Obese Vegetarians and Omnivores Show Different Metabolic Changes: Analysis of 1340 Individuals

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Aspects
2.2. Sample
2.3. Sample Classification Regarding Eating Groups
2.4. Sample Classification Regarding Nutritional Status
2.5. Analysis of Markers Associated with BMI
2.6. Statistical Analysis
3. Results
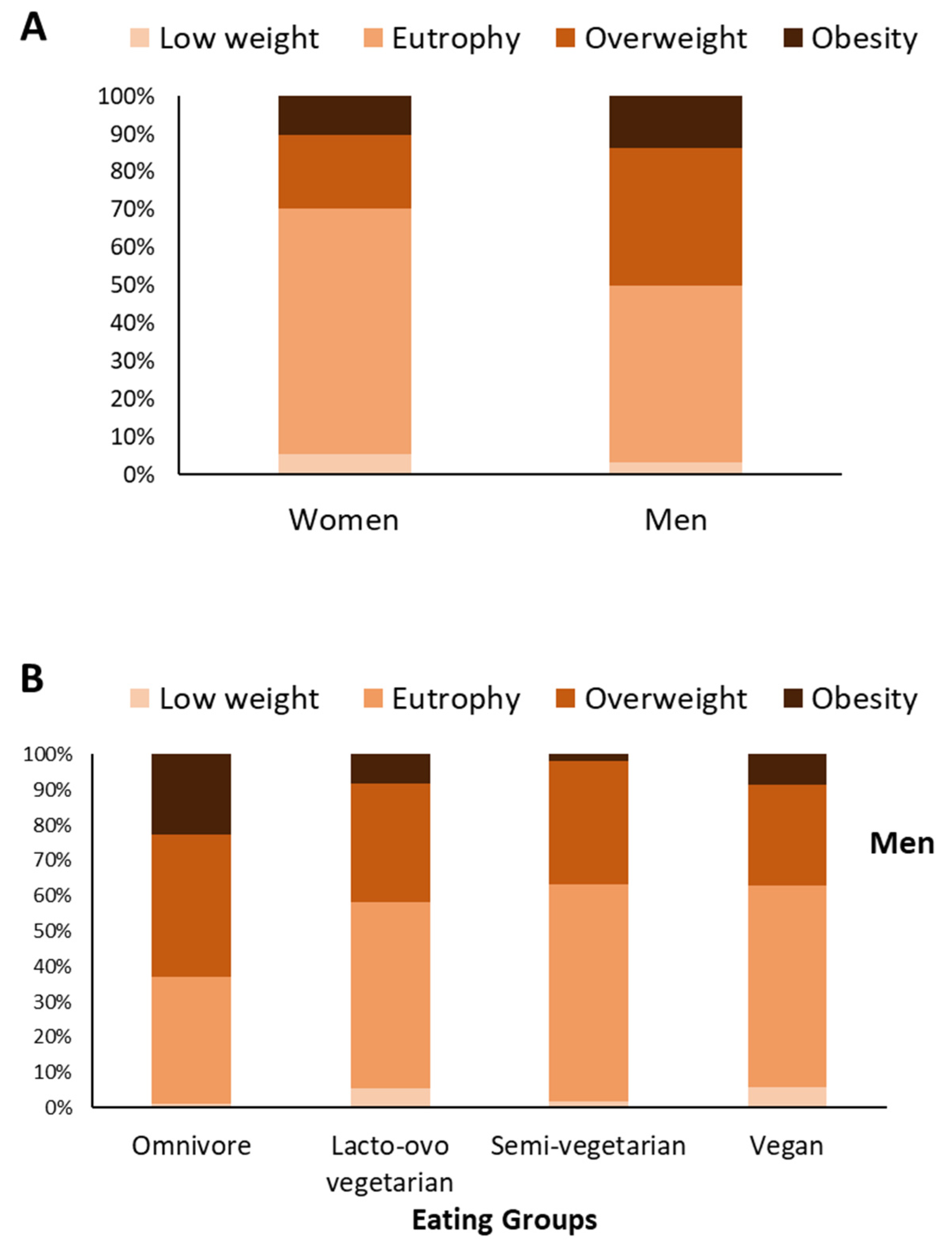
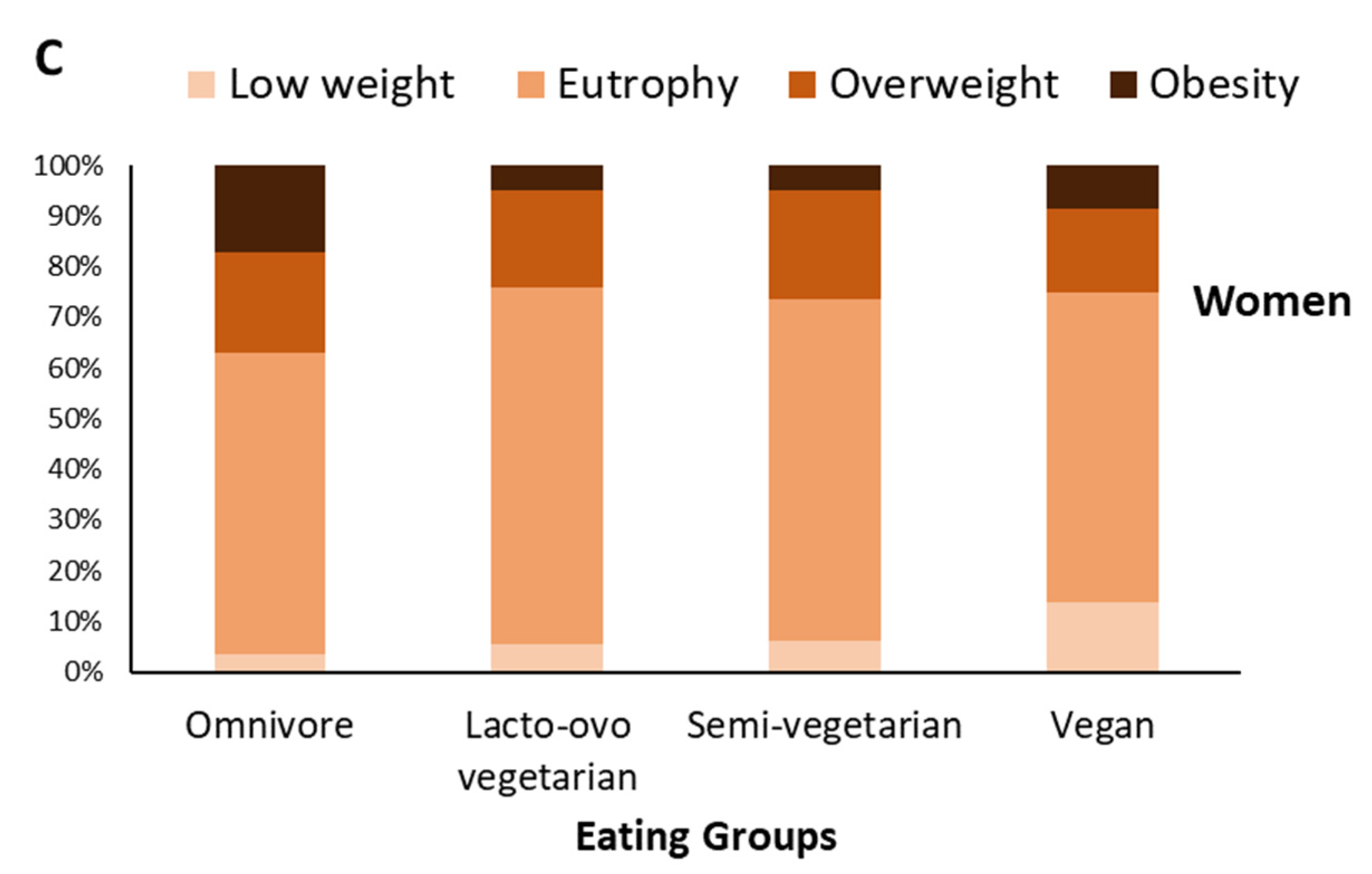
3.1. Omnivorous Men and Women Have a Higher Prevalence of Obesity; Vegan Women Have a Higher Prevalence of Underweight
3.2. Alcohol Consumption Did Not Influence Liver Enzyme Levels, as Physical Activity Did Not Influence HOMA-IR Levels
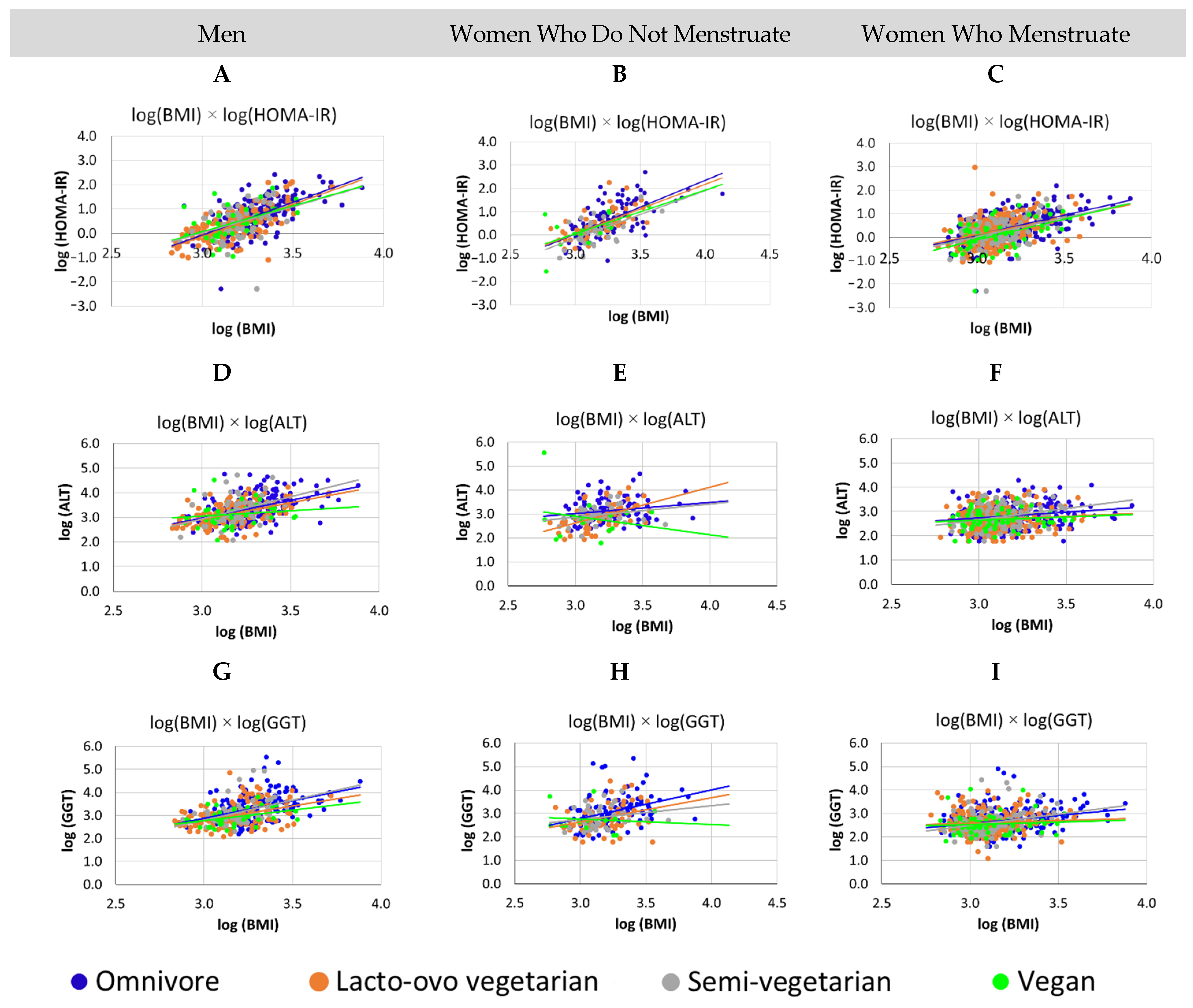
3.3. For All Groups, Increased BMI Was Associated with Worse Metabolism, Men Have Higher Levels of GGT and HOMA-IR Than Women, for Vegan Women, Who Do Not Menstruate, the Higher the BMI, the Lower the ALT
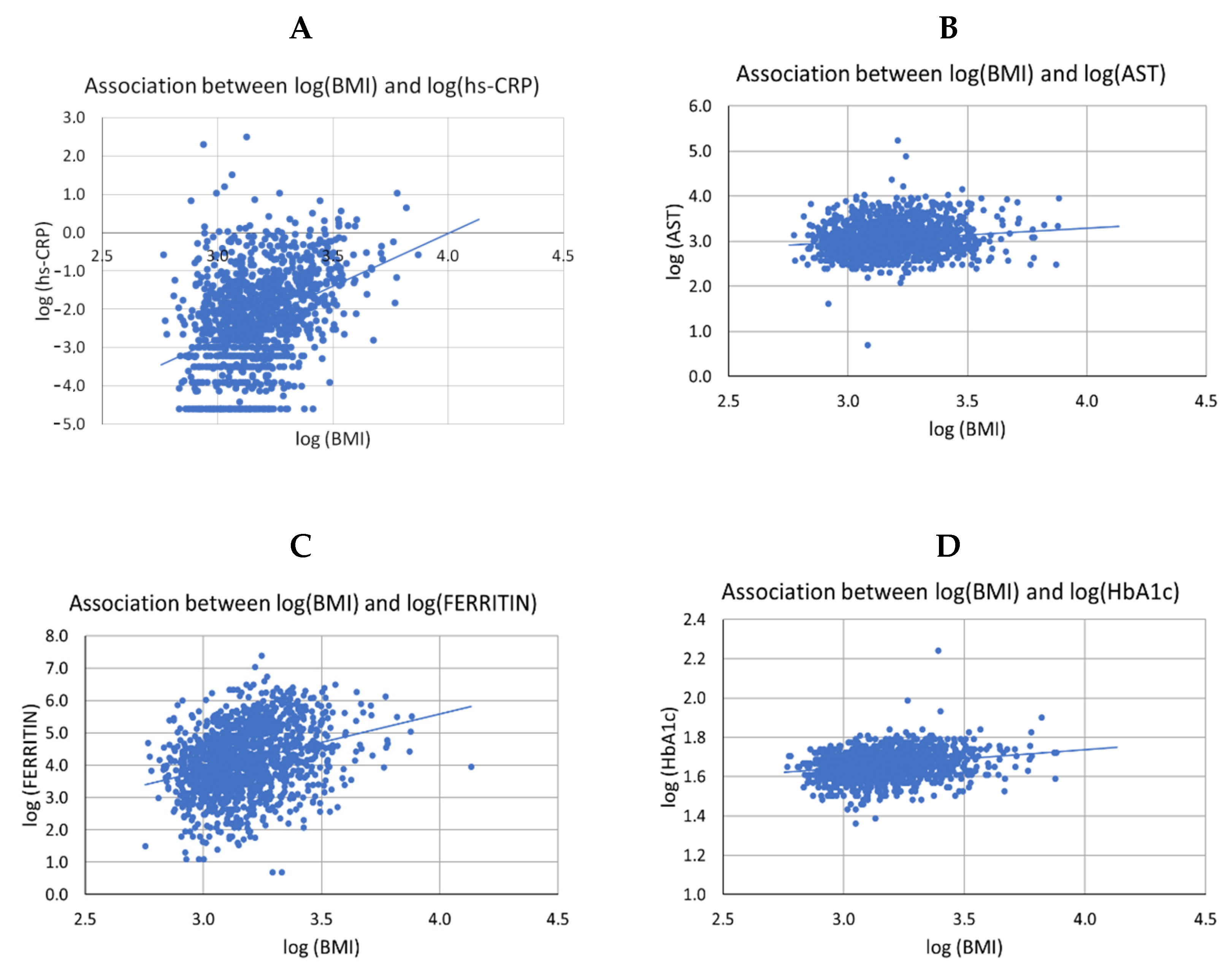
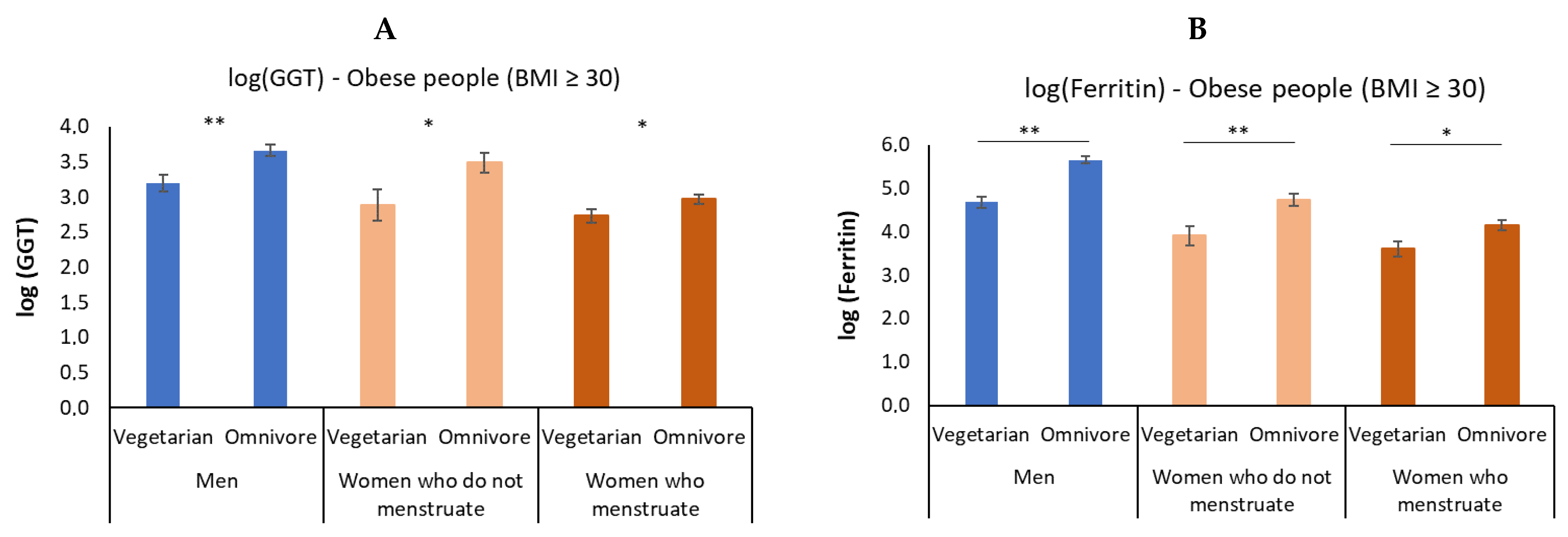
3.4. In Obesity, Vegetarian Individuals Have a Lower Elevation of GGT and Ferritin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- The Global BMI Mortality Collaboration. Body-mass index and all-cause mortality: Individual-participant-data meta-analysis of 239 prospec-tive studies in four continents. Lancet 2016, 388, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Favaretto, F.; Bettini, S.; Busetto, L.; Milan, G.; Vettor, R. Adipogenic progenitors in different organs: Pathophysiological implications. Rev. Endocr. Metab. Disord. 2021, 23, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Rao, H.; Liu, F.; Wei, L.; Li, H.; Wu, C. Recent Advances in Adipose Tissue Dysfunction and Its Role in the Pathogenesis of Non-Alcoholic Fatty Liver Disease. Cells 2021, 10, 3300. [Google Scholar] [CrossRef]
- Albhaisi, S.; Noureddin, M. Current and Potential Therapies Targeting Inflammation in NASH. Front. Endocrinol. 2021, 12, 767314. [Google Scholar] [CrossRef] [PubMed]
- Aragon, G.; Younossi, Z.M. When and how to evaluate mildly elevated liver enzymes in apparently healthy patients. Clevel. Clin. J. Med. 2010, 77, 195–204. [Google Scholar] [CrossRef]
- Rahmani, J.; Miri, A.; Namjoo, I.; Zamaninour, N.; Maljaei, M.B.; Zhou, K.; Cerneviciute, R.; Mousavi, S.M.; Varkaneh, H.K.; Salehisahlabadi, A.; et al. Elevated liver enzymes and cardiovascular mortality: A systematic review and dose-response meta-analysis of more than one million participants. Eur. J. Gastroenterol. Hepatol. 2019, 31, 555–562. [Google Scholar] [CrossRef]
- Kunutsor, S.K.; Apekey, T.A.; Seddoh, D.; Walley, J. Liver enzymes and risk of all-cause mortality in general populations: A systematic review and meta-analysis. Int. J. Epidemiol. 2014, 43, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.B. Gamma glutamyl transferase. Crit. Rev. Clin. Lab. Sci. 2001, 38, 263–355. [Google Scholar] [CrossRef]
- Targher, G. Elevated serum gamma-glutamyltransferase activity is associated with increased risk of mortality, incident type 2 diabetes, cardiovascular events, chronic kidney disease and cancer—A narrative review. Clin. Chem. Lab. Med. 2010, 48, 147–157. [Google Scholar] [CrossRef]
- Ndrepepa, G.; Kastrati, A. Gamma-glutamyl transferase and cardiovascular disease. Ann. Transl. Med. 2016, 4, 481. [Google Scholar] [CrossRef]
- Paolicchi, A.; Dominici, S.; Pieri, L.; Maellaro, E.; Pompella, A. Glutathione catabolism as a signaling mechanism. Biochem. Pharmacol. 2002, 64, 1027–1035. [Google Scholar] [CrossRef]
- Ndrepepa, G.; Colleran, R.; Kastrati, A. Gamma-glutamyl transferase and the risk of atherosclerosis and coronary heart disease. Clin. Chim. Acta 2018, 476, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-W.; Li, M.; Hou, W.-S.; Li, K.; Zhou, J.-R.; Tang, Z.-Y. Association between Gamma-Glutamyltransferase Level and Risk of Stroke: A Systematic Review and Meta-analysis of Prospective Studies. J. Stroke Cerebrovasc. Dis. 2015, 24, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Kunutsor, S.K.; Apekey, T.A.; Cheung, B.M. Gamma-glutamyltransferase and risk of hypertension: A systematic review and dose-response meta-analysis of prospective evidence. J. Hypertens. 2015, 33, 2373–2381. [Google Scholar] [CrossRef] [Green Version]
- Kunutsor, S.K.; Apekey, T.A.; Seddoh, D. Gamma glutamyltransferase and metabolic syndrome risk: A systematic review and dose-response meta-analysis. Int. J. Clin. Pract. 2014, 69, 136–144. [Google Scholar] [CrossRef]
- Kunutsor, S.K.; Abbasi, A.; Adler, A.I. Gamma-glutamyl transferase and risk of type II diabetes: An updated systematic review and dose-response meta-analysis. Ann. Epidemiol. 2014, 24, 809–816. [Google Scholar] [CrossRef]
- Liu, C.-F.; Gu, Y.-T.; Wang, H.-Y.; Fang, N.-Y. Gamma-Glutamyltransferase Level and Risk of Hypertension: A Systematic Review and Meta-Analysis. PLoS ONE 2012, 7, e48878. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-F.; Zhou, W.-N.; Fang, N.-Y. Gamma-glutamyltransferase levels and risk of metabolic syndrome: A meta-analysis of prospective cohort studies. Int. J. Clin. Pract. 2012, 66, 692–698. [Google Scholar] [CrossRef]
- Long, Y.; Zeng, F.; Shi, J.; Tian, H.; Chen, T. Gamma-glutamyltransferase predicts increased risk of mortality: A systematic review and meta-analysis of prospective observational studies. Free Radic. Res. 2014, 48, 716–728. [Google Scholar] [CrossRef]
- Bataller-Sifre, R.; Guiral-Olivan, V.; Bataller-Alberola, L. New clinical and toxicological scenario of gammaglutamyltranspepti-dase. Rev. Esp. Enferm. Dig. 2011, 103, 586–590. [Google Scholar] [CrossRef] [Green Version]
- Slywitch, E.; Savalli, C.; Duarte, A.C.G.; Escrivão, M.A.M.S. Iron Deficiency in Vegetarian and Omnivorous Individuals: Analysis of 1340 Individuals. Nutrients 2021, 13, 2964. [Google Scholar] [CrossRef]
- Uysal, S.; Armutcu, F.; Aydogan, T.; Akin, K.; Ikizek, M.; Yigitoglu, M.R. Some inflammatory cytokine levels, iron metabolism and oxidan stress markers in subjects with nonalcoholic steatohepatitis. Clin. Biochem. 2011, 44, 1375–1379. [Google Scholar] [CrossRef]
- Fraser, G.; Katuli, S.; Anousheh, R.; Knutsen, S.; Herring, P.; Fan, J. Vegetarian diets and cardiovascular risk factors in black members of the Adventist Health Study-2. Public Health Nutr. 2014, 18, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satija, A.; Bhupathiraju, S.N.; Rimm, E.B.; Spiegelman, D.; Chiuve, S.; Borgi, L.; Willett, W.C.; Manson, J.E.; Sun, Q.; Hu, F.B. Plant-Based Dietary Patterns and Incidence of Type 2 Diabetes in US Men and Women: Results from Three Prospective Cohort Studies. PLoS Med. 2016, 13, e1002039. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, Y.; Barnard, N.D.; Levin, S.M.; Watanabe, M. Vegetarian diets and glycemic control in diabetes: A systematic review and meta-analysis. Cardiovasc. Diagn. Ther. 2014, 4, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Craig, W.J. Health effects of vegan diets. Am. J. Clin. Nutr. 2009, 89, 1627S–1633S. [Google Scholar] [CrossRef] [Green Version]
- Barnard, N.D.; Cohen, J.; Jenkins, D.J.; Turner-McGrievy, G.; Gloede, L.; Jaster, B.; Seidl, K.; Green, A.A.; Talpers, S. A Low-Fat Vegan Diet Improves Glycemic Control and Cardiovascular Risk Factors in a Randomized Clinical Trial in Individuals with Type 2 Diabetes. Diabetes Care 2006, 29, 1777–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, N.D.; Katcher, H.I.; Jenkins, D.J.; Cohen, J.; Turner-McGrievy, G. Vegetarian and vegan diets in type 2 diabetes management. Nutr. Rev. 2009, 67, 255–263. [Google Scholar] [CrossRef]
- Kahleova, H.; Matoulek, M.; Malinska, H.; Oliyarnik, O.; Kazdova, L.; Neskudla, T.; Skoch, A.; Hajek, M.; Hill, M.; Kahle, M.; et al. Vegetarian diet improves insulin resistance and oxidative stress markers more than conventional diet in subjects with Type 2 diabetes. Diabet. Med. 2011, 28, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Dinu, M.; Abbate, R.; Gensini, G.F.; Casini, A.; Sofi, F. Vegetarian, vegan diets and multiple health outcomes: A systematic review with meta-analysis of observa-tional studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3640–3649. [Google Scholar] [CrossRef]
- Ornish, D.; Scherwitz, L.W.; Billings, J.H.; Gould, K.L.; Merritt, T.A.; Sparler, S.; Armstrong, W.T.; Ports, T.A.; Kirkeeide, R.L.; Hogeboom, C.; et al. Intensive Lifestyle Changes for Reversal of Coronary Heart Disease. JAMA J. Am. Med. Assoc. 1998, 280, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.; Newman, J.D.; Woolf, K.; Ganguzza, L.; Guo, Y.; Allen, N.; Zhong, J.; Fisher, E.A.; Slater, J. Anti-Inflammatory Effects of a Vegan Diet Versus the American Heart Association–Recommended Diet in Coronary Artery Disease Trial. J. Am. Heart Assoc. 2018, 7, e011367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trepanowski, J.F.; Varady, K.A. Veganism Is a Viable Alternative to Conventional Diet Therapy for Improving Blood Lipids and Glycemic Control. Crit. Rev. Food Sci. Nutr. 2014, 55, 2004–2013. [Google Scholar] [CrossRef] [PubMed]
- Orlich, M.J.; Fraser, G.E. Vegetarian diets in the Adventist Health Study 2: A review of initial published findings. Am. J. Clin. Nutr. 2014, 100, 353S–358S. [Google Scholar] [CrossRef] [Green Version]
- WHO. World Health Organization Obesity Preventing and Managing the Global Epidemic; Report of a WHO Consultation on Obesity; WHO: Geneva, Switzerland, 1997.
- Rosner, B. Fundamentals of Biostatistics, 2nd ed.; PWS Publishers: Boston, MA, USA, 1986; pp. 442–480. [Google Scholar]
- Oussalah, A.; Levy, J.; Berthezène, C.; Alpers, D.H.; Guéant, J.L. Health outcomes associated with vegetarian diets: An umbrella review of systematic reviews and meta-analyses. Clin. Nutr. 2020, 39, 3283–3307. [Google Scholar] [CrossRef]
- Turner-McGrievy, G.M.; Davidson, C.; Wingard, E.E.; Wilcox, S.; Frongillo, E.A. Comparative effectiveness of plant-based diets for weight loss: A randomized controlled trial of five different diets. Nutrition 2015, 31, 350–358. [Google Scholar] [CrossRef]
- Kahleova, H.; Petersen, K.F.; Shulman, G.I.; Alwarith, J.; Rembert, E.; Tura, A.; Hill, M.; Holubkov, R.; Barnard, N.D. Effect of a Low-Fat Vegan Diet on Body Weight, Insulin Sensitivity, Postprandial Metabolism, and In-tramyocellular and Hepatocellular Lipid Levels in Overweight Adults: A Randomized Clinical Trial. JAMA Netw. Open 2020, 3, e2025454. [Google Scholar] [CrossRef]
- Barnard, N.D.; Alwarith, J.; Rembert, E.; Brandon, L.; Nguyen, M.; Goergen, A.; Horne, T.; Nascimento, G.F.D.; Lakkadi, K.; Tura, A.; et al. A Mediterranean Diet and Low-Fat Vegan Diet to Improve Body Weight and Cardiometabolic Risk Factors: A Randomized, Cross-over Trial. J. Am. Nutr. Assoc. 2021, 41, 127–139. [Google Scholar] [CrossRef]
- Medawar, E.; Huhn, S.; Villringer, A.; Witte, A.V. The effects of plant-based diets on the body and the brain: A systematic review. Transl. Psychiatry 2019, 9, 226. [Google Scholar] [CrossRef]
- Jung, J.Y.; Shim, J.-J.; Park, S.K.; Ryoo, J.-H.; Choi, J.-M.; Oh, I.-H.; Jung, K.-W.; Cho, H.; Ki, M.; Won, Y.-J.; et al. Serum ferritin level is associated with liver steatosis and fibrosis in Korean general population. Hepatol. Int. 2018, 13, 222–233. [Google Scholar] [CrossRef]
- Sun, L.; Zong, G.; Pan, A.; Ye, X.; Li, H.; Yu, Z.; Zhao, Y.; Zou, S.; Yu, D.; Jin, Q.; et al. Elevated Plasma Ferritin Is Associated with Increased Incidence of Type 2 Diabetes in Middle-Aged and Elderly Chinese Adults. J. Nutr. 2013, 143, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Harvey, L.J.; Armah, C.N.; Dainty, J.R.; Foxall, R.J.; Lewis, D.J.; Langford, N.J.; Fairweather-Tait, S.J. Impact of menstrual blood loss and diet on iron deficiency among women in the UK. Br. J. Nutr. 2005, 94, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, A.L.M.; Skeaff, C.M.; Williams, S.; Gibson, R.S. The role of blood loss and diet in the aetiology of mild iron deficiency in premenopausal adult New Zea-land women. Public Health Nutr. 2001, 4, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, S.C.; Mahida, V.I.; Bhatti, A. Iron status of regular voluntary blood donors. Asian J. Transfus. Sci. 2008, 2, 9–12. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, L.-Q.; Liu, A.-P.; Wang, P.-Y. Prevalence of Risk Factors for Cardiovascular Disease and Their Associations with Diet and Physical Activity in Suburban Beijing, China. J. Epidemiol. 2010, 20, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Le, L.T.; Sabaté, J. Beyond Meatless, the Health Effects of Vegan Diets: Findings from the Adventist Cohorts. Nutrients 2014, 6, 2131–2147. [Google Scholar] [CrossRef] [Green Version]
- Glick-Bauer, M.; Yeh, M.-C. The Health Advantage of a Vegan Diet: Exploring the Gut Microbiota Connection. Nutrients 2014, 6, 4822–4838. [Google Scholar] [CrossRef] [Green Version]
- Ruengsomwong, S.; La-Ongkham, O.; Jiang, J.; Wannissorn, B.; Nakayama, J.; Nitisinprasert, S. Microbial Community of Healthy Thai Vegetarians and Non-Vegetarians, Their Core Gut Microbiota, and Pathogen Risk. J. Microbiol. Biotechnol. 2016, 26, 1723–1735. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.-W.; Lin, S.-X.; Shen, Z.-H.; Luo, W.-W.; Wang, X.-Y. Systematic Review with Meta-Analysis: The Effects of Probiotics in Nonalcoholic Fatty Liver Disease. Gastroenterol. Res. Pract. 2019, 2019, 1484598. [Google Scholar] [CrossRef]
- Loman, B.R.; Hernández-Saavedra, D.; An, R.; Rector, R.S. Prebiotic and probiotic treatment of nonalcoholic fatty liver disease: A systematic review and meta-analysis. Nutr. Rev. 2018, 76, 822–839. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, H.K.; Bae, S.J.; Park, J.Y.; Lee, K.U. Association of elevated serum ferritin concentration with insulin resistance and impaired glucose metabolism in Korean men and women. Metabolism 2011, 60, 414–420. [Google Scholar] [CrossRef]
- Hua, N.W.; Stoohs, R.A.; Facchini, F.S. Low iron status and enhanced insulin sensitivity in lacto-ovo vegetarians. Br. J. Nutr. 2001, 86, 515–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orban, E.; Schwab, S.; Thorand, B.; Huth, C. Association of iron indices and type 2 diabetes: A meta-analysis of observational studies. Diabetes Metab. Res. Rev. 2014, 30, 372–394. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.J.; Wang, L.; Zhuang, Z.J.; Yang, W.J.; Ding, J.P.; Shi, J.P. Studying the correlation between ferritin and non-alcoholic fatty liver disease. Zhonghua Gan Zang Bing Za Zhi 2021, 29, 1089–1094. [Google Scholar] [PubMed]
- Mousavi, S.R.M.; Geramizadeh, B.; Anushiravani, A.; Ejtehadi, F.; Anbardar, M.H.; Moini, M. Correlation between Serum Ferritin Level and Histopathological Disease Severity in Non-alcoholic Fatty Liver Disease. Middle East J. Dig. Dis. 2018, 10, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Koenig, G.; Seneff, S. Gamma-Glutamyltransferase: A Predictive Biomarker of Cellular Antioxidant Inadequacy and Disease Risk. Dis. Markers 2015, 2015, 818570. [Google Scholar] [CrossRef] [Green Version]
- Kunutsor, S.K.; Apekey, T.A.; Van Hemelrijck, M.; Calori, G.; Perseghin, G. Gamma glutamyltransferase, alanine aminotransferase and risk of cancer: Systematic review and me-ta-analysis. Int. J. Cancer 2015, 136, 1162–1170. [Google Scholar] [CrossRef]
- Wannamethee, G.; Ebrahim, S.; Shaper, A.G. Gamma-glutamyltransferase: Determinants and Association with Mortality from Ischemic Heart Disease and All Causes. Am. J. Epidemiol. 1995, 142, 699–708. [Google Scholar] [CrossRef]
- Suh, Y.J.; Park, S.K.; Choi, J.-M.; Ryoo, J.-H. The clinical importance of serum γ-glutamyltransferase level as an early predictor of obesity development in Korean men. Atherosclerosis 2013, 227, 437–441. [Google Scholar] [CrossRef]
- Yang, P.; Wu, P.; Liu, X.; Feng, J.; Zheng, S.; Wang, Y.; Fan, Z. Association Between γ-Glutamyltransferase Level and Cardiovascular or All-Cause Mortality in Patients With Coronary Artery Disease: A Systematic Review and Meta-Analysis. Angiology 2019, 70, 844–852. [Google Scholar] [CrossRef]
- Du, G.; Song, Z.; Zhang, Q. Gamma-glutamyltransferase is associated with cardiovascular and all-cause mortality: A meta-analysis of prospective cohort studies. Prev. Med. 2013, 57, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.E.; Kinter, M.T.; Oberley, T.D.; Freeman, M.L.; Frierson, H.F.; Ridnour, L.A.; Tao, Y.; Oberley, L.W.; Spitz, D.R. Enhanced gamma-glutamyl transpeptidase expression and selective loss of CuZn superoxide dismutase in hepatic iron overload. Free Radic. Biol. Med. 1998, 24, 545–555. [Google Scholar] [CrossRef]
- Wei, D.; Chen, T.; Gao, Y.; Tian, H. Serum Gamma-Glutamyltransferase and Ferritin are Related to Insulin Resistance: A Population-Based Study. Clin. Lab. 2015, 61, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ren, Y.; Liu, Y.; Long, Y.; Zhang, X.; Yu, H.; Xu, J.; Yu, T.; Tian, H. Serum gamma-glutamyl transferase, ferritin and the risk of type 2 diabetes in women from a Chinese minority. Diabetes Res. Clin. Pract. 2010, 90, 352–357. [Google Scholar] [CrossRef]
- Wei, D.; Chen, T.; Li, J.; Gao, Y.; Ren, Y.; Zhang, X.; Yu, H.; Tian, H. Association of Serum Gamma-Glutamyl Transferase and Ferritin with the Metabolic Syndrome. J. Diabetes Res. 2015, 2015, 741731. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Ren, Y.; Gao, Y.; Tian, H. Serum Gamma-Glutamyl Transferase and Ferritin Synergistically Associated with the Rate of Chronic Kidney Disease. Dis. Markers 2017, 2017, 9765259. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Evans, J.C.; Robins, S.J.; Wilson, P.W.; Albano, I.; Fox, C.S.; Wang, T.J.; Benjamin, E.J.; D’Agostino, R.B.; Vasan, R.S. Gamma Glutamyl Transferase and Metabolic Syndrome, Cardiovascular Disease, and Mortality Risk. Arter. Thromb. Vasc. Biol. 2007, 27, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, E.; Priya, R.; Nanda, R.; Patel, S. Serum GGT and serum ferritin as early markers for metabolic syndrome. J. Fam. Med. Prim. Care 2020, 9, 3458–3463. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- Kažimírová, A.; Barančoková, M.; Volkovová, K.; Staruchová, M.; Krajčovičová-Kudláčková, M.; Wsólová, L.; Collins, A.R.; Dušinská, M. Does a vegetarian diet influence genomic stability? Eur. J. Nutr. 2004, 43, 32–38. [Google Scholar] [CrossRef]
- Manjari, V.; Suresh, Y.; Devi, M.S.; Das, U. Oxidant stress, anti-oxidants and essential fatty acids in South Indian vegetarians and non-vegetarians. Prostaglandins Leukot. Essent. Fat. Acids 2001, 64, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Szeto, Y.T.; Kwok, T.C.; Benzie, I.F. Effects of a long-term vegetarian diet on biomarkers of antioxidant status and cardi-ovascular disease risk. Nutrition 2004, 20, 863–866. [Google Scholar] [CrossRef]
- Kim, M.K.; Cho, S.W.; Park, Y.K. Long-term vegetarians have low oxidative stress, body fat, and cholesterol levels. Nutr. Res. Pract. 2012, 6, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.H. A New Threat to Public Health: Organochlorines and Food. Nutr. Health 1992, 8, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Devoto, E.; Kohlmeier, L.; Heeschen, W. Some dietary predictors of plasma organochlorine concentrations in an elderly German population. Arch. Environ. Health Int. J. 1998, 53, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Battu, R.; Singh, B.; Kang, B.; Joia, B. Risk assessment through dietary intake of total diet contaminated with pesticide residues in Punjab, India, 1999–2002. Ecotoxicol. Environ. Saf. 2005, 62, 132–139. [Google Scholar] [CrossRef]
- Van Audenhaege, M.; Héraud, F.; Ménard, C.; Bouyrie, J.; Morois, S.; Calamassi-Tran, G.; Lesterle, S.; Volatier, J.-L.; Leblanc, J.-C. Impact of food consumption habits on the pesticide dietary intake: Comparison between a French vegetarian and the general population. Food Addit. Contam. Part A 2009, 26, 1372–1388. [Google Scholar] [CrossRef]
- Baudry, J.; Rebouillat, P.; Allès, B.; Cravedi, J.-P.; Touvier, M.; Hercberg, S.; Lairon, D.; Vidal, R.; Kesse-Guyot, E. Estimated dietary exposure to pesticide residues based on organic and conventional data in omnivores, pesco-vegetarians, vegetarians and vegans. Food Chem. Toxicol. 2021, 153, 112179. [Google Scholar] [CrossRef]
- Berman, T.; Göen, T.; Novack, L.; Beacher, L.; Grinshpan, L.; Segev, D.; Tordjman, K. Urinary concentrations of organophosphate and carbamate pesticides in residents of a vegetarian community. Environ. Int. 2016, 96, 34–40. [Google Scholar] [CrossRef]
- Dickman, M.; Leung, C.; Leong, M. Hong Kong male subfertility links to mercury in human hair and fish. Sci. Total Environ. 1998, 214, 165–174. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eating Habit | Men | Women Who Do Not Menstruate | Women Who Menstruate | ||||||
|---|---|---|---|---|---|---|---|---|---|
| HOMA-IR | GGT | ALT | HOMA-IR | GGT | ALT | HOMA-IR | GGT | ALT | |
| ONV | 2.70% | 1.53% | 1.45% | 2.30% | 1.22% | 0.47% | 1.70% | 0.70% | 0.47% |
| LOV | 2.67% | 1.24% | 1.35% | 2.11% | 1.02% | 1.50% | 1.45% | 0.22% | 0.30% |
| SMV | 2.16% | 1.73% | 1.79% | 2.05% | 0.58% | 0.55% | 1.63% | 0.94% | 0.92% |
| VGN | 2.13% | 0.90% | 0.43% | 1.86% | −0.23% | −0.77% | 1.78% | 0.28% | 0.25% |
| p Value | 0.7349 | 0.6113 | 0.2198 | 0.9119 | 0.2744 | 0.0078 * | 0.8428 | 0.0985 | 0.3325 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slywitch, E.; Savalli, C.; Duarte, A.C.; Escrivão, M.A.M.S. Obese Vegetarians and Omnivores Show Different Metabolic Changes: Analysis of 1340 Individuals. Nutrients 2022, 14, 2204. https://doi.org/10.3390/nu14112204
Slywitch E, Savalli C, Duarte AC, Escrivão MAMS. Obese Vegetarians and Omnivores Show Different Metabolic Changes: Analysis of 1340 Individuals. Nutrients. 2022; 14(11):2204. https://doi.org/10.3390/nu14112204
Chicago/Turabian StyleSlywitch, Eric, Carine Savalli, Antonio Cláudio Duarte, and Maria Arlete Meil Schimith Escrivão. 2022. "Obese Vegetarians and Omnivores Show Different Metabolic Changes: Analysis of 1340 Individuals" Nutrients 14, no. 11: 2204. https://doi.org/10.3390/nu14112204
APA StyleSlywitch, E., Savalli, C., Duarte, A. C., & Escrivão, M. A. M. S. (2022). Obese Vegetarians and Omnivores Show Different Metabolic Changes: Analysis of 1340 Individuals. Nutrients, 14(11), 2204. https://doi.org/10.3390/nu14112204