
Serum Metabolomics and Proteomics to Study the Antihypertensive Effect of Protein Extracts from Tenebrio molitor

 , , , , , and
, , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. TM Preparation
2.3. Animal Experiment
2.4. Sample Preparation
2.5. LC-HRMS Metabolomics Analysis
2.6. Proteomics Analysis: Protein Precipitation and Trypsin Digestion
2.7. LC-HRMS/MS Proteomics Analysis and Database Search
2.8. Bioinformatic Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of TM and Captopril Supplementation
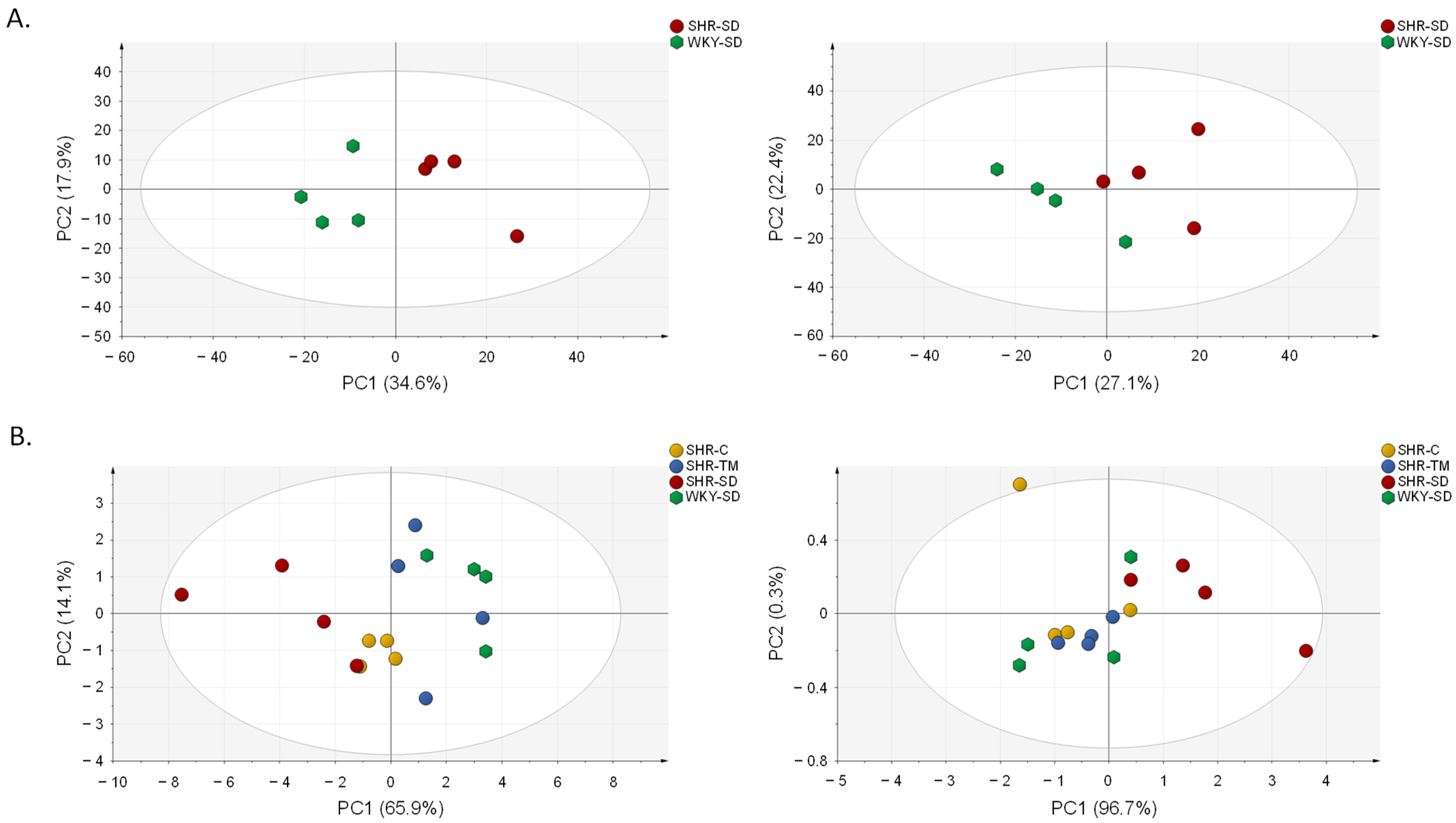
3.2. Metabolomics Profiling of Serum by LC-HRMS
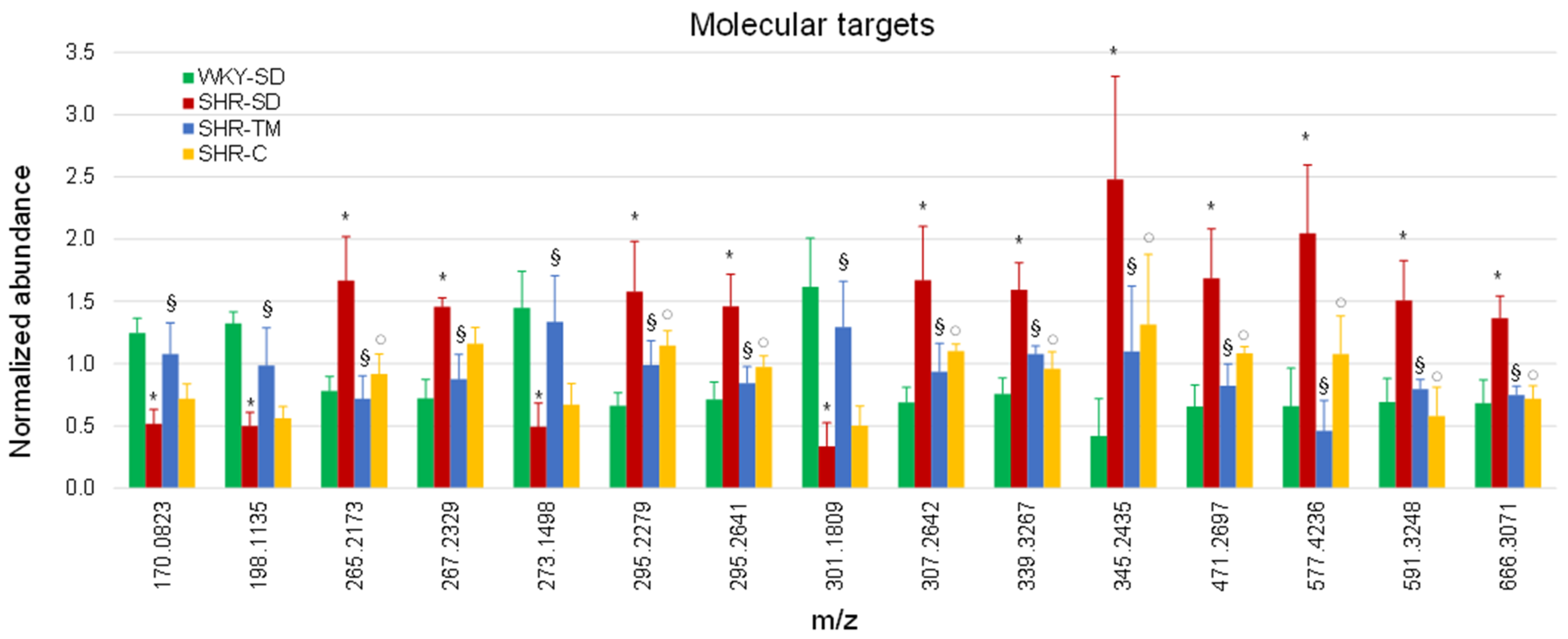
3.3. Annotation of Metabolite Markers
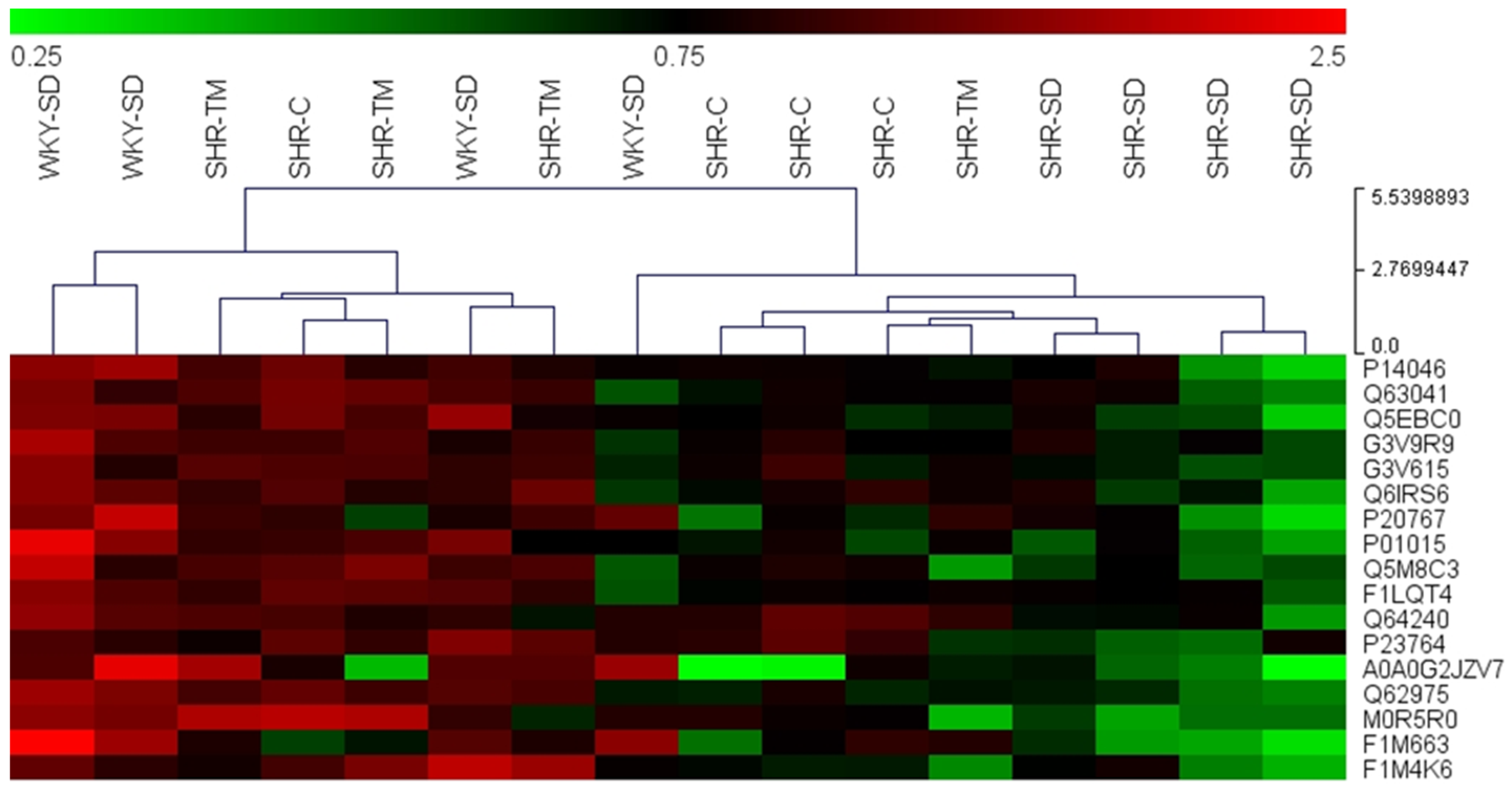
3.4. Proteomics Profiling of Serum by LC-HRMS/MS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Weber, M.A.; Schiffrin, E.L.; White, W.B.; Mann, S.; Lindholm, L.H.; Kenerson, J.G.; Flack, J.M.; Carter, B.L.; Materson, B.J.; Ram, C.V.S.; et al. Clinical Practice Guidelines for the Management of Hypertension in the Community. J. Clin. Hypertens. 2014, 16, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Bentham, J.; Di Cesare, M.; Bixby, H.; Danaei, G.; Cowan, M.J.; Paciorek, C.J.; Singh, G.; Hajifathalian, K.; Bennett, J.E.; et al. Worldwide Trends in Blood Pressure from 1975 to 2015: A Pooled Analysis of 1479 Population-Based Measurement Studies with 19·1 Million Participants. Lancet 2017, 389, 37–55. [Google Scholar] [CrossRef]
- Hwang, A.Y.; Dave, C.; Smith, S.M. Trends in Antihypertensive Medication Use Among US Patients with Resistant Hypertension, 2008 to 2014. Hypertension 2016, 68, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global Burden of Hypertension: Analysis of Worldwide Data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Yoon, S.S.; Gu, Q.; Nwankwo, T.; Wright, J.D.; Hong, Y.; Burt, V. Trends in Blood Pressure Among Adults with Hypertension. Hypertension 2015, 65, 54–61. [Google Scholar] [CrossRef]
- Sim, J.J.; Bhandari, S.K.; Shi, J.; Reynolds, K.; Calhoun, D.A.; Kalantar-Zadeh, K.; Jacobsen, S.J. Comparative Risk of Renal, Cardiovascular, and Mortality Outcomes in Controlled, Uncontrolled Resistant, and Nonresistant Hypertension. Kidney Int. 2015, 88, 622–632. [Google Scholar] [CrossRef]
- Patel, P.; Ordunez, P.; DiPette, D.; Escobar, M.C.; Hassell, T.; Wyss, F.; Hennis, A.; Asma, S.; Angell, S. Improved Blood Pressure Control to Reduce Cardiovascular Disease Morbidity and Mortality: The Standardized Hypertension Treatment and Prevention Project. J. Clin. Hypertens. 2016, 18, 1284–1294. [Google Scholar] [CrossRef]
- Beevers, G. ABC of Hypertension: The Pathophysiology of Hypertension. BMJ 2001, 322, 912–916. [Google Scholar] [CrossRef]
- Verdecchia, P.; Cavallini, C.; Angeli, F. Advances in the Treatment Strategies in Hypertension: Present and Future. J. Cardiovasc. Dev. Dis. 2022, 9, 72. [Google Scholar] [CrossRef]
- Kokkinos, P.; Panagiotakos, D.B.; Polychronopoulos, E. Dietary Influences on Blood Pressure: The Effect of the Mediterranean Diet on the Prevalence of Hypertension. J. Clin. Hypertens. 2005, 7, 165–172. [Google Scholar] [CrossRef]
- Feyh, A.; Bracero, L. Role of Dietary Components in Modulating Hypertension. J. Clin. Exp. Cardiolog. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.W.; Hall, J.E. Seventh Report of the Joint National Committee on Prevention, Detection, Evaluation, and Treatment of High Blood Pressure and Evidence from New Hypertension Trials. Hypertension 2004, 43, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Marte, F.; Sankar, P.; Cassagnol, M. Captopril; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Peng, H.; Carretero, O.A.; Vuljaj, N.; Liao, T.-D.; Motivala, A.; Peterson, E.L.; Rhaleb, N.-E. Angiotensin-Converting Enzyme Inhibitors. Circulation 2005, 112, 2436–2445. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Aoki, K. Development of a Strain of Spontaneously Hypertensive Rats. Jpn. Circ. J. 1963, 27, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Y. Lessons from Rat Models of Hypertension from Goldblatt to Genetic Engineering. Cardiovasc. Res. 1998, 39, 77–88. [Google Scholar] [CrossRef]
- Matsutomo, T.; Ushijima, M.; Kodera, Y.; Nakamoto, M.; Takashima, M.; Morihara, N.; Tamura, K. Metabolomic Study on the Antihypertensive Effect of S -1-Propenylcysteine in Spontaneously Hypertensive Rats Using Liquid Chromatography Coupled with Quadrupole-Orbitrap Mass Spectrometry. J. Chromatogr. B 2017, 1046, 147–155. [Google Scholar] [CrossRef]
- Tian, Y.; Jiang, F.; Li, Y.; Jiang, H.; Chu, Y.; Zhu, L.; Guo, W. Evaluation of the Anti-Hypertensive Effect of Tengfu Jiangya Tablet by Combination of UPLC-Q-Exactive-MS-Based Metabolomics and ITRAQ-Based Proteomics Technology. Biomed. Pharmacother. 2018, 100, 324–334. [Google Scholar] [CrossRef]
- Jiang, H.; Shen, Z.; Chu, Y.; Li, Y.; Li, J.; Wang, X.; Yang, W.; Zhang, X.; Ju, J.; Xu, J.; et al. Serum Metabolomics Research of the Anti-Hypertensive Effects of Tengfu Jiangya Tablet on Spontaneously Hypertensive Rats. J. Chromatogr. B 2015, 1002, 210–217. [Google Scholar] [CrossRef]
- Chu, Y.; Jiang, H.; Ju, J.; Li, Y.; Gong, L.; Wang, X.; Yang, W.; Deng, Y. A Metabolomic Study Using HPLC–TOF/MS Coupled with Ingenuity Pathway Analysis: Intervention Effects of Rhizoma Alismatis on Spontaneous Hypertensive Rats. J. Pharm. Biomed. Anal. 2016, 117, 446–452. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Snyder, M.P. Integrative Omics for Health and Disease. Nat. Rev. Genet. 2018, 19, 299–310. [Google Scholar] [CrossRef]
- Martínez-Espinosa, R. Impact of the “Omics Sciences” in Medicine: New Era for Integrative Medicine. J. Clin. Microbiol. Biochem. Technol. 2017, 3, 9–13. [Google Scholar] [CrossRef]
- Kordalewska, M.; Markuszewski, M.J. Metabolomics in Cardiovascular Diseases. J. Pharm. Biomed. Anal. 2015, 113, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, H.; Chang, X.; Cheng, W.; Zhao, X.; Ding, G.; Zhang, H.; Cai, D.; Zou, Z.-M. Metabolic Pathways Involved in Xin-Ke-Shu Protecting against Myocardial Infarction in Rats Using Ultra High-Performance Liquid Chromatography Coupled with Quadrupole Time-of-Flight Mass Spectrometry. J. Pharm. Biomed. Anal. 2014, 90, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhang, A.; Sun, H.; Dong, W.; Yan, G.; Li, T.; Wang, X. Metabolomic Study of Insomnia and Intervention Effects of Suanzaoren Decoction Using Ultra-Performance Liquid-Chromatography/Electrospray-Ionization Synapt High-Definition Mass Spectrometry. J. Pharm. Biomed. Anal. 2012, 58, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Geyer, P.E.; Holdt, L.M.; Teupser, D.; Mann, M. Revisiting Biomarker Discovery by Plasma Proteomics. Mol. Syst. Biol. 2017, 13, 942. [Google Scholar] [CrossRef] [PubMed]
- Pessina, F.; Frosini, M.; Marcolongo, P.; Fusi, F.; Saponara, S.; Gamberucci, A.; Valoti, M.; Giustarini, D.; Fiorenzani, P.; Gorelli, B.; et al. Antihypertensive, Cardio- and Neuro-Protective Effects of Tenebrio Molitor (Coleoptera: Tenebrionidae) Defatted Larvae in Spontaneously Hypertensive Rats. PLoS ONE 2020, 15, e0233788. [Google Scholar] [CrossRef]
- Costa, S.; Pedro, S.; Lourenço, H.; Batista, I.; Teixeira, B.; Bandarra, N.M.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio Molitor Larvae as an Alternative Food Source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Choi, R.-Y.; Ham, J.R.; Ryu, H.-S.; Lee, S.S.; Miguel, M.A.; Paik, M.-J.; Ji, M.; Park, K.-W.; Kang, K.-Y.; Lee, H.-I.; et al. Defatted Tenebrio Molitor Larva Fermentation Extract Modifies Steatosis, Inflammation and Intestinal Microflora in Chronic Alcohol-Fed Rats. Nutrients 2020, 12, 1426. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.-H.; Hong, S.-J.; Kim, N.-J. Nutritional Value of Mealworm, Tenebrio Molitor as Food Source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]
- Eikenburg, D.C. Effects of Captopril on Vascular Noradrenergic Transmission in SHR. Hypertension 1984, 6, 660–665. [Google Scholar] [CrossRef]
- Koike, H.; Ito, K.; Miyamoto, M.; Nishino, H. Effects of Long-Term Blockade of Angiotensin Converting Enzyme with Captopril (SQ14,225) on Hemodynamics and Circulating Blood Volume in SHR. Hypertension 1980, 2, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Levens, N.R.; Peach, M.J.; Vaughan, E.D.; Weed, W.C.; Carey, R.M. Responses of Blood Pressure and Angiotensin-Converting Enzyme Activity to Acute Captopril Administration in Normotensive and Hypertensive Rats. Endocrinology 1981, 108, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Low-Dose Captopril for the Treatment of Mild to Moderate Hypertension. I. Results of a 14-Week Trial. Veterans Administration Cooperative Study Group on Antihypertensive Agents. Arch. Intern. Med. 1984, 144, 1947–1953. [CrossRef]
- Marcolongo, P.; Gamberucci, A.; Tamasi, G.; Pardini, A.; Bonechi, C.; Rossi, C.; Giunti, R.; Barone, V.; Borghini, A.; Fiorenzani, P.; et al. Chemical Characterisation and Antihypertensive Effects of Locular Gel and Serum of Lycopersicum Esculentum L. Var. “Camone” Tomato in Spontaneously Hypertensive Rats. Molecules 2020, 25, 3758. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for Large-Scale Metabolic Profiling of Serum and Plasma Using Gas Chromatography and Liquid Chromatography Coupled to Mass Spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Lin, S.-Y.; Yeh, Y.-Y.; Hsiao, H.-H.; Wu, C.-Y.; Chen, S.-T.; Wang, A.H.-J. A Modified Protein Precipitation Procedure for Efficient Removal of Albumin from Serum. Electrophoresis 2005, 26, 2117–2127. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Cito, A.; Longo, S.; Mazza, G.; Dreassi, E.; Francardi, V. Chemical Evaluation of the Rhynchophorus Ferrugineus Larvae Fed on Different Substrates as Human Food Source. Food Sci. Technol. Int. 2017, 23, 529–539. [Google Scholar] [CrossRef]
- Brai, A.; Trivisani, C.I.; Vagaggini, C.; Stella, R.; Angeletti, R.; Iovenitti, G.; Francardi, V.; Dreassi, E. Proteins from Tenebrio Molitor: An Interesting Functional Ingredient and a Source Of ACE Inhibitory Peptides. Food Chem. 2022, 393, 133409. [Google Scholar] [CrossRef]
- Te Riet, L.; van Esch, J.H.M.; Roks, A.J.M.; van den Meiracker, A.H.; Danser, A.H.J. Hypertension. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef]
- Son, Y.-J.; Choi, S.Y.; Hwang, I.-K.; Nho, C.W.; Kim, S.H. Could Defatted Mealworm (Tenebrio Molitor) and Mealworm Oil Be Used as Food Ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.A.; Wright, J.W.; Harding, J.W. GABA and Bicuculline-Induced Blood Pressure Changes in Spontaneously Hypertensive Rats. J. Cardiovasc. Pharmacol. 1993, 21, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Hayakawa, K.; Sansawa, H. Involvement of γ-Aminobutyric Acid (GABA) B Receptors in the Hypotensive Effect of Systemically Administered GABA in Spontaneously Hypertensive Rats. Jpn. J. Pharmacol. 2002, 89, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Dang, H.; Wallner, M.; Olsen, R.; Kaufman, D.L. Homotaurine, a Safe Blood-Brain Barrier Permeable GABAA-R-Specific Agonist, Ameliorates Disease in Mouse Models of Multiple Sclerosis. Sci. Rep. 2018, 8, 16555. [Google Scholar] [CrossRef]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of Oral Gamma-Aminobutyric Acid (GABA) Administration on Stress and Sleep in Humans: A Systematic Review. Front. Neurosci. 2020, 14, 923. [Google Scholar] [CrossRef]
- Kawakami, K.; Yamada, K.; Yamada, T.; Nabika, T.; Nomura, M. Antihypertensive Effect of γ-Aminobutyric Acid-Enriched Brown Rice on Spontaneously Hypertensive Rats. J. Nutr. Sci. Vitaminol. 2018, 64, 56–62. [Google Scholar] [CrossRef]
- Aoki, H.; Furuya, Y.; Endo, Y.; Fujimoto, K. Effect of γ-Aminobutyric Acid-Enriched Tempeh-like Fermented Soybean (GABA-Tempeh) on the Blood Pressure of Spontaneously Hypertensive Rats. Biosci. Biotechnol. Biochem. 2003, 67, 1806–1808. [Google Scholar] [CrossRef]
- Rosendorff, C. Captopril—An Overview. S. Afr. Med. J. 1982, 62, 593–599. [Google Scholar]
- Sanrattana, W.; Maas, C.; de Maat, S. SERPINs—From Trap to Treatment. Front. Med. 2019, 6, 25. [Google Scholar] [CrossRef]
- Doolittle, R.F. Angiotensinogen Is Related to the Antitrypsin-Antithrombin-Ovalbumin Family. Science 1983, 222, 417–419. [Google Scholar] [CrossRef]
- Cleland, S.J.; Reid, J.L. The Renin-Angiotensin System and the Heart: A Historical Review. Heart 1996, 76, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.F.; Tofteng, S.S.; Madsen, K.; Jensen, B.L. Role of the Renin–Angiotensin System in Kidney Development and Programming of Adult Blood Pressure. Clin. Sci. 2020, 134, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Wan, J.; Read, R.J.; Carrell, R.W.; Zhou, A. Angiotensinogen and the Modulation of Blood Pressure. Front. Cardiovasc. Med. 2021, 8, 645123. [Google Scholar] [CrossRef]
- Chao, J.; Bledsoe, G.; Chao, L. Protective Role of Kallistatin in Vascular and Organ Injury. Hypertension 2016, 68, 533–541. [Google Scholar] [CrossRef]
- Huang, X.; Dementiev, A.; Olson, S.T.; Gettins, P.G.W. Basis for the Specificity and Activation of the Serpin Protein Z-Dependent Proteinase Inhibitor (ZPI) as an Inhibitor of Membrane-Associated Factor Xa. J. Biol. Chem. 2010, 285, 20399–20409. [Google Scholar] [CrossRef]
- Lip, G.Y. Hypertension and the Prothrombotic State. J. Hum. Hypertens. 2000, 14, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Ghozlan, M.F.; Mohamed, A.A.E.-H.; Eissa, D.S.; Eldawy, H.S. Low Protein Z Level: A Thrombophilic Risk Biomarker for Acute Coronary Syndrome. Indian J. Hematol. Blood Transfus. 2019, 35, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Denecke, B.; Gräber, S.; Schäfer, C.; Heiss, A.; Wöltje, M.; Jahnen-Dechent, W. Tissue Distribution and Activity Testing Suggest a Similar but Not Identical Function of Fetuin-B and Fetuin-A. Biochem. J. 2003, 376, 135–145. [Google Scholar] [CrossRef]
- Olivier, E.; Soury, E.; Ruminy, P.; Husson, A.; Parmentier, F.; Daveau, M.; Salier, J.-P. Fetuin-B, a Second Member of the Fetuin Family in Mammals. Biochem. J. 2000, 350, 589. [Google Scholar] [CrossRef]
- Jung, S.H.; Won, K.-J.; Lee, K.P.; Kim, H.-J.; Seo, E.-H.; Lee, H.M.; Park, E.S.; Lee, S.H.; Kim, B. The Serum Protein Fetuin-B Is Involved in the Development of Acute Myocardial Infarction. Clin. Sci. 2015, 129, 27–38. [Google Scholar] [CrossRef]
- González-Marrero, I.; Castañeyra-Ruiz, L.; González-Toledo, J.M.; Castañeyra-Ruiz, A.; de Paz-Carmona, H.; Castro, R.; Hernandez-Fernaud, J.R.; Castañeyra-Perdomo, A.; Carmona-Calero, E.M. High Blood Pressure Effects on the Blood to Cerebrospinal Fluid Barrier and Cerebrospinal Fluid Protein Composition: A Two-Dimensional Electrophoresis Study in Spontaneously Hypertensive Rats. Int. J. Hypertens. 2013, 2013, 164653. [Google Scholar] [CrossRef] [PubMed]
- Fries, E.; Blom, A.M. Bikunin--Not Just a Plasma Proteinase Inhibitor. Int. J. Biochem. Cell Biol. 2000, 32, 125–137. [Google Scholar] [CrossRef]
- Erlandsson, L.; Ducat, A.; Castille, J.; Zia, I.; Kalapotharakos, G.; Hedström, E.; Vilotte, J.-L.; Vaiman, D.; Hansson, S.R. Alpha-1 Microglobulin as a Potential Therapeutic Candidate for Treatment of Hypertension and Oxidative Stress in the STOX1 Preeclampsia Mouse Model. Sci. Rep. 2019, 9, 8561. [Google Scholar] [CrossRef] [PubMed]
- Ten Kate, M.K.; van der Meer, J. Protein S Deficiency: A Clinical Perspective. Haemophilia 2008, 14, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Lichenstein, H.S.; Lyons, D.E.; Wurfel, M.M.; Johnson, D.A.; McGinley, M.D.; Leidli, J.C.; Trollinger, D.B.; Mayer, J.P.; Wright, S.D.; Zukowski, M.M. Afamin Is a New Member of the Albumin, Alpha-Fetoprotein, and Vitamin D-Binding Protein Gene Family. J. Biol. Chem. 1994, 269, 18149–18154. [Google Scholar] [CrossRef]
- Dieplinger, H.; Dieplinger, B. Afamin—A Pleiotropic Glycoprotein Involved in Various Disease States. Clin. Chim. Acta 2015, 446, 105–110. [Google Scholar] [CrossRef]
- Kurdiova, T.; Balaz, M.; Kovanicova, Z.; Zemkova, E.; Kuzma, M.; Belan, V.; Payer, J.; Gasperikova, D.; Dieplinger, H.; Ukropcova, B.; et al. Serum Afamin a Novel Marker of Increased Hepatic Lipid Content. Front. Endocrinol. 2021, 12, 670425. [Google Scholar] [CrossRef]
- Björnstedt, M.; Xue, J.; Huang, W.; Akesson, B.; Holmgren, A. The Thioredoxin and Glutaredoxin Systems Are Efficient Electron Donors to Human Plasma Glutathione Peroxidase. J. Biol. Chem. 1994, 269, 29382–29384. [Google Scholar] [CrossRef]
- Nirgude, S.; Choudhary, B. Insights into the Role of GPX3, a Highly Efficient Plasma Antioxidant, in Cancer. Biochem. Pharmacol. 2021, 184, 114365. [Google Scholar] [CrossRef]
- Voetsch, B.; Jin, R.C.; Bierl, C.; Benke, K.S.; Kenet, G.; Simioni, P.; Ottaviano, F.; Damasceno, B.P.; Annichino-Bizacchi, J.M.; Handy, D.E.; et al. Promoter Polymorphisms in the Plasma Glutathione Peroxidase (GPx-3) Gene. Stroke 2007, 38, 41–49. [Google Scholar] [CrossRef]
- Lassegue, B. Reactive Oxygen Species in Hypertension*1An Update. Am. J. Hypertens. 2004, 17, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yu, Y.-S. Chronic Hydrogen-Rich Saline Treatment Attenuates Vascular Dysfunction in Spontaneous Hypertensive Rats. Biochem. Pharmacol. 2012, 83, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Robaczewska, J.; Kędziora-Kornatowska, K.; Kucharski, R.; Nowak, M.; Muszalik, M.; Kornatowski, M.; Kędziora, J. Decreased Expression of Heme Oxygenase Is Associated with Depressive Symptoms and May Contribute to Depressive and Hypertensive Comorbidity. Redox Rep. 2016, 21, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Flohé, L. Regulatory Phenomena in the Glutathione Peroxidase Superfamily. Antioxid. Redox Signal. 2020, 33, 498–516. [Google Scholar] [CrossRef]
- Vangaveti, V.N.; Jansen, H.; Kennedy, R.L.; Malabu, U.H. Hydroxyoctadecadienoic Acids: Oxidised Derivatives of Linoleic Acid and Their Role in Inflammation Associated with Metabolic Syndrome and Cancer. Eur. J. Pharmacol. 2016, 785, 70–76. [Google Scholar] [CrossRef]
- Mitchell, L.A.; Grant, D.F.; Melchert, R.B.; Petty, N.M.; Kennedy, R.H. Linoleic Acid Metabolites Act to Increase Contractility in Isolated Rat Heart. Cardiovasc. Toxicol. 2002, 2, 219–229. [Google Scholar] [CrossRef]
- Sousa, T.; Afonso, J.; Carvalho, F.; Albino-Teixeir, A. Lipid Peroxidation and Antioxidants in Arterial Hypertension. In Lipid Peroxidation; InTech: Nappanee, IN, USA, 2012. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z | Retention Time (min) | Elemental Composition | Adduct | Annotation or Identification * | Class | Ionization Polarity | Ratio SHR-SD/SHR-TM |
|---|---|---|---|---|---|---|---|
| 170.0823 | 4.7 | C8H15NO4 | [M-H2O-H]− | hydroxyhexanoylglycine | acyl aminoacids | Negative | 0.48 |
| 198.1135 | 7.2 | C10H19NO4 | [M-H2O-H]− | propionylcarnitine | short chain acylcarnitine | Negative | 0.51 |
| 265.2173 | 13.6 | - | - | - | Negative | 2.32 | |
| 267.2329 | 14.3 | - | - | - | Negative | 1.66 | |
| 273.1498 | 8.7 | - | - | - | Negative | 0.37 | |
| 295.2279 | 11.1 | C18H32O3 | [M-H]− | epoxyoctadecenoic acid | peroxidation product of linoleic acid | Negative | 1.60 |
| 295.2641 | 15.2 | C19H36O2 | [M-H]− | nonadecenoic acid | long chain fatty acids | Negative | 1.73 |
| 301.1809 | 10.2 | C19H26O3 | [M-H]− | 19-hydroxyandrost-4-ene-3,17-dione | steroid | Negative | 0.26 |
| 307.2642 | 14.9 | C20H36O2 | [M-H]− | eicosadienoic acid | unsaturated fatty acids | Negative | 1.79 |
| 339.3267 | 16.9 | C22H44O2 | [M-H]- | docosanoic acid * | unsaturated fatty acids | Negative | 1.48 |
| 345.2435 | 12.1 | - | - | - | Negative | 2.25 | |
| 471.2697 | 14.9 | C11H19NO3 | [2M+HCOO]− | nonenoylglycine | acyl aminoacids | Negative | 2.05 |
| 577.4236 | 13.4 | C31H63O7P | [M-H]− | 1-hexadecyl-2-dodecanoyl-glycero-3-phosphate | glycerophospholipids | Negative | 4.45 |
| 591.3248 | 11.6 | C27H39N7O7 | [M+NH4]+ | - | peptides | Positive | 1.89 |
| 666.3071 | 11.6 | C28H41N11O7 | [M+Na]+ | - | peptides | Positive | 1.83 |
| UniProtKB Accession | Description | Gene Name | Sequence Coverage (%) | Unique Peptides | Ratio SHR-SD/SHR-TM |
|---|---|---|---|---|---|
| P14046 | Alpha-1-inhibitor 3 | A1i3 | 64.0 | 31 | 0.33 |
| Q5EBC0 | Inter alpha-trypsin inhibitor, heavy chain 4 | Itih4 | 73.3 | 107 | 0.32 |
| Q6IRS6 | Fetub protein | Fetub | 62.7 | 47 | 0.43 |
| P20767 | Ig lambda-2 chain C region | ENSRNOG00000050000 | 64.5 | 45 | 0.31 |
| P01015 | Angiotensinogen | Agt | 48.6 | 36 | 0.28 |
| F1LQT4 | Carboxypeptidase N subunit 2 | Cpn2 | 64.6 | 20 | 0.51 |
| Q64240 | Protein AMBP | Ambp | 89.4 | 9 | 0.46 |
| A0A0G2JZV7 | Ig-like domain-containing protein | ENSRNOG00000047464 | 42.1 | 15 | 0.42 |
| Q62975 | Protein Z-dependent protease inhibitor | Serpina10 | 39.7 | 12 | 0.44 |
| M0R5R0 | Protein S (Alpha), isoform CRA_b | Pros1 | 32.2 | 13 | 0.36 |
| F1M663 | Ig-like domain-containing protein | ENSRNOG00000048017 | 24.4 | 8 | 0.28 |
| F1M4K6 | Leucine-rich repeat-containing protein 7 | Lrrc7 | 36.3 | 8 | 0.36 |
| Q63041 | Alpha-1-macroglobulin | A1m | 52.3 | 5 | 0.49 |
| G3V9R9 | Afamin | Afm | 13.5 | 4 | 0.51 |
| G3V615 | Complement factor B | Cfb | 13.9 | 5 | 0.51 |
| Q5M8C3 | Serine (Or cysteine) proteinase inhibitor, clade A (Alpha-1 anti-proteinase, antitrypsin), member 4 | Serpina4 | 6.5 | 5 | 0.42 |
| P23764 | Glutathione peroxidase 3 | Gpx3 | 33.9 | 3 | 0.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stella, R.; Peggion, C.; Bergantin, C.; Biancotto, G.; Frosini, M.; Dreassi, E.; Marcolongo, P.; Aloisi, A.M.; Pessina, F. Serum Metabolomics and Proteomics to Study the Antihypertensive Effect of Protein Extracts from Tenebrio molitor. Nutrients 2022, 14, 3288. https://doi.org/10.3390/nu14163288
Stella R, Peggion C, Bergantin C, Biancotto G, Frosini M, Dreassi E, Marcolongo P, Aloisi AM, Pessina F. Serum Metabolomics and Proteomics to Study the Antihypertensive Effect of Protein Extracts from Tenebrio molitor. Nutrients. 2022; 14(16):3288. https://doi.org/10.3390/nu14163288
Chicago/Turabian StyleStella, Roberto, Caterina Peggion, Caterina Bergantin, Giancarlo Biancotto, Maria Frosini, Elena Dreassi, Paola Marcolongo, Anna Maria Aloisi, and Federica Pessina. 2022. "Serum Metabolomics and Proteomics to Study the Antihypertensive Effect of Protein Extracts from Tenebrio molitor" Nutrients 14, no. 16: 3288. https://doi.org/10.3390/nu14163288