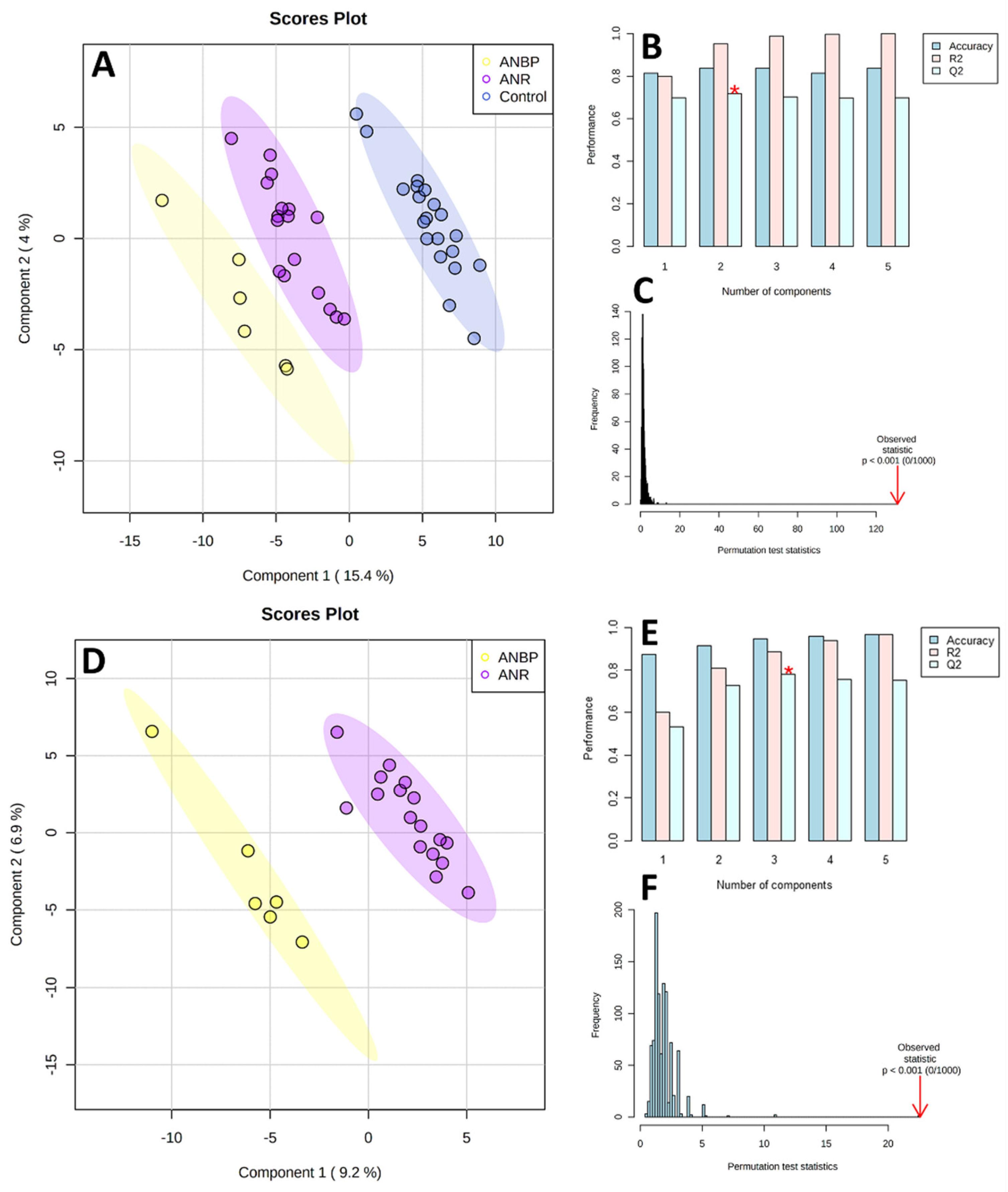
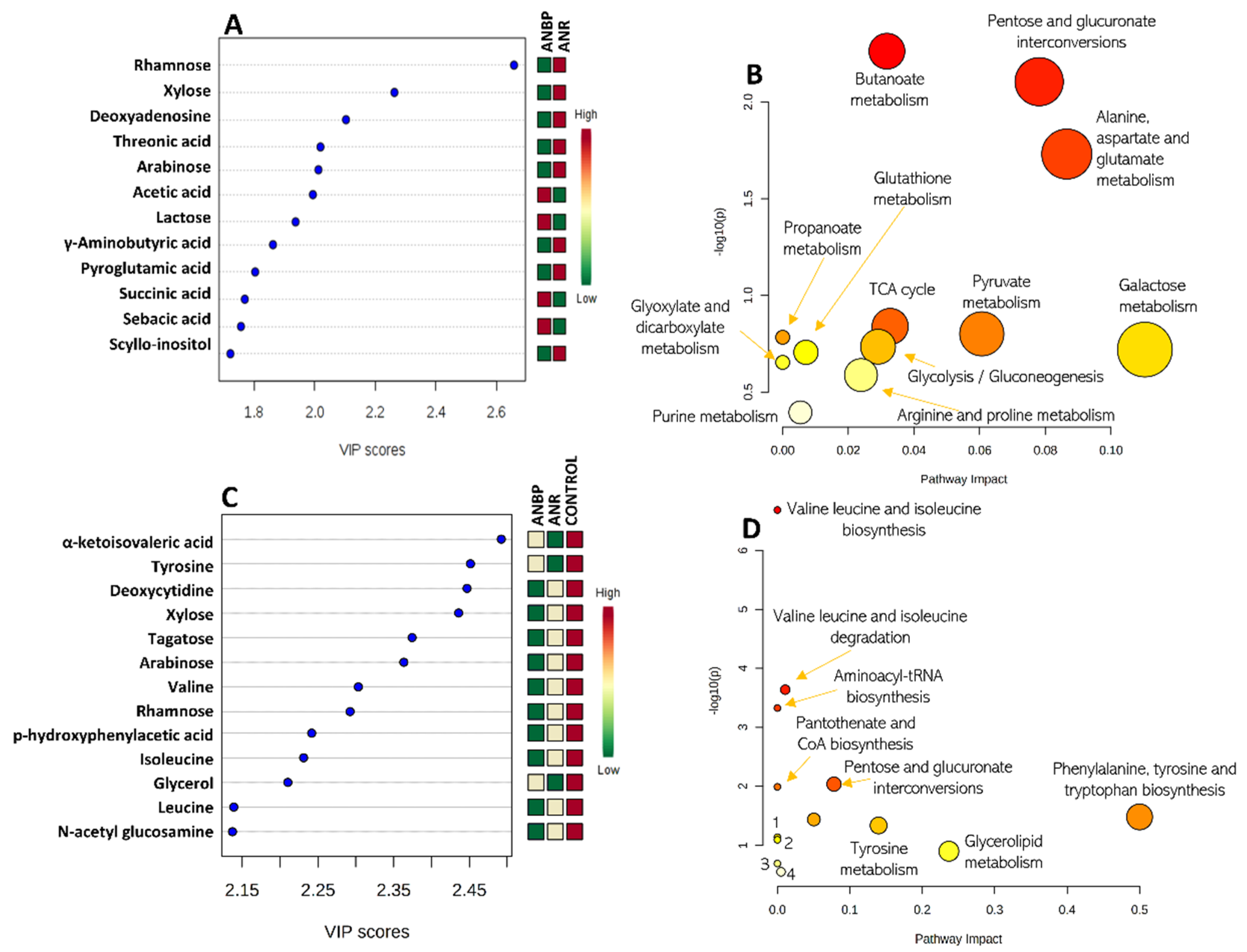
3.2. Metabolomics
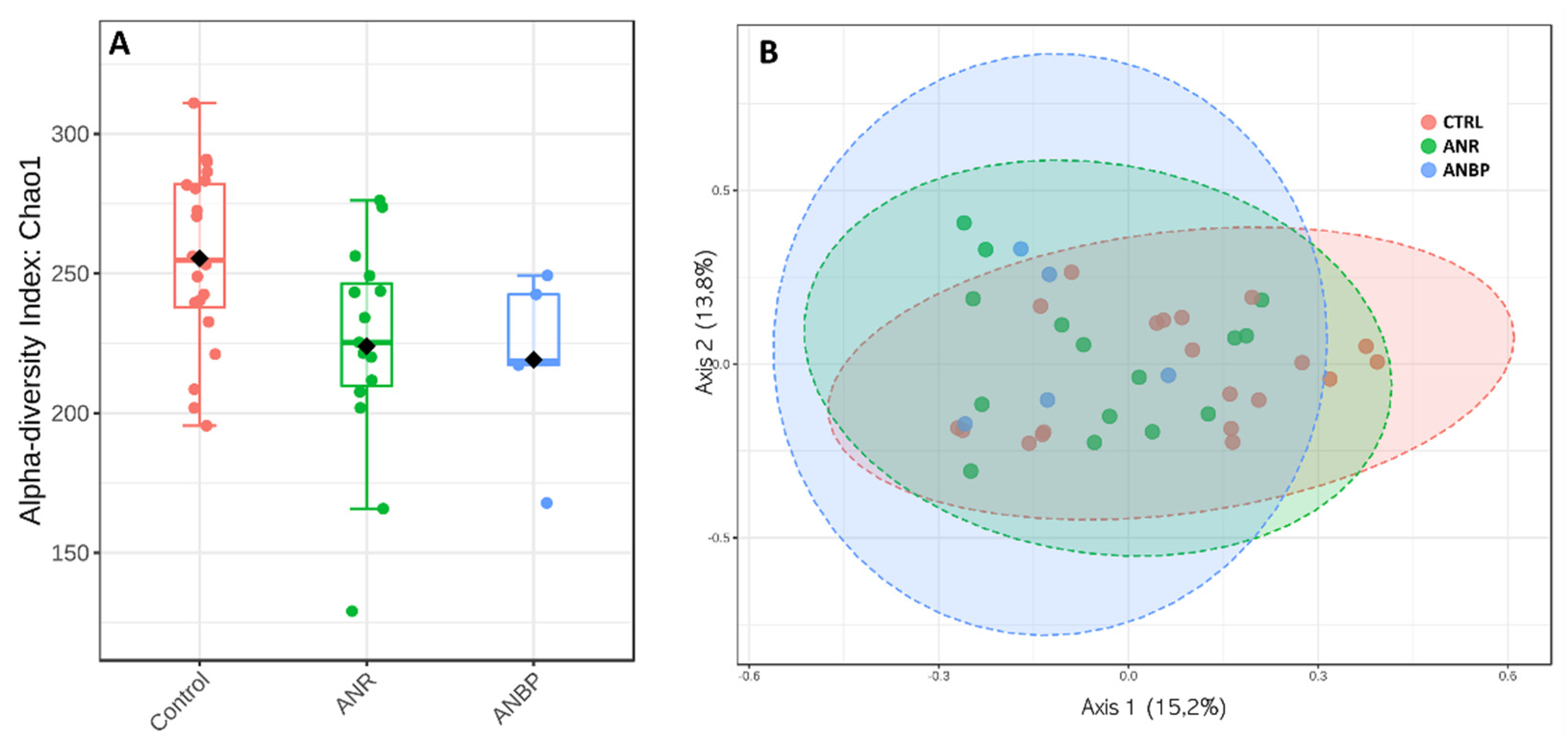
3.3. Microbiome
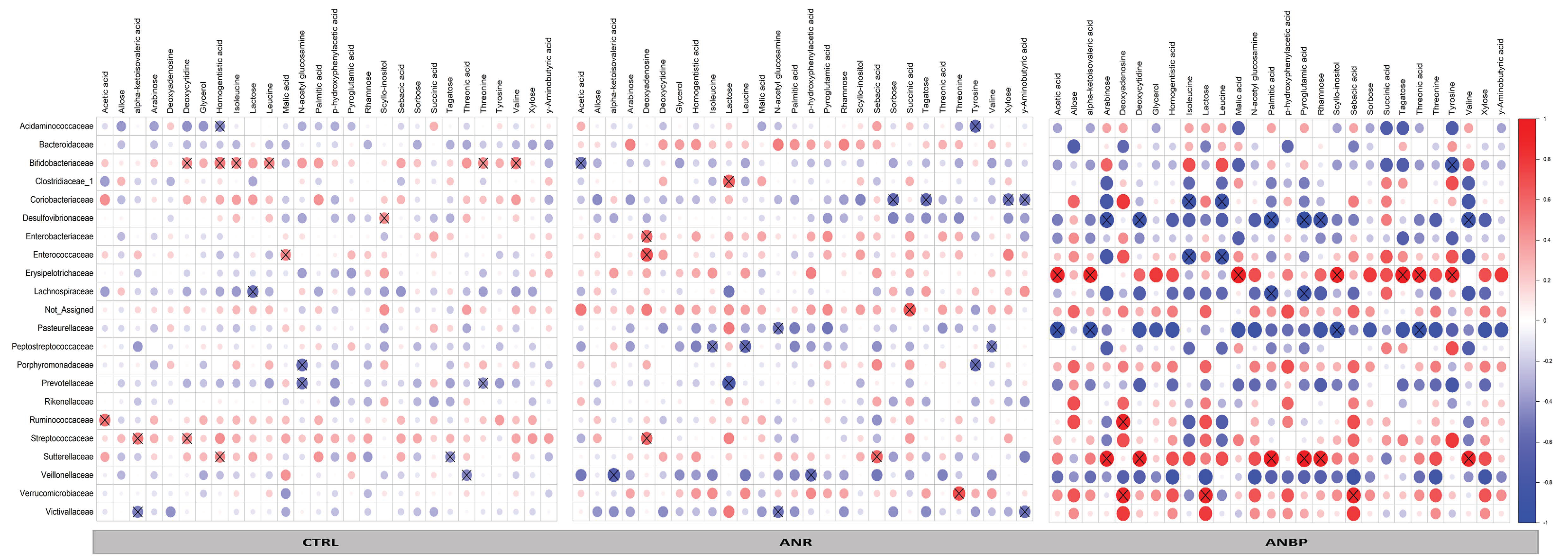
3.4. Microbiome-Metabolomics Integration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turnbaugh, P.J.; Gordon, J.I. The core gut microbiome, energy balance and obesity. J. Physiol. 2009, 587, 4153–4158. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Gut instincts: Microbiota as a key regulator of brain development, ageing and neurodegeneration. J. Physiol. 2017, 595, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Crumeyrolle-Arias, M.; Jaglin, M.; Bruneau, A.; Vancassel, S.; Cardona, A.; Daugé, V.; Naudon, L.; Rabot, S. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology 2014, 42, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Więdłocha, M.; Marcinowicz, P.; Janoska-Jaździk, M.; Szulc, A. Gut microbiota, kynurenine pathway and mental disorders—Review. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110145. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, M.; Jonkers, D.M.; Troost, F.J.; Roeselers, G.; Venema, K. In vitro characterization of the impact of different substrates on metabolite production, energy extraction and composition of gut microbiota from lean and obese subjects. PLoS ONE 2014, 9, e113864. [Google Scholar] [CrossRef] [Green Version]
- Treasure, J.; Duarte, T.A.; Schmidt, U. Eating disorders. Lancet 2020, 395, 899–911. [Google Scholar] [CrossRef]
- Di Lodovico, L.; Mondot, S.; Doré, J.; Mack, I.; Hanachi, M.; Gorwood, P. Anorexia nervosa and gut microbiota: A systematic review and quantitative synthesis of pooled microbiological data. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 106, 110114. [Google Scholar] [CrossRef]
- Seitz, J.; Dahmen, B.; Keller, L.; Herpertz-Dahlmann, B. Gut Feelings: How Microbiota Might Impact the Development and Course of Anorexia Nervosa. Nutrients 2020, 12, 3295. [Google Scholar] [CrossRef]
- Monteleone, P.; Monteleone, A.M.; Troisi, J.; Dalle Grave, R.; Corrivetti, G.; Calugi, S.; Scala, G.; Patriciello, G.; Zanetti, A.; Maj, M. Metabolomics signatures of acutely ill and short-term weight recovered women with anorexia nervosa. Mol. Psychiatry 2019. [Google Scholar] [CrossRef]
- Monteleone, A.M.; Troisi, J.; Fasano, A.; Dalle Grave, R.; Marciello, F.; Serena, G.; Calugi, S.; Scala, G.; Corrivetti, G.; Cascino, G.; et al. Multi-omics data integration in anorexia nervosa patients before and after weight regain: A microbiome-metabolomics investigation. Clin. Nutr. 2020. [Google Scholar] [CrossRef]
- APA. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Mack, I.; Cuntz, U.; Grämer, C.; Niedermaier, S.; Pohl, C.; Schwiertz, A.; Zimmermann, K.; Zipfel, S.; Enck, P.; Penders, J. Weight gain in anorexia nervosa does not ameliorate the faecal microbiota, branched chain fatty acid profiles and gastrointestinal complaints. Sci. Rep. 2016, 6, 26752. [Google Scholar] [CrossRef] [Green Version]
- Fairburn, C.; Cooper, Z. The Eating Disorders Examination In: Binge Eating: Nature, Assesment and Treatment; Fairburn, C.G., Wilson, G.T., Eds.; Guilford Press: New York, NY, USA, 1993; Volume 160, p. 192. [Google Scholar]
- Calugi, S.; Ricca, V.; Castellini, G.; Sauro, C.L.; Ruocco, A.; Chignola, E.; El Ghoch, M.; Dalle Grave, R. The eating disorder examination: Reliability and validity of the Italian version. Eat. Weight Dis. 2015, 20, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Derogatis, L.R. Brief Symptom Inventory (BSI) Administration, Scoring, and Procedures Manual; NCS Pearson: Minneap, MN, USA, 1993. [Google Scholar]
- De Leo, D.; Frisoni, G.B.; Rozzini, R.; Trabucchi, M. Italian community norms for the Brief Symptom Inventory in the elderly. Br. J. Clin. Psychol. 1993, 32, 209–213. [Google Scholar] [CrossRef] [PubMed]
- First, M.; Williams, J.; Karg, R.; Spitzer, R. Structured Clinical Interview for DSM-5 Disorders, Clinician Version (SCID-5-CV); Clinical Trials Version; American Psychiatric Association: Washington, DC, USA, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
- Albanese, D.; Fontana, P.; De Filippo, C.; Cavalieri, D.; Donati, C. MICCA: A complete and accurate software for taxonomic profiling of metagenomic data. Sci. Rep. 2015, 5, 9743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MICrobial Community Analysis. Available online: https://micca.org/ (accessed on 16 November 2020).
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Domingos, P. MetaCost: A general method for making classifiers cost-sensitive. In Proceedings of the Fifth ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Diego, CA, USA, 15–18 August 1999; pp. 155–164. [Google Scholar]
- Mitchell, M. An Introduction to Genetic Algorithms; MIT Press: Cambridge, MA, USA, 1998; ISBN 0-262-63185-7. [Google Scholar]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Kawai, K.; Yamashita, M.; Shoji, M.; Takakura, S.; Hata, T.; Nakashima, M.; Tatsushima, K.; Tanaka, K.; Sudo, N. Very long chain fatty acids are an important marker of nutritional status in patients with anorexia nervosa: A case control study. Biopsychosoc. Med. 2020, 17, 14. [Google Scholar] [CrossRef]
- Bulant, J.; Hill, M.; Velíková, M.; Yamamotová, A.; Martásek, P.; Papežová, H. Changes of BMI, steroid metabolome and psychopathology in patients with anorexia nervosa during hospitalization. Steroids 2020, 153, 108523. [Google Scholar] [CrossRef] [PubMed]
- Satogami, K.; Tseng, P.T.; Su, K.P.; Takahashi, S.; Ukai, S.; Li, D.J.; Chen, T.Y.; Lin, P.Y.; Chen, Y.W.; Matsuoka, Y.J. Relationship between polyunsaturated fatty acid and eating disorders: Systematic review and meta-analysis. Prostaglandins Leukot. Essent. Fatty Acids. 2019, 142, 11–19. [Google Scholar] [CrossRef]
- Shih, P.B.; Yang, J.; Morisseau, C.; German, J.B.; Zeeland, A.A.; Armando, A.M.; Quehenberger, O.; Bergen, A.W.; Magistretti, P.; Berrettini, W.; et al. Dysregulation of soluble epoxide hydrolase and lipidomic profiles in anorexia nervosa. Mol. Psychiatry 2016, 21, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Focker, M.; Timmesfeld, N.; Scherag, S.; Knoll, N.; Singmann, P.; Wang-Sattler, R.; Bühren, K.; Schwarte, R.; Egberts, K.; Fleischhaker, C.; et al. Comparison of metabolic profiles of acutely ill and short-term weight recovered patients with anorexia nervosa reveals alterations of 33 out of 163 metabolites. J. Psychiatr. Res. 2012, 46, 1600–1609. [Google Scholar] [CrossRef]
- Drewnowski, A.; Pierce, B.; Halmi, K.A. Fat aversion in eating disorders. Appetite 1988, 10, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Simon, Y.; Bellisle, F.; Monneuse, M.O.; Samuel-Lajeunesse, B.; Drewnowski, A. Taste responsiveness in anorexia nervosa. Br. J. Psychiatry 1993, 162, 244–246. [Google Scholar] [CrossRef]
- Jauregui Lobera, I.; Bolanos Rios, P. Choice of diet in patients with anorexia nervosa. Nutr. Hosp. 2009, 24, 682–687. [Google Scholar]
- Mörkl, S.; Lackner, S.; Müller, W.; Gorkiewicz, G.; Kashofer, K.; Oberascher, A.; Painold, A.; Holl, A.; Holzer, P.; Meinitzer, A.; et al. Gut microbiota and body composition in anorexia nervosa inpatients in comparison to athletes, overweight, obese, and normal weight controls. Int. J. Eat. Disord. 2017, 50, 1421–1431. [Google Scholar] [CrossRef]
- Kleiman, S.C.; Watson, H.J.; Bulik-Sullivan, E.C.; Huh, E.Y.; Tarantino, L.M.; Bulik, C.M.; Carroll, I.M. The Intestinal Microbiota in Acute Anorexia Nervosa and During Renourishment. Psychosom. Med. 2015, 77, 969–981. [Google Scholar] [CrossRef] [Green Version]
- Borgo, F.; Riva, A.; Benetti, A.; Casiraghi, M.C.; Bertelli, S.; Garbossa, S.; Anselmetti, S.; Scarone, S.; Pontiroli, A.E.; Morace, G.; et al. Microbiota in anorexia nervosa: The triangle between bacterial species, metabolites and psychological tests. PLoS ONE 2017, 12, e0179739. [Google Scholar] [CrossRef]
- Morita, C.; Tsuji, H.; Hata, T.; Gondo, M.; Takakura, S.; Kawai, K.; Yoshihara, K.; Ogata, K.; Nomoto, K.; Miyazaki, K.; et al. Gut Dysbiosis in Patients with Anorexia Nervosa. PLoS ONE 2015, 10, e0145274. [Google Scholar] [CrossRef] [Green Version]
- Wade, T.D. A retrospective comparison of purging type disorders, eating disorder not otherwise specified and bulimia nervosa. Int. J. Eat. Dis. 2007, 40, 1–6. [Google Scholar] [CrossRef]
- Nunez-Navarro, A.; Jimenez-Murcia, S.; Alvarez-Moya, E.; Villarejo, C.; Sanchez Diaz, I.; Masuet Augmantell, C.; Granero, R.; Penelo, E.; Krug, I.; Tinahones, F.J.; et al. Differentiating purging and nonpurging bulimia nervosa and binge eating disorder. Int. J. Eat. Dis. 2011, 44, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, A.M.; Monteleone, P.; Marciello, F.; Pellegrino, F.; Castellini, G.; Maj, M. Differences in Cortisol Awakening Response between Binge-Purging and Restrictive Patients with Anorexia Nervosa. Eur. Eat. Disord. Rev. 2017, 25, 13–18. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, N.C.; Frissen, M.N.; Davids, M.; Groen, A.K.; Nieuwdorp, M. Weight Gain after Fecal Microbiota Transplantation in a Patient with Recurrent Underweight following Clinical Recovery from Anorexia Nervosa. Psychother. Psychosom. 2019, 88, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Prochazkova, P.; Roubalova, R.; Dvorak, J.; Tlaskalova-Hogenova, H.; Cermakova, M.; Tomasova, P.; Sediva, B.; Kuzma, M.; Bulant, J.; Bilej, M.; et al. Microbiota, Microbial Metabolites, and Barrier Function in a Patient with Anorexia Nervosa after Fecal Microbiota Transplantation. Microorganisms 2019, 7, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Women with ANR (n = 17) | Women with ANBP (n = 6) | Healthy Women (n = 20) | F2,40 | p-Value | |
|---|---|---|---|---|---|
| Age, yrs | 20.5 ± 3.1 | 25.2 ± 5.2 | 23.0 ± 3.3 | 4.94 | 0.02 |
| Body weight, kg | 38.6 ± 6.0 | 39.8 ± 4.3 | 55.2 ± 6.3 | 39.59 | 3.29 × 10−10 |
| BMI | 15.0 ± 1.8 | 14.7 ± 1.5 | 20.3 ± 1.4 | 59.29 | 1.08 × 10−12 |
| EDE restraint | 4.5 ± 0.9 | 4.4 ± 1.2 | 0.2 ± 0.6 | 146.07 | 4.12 × 10−19 |
| EDE eating concern | 3.7 ± 1.1 | 3.8 ± 1.3 | 0.1 ± 0.4 | 91.61 | 1.17 × 10−15 |
| EDE weight concern | 4.2 ± 1.5 | 3.7 ± 0.8 | 0.3 ± 0.7 | 63.55 | 3.82 × 10−13 |
| EDE shape concern | 4.8 ± 1.3 | 4.7 ± 1.5 | 0.5 ±1.0 | 66.19 | 2.05 × 10−13 |
| BSI somatization | 1.8 ± 0.9 | 2.5 ± 1.1 | 0.1 ± 0.2 | 36.45 | 9.69 × 10−10 |
| BSI obsessive-compulsive | 2.2 ± 0.2 | 3.0 ± 0.3 | 0.3 ± 0.4 | 43.26 | 9.96 × 10−11 |
| BSI interpersonal sensitivity | 2.7 ± 0.8 | 3.3 ± 0.8 | 0.1 ± 0.2 | 116.57 | 2.05 × 10−17 |
| BSI depression | 2.7 ± 0.7 | 3.2 ± 0.5 | 0.2 ± 0.2 | 145.62 | 4.34 × 10−19 |
| BSI anxiety | 2.6 ± 0.8 | 3.3 ± 0.5 | 0.4 ± 0.2 | 93.17 | 8.83 × 10−16 |
| BSI hostility | 1.6 ± 0.8 | 2.4 ± 1.2 | 0.2 ± 0.3 | 27.80 | 2.69 × 10−8 |
| BSI phobic anxiety | 1.5 ± 1.0 | 1.9 ± 0.9 | 0.1 ± 0.1 | 24.78 | 9.94 × 10−8 |
| BSI paranoid ideation | 1.8 ± 0.9 | 2.3 ± 0.8 | 0.1 ± 0.3 | 36.55 | 9.37 × 10−10 |
| BSI psychoticism | 2.1 ± 0.8 | 2.6 ± 0.9 | 0.1 ± 0.2 | 64.62 | 2.95 × 10−13 |
| Metabolite | HMDB Code | ANR | ANBP | Selection Criteria |
|---|---|---|---|---|
| Acetic acid | HMDB0000042 | ↓ | ↑ | PLS-DA-2classes |
| Allose | HMDB0001151 | ↓ | ↑ | ANOVA |
| alpha-ketoisovaleric acid | HMDB0000019 | ↓ | ↑ | PL-SDA-3classes |
| Arabinose | HMDB0029942 | ↓ | ↓↓ | PLS-DA-3classes, PLS-DA-2classes |
| Deoxyadenosine | HMDB0000101 | ↓ | ↓↓ | PLS-DA-2classes |
| Deoxycytidine | HMDB0000014 | ↓ | ↓↓ | ANOVA, PLS-DA-3classes |
| Glycerol | HMDB0000131 | ↓ | ↑ | ANOVA, PLS-DA-3classes |
| Homogentistic acid | HMDB0000130 | ↓ | ↑ | ANOVA |
| Isoleucine | HMDB0000172 | ↓ | ↓↓ | ANOVA, PLS-DA-3classes |
| Lactose | HMDB0000186 | ↓ | ↑ | PLS-DA-2classes |
| Leucine | HMDB0000687 | ↓ | ↓↓ | PLS-DA-3classes |
| Malic acid | HMDB0000156 | ↓ | ↓↓ | ANOVA |
| N-acetyl glucosamine | HMDB0000803 | ↓↓ | ↓ | ANOVA, PLS-DA-3classes |
| Palmitic acid | HMDB0000220 | ↓↓ | ↓ | ANOVA |
| p-hydroxyphenylacetic acid | HMDB0000020 | ↓ | ↑ | ANOVA, PLS-DA-3classes |
| Pyroglutamic acid | HMDB0000267 | ↓ | ↓↓ | PLS-DA-2classes |
| Rhamnose | HMDB0000849 | ↓ | ↓↓ | ANOVA, PLS-DA-3classes PLSDA-2classes |
| Scyllo-inositol | HMDB0006088 | ↓ | ↑ | PLS-DA-2classes |
| Sebacic acid | HMDB0000792 | ↓ | ↑ | PLS-DA-2classes |
| Sorbose | HMDB0001266 | ↓↓ | ↓ | ANOVA |
| Succinic acid | HMDB0000254 | ↓ | ↑ | ANOVA, PLS-DA-2classes |
| Tagatose | HMDB0003418 | ↓↓ | ↓ | ANOVA, PLS-DA-3classes |
| Threonic acid | HMDB0000943 | ↓ | ↑ | PLS-DA-2classes |
| Threonine | HMDB0000167 | ↓ | ↑ | ANOVA |
| Tyrosine | HMDB0000158 | ↓ | ↑ | PLS-DA-3classes |
| Valine | HMDB0000883 | ↓ | ↓↓ | PLS-DA-3classes |
| Xylose | HMDB0000098 | ↓ | ↓↓ | ANOVA, PLS-DA-3classes PLSDA-2classes |
| γ-Aminobutyric acid | HMDB0000112 | ↓ | ↓ | PLS-DA-2classes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteleone, A.M.; Troisi, J.; Serena, G.; Fasano, A.; Dalle Grave, R.; Cascino, G.; Marciello, F.; Calugi, S.; Scala, G.; Corrivetti, G.; et al. The Gut Microbiome and Metabolomics Profiles of Restricting and Binge-Purging Type Anorexia Nervosa. Nutrients 2021, 13, 507. https://doi.org/10.3390/nu13020507
Monteleone AM, Troisi J, Serena G, Fasano A, Dalle Grave R, Cascino G, Marciello F, Calugi S, Scala G, Corrivetti G, et al. The Gut Microbiome and Metabolomics Profiles of Restricting and Binge-Purging Type Anorexia Nervosa. Nutrients. 2021; 13(2):507. https://doi.org/10.3390/nu13020507
Chicago/Turabian StyleMonteleone, Alessio Maria, Jacopo Troisi, Gloria Serena, Alessio Fasano, Riccardo Dalle Grave, Giammarco Cascino, Francesca Marciello, Simona Calugi, Giovanni Scala, Giulio Corrivetti, and et al. 2021. "The Gut Microbiome and Metabolomics Profiles of Restricting and Binge-Purging Type Anorexia Nervosa" Nutrients 13, no. 2: 507. https://doi.org/10.3390/nu13020507
APA StyleMonteleone, A. M., Troisi, J., Serena, G., Fasano, A., Dalle Grave, R., Cascino, G., Marciello, F., Calugi, S., Scala, G., Corrivetti, G., & Monteleone, P. (2021). The Gut Microbiome and Metabolomics Profiles of Restricting and Binge-Purging Type Anorexia Nervosa. Nutrients, 13(2), 507. https://doi.org/10.3390/nu13020507